
Abstract
Introduction
APOE is the strongest risk gene for sporadic Alzheimer's disease (AD) so far. Recent genome wide association studies found links for sporadic AD with CLU and CR1 involved in Aβ clearance, and PICALM affecting intracellular trafficking.
Methods
We investigated the associations of senile plaques (SP) and neurofibrillary tangles (NFT) with the proposed risk genes and APOE, in the Tampere Autopsy Study (TASTY) series (603 cases), a sample of the general population (0 to 97 yrs), who died out-of-hospital.
Results
Age and the APOEε4 allele associated strongly with all phenotypes of SP, as expected. In age and APOEε4 adjusted analyses, compared to the most common homozygous genotype, burnt out SP were more common among carriers of the C-allele of CLU, whereas the T-allele of PICALM and C-allele of CR1 were linked with lower SP coverage. We found no significant associations between any of the genetic variants and NFT.
Conclusions
Marginal effects from CLU, CR1 and PICALM suggest that these genes have minimal effects on the development of AD lesions.
Similar content being viewed by others

Introduction
Alzheimer's disease (AD) is the most common form of dementia in Western society and is, and will continue to be, a burden on health systems in the future as the population ages. Age is the largest risk factor for the disease, with higher incidences in older populations [1, 2].
Identification of genes related to sporadic AD risk has been slow with study groups isolating only one strongly associated gene: APOE [3, 4]. The epsilon 4 allele of apolipoprotein E (APOEε4) provides odds ratios (ORs) of between 3 and 25 [5, 6] for disease association. APOEε4 is suspected to have a lower effectiveness at transporting cholesterol and is not as efficient at repairing neuronal damage as APOEε3 [7]. One or even two copies of the allele, however, are not sufficient to cause the disease, as many carriers of two ε4 alleles do not develop AD [5].
Studies aiming to detect genes associated with disease risk have used heterogeneous AD cohorts and ascertained few polymorphisms with only a minor impact on disease incidence. One of the problems is to distinguish between pure AD, vascular dementia and other dementia types in clinical cohorts [8–10]. The only consistent and currently accepted method for confirming AD is with post-mortem assessment of the neuropathological lesions neurofibrillary tangles (NFT) and senile plaques (SP) [11–14].
Demented individuals do not always exhibit large enough numbers of SP to warrant an AD diagnosis [15] and NFT and SP are both relatively common in the general population [16–19]. Furthermore, these lesions do not provide a clear-cut explanation as to the cause of AD, with different theories advocating amyloid beta (Aβ) accumulation [12, 20] or hyperphosphorylated NFT-causing tau protein [21] as the underlying initiating mechanisms that trigger the disease.
Two recent extensive genome-wide association studies (GWAS), comprising 12,000 probable AD cases and 18,000 age-matched non-demented controls [22, 23], revealed three new candidates for genetic risk of developing late onset or sporadic AD: CLU, CR1 and PICALM. Phosphatidylinositol-binding clathrin assembly protein (PICALM) is involved in synaptic neurotransmitter release and intracellular trafficking [24–26], whilst complement component (3b/4b) receptor 1 (CR1), the main receptor of complement C3b protein, binds Aβ and thus may promote clearance [27–30]. Clusterin (CLU, and also known as ApoJ), was replicated independently in the two studies and is thought to bind and remove Aβ from the brain, as well as assist in re-entry of Aβ [31–34].
We have previously shown that as many as one-third of non-demented individuals in an autopsy series-based sample carry SP and more than 40% NFT, with strong age dependence [16]. This suggests that in clinical study series, non-demented control patients may not be free of AD-related neuropathological lesions. Utilising this same cohort, we aimed to investigate whether SP and NFT are associated with any of the recently identified GWAS single nucleotide polymorphisms (SNPs); CLU, CR1 and PICALM to examine their involvement in the development of these brain lesions.
Materials and methods
Autopsy series
The Tampere Autopsy Study (TASTY) cohort consisted of 603 autopsy cases, of which the majority died out-of-hospital within Tampere, Finland and surroundings, collected during the years 2002 to 2004 (described in detail elsewhere [16]). The study was approved by the Board of Medicolegal Affairs of Finland. Females within the cohort accounted for 35.8% and the ages for the entire series ranged from 0 to 97, with an average of 63 years (59 years for males and 68 years for females). Of the cases, 6 (1%) had a clinical AD diagnosis, 16 (2.7%) undefined dementia, 10 (1.7%) had memory disorders and 1 (0.2%) had Parkinson's disease prior to death (according to available hospital records and next of kin reports). In some cases it was impossible to obtain all variables due to technical difficulties and sample damage.
