
Abstract
Background
Zinc is an essential metal involved in many physiological processes. Previous work has identified a set of zinc transporters involved in Drosophila dietary zinc absorption. However, zinc excretion and reabsorption, the other two important processes to maintain zinc homeostasis, are not as well understood. In this work, we screened all the potential zinc transporter Zip (SLC39) and ZnT (SLC30) members for their likely roles in zinc excretion in Malpighian tubules, an insect organ functionally analogous to mammalian kidneys.
Results
Zip71B (CG10006, most homologous to hZIP5), in addition to the previously characterized ZnT35C (CG3994), was identified as being critical in zinc excretion. Tubule-specific knockdown of Zip71B/dZip5 reduces zinc accumulation in the tubules, but increases zinc levels in the body, resulting in survival defect under zinc excess conditions. Zip71B/dZip5 is localized to the plasma membrane at the basolateral side of the tubules, and is functionally epistatic to the apically localized ZnT35C in regulating the tubule zinc homeostasis. Our results indicate that Zip71B/dZip5 is involved in zinc import into the tubular cells from the circulation, and ZnT35C in turn effluxes the tubular zinc out. Notably, mammalian ZIP5, which is expressed in the kidney, functions analogously to Zip71B/dZip5 in the fly while hZIP4 cannot complement the loss of Zip71B/dZip5 function. Furthermore, Zip71B/dZip5 expression is regulated by zinc so that, in response to toxic levels of zinc, the tubules can increase zinc efflux capability. We also characterized the role of dZnT1 (CG17723) in zinc reabsorption in Malpighian tubules. Finally, using a tubule calcification model, we were able to show that knockdown of Zip71B/dZip5 or ZnT35C was able to mitigate stone formation, consistent with their roles in tubular zinc homeostasis.
Conclusions
Our results start to sketch out a relatively complete picture of the zinc excretion process in Drosophila Malpighian tubules, and may provide a reference for relevant mammalian studies.
Similar content being viewed by others


Background
Zinc is an essential micronutrient that serves as a catalytic or structural component of numerous enzymes and other proteins and accordingly contributes to a variety of fundamental biological processes [1–5]. Maintenance of zinc homeostasis is very important for human health. Either zinc deficiency or zinc toxicosis can lead to physiologic abnormalities [6–8]. Many common diseases including diabetes mellitus, Crohn’s disease, and Alzheimer’s disease are also linked with zinc dyshomeostasis [9–14].
Zinc homeostasis is tightly controlled by a series of transporter proteins that are broadly categorized into two major families, the Slc39a or Zip family and the Slc30a or ZnT family. Generally speaking, Zip proteins function in transporting zinc into the cytoplasm from extra- or intracellular vesicles, while ZnT proteins transport zinc in opposite orientations to efflux zinc out of the cytosol [15, 16]. Notably, some of these members also transport other metals like manganese, iron or cadmium, and sometimes in opposite directions [17–21]. In addition to these transporters, metallothioneins (MTs), a group of small, ubiquitous, cysteine-rich Zn-sensing molecules with high affinities for heavy metals, also play an essential role in zinc detoxification and in maintaining zinc homeostasis by buffering and storing intracellular zinc [22, 23]. The transcription of MT is regulated by metal-regulatory transcription factor 1 (MTF-1), which binds to the metal-response elements in the enhancer/promoter regions of MT genes [24–26].
At the organismal level, the physiological function of zinc transporters was shown to be essential for systemic zinc homeostasis and health [27, 28]. For example, in mammals, ZnT1 knockout mice die at an embryonic stage, suggesting a key function for ZnT1 in fetal development [29]; hZIP13 mutations have been shown to cause Ehlers–Danlos syndrome [30]; and human ZIP4 has been found to be responsible for acrodermatitis enteropathica (AE), a human inherited disease caused by impaired zinc uptake [31–33].
Drosophila provides a valid model for studying metal homeostasis [34–41], and the core zinc transport and regulation machinery discovered in yeast and mammals is conserved in the fly [42–44]. The Drosophila genome encodes 10 Zip proteins and 7 ZnT proteins. In comparison, mammalian zinc transporters consist of 14 Zip members and 10 ZnT members. Phylogenetic analysis indicates that these proteins cluster into distinct subgroups [45]. Drosophila also has five MT genes, as well as the zinc-finger-containing transcription factor MTF-1 gene [46–48]. Molecular and cellular functions of Drosophila Zip/ZnT members were systematically analyzed via both over-expression and RNA interference using the GAL4-UAS system [45]. These zinc transporters were further analyzed based on their subcellular localizations and genetic interactions [49–51]. At the organismal level, a set of plasma membrane-resident zinc transporters was identified to participate in dietary zinc absorption. Zip42C.1 (CG9428), Zip42C.2 (CG9430) and Zip89B (CG6898) appear to be involved in uptaking zinc from the lumen into the enterocyte, and ZnT63C (CG17723) and ZnT77C (CG5130) work together to move absorbed zinc from the enterocyte into the circulatory system. The intracellular zinc transporters, such as ZnT86D (CG6672), were not directly involved in dietary zinc absorption [52–54].
Besides zinc absorption, zinc excretion and reabsorption are two other aspects to control body zinc homeostasis. Unfortunately, our understanding of these two important processes is still limited [55, 56]. Drosophila Malpighian tubules are renal organs that perform excretory functions like the kidney in vertebrates. The four tubules originate from the junction between the midgut and hindgut, and are packed as two pairs, with one pair migrating anteriorly and the other pair posteriorly [57–59]. The only player that was known to be involved in zinc excretion in Malpighian tubules is ZnT35C [60], which is homologous to a set of mammalian intracellular zinc transporters including ZnT2, 3, 4, and 8; it was not known what other players might be involved. The zinc exporter ZnT35C was found localized to the apical membrane of tubular cells and proposed to mediate zinc efflux from tubular cells. Interestingly, knocking down ZnT35C also significantly reduced kidney stone formation in a fly calcification model generated by inhibiting xanthine dehydrogenase (Xdh), suggesting that zinc homeostasis in tubules is important to this physiologic process [61].
Theoretically, there should be at least one zinc importer involved in zinc excretion in fly Malpighian tubules. In addition, from a physiological point of view, very likely one or more additional zinc transporters might be involved in zinc reabsorption, preventing the loss of this precious nutritional element under normal or low zinc conditions. In this study, we systematically studied the function of zinc transporters for their roles in zinc excretion in Drosophila. Our results identified Zip71B as a key player in zinc excretion and dZnT1 in zinc reabsorption. This study, together with previous work, began to reveal a primitive picture of players orchestrating the control of zinc excretion in Drosophila Malpighian tubules.
