
Abstract
"Natura non facit saltum" (nature makes no leap) the Latins used to say, meaning that nature does not like discontinuities. Cells make no exception and indeed any discontinuity in the DNA double helix is promptly detected, triggering an alteration of cell proliferation and an attempt to repair. Yet, linear chromosomes bear DNA ends that are compatible with normal cell proliferation and they escape, under normal conditions, any repair. How telomeres, the chromosomes tips, achieve that is not fully understood. We recently observed that the Rad9/Hus1/Rad1 (911) complex, previously known for its functions in DNA metabolism and DNA damage responses, is constitutively associated with telomeres and plays an important role in their maintenance. Here, we summarize the available data and discuss the potential mechanisms of 911 action at telomeres.
Similar content being viewed by others

Background
Any discontinuity within the DNA double helix is perceived by the cell as a threat to its genetic integrity. Consequently, cells respond promptly to the generation of DNA interruptions by mounting a coordinated set of actions collectively known as the DNA-damage response (DDR) with the intent of arresting cell cycle progression and initiating DNA repair. In higher eukaryotes, the large protein kinases Atm and Atr play central roles in initiating the DDR [1]. Atm responds primarily to double-stranded DNA breaks (DSB). With assistance from the Mrell/Rad50/Nbsl complex, as well as additional mediator and adaptor proteins, Atm phosphorylates and thereby modulates the activity of several substrates involved in cell-cycle control and DNA replication. Bulky DNA lesions, processed DSB, and DNA replication blockage trigger a second DDR pathway involving Atr. These insults lead to the accumulation of single-stranded DNA (ssDNA) coated with the replication protein A (Rpa), a DNA structure that separately attracts Atr and a trimeric complex of Rad9, Rad1, and Hus1 (911) [2]. Atr, in association with its cofactor Atrip, is further stimulated through interactions with Topbpl and, with assistance from the 911 complex and Claspin, phosphorylates and activates the transducer kinase Chkl [3]. Chkl and other Atr targets then mediate a host of DDR outputs, including cell cycle arrest, replication fork stabilization, and DNA repair.
The 911 trimer resembles the toroidal sliding clamp proliferating cell nuclear antigen (PCNA) and is loaded onto DNA by a clamp loader composed of Rad17 and replication factor C (RFC) subunits [4]. While many DDR factors function exclusively in DNA damage signaling, the 911 complex also directly participates in DNA metabolism. The checkpoint clamp physically associates with several factors required for base excision repair (BER) [5–11] and additionally interacts with translesion DNA polymerases [12, 13]. 911 function is also required for homologous recombinational repair (HR) [14], although the precise molecular details of this role have not been elucidated. These findings highlight a broad role for the 911 complex in a variety of DNA transactions and suggest a model in which this trimer may act as a scaffold to recruit checkpoint proteins and DNA modifying enzymes to their sites of action.
Telomeres: an unexpected home for DDR factors
Telomeres, the ends of linear chromosomes, are exceptional, as they are DNA ends that do not normally trigger a robust DDR and are compatible with normal cellular proliferation in checkpoint proficient cells. They contain long stretches of DNA tandem repeats (TTAGGG in vertebrates) and terminate in a 3' protruding single-stranded DNA overhang. Due to the inability of the standard DNA replication machinery to fully replicate DNA ends, many cells maintain their telomeres by the action of telomerase (Tert), a specialized reverse transcriptase that uses its associated RNA component (Terc) as a template to elongate chromosome ends (see [15] for a historical perspective). In the absence of such a mechanism, some tumor cell lines maintain telomeres through homologous recombination, a mechanism known as ALT, for Alternative Lenghtening of the Telomeres [16]. The telomeric repeat sequences are recognized by a specific set of sequence- and structure-specific DNA-binding factors that are essential for many of the key biological features of telomeres, including their ability to avoid triggering a DDR. Some of these factors, such as Trf1 and Trf2, bind to the double-stranded portion of the telomeric DNA and are involved in telomere length regulation and protection, while others, such as Pot1 have important roles in capping the very end of the chromosome by virtue of their ability to recognize the telomeric 3' overhang [17–19]. In addition, the recently discovered KEOPS complex has also been shown to control telomere capping [20]. The intrinsic ability of telomeric tracts to prevent full DDR activation has been demonstrated in the yeast Saccharomyces cerevisiae in which an internal tract of telomeric repeats inhibits DDR signalling [21]. This anti-checkpoint function is likely dependent on telomere DNA binding proteins, and indeed in mammals inactivation of TRF2 triggers a robust DDR emanating from telomeres [22–24]; conversely, Trf2 overexpression can dampen DDR signalling, possibly by directly binding and inhibiting Atm [25].