Alzheimer-related lesion measurements
SP and NFT staining and measures have be portrayed previously [16]. Briefly, the Bielschowsky argyrophilic silver impregnation method was performed on samples and measured by two researchers to acquire SP (neocortex) and NFT (hippocampus) counts. Each area was screened to find the highest density of SP (neocortical area at 100 × magnification) and NFT (hippocampus - CA1 area at 200 × magnification) and then scored using a square microscopic grid (SP - 100 intersections covering 1 mm2, NFT - four to six random columns), before creating an average percentage of coverage (SP) or average number in 1 mm2 (NFT). Bielschowsky staining was highly correlated with Amyloid beta (Aβ) staining (antibody from Cell Signaling Technology, Danvers, MA, USA; concentration 1:150), which was used to verify our previous results [16], and used the same cortical coverage of SP method. SP and NFT variables included the following categorisations as measured by a neuropathologist: SP (No, Yes), SP type (No Plaques, Diffuse, Primitive, Classic, Burnt Out), SP type 2 (No Plaques, Non-neuritic SP, Neuritic SP), NFT (No, Yes), where reference groups were those with 'No SP' or 'No NFT' and those with either brain lesion were considered 'affected'. Semi-quantitative data for SP utilised the categories 'no', 'sparse', 'moderate' and 'frequent' SP.
Genotyping
The ABI Prism 7900HT Sequence Detection System used 1 μl DNA with PCR primers (Applied Biosystems, Espoo, Finland) for rs11136000 (CLU), rs1408077 (CR1) and rs3851179 (PICALM). All SNPs were in Hardy-Weinberg equilibrium and genotyping confirmed using SDS version 2.2 (Applied Biosystems). Genotyping for APOE has been previously described [16]. Genotyping for the polymorphisms of CLU, CR1 and PICALM were successful for 94%, 97% and 97% of the TASTY cases, respectively.
Statistics
Logistic regression analyses, with continuous age and APOEε4 carriership as covariates (where possible), were used with SPSS (version 14.0 for Windows; SPSS Finland Oy, Espoo, Finland) to determine associations between the SNPs and AD-related neuropathological lesions. For all SNPs, the most common homozygous genotype was used as the reference group. As previously mentioned, those unaffected by SP or NFT were considered the reference group for the brain lesion categories. When analysing with the cohort split by age groups, the following categories were used: 0 to 49 years, 50 to 59 years, 60 to 69 years, 70 to 79 years, 80+ years, with the youngest group (0 to 49 years) considered the reference group with respect to age, in analyses. The cohort was also split by gender, where mentioned.
Results
Autopsy series characteristics
The Tampere Autopsy Series (TASTY) (n = 603) comprises consecutive autopsies on males and females aged 0 to 97 years that lived outside institutions or hospitals (see Table 1). Females were on average 10 years older than males, but males were more likely to have SP compared to females (odds ratio (OR) 2.15, P < 0.0001, 95% confidence intervals (CI) 1.49 to 3.11). When age was divided into five equal-sized groups, each age group was consistently more likely to have SP compared to the youngest group, with each association also highly statistically significant (see Table 2). This was also true for NFT prevalence (see Table 2), with females more likely than males to have NFT (OR 2.18, P < 0.0001, CI 1.49 to 3.18).
APOE, CLU, CR1 and PICALMassociations with SP
As expected, APOEε4 carriership was significantly associated with increased risk of having SP (OR 2.52, P < 0.0001, CI 1.72 to 3.68); having both non-neuritic (OR 2.42, P = 0.003, CI 1.36 to 4.29) and neuritic SP (OR 2.58, P < 0.0001, CI 1.66 to 4.02) compared to no SP; and having primitive (OR 2.53, P = 0.010, CI 1.25 to 5.10), classic (OR 2.52, P < 0.0001, CI 1.54 to 4.13) and burnt out SP (OR 2.77, P = 0.014, CI 1.22 to 6.27) compared to no SP, when evaluated against non ε4 carriers (see Table 3). Results showed similar trends when the cohort was split by gender (data not shown).