Results
Tubule-specific Zip71B-RNAi and ZnT35C-RNAi flies are sensitive to zinc
Drosophila homologs of mammalian zinc transporters in the Zip and ZnT family include seven potential ZnT and ten Zip proteins [45]. Our previous work systematically analyzed Drosophila zinc transporters for their roles in dietary zinc absorption, enabling us to form a relatively clear picture of the zinc absorption [52, 53]. However, our knowledge is still limited for another important process, zinc excretion. To identify zinc transporters involved in zinc excretion, we used a Malpighian tubule-specific GAL4 line NP1093 (Fig. 1a), crossed to RNAi lines of all potential Zip and ZnT members to knock down each of them, and tested sensitivities of the larvae to zinc overload. Among these zinc transporters, only Zip71B-RNAi and ZnT35C-RNAi flies are hypersensitive to zinc overload (Additional file 1: Table S1).

Tubule-specific modulation of Zip71B and ZnT35C expressions modifies flies’ zinc sensitivity. A Expression pattern of NP1093-GAL4 > UAS-GFP in the gut of third-instar larvae. NP1093-GAL4 is expressed specifically in the Malpighian tubules. Arrows denote the Malpighian tubules. Scale bars = 100 μm. B, C RT-PCR analysis of the knockdown effect of Zip71B-RNAi and ZnT35C-RNAi flies. rp49 was used as internal control. D, E Survival rate of tubule-specific Zip71B-RNAi and ZnT35C-RNAi flies under different food conditions. Both Zip71B-RNAi and ZnT35C-RNAi flies show increased sensitivity toward zinc (means ± SEM, n = 6, P < 0.001; two-tailed Student’s t test), but not other metals or metal chelator EDTA. Genotypes of the flies are NP1093-GAL4/+ for control, Zip71B-RNAi/+; NP1093-GAL4/+ for Zip71B-RNAi, and ZnT35C-RNAi/+; NP1093-GAL4/+ for ZnT35C-RNAi. F (a) RT-PCR analysis of the overexpression effect of Zip71B-OE and ZnT35C-OE flies. rp49 was used as internal control. (b) Statistical analysis of RT-PCR result in (a); n = 3. Genotypes of the flies are NP1093-GAL4/+ for control, UAS-Zip71B/+; NP1093-GAL4/+ for Zip71B-OE, and UAS-ZnT35C/+; NP1093-GAL4/+ for ZnT35C-OE. G Tubule-specific over-expression of Zip71B and ZnT35C confers resistance to high dietary zinc. While both Zip71B-OE and ZnT35C-OE flies showed resistance to high dietary zinc, Zip71B-OE and ZnT35C-OE flies displayed a synergistic effect. Data are presented as means ± SEM, n = 6. * P < 0.05, *** P < 0.001; two-tailed Student’s t test. Genotypes of the flies are NP1093-GAL4/+ for control, UAS-Zip71B/+; NP1093-GAL4/+ for Zip71B-OE, UAS-ZnT35C/+; NP1093-GAL4/+ for ZnT35C-OE, and UAS-ZnT35C/+; NP1093-GAL4/+ for ZnT35C-OE
To confirm the sensitivity phenotype of Zip71B-RNAi is zinc specific, we analyzed the survival rate of Zip71B-RNAi flies on food with zinc or other metal addition. To reduce the possibility that the observed sensitivity arose from off-target effects of RNAi, we used two independent RNAi lines of Zip71B. On normal food, Zip71B-RNAi flies emerged with no aberrances compared to the control. However, as the concentration of zinc in the food increased, the survival rate of Zip71B-RNAi flies decreased dramatically; 4 mM zinc addition prevented almost all Zip71B-RNAi flies from surviving to the adult stage, suggesting that the Zip71B-RNAi flies are extremely sensitive to zinc overload. ZnT35C-RNAi flies also showed increased sensitivity to excess zinc, similar to ZnT35C mutant flies as reported before [60]. Iron, copper or zinc chelator EDTA addition has no obvious effects on Zip71B-RNAi or ZnT35C-RNAi flies, indicating that this sensitivity is specific to zinc overload (Fig. 1d, e).
Next, we examined the effects of tubule-specific overexpression of Zip71B on survival under excessive dietary zinc conditions. Consistently, overexpression of Zip71B confers resistance to high dietary zinc levels (5–15 mM zinc), similar to ZnT35C overexpression. Notably, overexpressing Zip71B and ZnT35C simultaneously enabled significantly stronger resistance to high levels of zinc, suggesting a synergistic effect in zinc excretion (Fig. 1g).
Zinc is accumulated in the body when Zip71B is knocked down in the tubule
The above study showed that tubule-specific RNAi of Zip71B resulted in sensitivity to zinc overload. To analyze possible zinc homeostasis defects of Zip71B-RNAi flies, we measured total zinc content of the whole larvae using inductively coupled plasma-mass spectrometry (ICP-MS). Zinc content in Zip71B-RNAi flies increased dramatically compared to the control flies under either normal or zinc-supplement conditions. As control, levels of other metals including iron, copper and manganese were little changed (Fig. 2a, b). These results suggest that the sensitivity of Zip71B-RNAi larvae to excess zinc is due to failure of zinc excretion in the tubules, resulting in excessive zinc accumulation in the body.

Zinc is accumulated in the body of Zip71B-RNAi fly. a, b Metal contents in Zip71B-RNAi larvae on normal and 1 mM zinc-supplemented food. Tubule-specific RNAi of Zip71B led to a significant increase in whole-body zinc content but no other metal content in both normal food and 1 mM zinc food. Metal content values represent three independent ICP-MS measurements and are normalized to dry body weight; data are presented as means ± SEM, n = 3. *** P < 0.001; two-tailed Student’s t test. c Real-time analysis showing up-regulation of MtnB in Zip71B-RNAi larvae on normal and 1 mM zinc-supplemented food. rp49 was used as the reference gene for normalization. MtnB expression was shown as ratios relative to that of wild-type larvae raised on normal food. Values are presented as means ± SEM; n = 3. ** P < 0.01, *** P < 0.001, two-tailed Student’s t test. Genotypes of the flies are NP1093-GAL4/+ for control and Zip71B-RNAi/+; NP1093-GAL4/+ for Zip71B-RNAi
MT B (MtnB) expression is sensitive to zinc increases in the cell. We thus preformed a real-time PCR analysis to test the transcription level of MtnB in Zip71B-RNAi and wild-type larvae. Compared to that in normal food, both wild-type and Zip71B-RNAi flies show a dramatic increase in MtnB transcription when reared on 1 mM zinc. In the normal food, the MtnB RNA level of Zip71B-RNAi larvae was about 400% of that of the normal wild-type, while in the presence of 1 mM zinc supplementation the MtnB RNA level of Zip71B-RNAi larvae was about 200% of that of the normal wild-type (Fig. 2c). This result is consistent with the notion that an increase of zinc level occurred in Zip71B-RNAi flies.