Despite a tight inhibitory control of DDR at telomeres, mammalian chromosome ends are nevertheless a physiological substrate of the DDR apparatus as the inactivation of genes encoding certain DDR factors involved in DSB repair such as DNA-PK [26–30], Parp-1 [31, 32], Rad51D [33] and Wrn [34], as well as components of the Atm-Mrell/Rad50/Nbsl pathway [35–38] results in telomere dysfunction. Thus, these DDR proteins, which ordinarily would vigorously oppose a free DNA end, actually serve to protect telomere integrity. Specifically how DDR factors are integrated with telomere physiology is, however, still largely unclear [19, 39]. The observation that telomere DNA ends become transiently accessible in the G2 phase of the cell cycle [40] suggests that telomeres can engage DDR factors in a controlled and time-restricted fashion, without nevertheless enforcing a full blown cell-cycle checkpoint. It is therefore possible that telomeres are recognized only by the upstream elements of the DDR machinery, but do not engage the downstream elements of the cascade that would lead to cell-cycle checkpoint enforcement. It remains unclear at which step the transduction of the signal is inhibited. Overall, this strategy may allow DDR factors to mediate a subset of their functions at telomeres in order to promote telomere replication and stability, without triggering other unnecessary or even detrimental responses. These observations also raise the possibility that some DDR proteins might have special functions at telomeres in addition to their established activities in checkpoint signaling and DNA repair.
911, a new component of mammalian telomeres
Recently, we identified an important role for 911 at telomeres in mammals [41]. We reported that the 911 complex is physically associated with telomeric DNA in human and mouse cells. This association engages all three individual subunits of the complex and occurs both in normal and transformed cells, independent of telomerase expression or cell cycle stage. Therefore, 911 is a novel constitutive component of the mammalian telomere. In order to investigate the physiological significance of this association, we analyzed telomere length in cells bearing a genetic deletion of Hus1. Unfortunately, mouse telomere length cannot be reliably measured by Southern blotting techniques, because mouse telomeres are very long (10–50 Kb) and heterogeneous, bearing restriction enzymes cleavage sites interspersed within the telomeric tracts [42]. Techniques based on fluorescence in situ hybridization (FISH) are more reliable because they are insensitive to the polymorphisms associated with mouse telomeres and they can be used to specifically detect and measure only the telomeric repeats signal at chromosome ends [43]. By quantitative use of this technique, we demonstrated that Hus1 deficiency leads to dramatic telomere shortening, from a mean telomere length of 48 Kb in wild type fibroblasts to 27 Kb in Hus1-null fibroblasts. This result was supported by an independent analysis of telomere length by flow-FISH, another FISH-based technology, in unpassaged thymocytes from conditional Hus1 knockout mice. Therefore, 911 is associated with mammalian telomeres and the lack of one of its components, Hus1, leads to a dramatic telomere shortening. These data are in agreement with those showing telomere loss and chromosomal fusions in mouse and human cells lacking Rad9 [14]. Notably, the role for 911 at telomeres is evolutionarily conserved. Caenorhabditis elegans strains lacking HUS-1 or MRT-2, a functional ortholog of mammalian Rad1, display progressive telomere shortening and loss of germ-line immortality [44, 45]. In Schizosaccharomyces pombe orthologues of this complex have been reported to associate with telomeric repeats and are required for telomere length maintenance [46]. The deletion of components of the orthologous complex in Saccharomyces cerevisiae causes mild telomere length changes, although some effects appear to be laboratory or strain specific [47–49].