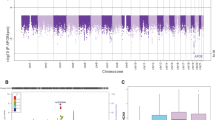
APOEε4 carriers, compared to ε3-ε3 carriers, were significantly associated with an increased risk of having SP in all age groups except the youngest and oldest (Figure 1). There was a trend of age-related increases in SP, especially of the neuritic type, across all studied genotypes. The APOEε2 carrier group was too small to investigate supposed protective effects, although previously published results suggest tendencies towards protection [16]. In APOEε4 adjusted analyses, 80+ year old carriers of the rare TT genotype of PICALM had a significantly lower incidence of SP compared to the common CC carriers (OR 0.18, P = 0.025, CI 0.04 to 0.81) (see Figure 1). This association was not seen among younger age groups. There were no significant associations between genotypes of CLU and CR1 and SP prevalence.

Senile plaque prevalence by age and genotype ( APOE, CLU, CR1 and PICALM ). CI = confidence interval; OR = odds ratio.
Grouping the rare homozygote and heterozygotes versus the common homozygotes for the SNPs uncovered statistically significant associations between the T allele of PICALM and SP (OR 0.62, P = 0.028, CI 0.41 to 0.95, versus CC genotype). When we divided the SP into diffuse, primitive, classic and burnt out phenotypes (to investigate the particular phases of the SP life cycle), we found that the rare C allele of CLU was significantly associated with the presence of late stage SP (OR 4.4, P = 0.004, CI 1.61 to 12.2) compared to the common TT genotype (Table 3). In that setting, the statistically significant association of the PICALM T allele was lost.
APOE, CLU, CR1 and PICALMassociations with SP frequency
When analyses were performed with SP frequency as the dependent variable, APOEε4 carriership was again found to be highly significantly associated with increasing SP coverage, compared to ε3-ε3 carriers (see Table 4). PICALM TC genotypes (versus CC genotype) were significantly less likely to have moderate SP compared to no SP (OR 0.42, P = 0.012, CI 0.21 to 0.83), whilst CR1 CC genotype carriers (compared to AA genotype) were more likely to have sparse SP than no SP (OR 2.1, P = 0.048, CI 1.01 to 4.43).
When we grouped the rare homozygote and heterozygotes together versus the common homozygotes (Table 4), significance was lost for CR1, however PICALM T allele carriers remained less likely to have coverage of SP versus no SP compared to CC genotype, however again statistical significance was only reached for moderate SP (OR 0.43, P = 0.010, CI 0.23 to 0.82).
Associations with gender
Reanalysing the significant associations with the cohort split by gender gave similar results (data not shown), with females generally more strongly associated, most likely due to their older age. No further significant associations were uncovered.
Associations with Aβ staining
A subpopulation of the cohort were assessed for associations with immunohistochemical staining (n = 152). None of the newly identified SNPs were statistically significantly associated with Aβ staining, as seen in Figure 2. APOEε4 carriership, however, was significantly associated with higher cortical coverage of Aβ staining (P < 0.0001).

Boxplots of cortical SP coverage (%) according to Aβ staining and genotype ( APOE, CLU, CR1 and PICALM ).
Discussion
AD is the most common form of dementia, but to date its aetiology has remained elusive, despite intensive research. The proposed causes of AD relate to neuropathological findings post-mortem, which is the only way to definitively confirm a patient's diagnosis [11–14]. Diagnosis of the first AD patient, back in 1906, revealed large numbers of SP and NFT; however, although new treatments aimed at reversing the disease by reducing SP have proven successful, they have been without improvements in cognitive abilities of patients [35]. Furthermore, studies have shown cognitively normal elderly can also have large numbers of these brain lesions [16–19] and not all AD cases have the required amounts to corroborate cognitive dysfunction [15].
Genome wide association studies (GWAS) investigating AD have in the past not been powerful enough to reveal anything except APOE. Two recent large GWAS [22, 23], however, collectively investigated over 30,000 individuals (with almost 12,000 probable AD cases) and examined around 500,000 SNPs that may influence AD risk.
We recently showed that SP and NFT were surprisingly common in a non-demented autopsy series, which represents the closest model to a population sample and that the occurrence of SP, but not NFT, was strongly affected by the APOEε4 allele, regardless of age [16]. Because of the GWAS' discoveries of three potential new candidates for AD risk, we decided to look at their associations with the neuropathological lesions SP and NFT in our cohort to investigate their involvement in the development of these brain lesions.