Endogenous Zip71B localizes to the basolateral membrane of Malpighian tubule cells
According to FlyAtlas data [62], Zip71B transcript is mainly found in the Malpighian tubules. In order to examine its endogenous protein expression pattern, especially in subcellular locations, we generated an antibody against Zip71B. In dissected guts of wild-type third-instar larvae, Zip71B immunoreactivity was most prominent in the Malpighian tubules (Fig. 3a). The specificity of the antibody was confirmed by a significantly reduced staining in Zip71B-RNAi larvae (Fig. 3b).

Zip71B is localized to the basolateral membrane to import zinc into the tubular cells. a–c Expression pattern and intracellular localization of Zip71B. Zip71B expression (immunofluorescence in red) in the Malpighian tubules of control larvae (a) and Zip71B-RNAi larvae (b) grown on normal food. Arrows denote Malpighian tubules. Scale bars = 100 μm. c Zip71B (red) is localized on plasma membrane and restricted to the basolateral side of tubule cells. DAPI (blue) shows the nucleus. Arrowheads indicate the basolateral membranes, and arrows indicate the apical membranes. Scale bars = 20 μm. Genotypes of the flies used are NP1093-GAL4/+ for control and Zip71B-RNAi/+; NP1093-GAL4/+ for Zip71B-RNAi. d, e Zip71B RNAi, in contrast to ZnT35C RNAi, resulted in zinc reduction in Malpighian tubules. Zinc changes are indicated by alterations of MtnB-EYFP fluorescence; (d) is on normal food and (e) is on zinc food. When reared on normal food, Zip71B-RNAi and Zip71B, ZnT35C-RNAi flies showed no obvious difference in fluorescence signal compared to controls, while ZnT35C-RNAi flies showed dramatically increased MtnB-EYFP fluorescence in Malpighian tubules. When reared on 1 mM zinc food. ZnT35C-RNAi flies showed further increased MtnB-EYFP fluorescence signal in tubules compared to control. In Zip71B-RNAi and Zip71B, ZnT35C-RNAi flies, MtnB-EYFP fluorescence was significantly decreased in tubules. Scale bars = 100 μm. Genotypes of the flies used are MtnB-EYFP/+; NP1093-GAL4/+ for the control, MtnB-EYFP/+; Zip71B-RNAi/+; NP1093-GAL4/+ for Zip71B-RNAi, MtnB-EYFP/+; ZnT35C-RNAi/+; NP1093-GAL4/+ for ZnT35C-RNAi, and MtnB-EYFP/+; Zip71B, ZnT35C-RNAi/+; NP1093-GAL4/+ for the double RNAi flies
The subcellular localization of endogenous Zip71B was revealed by immunofluorescence microscopy under high magnification. Zip71B was found to be localized on the plasma membrane and restricted to the basolateral side of the tubular cells (Fig. 3c). Considering that Zip71B is a Zip member, the basolateral distribution of Zip71B is consistent with its role in zinc excretion, i.e., absorbing zinc from circulation into the tubular cells.
Given that immunostaining of Zip71B also revealed slight expression in the intestine, implying a possible excretion role of Zip71B/dZip5 in the midgut, we additionally analyzed the roles of Zip71B/dZip5 and ZnT35C for zinc excretion in the intestine. To address this, we used a midgut-specific driver NP3084. NP3084 predominantly activates expression in the midgut, with undetectable activity in the Malpighian tubes, as confirmed with UAS-GFP as the activity reporter (Additional file 2: Figure S1A). Knocking down Zip71B/dZip5 or ZnT35C in the midgut made the flies slightly more susceptible to zinc overdose (Additional file 2: Figure S1B, C), suggesting that under zinc stress, the Malpighian tubes are the predominant organ for detoxification whereas the intestine likely plays an auxiliary role.
Zip71B acts in concert with ZnT35C to move zinc across the tubular cells in zinc excretion
MtnB-EYFP transgene flies, wherein an enhanced yellow fluorescent protein (EYFP) cDNA is inserted behind the regulatory region of the zinc responsive gene MtnB, can be used as an indicator of intracellular zinc levels. MtnB-EYFP flies have been successfully used in monitoring zinc levels in the midgut constriction before [47, 52–54]. To analyze possible alterations of tubular zinc level in ZnT35C-RNAi and Zip71B-RNAi larvae, we crossed in the MtnB-EYFP transgene. Normal larvae raised on zinc-supplemented food show dramatically induced signal of MtnB-EYFP in both the tubules and posterior midgut, confirming that zinc accumulation in these tissues can be reflected by an increase of MtnB-EYFP fluorescence signal (Fig. 3d, e).
We then tested the MtnB-EYFP fluorescence signal in ZnT35C-RNAi and Zip71B-RNAi larvae. ZnT35C is reported to be localized to the apical membrane of Malpighian tubule cells, so our prediction is that knockdown of ZnT35C might accumulate zinc in tubule cells. Indeed, on both normal and zinc-supplemented food, ZnT35C-RNAi flies showed dramatically increased MtnB-EYFP fluorescence signal in tubules compared to the control (Fig. 3d, e), consistent with its role in moving zinc out from tubule cells to the lumen. In contrast, in Zip71B-RNAi flies, no significant change in MtnB-EYFP signal was seen in the normal food condition compared to the control (Fig. 3d, e). Further, when 1 mM zinc was supplemented in the diet, the MtnB-EYFP signal induction normally observed in the wild-type larvae was much less remarkable in the Zip71B-RNAi tubules, while the MtnB-EYFP signal intensity in the posterior midgut remains unchanged (Fig. 3e). These results indicate that zinc failed to be imported into the tubule cells in Zip71B-RNAi flies.
If Zip71B is responsible for zinc import into, and ZnT35C for zinc efflux out of, the tubule cells, we expect ZnT35C functions downstream of Zip71B. To confirm this epistatic relationship, we checked MtnB-EYFP signal in Zip71B and ZnT35C double RNAi animals. As expected, in the double RNAi larvae, MtnB-EYFP signal was significantly decreased in Malpighian tubules on zinc-supplemented food (Fig. 3e), similar to that in Zip71B-RNAi animals, indicating that Zip71B is indeed epistatic to ZnT35C. Taken together, we propose that Zip71B functions upstream of ZnT35C to transport zinc into tubule cells, and collaborates with ZnT35C to excrete zinc out of the body to maintain zinc homeostasis.
Zip71B expression is regulated by dietary zinc
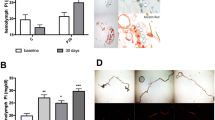
From a physiological point of view, zinc excretion activity increases in response to whole-body zinc stress. We therefore asked whether the expression of Zip71B in Malpighian tubules is subject to zinc regulation. We reared normal flies on standard and zinc supplemented diets, and examined the mRNA level of Zip71B in dissected Malpighian tubules using real-time PCR analysis. The expression levels of Zip71B were higher in flies fed zinc supplemented food, similar to that of the other zinc excretion gene ZnT35C (Fig. 4a). The zinc-inducibility of Zip71B expression was further confirmed by immunostaining in Malpighian tubules (Fig. 4b).