Possible mechanisms for 911 action at telomeres
A key question raised by the above-mentioned studies centres precisely on how the 911 complex participates in telomere maintenance. To determine whether 911 affected the function of telomerase, the enzyme that catalyzes telomere extension, we compared telomerase activity in wild type cells and cells lacking Hus1 or other 911 components. Excitingly, we discovered that lack of 911 leads to reduced telomerase activity without affecting the expression of the core components of the telomerase complex. We therefore conclude that 911 controls telomerase activity. The notion that 911 promotes telomerase activity is consistent with findings indicating that defects in 911 and Tert have equivalent and non-additive effects on telomere length in C. elegans [50]. Interestingly, other elements of the mammalian 911 signaling cascade such as Atr neither interacted with telomerase nor detectably affected telomerase activity in vitro. These results suggest a unique functional role for 911 in the regulation of telomerase activity. Meanwhile, additional DDR proteins further contribute to telomere homeostasis through other important functions such as promoting T-loop formation and suppressing chromosomal end joining as well as sister telomere exchanges [17, 40].
The structural similarity that the 911 trimer shares with PCNA provides a useful starting point for considering how the 911 complex might influence telomerase activity. Best known for its role as a processivity factor for DNA polymerases during DNA synthesis, PCNA functions as a molecular scaffold that recruits replication and repair proteins to DNA [51]. Evidence suggests that the 911 complex might function in an analogous manner as a landing pad for replication, repair, and checkpoint proteins. For instance, the 911 complex has been shown to directly interact with two different polymerases involved in translesion DNA synthesis (TLS) Pol Kappa [13] and Pol Zeta [12]. Use of the 911 complex as an alternative clamp for damaged templates may provide a mechanism for controlling the usage of TLS polymerases, which have a low fidelity on undamaged DNA templates. Indeed, loss of MEC3 or DDC1, budding yeast orthologs of Hus1 and Rad9, respectively, results in decreased Pol Zeta-dependent spontaneous mutagenesis [12]. These results suggest that the 911 complex may directly control the access of Pol Zeta to damaged DNA, although stimulation of Pol Zeta-mediated translesion synthesis by the checkpoint clamp has not yet been recapitulated in vitro [52]. A scaffolding function also has been identified for the 911 complex during the process of base excision repair. 911 associates with proteins that act throughout the multi-step BER process, including the DNA glycosylase MutY homolog (Myh) [5, 11], the flap endonuclease (Fenl) [7, 10], DNA polymerase Beta [8], and DNA ligase I [6, 9]. In some of these instances, 911 is known to directly stimulate the activity of the associated repair enzyme. Taken together, these findings establish a recurrent theme suggesting that a primary function of the 911 complex may be to recruit DNA modifying enzymes to appropriate substrates and to stimulate their activities.
When applied to the issue of telomere maintenance, this model would suggest that the 911 complex might act by directly binding and regulating the activity of telomerase. Consistent with this possibility, in a number of tests, 911 biochemically associates with catalytically active telomerase complex. The 911 complex is appropriately situated for such an interaction, as in vitro experiments suggest that 5'-recessed DNA ends are preferential sites for loading of the 911 complex, making telomeres a natural candidate substrate in vivo as well [53, 54]. An association between 911 and telomerase could conceivably recruit telomerase to telomeres and/or enhance its activity. The latter could encompass improvements to telomerase processivity, similar to the effect of PCNA on DNA polymerase. However, it is worth noticing that the association of 911 with DNA polymerase Beta does not increase processivity but instead enhances primer utilization [8].
Although the available data are consistent with a model in which 911 directly interacts with and stimulates telomerase (as shown in figure 1), it remains possible that 911 instead impacts telomerase through an indirect mechanism. 911 could fulfill such a role by helping create a telomeric substrate that is more accessible to telomerase or can be more readily extended. DNA polymerases are highly sensitive to misaligned primer/template substrates, terminal mismatches and other abnormal DNA structures. At telomeres, the G-rich single-stranded DNA overhang is prone to form G-quartet structures that are inhibitory for telomerase activity [55, 56]. Optimal telomerase activity might require a mechanism for resolving such impediments at chromosome ends, a role that could be fulfilled by 911. In addition, the observation that DNA polymerase Beta localizes at telomeres [57] suggests a potential additional contribution of 911 to telomeric DNA replication.