SP associated with both age and gender, and the APOEε4 allele was highly associated with SP in many of our analyses. Additional analyses showed that the APOEε4 associations were extremely robust in the TASTY series, thus validating our cohort's ability to detect associations with the measured brain lesions. However, whilst NFT were found to associate with age and gender, they were not associated with any of the SNPs investigated. The strong association seen between males and SP prevalence, and females and NFT occurrence may be a confounding factor, due to the older average age of females in the cohort. Additionally, our cohort may over-represent early and violent deaths; however, all cases were included to best represent a population-based investigation.
We hypothesised that the three other SNPs (CLU, CR1 and PICALM) would also associate with SP, as they are involved in AD pathways and most likely would be associated with the development of brain lesions [24–34]. Our results indicate that we did not find as many robust correlations as for APOE.
Genetic variants of CLU, PICALM and CR1 genes were associated with SP and remained so with the inclusion of APOEε4 carriers and age as covariates. The appearance of an increased risk for CLU C carriers versus TT is unusual in that it only applies for Burnt Out SP - a group in the cohort that is relatively small and are majority females. This suggests that the effect of CLU could be on the later stages of SP development and related to removal of Aβ [31–34] being reduced in efficiency.
The PICALM T allele appears to have a protective effect on SP prevalence, true also for TT genotypes, versus CC genotypes in the oldest age group. This may be due to more efficient intracellular trafficking and clear-up of Aβ, or the components or pathways that induce Aβ build-up or production [24–27]. The protective effect of the T carriers was seen also for SP coverage; however, it was only significant for moderate SP.
CR1 CC genotype carriers were associated with an increased risk of having sparse compared to no SP; however, the trend was not consistent for increasing coverage of SP (data not shown), which was also true for the analysis grouping the rare homozygote with the heterozygotes. This suggests the effect of CR1 is complex and not as straight-forward as increasing SP risk and requires further investigation.
The lack of robust and numerous associations with the GWAS SNPs and brain lesions, alongside the strong APOEε4 results, questions the involvement of SP in AD pathology. It may be a coincidence that SP are found in AD brains with evidence suggesting that they may be a part of normal aging [16, 17]. In light of this, SP treatments have so far failed to improve patients' cognitive abilities [35] and current theories are moving away from SP and suggest soluble oligomeric Aβ may be the culprit in AD [36–39]. The scarcity of associations may be due to the small number of cases with SP within the TASTY series (31.1%), resulting in low power; however, we have a 600 case-strong cohort, which revealed strong associations between APOE with SP. It may also be due to the low strength of these prior associations in the original studies, which should be investigated in future cohorts of a similar nature.
Some may question the use of an autopsy series to investigate an age-dependent illness such as AD; however, the TASTY cohort provides a unique view into the early midlife prevalence of well-defined neuropathological lesions, showing their common prevalence.
Conclusions
We have an interesting window into the development of neuropathological lesions and their associations with AD-risk genes in the general population, and as far as we know, this is the first study of its kind. SP were found to associate with age, gender, and APOEε4, but not consistently with CLU, CR1 or PICALM, suggesting that these SNPs most likely do not affect the development of the studied neuropathological lesions. Further studies should replicate these findings in a larger autopsy series of the same kind, both with and without AD cases, to define the occurrence of these neuropathological lesions within the context of normal aging.
Our results suggest that whilst these SNPs are associated with probable AD cases (in the GWAS), they do not strongly relate to SP prevalence, or at all to NFT, in an autopsy series most representative of the general population, possibly indicating their complex involvement in the disease.
Abbreviations
- AD:
-
Alzheimer's disease
- APOE:
-
apolipoprotein E
- CI:
-
confidence interval
- CLU:
-
clusterin
- CR1:
-
complement component (3b/4b) receptor 1
- GWAS:
-
genome wide association studies
- NFT:
-
neurofibrillary tangles
- OR:
-
odds ratio
- PICALM:
-
phosphatidylinositol binding clathrin assembly protein
- SNPs:
-
single nucleotide polymorphisms
- SP:
-
senile plaques
- TASTY:
-
Tampere autopsy study.
References
Polvikoski T, Sulkava R, Rastas S, Sutela A, Niinistö L, Notkola I, Verkkoniemi A, Viramo P, Juva K, Haltia M: Incidence of dementia in very elderly individuals: a clinical, neuropathological and molecular genetic study. Neuroepidemiology. 2006, 26: 76-82. 10.1159/000090252.