Both Zip71B and ZnT35C expressions are responsive to zinc. a Real-time analysis of mRNA levels of Zip71B and ZnT35C in dissected Malpighian tubules of wild-type animals cultured with normal food or 1 mM zinc-supplemented food. Zip71B was significantly induced by 1 mM zinc (** P < 0.01), as ZnT35C (** P < 0.01). Means ± SEM; n = 3, two-tailed Student’s t test. b Immunostaining reveals that Zip71B is upregulated by dietary zinc addition in Malpighian tubules (arrows). Scale bars = 50 μm
Human ZIP5 can complement loss of Zip71B function
The protein encoded by Zip71B shares the highest homology with human ZIP5 (32% identity and 48% similarity; Fig. 5a). Interestingly, rodent ZIP5 (mZIP5) was also found to be localized to the basolateral membrane of mouse intestinal enterocytes where it plays an important role in zinc excretion [55]. To test whether hZIP5 can functionally rescue the zinc sensitive phenotype of Zip71B-RNAi flies, we generated UAS-hZIP5 transgenic flies, introduced the transgene by crossing them with Zip71B-RNAi flies, and used the NP1093-GAL4 line to drive the tubular expression. As shown in Fig. 5b, expression of hZIP5 can partially but significantly rescue the survival of Zip71B-RNAi flies on zinc-supplemented food, while as control, expression of hZIP4 showed no obvious rescue effect. These results suggest Zip71B is not only most homologous to mammalian ZIP5 in protein sequence, but functions analogously. We thus propose Zip71B as dZip5.

Zip71B loss of function can be complemented by hZIP5 but not hZIP4. a Phylogenetic tree revealing the relationship among human and Drosophila Zip members. Tree was generated and displayed by using MEGA software version 4. Zip71B shares the highest homology with human ZIP5. b hZIP5 expression can rescue the sensitivity of Zip71B-RNAi larvae on food supplemented with 1 mM Zinc. Data are presented as means ± SEM; n = 6. ** P < 0.01, *** P < 0.001, two-tailed Student’s t test. Genotypes of the flies used are UAS-EGFP/+; Zip71B-RNAi/+; NP1093-GAL4/+ (Zip71B-RNAi fly), UAS-hZIP5/+; Zip71B-RNAi/+; NP1093-GAL4/+ (Zip71B-RNAi, UAS-hZIP5 fly), and UAS-hZIP4/+; Zip71B-RNAi/+; NP1093-GAL4/+ (Zip71B-RNAi, UAS-hZIP4 fly)
dZnT1 functions in zinc reabsorption in Drosophila Malpighian tubules
Zinc reabsorption is another important process for zinc homeostasis; however, it remains unknown which zinc transporter is involved in zinc reabsorption in Malpighian tubules. dZnT1 was reported to be localized on the basolateral membrane of Malpighian tubule cells. This distribution pattern of dZnT1 suggested its role in zinc reabsorption [52]. To verify this hypothesis, we examined the survival rate of dZnT1-RNAi flies driven by NP1093-GAL4. While dZnT1-RNAi flies showed no obvious defect on normal food and zinc-chelated food, they were much more resistant to high levels of dietary zinc (such as 10–15 mM) (Fig. 6a).

Tubule-specific dZnT1 RNAi flies display resistance to high dietary zinc. a Survival rate of dZnT1-RNAi flies on different food conditions. There is no effect on survival of dZnT1-RNAi flies on normal and zinc-chelated (100 μM TPEN) food compared to control flies. However, dZnT1-RNAi flies survived significantly better on high dietary zinc (10 and 15 mM zinc) compared to wild-type flies. b Knocking down dZnT1 in the tubule could not suppress zinc sensitivity of Zip71B-RNAi flies. Data are presented as means ± SEM; n = 6. * P < 0.05, ** P < 0.01, two-tailed Student’s t test. Genotypes of the flies used are NP1093-GAL4/+ (control fly), dZnT1-RNAi/+; NP1093-GAL4/+ (dZnT1-RNAi fly), Zip71B-RNAi/+; NP1093-GAL4/+ (Zip71B-RNAi fly) and dZnT1-RNAi/Zip71B-RNAi; NP1093-GAL4/+ (Zip71B, dZnT1-RNAi fly)
To investigate the genetic interaction between dZnT1 and Zip71B (dZip5) in Malpighian tubules, we made dZnT1, Zip71B double RNAi flies by recombining them together. Since dZnT1-RNAi flies are more resistant to zinc overload presumably due to reduced zinc reabsorption, we asked whether knockdown of dZnT1 in tubules could rescue the zinc sensitivity phenotype of Zip71B-RNAi flies. The survival assay on 1 mM zinc-supplemented food showed that the double RNAi flies are still sensitive to excess zinc, to a similar extent as the Zip71B-RNAi flies (Fig. 6b). These results indicate that Zip71B (dZip5) is epistatic to dZnT1, and in this case, dZnT1 functions downstream of Zip71B/dZip5 in tubules for zinc recovery from excretion.
Tubule-specific inhibition of Zip71B (dZip5) and ZnT35C suppresses the formation of tubule stones caused by inhibition of xanthine dehydroxylase
A recent work showed that zinc metabolism in Malpighian tubules is related to ‘kidney stone’ formation in Drosophila; a reduction of the zinc level in the tubular lumen led to decreased stone formation. Three ZnT members (ZnT35C, dZnT1, and ZnT41F), when ubiquitously knocked down, were identified to be involved in this process [61]. We used tubule-specific NP1093 instead of the original ubiquitous GAL4 line to knock down these genes, and found that tubule-specific suppression of ZnT35C, but not dZnT1 and ZnT41F, affected tubular stone formation. These results suggest that dZnT1 and ZnT41F may mediate tubular zinc homeostasis mostly in an indirect way. dZnT1, for example, is a key protein for dietary zinc absorption in addition to its role in the tubular zinc reabsorption. Consistent with its essential role in the tubular zinc homeostasis, tubule-specific knockdown of Zip71B(dZip5) also suppressed the stone formation (Fig. 7).