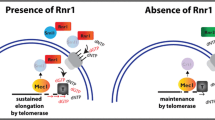
Models for the function of the 911 complex at telomeres. During most of the cell cycle, telomeres are believed to be folded into T-loop structures in which the 3' single-stranded overhang is tucked back into the adjacent double-stranded DNA, forming a D-loop. 911 is localized to telomeres in both quiescent and replicating cells, suggesting that it associates with T-loop structures. The precise DNA structure necessary for 911 loading at telomeres is unknown. In vitro studies suggest preferential loading of the 911 complex onto 5' recessed DNA ends, but loading onto 3' recessed DNA ends is observed under some conditions. The telomere shortening observed in 911-deficient cells as well as additional data implicate 911 in telomere replication mechanisms, including telomerase-based and alternative lengthening of telomeres (ALT) recombinational mechanisms. The speculative models shown illustrate direct interactions between 911 and telomerase or DNA polymerase, but the precise molecular function of 911 in these processes remains to be determined (see text for details).
Notably, the rate of telomere shortening that we observe in Hus1-null cells is higher than that reported for mice lacking telomerase, suggesting that additional mechanisms of telomere shortening might be at play in the absence of the 911 complex. Rapid telomere shortening could be caused by nucleolytic degradation or aberrant recombination events at uncapped telomeres. The 911 complex has been implicated previously in telomeric recombination. In ALT cells, telomeres are maintained by a recombinational mechanism in which short telomeres invade homologous telomeric sequences and use this as a template for lengthening [58]. Interestingly, ALT cells have been shown to display cytologically detectable accumulation of 911 at telomeres [59], suggesting that, in the absence of telomerase, the presence of the 911 complex at telomeres could promote telomere lengthening through HR. Moreover, Rad9 or Hus1 inactivation in mammals is associated with defective HR [6, 14], which in some cases might result in increased usage of alternative, error-prone mechanisms. Thus, aberrant recombination events at telomeres could contribute to rapid telomere shortening in cells lacking the 911 complex.
Finally, it is worth considering that the presence of a poorly functioning telomerase on DNA might be more detrimental to telomere stability than not having telomerase at all. A poorly processive polymerase on DNA can cause DNA replication fork stalling and such an event in checkpoint-deficient cells has been shown to lead to fork reversal and hyper-recombination [60]. In agreement with this, budding yeast cells replicating with a limited amount of DNA polymerase show chromosomal instability at specific genomic regions [61]. Stalled forks promote checkpoint activation by exposing significant amounts of single-stranded DNA (ssDNA) coated by Rpa [60, 62]. These Rpa-ssDNA complexes recruit Atr and 911 to stalled forks and cause the activation of replication checkpoints [63]. In checkpoint mutants, stalled forks rapidly degenerate, accumulating gapped and hemireplicated molecules that often end up processed by unscheduled recombinogenic events [64]. Therefore, it is possible that in cells lacking Hus1 the presence at the telomere of a poorly functioning telomerase complex leads to generation of aberrant DNA structures that can result in the rapid loss of telomeric repeats through low fidelity DNA recombination and processing events.
Summary
The linear structure of eukaryotic chromosomes creates a requirement for special mechanisms to replicate and protect chromosome ends. Over the past several years, a seemingly unlikely caretaker of telomeres has emerged. DDR proteins, which normally act to eradicate free DNA ends, have proven to be critical to telomere homeostasis. As reviewed here, the 911 complex is the latest example of a mammalian DDR factor that localizes to telomeres and acts to maintain telomere integrity. Consistent with results from S. pombe and C. elegans, loss of 911 components leads to telomere shortening and end-to-end chromosome fusions. Moreover, the 911 complex can be co-immunoprecipitated with telomerase activity and is required for optimal telomerase activity in in vitro assays. The precise molecular function of 911 at telomeres, which may involve stimulation of telomerase directly through a physical interaction or indirectly through effects on the telomeric DNA substrate, has yet to be completely resolved. This and other aspects of telomere maintenance undoubtedly will continue to be areas of intense interest given the principal role of telomeres as determinants of cellular lifespan and genome stability.