Harvey DJ, Beckett LA, Mungas DM: Multivariate modeling of two associated cognitive outcomes in a longitudinal study. J Alzheimers Dis. 2003, 5: 357-365.
Corder E, Saunders A, Strittmatter W, Schmechel D, Gaskell P, Small G, Roses A, Haines J, Pericak-Vance M: Gene dose of apolipoprotein E type 4 allele and the risk of Alzheimer's disease in late onset families. Science. 1993, 261: 921-923. 10.1126/science.8346443.
van Duijn C, Wehnert A, Van Broeckhoven C, Havekes LM, de Knijff P, Cruts M, Hofman A: Apolipoprotein E4 allele in a population-based study of early-onset Alzheimer's disease. Nat Genet. 1994, 7: 74-78. 10.1038/ng0594-74.
Farrer L, Cupples L, Haines J, Hyman B, Kukull W, Mayeux R, Myers R, Pericak-Vance M, Risch N, van Duijn C: Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. JAMA. 1997, 278: 1349-1356. 10.1001/jama.278.16.1349.
Coon KD, Myers AJ, Craig DW, Webster JA, Pearson JV, Lince DH, Zismann VL, Beach TG, Leung D, Bryden L, Halperin RF, Marlowe L, Kaleem M, Walker DG, Ravid R, Heward CB, Rogers J, Papassotiropoulos A, Reiman EM, Hardy J, Stephan DA: A high-density whole-genome association study reveals that APOE is the major susceptibility gene for sporadic late-onset Alzheimer's disease. J Clin Psychiatry. 2007, 68: 613-618. 10.4088/JCP.v68n0419.
Mahley RW, Rall SC: Apolipoprotein E: far more than a lipid transport protein. Annu Rev Genomics Hum Genet. 2000, 1: 507-537. 10.1146/annurev.genom.1.1.507.
Jellinger KA, Attems J: Prevalence of dementia disorders in the oldest-old: an autopsy study. Acta Neuropathol. 2010, 119: 421-433. 10.1007/s00401-010-0654-5.
Galasko D: Biomarkers in non-Alzheimer dementias. Clin Neurosci Res. 2004, 3: 375-381. 10.1016/j.cnr.2004.04.004.
Cavalieri M, Enzinger C, Petrovic K, Pluta-Fuerst A, Homayoon N, Schmidt H, Fazekas F, Schmidt R: Vascular dementia and Alzheimer's disease - are we in a dead-end road?. Neurodegener Dis. 2010, 7: 122-126. 10.1159/000285521.
Alafuzoff I, Pikkarainen M, Al-Sarraj S, Arzberger T, Bell J, Bodi I, Bogdanovic N, Budka H, Bugiani O, Ferrer I, Gelpi E, Giaccone G, Graeber MB, Hauw J, Kamphorst W, King A, Kopp N, Korkolopoulou P, Kovács GG, Meyronet D, Parchi P, Patsouris E, Preusser M, Ravid R, Roggendorf W, Seilhean D, Streichenberger N, Thal DR, Kretzschmar H: Interlaboratory comparison of assessments of Alzheimer disease-related lesions: a study of the BrainNet Europe Consortium. J Neuropathol Exp Neurol. 2006, 65: 740-757. 10.1097/01.jnen.0000229986.17548.27.
Duyckaerts C, Delatour B, Potier M: Classification and basic pathology of Alzheimer disease. Acta Neuropathol. 2009, 118: 5-36. 10.1007/s00401-009-0532-1.
Braak H, Braak E: Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82: 239-259. 10.1007/BF00308809.
Mirra SS, Heyman A, McKeel D, Sumi SM, Crain BJ, Brownlee LM, Vogel FS, Hughes JP, van Belle G, Berg L: The Consortium to Establish a Registry for Alzheimer's Disease (CERAD). Part II. Standardization of the neuropathologic assessment of Alzheimer's disease. Neurology. 1991, 41: 479-486.
Iqbal K, Flory M, Khatoon S, Soininen H, Pirttila T, Lehtovirta M, Alafuzoff I, Blennow K, Andreasen N, Vanmechelen E, Grundke-Iqbal I: Subgroups of Alzheimer's disease based on cerebrospinal fluid molecular markers. Ann Neurol. 2005, 58: 748-757. 10.1002/ana.20639.