Tubule-specific inhibition of Zip71B and ZnT35C suppresses the formation of tubule stones. Representative tubule images taken from different groups of flies. Scale bars = 200 μm. Tubule-specific RNAi of Xdh resulted in significantly increased tubule stone formation compared to controls. Knockdown of Zip71B or ZnT35C in Malpighian tubules significantly suppressed the stone formation, while knockdown of dZnT1 or ZnT41F displayed no obvious fly stone reduction. Genotypes of the flies are NP1093-GAL4/+ (control fly), Xdh-RNAi/+; NP1093-GAL4/+ (Xdh-RNAi fly), ZnT35C-RNAi/+; Xdh-RNAi/+; NP1093-GAL4/+ (Xdh-RNAi ZnT35C-RNAi fly), dZnT1-RNAi/+; Xdh-RNAi/+; NP1093-GAL4/+ (Xdh-RNAi dZnT1-RNAi fly), ZnT41F-RNAi/+; Xdh-RNAi/+; NP1093-GAL4/+ (Xdh-RNAi ZnT41F-RNAi fly), Zip71B-RNAi/+; Xdh-RNAi/+; NP1093-GAL4/+ (Xdh-RNAi Zip71B-RNAi fly)
Discussion
Zinc homeostasis in Drosophila is controlled by multiple biological processes. In this work, we focused on zinc excretion in Drosophila Malpighian tubules. Using the tubule-specific GAL4 line NP1093, we knocked down all the ten Zip members and seven ZnT members individually to screen for the zinc transporters participating in zinc excretion in Malpighian tubules. Zip71B/dZip5 was found to be involved in this process. Zip71B-RNAi flies displayed no developmental aberrance on normal media; however, knockdown of Zip71B/dZip5 led to hypersensitivity to zinc overload, similar to ZnT35C-RNAi flies. Fine analysis revealed that, in contrast to ZnT35C RNAi, knockdown of Zip71B/dZip5 resulted in decreased zinc content in the tubules. We concluded that Zip71B/dZip5 acts upstream of ZnT35C to drive zinc excretion from the body, while dZnT1 fulfills the task of zinc reabsorption. All three players work together to regulate the zinc excretory process in the Malpighian tubules.
It is quite remarkable that Zip71B is most homologous to hZIP5, and hZIP5 can partially substitute Drosophila Zip71B’s function in Malpighian tubules while hZIP4 cannot. mZIP5 has relatively high expression level in mouse kidneys [63], just like Zip71B in the Malpighian tubules. Why can hZIP4 not rescue dZip71B/dZip5, given that both are zinc importers? We think this is likely due to the fact that Zip71B/dZip5 has to be located to the basolateral side of the tubule cells to function properly, and hZIP4 may lack the proper signal sequence or structure to be targeted there. If a zinc importer is not correctly positioned, it is hard to fulfill its task of unidirectional mobilization of zinc from the circulation to the tubular cells.
Malpighian tubules, affiliated with the intestine, are renal organs of Drosophila that perform excretory and osmoregulatory functions, similar to kidney in vertebrates and mammals. In zinc excretion studies conducted in mammals, the results of isotropic tracer studies suggest that most zinc is excreted through intestinal cells under normal zinc conditions [64], and mZIP5 is involved in this process [55]. Similar results were also found in C. elegans; ttm-1, one of the Cation Diffusion Facilitator family members, was shown to work in intestinal cells to promote zinc excretion, protecting animals from zinc toxicity [56]. In Drosophila, we found that main zinc excretion occurs in Malpighian tubules. Why this difference? In our experiment, zinc excretion was functionally analyzed under zinc excess or toxic conditions. It is possible that under normal conditions, most zinc excreted will be re-absorbed by the tubules; in this case, little will be really excreted, similar to what was found in mammalian zinc isotope studies. Notably, ZnT35C and Zip71B/dZip5 are also expressed in Drosophila midgut, and functionally participate in zinc excretion as midgut-specific knockdown of these two genes resulted in sensitivity to zinc excess, albeit to a lesser extent. In light of this, we speculate that, under normal conditions, more zinc might be excreted in the midgut while zinc is mostly reabsorbed in the Malpighian tubules; and when zinc is in excess, most zinc is detoxified through the Malpighian tubules; this might also happen in mammals. This notion is supported by the observation that whole body knockout of mZIP5 displays additional zinc accumulation in the liver, compared to intestine-specific knockout of mZIP5 [55], suggesting intestine is not the only important zinc excretion route in mammals. It is possible that mZIP5, as a zinc importer, is involved in mouse kidney zinc excretion. Therefore, current data suggests a somewhat similar process of zinc excretion and control may occur in mammals and the fly. Proteins playing analogous roles to Drosophila ZnT35C and dZnT1 during zinc excretion/reabsorption in mammalian kidneys remain to be identified.
How regulation of Zip5 and Zip71B/dZip5 expressions is achieved might be a complex story, and not yet well characterized. It has been reported that mZIP5 gets internalized and degraded under zinc deficiency conditions [63]. Burke’s lab has reported that Zip71B/dZip5 is a highly efficient transporter, much more potent than other Zip members; it is the only Zip protein that was found to cause zinc toxicity phenotype in eyes when expressed alone [45, 49, 50]. The absence of any detectable Zip71BeGFP signal in larval salivary glands was interpreted by the fact that Zip71B/dZip5 might be down-regulated at the protein level to prevent zinc toxicity caused by excess zinc uptake. We found that Zip71B/dZip5 responds to zinc excess in the Malpighian tubules by an increase in transcription. It is possible that Zip5/Zip71B expression is differentially regulated by zinc in different tissues. Despite this difference, they are alternative strategies to achieve the same goal, i.e., to effectively and economically regulate zinc transport in response to zinc levels or body zinc needs.
Conclusions
Zinc excretion and reabsorption are important processes for whole-body zinc homeostasis control; however, the mechanisms of these controls are not well understood. Drosophila Malpighian tubules, functionally similar to mammalian kidneys, offer a model to study this intricate process. By systematically analyzing the roles of zinc transporters in Drosophila zinc excretion processes, we found that Zip71B (CG10006)/dZip5, which is localized to the basolateral membrane of tubule cells, functions in transporting zinc from body into Malpighian tubules, and acts in concert with apically resident ZnT35C (CG3994) to pump zinc out of the body to maintain zinc homeostasis. Both Zip71B/dZip5 and ZnT35C are subject to zinc regulation. dZnT1 (CG17723), on the other hand, is responsible for zinc reabsorption in Malpighian tubules. This study helps sketch out, for the first time in animals, a model of zinc excretion process (Fig. 8). These findings will be helpful for zinc excretion research in general, and provide a reference for future understanding of zinc metabolism in higher animals.

A model for zinc excretion in Drosophila Malpighian tubules. Zinc homeostasis in Drosophila Malpighian tubules is mediated by a set of plasma membrane-resident zinc transporters. Zip71B (dZip5) is localized to the basolateral membrane of tubule cells and responsible for zinc import from the body into Malpighian tubules. ZnT35C is localized to the apical membrane of tubule cells and pumps zinc out into tubule lumen. Zip71B/dZip5 and ZnT35C act in concert for moving zinc across the tubule cells in zinc excretion. dZnT1 is localized to the basolateral membrane of tubule cells and functions in zinc reabsorption to recover zinc from excretion. The transcription of Zip71B/dZip5 and ZnT35C is subject to the regulation of zinc ion
Methods
Plasmid
UAS-Zip71B and UAS-ZnT35C were generated by PCR amplification of the coding region of Zip71B and ZnT35C from Drosophila cDNA, and cloned into the vector pUAST. UAS-hZIP5 and UAS-hZIP4 were generated by PCR amplification of the coding region of hZIP5 and hZIP4 from human liver cDNA (Gibco BRL, Grand Island, NY, USA) and cloned into pUAST. All the constructs were verified by sequencing.