References
Bakkenist CJ, Kastan MB: DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature 2003, 421: 499–506. 10.1038/nature01368
Cortez D: Unwind and slow down: checkpoint activation by helicase and polymerase uncoupling. Genes Dev 2005, 19: 1007–1012. 10.1101/gad.1316905
Shechter D, Costanzo V, Gautier J: ATR and ATM regulate the timing of DNA replication origin firing. Nat Cell Biol 2004.
Parrilla-Castellar ER, Arlander SJH, Karnitz L: Dial 9–1-1 for DNA damage: the Rad9- Hus1-Rad1 (9–1-1) clamp complex. DNA Repair 2004, 3: 1009–1014. 10.1016/j.dnarep.2004.03.032
Shi G, Chang DY, Cheng CC, Guan X, Venclovas C, Lu AL: Physical and functional interactions between MutY glycosylase homologue (MYH) and checkpoint proteins Rad9-Rad1-Hus1. Biochem J 2006, 400: 53–62. 10.1042/BJ20060774
Wang W, Lindsey-Boltz LA, Sancar A, Bambara RA: Mechanism of stimulation of human DNA ligase I by the Rad9-Rad1-Hus1 checkpoint complex. J Biol Chem 2006, 281: 20865–20872. 10.1074/jbc.M602289200
Wang W, Brandt P, Rossi ML, Lindsey-Boltz L, Podust V, Fanning E, Sancar A, Bambara RA: The human Rad9-Rad1-Hus1 checkpoint complex stimulates flap endonuclease 1. PNAS 2004, 101: 16762–16767. 10.1073/pnas.0407686101
Toueille M, El-Andaloussi N, Frouin I, Freire R, Funk D, Shevelev I, Friedrich-Heineken E, Villani G, Hottiger MO, Hubscher U: The human Rad9/Rad1/Hus1 damage sensor clamp interacts with DNA polymerase {beta} and increases its DNA substrate utilisation efficiency: implications for DNA repair. Nucl Acids Res 2004, 32: 3316–3324. 10.1093/nar/gkh652
Smirnova E, Toueille M, Markkanen E, Hubscher U: The human checkpoint sensor and alternative DNA clamp Rad9-Rad1-Hus1 modulates the activity of DNA ligase I, a component of the long-patch base excision repair machinery. Biochem J 2005, 389: 13–17. 10.1042/BJ20050211
Friedrich-Heineken E, Toueille M, Tannler B, Burki C, Ferrari E, Hottiger MO, Hubscher U: The two DNA clamps Rad9/Rad1/Hus1 complex and proliferating cell nuclear antigen differentially regulate flap endonuclease 1 activity. J Mol Biol 2005, 353: 980–989. 10.1016/j.jmb.2005.09.018
Chang DY, Lu AL: Interaction of checkpoint proteins Hus1/Rad1/Rad9 with DNA base excision repair enzyme MutY homolog in fission yeast, Schizosaccharomyces pombe. J Biol Chem 2005, 280: 408–417.