Kok E, Haikonen S, Luoto T, Huhtala H, Goebeler S, Haapasalo H, Karhunen P: Apolipoprotein E-dependent accumulation of Alzheimer disease-related lesions begins in middle age. Ann Neurol. 2009, 65: 650-657. 10.1002/ana.21696.
Ghebremedhin E, Schultz C, Thal D, Rub U, Ohm T, Braak E, Braak H: Gender and age modify the association between APOE and AD-related neuropathology. Neurology. 2001, 56: 1696-1701.
Savva GM, Wharton SB, Ince PG, Forster G, Matthews FE, Brayne C: Age, neuropathology, and dementia. N Engl J Med. 2009, 360: 2302-2309. 10.1056/NEJMoa0806142.
Nicoll J, Savva GM, Stewart J, Matthews FE, Brayne C, Ince P: Association between APOE genotype, neuropathology and dementia in the older population of England and Wales. Neuropathol Appl Neurobiol. 2011, 37: 285-294. 10.1111/j.1365-2990.2010.01130.x.
Tanzi R, Kovacs D, Kim T, Moir K, Guenette S, Wasco W: The gene defects responsible for familial Alzheimer's disease. Neurobiol Dis. 1996, 3: 159-168. 10.1006/nbdi.1996.0016.
Rapoport M, Dawson HN, Binder LI, Vitek MP, Ferreira A: Tau is essential to beta-amyloid-induced neurotoxicity. Proc Natl Acad Sci USA. 2002, 99: 6364-6369. 10.1073/pnas.092136199.
Lambert JC, Heath S, Even G, Campion D, Sleegers K, Hiltunen M, Combarros O, Zelenika D, Bullido MJ, Tavernier B, Letenneur L, Bettens K, Berr C, Pasquier F, Fiévet N, Barberger-Gateau P, Engelborghs S, De Deyn P, Mateo I, Franck A, Helisalmi S, Porcellini E, Hanon O, the European Alzheimer's Disease Initiative Investigators, de Pancorbo MM, Lendon C, Dufouil C, Jaillard C, Leveillard T, Alvarez V, et al: Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer's disease. Nat Genet. 2009, 41: 1094-1099. 10.1038/ng.439.
Harold D, Abraham R, Hollingworth P, Sims R, Gerrish A, Hamshere ML, Pahwa JS, Moskvina V, Dowzell K, Williams A, Jones N, Thomas C, Stretton A, Morgan AR, Lovestone S, Powell J, Proitsi P, Lupton MK, Brayne C, Rubinsztein DC, Gill M, Lawlor B, Lynch A, Morgan K, Brown KS, Passmore PA, Craig D, McGuinness B, Todd S, Holmes C, et al: Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat Genet. 2009, 41: 1088-1093. 10.1038/ng.440.
Tebar F, Bohlander S, Sorkin A: Clathrin assembly lymphoid myeloid leukemia (CALM) protein: localization in endocytic-coated pits, interactions with clathrin, and the impact of overexpression on clathrin-mediated traffic. Mol Biol Cell. 1999, 10: 2687-2702.
Yao P, Petralia R, Bushlin I, Wang Y, Furukawa K: Synaptic distribution of the endocytic accessory proteins AP180 and CALM. J Comp Neurol. 2005, 481: 58-69. 10.1002/cne.20362.
Dreyling M, Martinez-Climent J, Zheng M, Mao J, Rowley J, Bohlander S: The t(10;11)(p13;q14) in the U937 cell line results in the fusion of the AF10 gene and CALM, encoding a new member of the AP-3 clathrin assembly protein family. Proc Natl Acad Sci USA. 1996, 93: 4804-4809. 10.1073/pnas.93.10.4804.
Kuo Y, Kokjohn T, Kalback W, Luehrs D, Galasko D, Chevallier N, Koo E, Emmerling M, Roher A: Amyloid-beta peptides interact with plasma proteins and erythrocytes: implications for their quantitation in plasma. Biochem Biophys Res Commun. 2000, 268: 750-756. 10.1006/bbrc.2000.2222.
Rogers J, Li R, Mastroeni D, Grover A, Leonard B, Ahern G, Cao P, Kolody H, Vedders L, Kolb W, Sabbagh M: Peripheral clearance of amyloid beta peptide by complement C3-dependent adherence to erythrocytes. Neurobiol Aging. 2006, 27: 1733-1739. 10.1016/j.neurobiolaging.2005.09.043.