Fly stocks, culture media, and transgenics
Fly stocks were raised at 18 °C, and all the experiments were performed at 25 °C on standard cornmeal food. When necessary, the food was supplemented with metals or metal chelators at stated concentrations. Fly stocks act-GAL4/CyO (Bloomington #4414), da-GAL4 (Bloomington #8641) and UAS-EGFP (Bloomington#5130) were obtained from the Bloomington Stock Center (Bloomington, IN, USA); NP3084 (DGRC# 113094) and NP1093 (DGRC# 103880) lines were obtained from the Drosophila Genetic Resource Center at the Kyoto Institute of Technology (Kyoto, Japan). The MtnB-EYFP line was a kind gift from Dr. Walter Schaffner (University of Zurich, Zurich, Switzerland). The RNAi lines were provided by the VDRC (Vienna, Austria) or custom made at the Tsinghua Fly Center (Beijing, China), Zip71B-RNAi (VDRC#44539 and TH00155.N), ZnT35C-RNAi (VDRC#103263 and TH00119.N), ZnT41F-RNAi (VDRC#5390), and Xdh-RNAi (THU5626). Transgenic flies were generated in w 1118 background by P-element-mediated transformation.
Fly eclosion assays
NP1093 or NP3084 homozygous flies were crossed with different transgenic flies, and the progeny were reared on food containing different metals or metal chelators, as indicated in each experiment. The density of each vial was controlled to 70 larvae, and the total number of emerged adults of each genotype was counted.
Metal content assay
Flies were reared on normal food or food supplemented with 1 mM zinc from eggs until late third-instar larval stage. About 30 larvae were collected, weighed, and dissolved in 1 mL 65% HNO3, boiled in 100 °C water bath for 8 min, and diluted to 10 mL for metal content analysis with ICP-MS (IRIS Intrepid II XSP, Thermo Electron Corporation, Waltham, MA, USA). Data were collected and analyzed using the native software (ICP Expert).
RNA isolation, semi-quantitative RT-PCR, and quantitative real-time PCR
Total RNA was extracted from adults, larvae or Malpighian tubules from third-instar larvae using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). cDNA was reverse-transcribed from 1 μg total RNA with TransScript First-Strand cDNA Synthesis SuperMix (TransGen Biotech Co., Ltd., Beijing, China), in accordance with the manufacturer’s instructions.
Semi-quantitative RT-PCR was performed using the specific primers corresponding to partial regions of the analyzed genes. Intensity of gel bands was quantitated using ImageJ densitometry. Real-time PCR was performed on StepOnePlusTM (Life Technologies, USA) using SYBR Green PCR Master Mix (Applied Biosystems). Fold change was determined by comparing target gene expression with the reference gene expression (rp49) under the same conditions. The primers used for PCR were: Zip71B, GACCGCTCACCCCAGTAG and ACCCACCGACATGCCCAC; ZnT35C, GTGTAAGCGGTATCAGAACTC and CTCCCAACTATATGCACTACG; mtnB: ATCAGTTCGCCTCAGCCAAG and GCAAACGCACTGGCAATCCT; rp49: GCACCAAGCACTTCATCC and CGATCTCGCCGCAGTAAA.
Antiserum preparation
Rabbit polyclonal antibody was raised against His-Zip71B fusion protein. Briefly, the cDNA fragment encoding the N-terminal 240 aa of Zip71B was synthesized and cloned into pQE80L vector (Qiagen, Hilden, Germany). The recombinant protein was purified and injected into New Zealand White rabbits for antibody generation.
Immunohistochemistry and microscopy
For immunohistochemistry, antibody for Zip71B was preabsorbed with fixed w 1118 embryos before performing staining on fly tissues. Third-instar larvae were collected and dissected in phosphate-buffered saline (PBS), fixed with paraformaldehyde and stained following standard procedures. Anti-Zip71B were used (1:250 dilution) in combination with TRITC -conjugated goat anti-rabbit IgG (ZF-0316, Zhongshan Goldenbridge Biotechnology Co. Ltd., Beijing, China). For nuclear staining, samples were incubated in 50 ng/mL DAPI for 10 min and washed three times in PBS. Slides were mounted with 50% glycerol/PBS.
The immunostainings of Zip71B were examined using a Zeiss LSM710 Meta confocal microscope (Zeiss, Germany). The fluorescence of MtnB-EYFP was captured with an ECLIPSE 80i microscope attached to a DXM1200F digital camera (both Nikon, Tokyo, Japan).
Statistical analysis
All data were analyzed by Student’s t-test using GraphPad Prism version 6.00 for Windows (GraphPad Software, La Jolla California USA). Statistical results were presented as means ± SEM. Asterisks indicate critical levels of significance (* P < 0.05, ** P < 0.01, and *** P < 0.001).
References
Vallee BL, Falchuk KH. The biochemical basis of zinc physiology. Physiol Rev. 1993;73(1):79–118.
Berg JM, Shi Y. The galvanization of biology: a growing appreciation for the roles of zinc. Science. 1996;271(5252):1081–5.
Vallee BL, Auld DS. Cocatalytic zinc motifs in enzyme catalysis. Proc Natl Acad Sci U S A. 1993;90(7):2715–8.
Prasad AS. Zinc: an overview. Nutrition (Burbank, Los Angeles County, Calif). 1995;11(1 Suppl):93–9.
Murakami M, Hirano T. Intracellular zinc homeostasis and zinc signaling. Cancer Sci. 2008;99(8):1515–22.
Fukada T, Yamasaki S, Nishida K, Murakami M, Hirano T. Zinc homeostasis and signaling in health and diseases: Zinc signaling. J Biol Inorg Chem. 2011;16(7):1123–34.
Hambidge M. Human zinc deficiency. J Nutr. 2000;130(5S Supp):1344s–9s.
Fosmire GJ. Zinc toxicity. Am J Clin Nutr. 1990;51(2):225–7.
Chen MD, Song YM, Lin PY. Zinc effects on hyperglycemia and hypoleptinemia in streptozotocin-induced diabetic mice. Horm Metab Res. 2000;32(3):107–9.
Pearson E. Zinc transport and diabetes risk. Nat Genet. 2014;46(4):323–4.
Sturniolo GC, Di Leo V, Ferronato A, D’Odorico A, D’Inca R. Zinc supplementation tightens “leaky gut” in Crohn’s disease. Inflamm Bowel Dis. 2001;7(2):94–8.
Friedlich AL, Lee JY, van Groen T, Cherny RA, Volitakis I, Cole TB, Palmiter RD, Koh JY, Bush AI. Neuronal zinc exchange with the blood vessel wall promotes cerebral amyloid angiopathy in an animal model of Alzheimer’s disease. J Neurosci. 2004;24(13):3453–9.