Zhou J, Fonseca M, Pisalyaput K, Tenner A: Complement C3 and C4 expression in C1q sufficient and deficient mouse models of Alzheimer's disease. J Neurochem. 2008, 106: 2080-2092. 10.1111/j.1471-4159.2008.05558.x.
Wyss-Coray T, Yan F, Lin AH, Lambris JD, Alexander JJ, Quigg R, Masliah E: Prominent neurodegeneration and increased plaque formation in complement-inhibited Alzheimer's mice. Proc Natl Acad Sci USA. 2002, 99: 10837-10842. 10.1073/pnas.162350199.
Jenne D, Tschopp J: Clusterin: the intriguing guises of a widely expressed glycoprotein. Trends Biochem Sci. 1992, 17: 154-159. 10.1016/0968-0004(92)90325-4.
Jones S, Jomary C: Clusterin. Int J Biochem Cell Biol. 2002, 34: 427-431. 10.1016/S1357-2725(01)00155-8.
DeMattos RB, Cirrito JR, Parsadanian M, May PC, O'Dell MA, Taylor JW, Harmony JA, Aronow BJ, Bales KR, Paul SM, Holtzman DM: ApoE and clusterin cooperatively suppress Abeta levels and deposition: evidence that ApoE regulates extracellular Abeta metabolism in vivo. Neuron. 2004, 41: 193-202. 10.1016/S0896-6273(03)00850-X.
Calero M, Rostagno A, Matsubara E, Zlokovic B, Frangione B, Ghiso J: Apolipoprotein J (clusterin) and Alzheimer's disease. Microsc Res Tech. 2000, 50: 305-315. 10.1002/1097-0029(20000815)50:4<305::AID-JEMT10>3.0.CO;2-L.
Holmes C, Boche D, Wilkinson D, Yadegarfar G, Hopkins V, Bayer A, Jones RW, Bullock R, Love S, Neal JW, Zotova E, Nicoll JA: Long-term effects of Abeta42 immunisation in Alzheimer's disease: follow-up of a randomised, placebo-controlled phase I trial. Lancet. 2008, 372: 216-223. 10.1016/S0140-6736(08)61075-2.
Gandy S, Simon AJ, Steele JW, Lublin AL, Lah JJ, Walker LC, Levey AI, Krafft GA, Levy E, Checler F, Glabe C, Bilker WB, Abel T, Schmeidler J, Ehrlich ME: Days to criterion as an indicator of toxicity associated with human Alzheimer amyloid-beta oligomers. Ann Neurol. 2010, 68: 220-230.
Moir RD, Tseitlin KA, Soscia S, Hyman BT, Irizarry MC, Tanzi RE: Autoantibodies to redox-modified oligomeric Abeta are attenuated in the plasma of Alzheimer's disease patients. J Biol Chem. 2005, 280: 17458-17463. 10.1074/jbc.M414176200.
Tomiyama T, Matsuyama S, Iso H, Umeda T, Takuma H, Ohnishi K, Ishibashi K, Teraoka R, Sakama N, Yamashita T, Nishitsuji K, Ito K, Shimada H, Lambert MP, Klein WL, Mori H: A mouse model of amyloid beta oligomers: their contribution to synaptic alteration, abnormal tau phosphorylation, glial activation, and neuronal loss in vivo. J Neurosci. 2010, 30: 4845-4856. 10.1523/JNEUROSCI.5825-09.2010.
Hardy J, Selkoe DJ: The amyloid hypothesis of Alzheimer's disease: progress and problems on the road to therapeutics. Science. 2002, 297: 353-356. 10.1126/science.1072994.
Acknowledgements
Many thanks to Heini Huhtala (for assistance with statistical analyses) and Sari Tuomisto (for assistance with genotyping). This work was supported by funds from the Medical Research Fund of Tampere University Hospital, the Pirkanmaa Regional Fund of the Finnish Cultural Foundation, the Finnish Foundation for Cardiovascular Research, and the Yrjö Jahnsson Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
All authors contributed to this manuscript. EHK performed experiments and analyses and wrote the manuscript. HH, TL and SH measured the neuropathological lesions. SG and PJK collected the autopsy series. SG, HH and PJK provided comments and discussions on the progress of the manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Kok, E.H., Luoto, T., Haikonen, S. et al. CLU, CR1 and PICALM genes associate with Alzheimer's-related senile plaques. Alz Res Therapy 3, 12 (2011). https://doi.org/10.1186/alzrt71
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/alzrt71