Lang M, Wang L, Fan Q, Xiao G, Wang X, Zhong Y, Zhou B. Genetic inhibition of solute-linked carrier 39 family transporter 1 ameliorates abeta pathology in a Drosophila model of Alzheimer’s disease. PLoS Genet. 2012;8(4):e1002683.
Huang Y, Wu Z, Cao Y, Lang M, Lu B, Zhou B. Zinc binding directly regulates tau toxicity independent of tau hyperphosphorylation. Cell Rep. 2014;8(3):831–42.
Jeong J, Eide DJ. The SLC39 family of zinc transporters. Mol Asp Med. 2013;34(2-3):612–9.
Huang L, Tepaamorndech S. The SLC30 family of zinc transporters - a review of current understanding of their biological and pathophysiological roles. Mol Asp Med. 2013;34(2-3):548–60.
Chen P, Bowman AB, Mukhopadhyay S, Aschner M. SLC30A10: A novel manganese transporter. Worm. 2015;4(3):e1042648.
Pinilla-Tenas JJ, Sparkman BK, Shawki A, Illing AC, Mitchell CJ, Zhao N, Liuzzi JP, Cousins RJ, Knutson MD, Mackenzie B. Zip14 is a complex broad-scope metal-ion transporter whose functional properties support roles in the cellular uptake of zinc and nontransferrin-bound iron. Am J Physiol Cell Physiol. 2011;301(4):C862–71.
Xiao G, Wan Z, Fan Q, Tang X, Zhou B. The metal transporter ZIP13 supplies iron into the secretory pathway in Drosophila melanogaster. eLife. 2014;3:e03191.
Wang B, Schneider SN, Dragin N, Girijashanker K, Dalton TP, He L, Miller ML, Stringer KF, Soleimani M, Richardson DD, et al. Enhanced cadmium-induced testicular necrosis and renal proximal tubule damage caused by gene-dose increase in a Slc39a8-transgenic mouse line. Am J Physiol Cell Physiol. 2007;292(4):C1523–35.
Valentine RA, Jackson KA, Christie GR, Mathers JC, Taylor PM, Ford D. ZnT5 variant B is a bidirectional zinc transporter and mediates zinc uptake in human intestinal Caco-2 cells. J Biol Chem. 2007;282(19):14389–93.
Dalton T, Fu K, Palmiter RD, Andrews GK. Transgenic mice that overexpress metallothionein-I resist dietary zinc deficiency. J Nutr. 1996;126(4):825–33.
Andrews GK, Geiser J. Expression of the mouse metallothionein-I and -II genes provides a reproductive advantage during maternal dietary zinc deficiency. J Nutr. 1999;129(9):1643–8.
Radtke F, Heuchel R, Georgiev O, Hergersberg M, Gariglio M, Dembic Z, Schaffner W. Cloned transcription factor MTF-1 activates the mouse metallothionein I promoter. EMBO J. 1993;12(4):1355–62.
Palmiter RD. Regulation of metallothionein genes by heavy metals appears to be mediated by a zinc-sensitive inhibitor that interacts with a constitutively active transcription factor, MTF-1. Proc Natl Acad Sci U S A. 1994;91(4):1219–23.
Heuchel R, Radtke F, Georgiev O, Stark G, Aguet M, Schaffner W. The transcription factor MTF-1 is essential for basal and heavy metal-induced metallothionein gene expression. EMBO J. 1994;13(12):2870–5.
Kambe T. Molecular architecture and function of ZnT transporters. Curr Top Membr. 2012;69:199–220.
Lichten LA, Cousins RJ. Mammalian zinc transporters: nutritional and physiologic regulation. Annu Rev Nutr. 2009;29:153–76.
Andrews GK, Wang H, Dey SK, Palmiter RD. Mouse zinc transporter 1 gene provides an essential function during early embryonic development. Genesis. 2004;40(2):74–81.
Fukada T, Civic N, Furuichi T, Shimoda S, Mishima K, Higashiyama H, Idaira Y, Asada Y, Kitamura H, Yamasaki S, et al. The zinc transporter SLC39A13/ZIP13 is required for connective tissue development; its involvement in BMP/TGF-beta signaling pathways. PLoS One. 2008;3(11):e3642.
Wang K, Zhou B, Kuo YM, Zemansky J, Gitschier J. A novel member of a zinc transporter family is defective in acrodermatitis enteropathica. Am J Hum Genet. 2002;71(1):66–73.
Kury S, Dreno B, Bezieau S, Giraudet S, Kharfi M, Kamoun R, Moisan JP. Identification of SLC39A4, a gene involved in acrodermatitis enteropathica. Nat Genet. 2002;31(3):239–40.
Dufner-Beattie J, Wang F, Kuo YM, Gitschier J, Eide D, Andrews GK. The acrodermatitis enteropathica gene ZIP4 encodes a tissue-specific, zinc-regulated zinc transporter in mice. J Biol Chem. 2003;278(35):33474–81.
Norgate M, Lee E, Southon A, Farlow A, Batterham P, Camakaris J, Burke R. Essential roles in development and pigmentation for the Drosophila copper transporter DmATP7. Mol Biol Cell. 2006;17(1):475–84.
Xiao G, Fan Q, Wang X, Zhou B. Huntington disease arises from a combinatory toxicity of polyglutamine and copper binding. Proc Natl Acad Sci U S A. 2013;110(37):14995–5000.
Southon A, Burke R, Camakaris J. What can flies tell us about copper homeostasis? Metallomics. 2013;5(10):1346–56.
Tang X, Zhou B. Ferritin is the key to dietary iron absorption and tissue iron detoxification in Drosophila melanogaster. FASEB J. 2013;27(1):288–98.
Bettedi L, Aslam MF, Szular J, Mandilaras K, Missirlis F. Iron depletion in the intestines of Malvolio mutant flies does not occur in the absence of a multicopper oxidase. J Exp Biol. 2011;214(Pt 6):971–8.
Mandilaras K, Pathmanathan T, Missirlis F. Iron absorption in Drosophila melanogaster. Nutrients. 2013;5(5):1622–47.
Tang X, Zhou B. Iron homeostasis in insects: insights from Drosophila studies. IUBMB life. 2013;65(10):863–72.
Cui Y, Zhao S, Wang X, Zhou B. A novel Drosophila mitochondrial carrier protein acts as a Mg(2+) exporter in fine-tuning mitochondrial Mg(2+) homeostasis. Biochim Biophys Acta. 2016;1863(1):30–9.
Wang X, Zhou B. Dietary zinc absorption: a play of Zips and ZnTs in the gut. IUBMB life. 2010;62(3):176–82.
Richards CD, Burke R. A fly’s eye view of zinc homeostasis: Novel insights into the genetic control of zinc metabolism from Drosophila. Arch Biochem Biophys. 2016;611:142–9.
Xiao G, Zhou B. What can flies tell us about zinc homeostasis? Arch Biochem Biophys. 2016;611:134–41.
Lye JC, Richards CD, Dechen K, Paterson D, de Jonge MD, Howard DL, Warr CG, Burke R. Systematic functional characterization of putative zinc transport genes and identification of zinc toxicosis phenotypes in Drosophila melanogaster. J Exp Biol. 2012;215(Pt 18):3254–65.
Zhang B, Egli D, Georgiev O, Schaffner W. The Drosophila homolog of mammalian zinc finger factor MTF-1 activates transcription in response to heavy metals. Mol Cell Biol. 2001;21(14):4505–14.
Egli D, Yepiskoposyan H, Selvaraj A, Balamurugan K, Rajaram R, Simons A, Multhaup G, Mettler S, Vardanyan A, Georgiev O, et al. A family knockout of all four Drosophila metallothioneins reveals a central role in copper homeostasis and detoxification. Mol Cell Biol. 2006;26(6):2286–96.
Atanesyan L, Gunther V, Celniker SE, Georgiev O, Schaffner W. Characterization of MtnE, the fifth metallothionein member in Drosophila. J Biol Inorg Chem. 2011;16(7):1047–56.
Lye JC, Richards CD, Dechen K, Warr CG, Burke R. In vivo zinc toxicity phenotypes provide a sensitized background that suggests zinc transport activities for most of the Drosophila Zip and ZnT genes. J Biol Inorg Chem. 2013;18(3):323–32.
Dechen K, Richards CD, Lye JC, Hwang JE, Burke R. Compartmentalized zinc deficiency and toxicities caused by ZnT and Zip gene over expression result in specific phenotypes in Drosophila. Int J Biochem Cell Biol. 2015;60:23–33.
Richards CD, Burke R. Local and systemic effects of targeted zinc redistribution in Drosophila neuronal and gastrointestinal tissues. Biometals. 2015;28(6):967–74.
Wang X, Wu Y, Zhou B. Dietary zinc absorption is mediated by ZnT1 in Drosophila melanogaster. FASEB J. 2009;23(8):2650–61.
Qin Q, Wang X, Zhou B. Functional studies of Drosophila zinc transporters reveal the mechanism for dietary zinc absorption and regulation. BMC Biol. 2013;11:101.
Richards CD, Warr CG, Burke R. A role for dZIP89B in Drosophila dietary zinc uptake reveals additional complexity in the zinc absorption process. Biometals. 2015;69:11–9.
Geiser J, De Lisle RC, Andrews GK. The zinc transporter Zip5 (Slc39a5) regulates intestinal zinc excretion and protects the pancreas against zinc toxicity. PLoS One. 2013;8(11):e82149.
Roh HC, Collier S, Deshmukh K, Guthrie J, Robertson JD, Kornfeld K. ttm-1 encodes CDF transporters that excrete zinc from intestinal cells of C. elegans and act in a parallel negative feedback circuit that promotes homeostasis. PLoS Genet. 2013;9(5):e1003522.
Dow JA, Maddrell SH, Gortz A, Skaer NJ, Brogan S, Kaiser K. The malpighian tubules of Drosophila melanogaster: a novel phenotype for studies of fluid secretion and its control. J Exp Biol. 1994;197:421–8.
Beyenbach KW, Skaer H, Dow JA. The developmental, molecular, and transport biology of Malpighian tubules. Annu Rev Entomol. 2010;55:351–74.
Naikkhwah W, O’Donnell MJ. Salt stress alters fluid and ion transport by Malpighian tubules of Drosophila melanogaster: evidence for phenotypic plasticity. J Exp Biol. 2011;214(Pt 20):3443–54.
Yepiskoposyan H, Egli D, Fergestad T, Selvaraj A, Treiber C, Multhaup G, Georgiev O, Schaffner W. Transcriptome response to heavy metal stress in Drosophila reveals a new zinc transporter that confers resistance to zinc. Nucleic Acids Res. 2006;34(17):4866–77.
Chi T, Kim MS, Lang S, Bose N, Kahn A, Flechner L, Blaschko SD, Zee T, Muteliefu G, Bond N, et al. A Drosophila model identifies a critical role for zinc in mineralization for kidney stone disease. PLoS One. 2015;10(5):e0124150.
FlyAtlas: the Drosophila gene expression atlas. http://flyatlas.org/. Accessed 29 May 2007.
Dufner-Beattie J, Kuo YM, Gitschier J, Andrews GK. The adaptive response to dietary zinc in mice involves the differential cellular localization and zinc regulation of the zinc transporters ZIP4 and ZIP5. J Biol Chem. 2004;279(47):49082–90.
Methfessel AH, Spencer H. Zinc metabolism in the rat. II. Secretion of zinc into intestine. J Appl Physiol. 1973;34(1):63–7.
Acknowledgements
We greatly appreciate the fly stocks from the Drosophila Genetic Resource Center at the Kyoto Institute of Technology, the Bloomington Stock, Vienna Drosophila RNAi Center and Tsinghua Fly Center (Beijing, China). This work was supported by grants from the National Science Foundation of China (31123004) and the National Basic Research Program of China (2013CB910700). We also thank Linpu Zhang for help in Drosophila microinjection.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files. Raw data can be found in Additional file 3.
Authors’ contributions
SY and QQ performed the experiments; SY and BZ conceived the experiments; all authors contributed to the data analysis and manuscript writing. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1: Table S1.
Phenotype screening of RNAi lines covering all the 10 ZnT and 7 Zip members for zinc excretion in the Malpighian tubules. Expression was driven by tubule-specific GAL4, NP1093. (TIF 319 kb)
Additional file 2: Figure S1.
Gut-specific Zip71B and ZnT35C RNAi flies display only slight sensitivity to zinc overload. (A) Expression pattern of UAS-GFP driven by NP3084-GAL4 line was visualized in dissected third-instar larvae. NP3084 directs expression all over the midgut, but not in the Malpighian tubules. The yellow arrow denotes the Malpighian tubules, arrowhead denotes the hindgut, and star denotes the midgut. Scale bars = 100 μm. (B, C) Survival rate of gut-specific Zip71B-RNAi and ZnT35C-RNAi flies under different food conditions (means ± SEM, n = 6). Genotypes of the flies are NP3084-GAL4/+ for control, Zip71B-RNAi/+; NP3084-GAL4/+ for Zip71B-RNAi, and ZnT35C-RNAi/+; NP3084-GAL4/+ for ZnT35C-RNAi. (TIF 890 kb)
Additional file 3: Table S2.
Original data for RT-PCR and metal content analyses. (XLSX 13 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Yin, S., Qin, Q. & Zhou, B. Functional studies of Drosophila zinc transporters reveal the mechanism for zinc excretion in Malpighian tubules. BMC Biol 15, 12 (2017). https://doi.org/10.1186/s12915-017-0355-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12915-017-0355-9