Sabbioneda S, Minesinger BK, Giannattasio M, Plevani P, Muzi-Falconi M, Jinks-Robertson S: The 9–1-1 Checkpoint Clamp Physically Interacts with Pol{zeta} and Is Partially Required for Spontaneous Pol{zeta}-dependent Mutagenesis in Saccharomyces cerevisiae. J Biol Chem 2005, 280: 38657–38665. 10.1074/jbc.M507638200
Kai M, Wang TS: Checkpoint activation regulates mutagenic translesion synthesis. Genes Dev 2003, 17: 64–76. 10.1101/gad.1043203
Pandita RK, Sharma GG, Laszlo A, Hopkins KM, Davey S, Chakhparonian M, Gupta A, Wellinger RJ, Zhang J, Powell SN, et al.: Mammalian Rad9 Plays a Role in Telomere Stability, S- and G2-Phase-Specific Cell Survival, and Homologous Recombinational Repair. Mol Cell Biol 2006, 26: 1850–1864. 10.1128/MCB.26.5.1850-1864.2006
Blackburn EH, Greider CW, Szostak JW: Telomeres and telomerase: the path from maize, Tetrahymena and yeast to human cancer and aging. Nat Med 2006, 12: 1133–1138. 10.1038/nm1006-1133
Varley H, Pickett HA, Foxon JL, Reddel RR, Royle NJ: Molecular characterization of inter-telomere and intra-telomere mutations in human ALT cells. Nat Genet 2002, 30: 301–305. 10.1038/ng834
de Lange T: Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev 2005, 19: 2100–2110. 10.1101/gad.1346005
Price CM: Stirring the POT1: surprises in telomere protection. Nat Struct Mol Biol 2006, 13: 673–674. 10.1038/nsmb0806-673
d'Adda di Fagagna F, Teo S-H, Jackson SP: Functional links between telomeres and proteins of the DNA-damage response. Genes Dev 2004, 18: 1781–1799. 10.1101/gad.1214504
Downey M, Houlsworth R, Maringele L, Rollie A, Brehme M, Galicia S, Guillard S, Partington M, Zubko MK, Krogan NJ, et al.: A genome-wide screen identifies the evolutionarily conserved KEOPS complex as a telomere regulator. Cell 2006, 124: 1155–1168. 10.1016/j.cell.2005.12.044
Michelson RJ, Rosenstein S, Weinert T: A telomeric repeat sequence adjacent to a DNA double-stranded break produces an anticheckpoint. Genes Dev 2005, 19: 2546–2559. 10.1101/gad.1293805
Karlseder J, Broccoli D, Dai Y, Hardy S, de Lange T: p53- and ATM-Dependent Apoptosis Induced by Telomeres Lacking TRF2. Science 1999, 283: 1321–1325. 10.1126/science.283.5406.1321
Karlseder J, Smogorzewska A, de Lange T: Senescence induced by altered telomere state, not telomere loss. Science 2002, 295: 2446–2449. 10.1126/science.1069523
d'Adda di Fagagna F, Reaper PM, Clay-Farrace L, Fiegler H, Carr P, Von Zglinicki T, Saretzki G, Carter NP, Jackson SP: A DNA damage checkpoint response in telomere- initiated senescence. Nature 2003, 426: 194–198. 10.1038/nature02118
Karlseder J, Hoke K, Mirzoeva OK, Bakkenist C, Kastan MB, Petrini JH, Lange Td T: The Telomeric Protein TRF2 Binds the ATM Kinase and Can Inhibit the ATM- Dependent DNA Damage Response. PLoS Biol 2004, 2: E240. 10.1371/journal.pbio.0020240
Hande P, Slijepcevic P, Silver A, Bouffler S, van Buul P, Bryant P, Lansdorp P: Elongated telomeres in scid mice. Genomics 1999, 56: 221–223. 10.1006/geno.1998.5668
Gilley D, Tanaka H, Hande MP, Kurimasa A, Li GC, Oshimura M, Chen DJ: DNA-PKcs is critical for telomere capping. Proc Natl Acad Sci USA 2001, 98: 15084–15088. 10.1073/pnas.261574698
d'Adda di Fagagna F, Hande MP, Tong WM, Roth D, Lansdorp PM, Wang ZQ, Jackson SP: Effects of DNA nonhomologous end-joining factors on telomere length and chromosomal stability in mammalian cells. Curr Biol 2001, 11: 1192–1196. 10.1016/S0960-9822(01)00328-1
Espejel S, Martin M, Klatt P, Martin-Caballero J, Flores JM, Blasco MA: Shorter telomeres, accelerated ageing and increased lymphoma in DNA-PKcs-deficient mice. EMBO Rep 2004, 5: 503–509. 10.1038/sj.embor.7400127
Samper E, Goytisolo FA, Slijepcevic P, van Buul PP, Blasco MA: Mammalian Ku86 protein prevents telomeric fusions independently of the length of TTAGGG repeats and the G-strand overhang. EMBO Rep 2000, 1: 244–252. 10.1093/embo-reports/kvd051
d'Adda di Fagagna F, Hande MP, Tong WM, Lansdorp PM, Wang ZQ, Jackson SP: Functions of poly (ADP-ribose) polymerase in controlling telomere length and chromosomal stability. Nat Genet 1999, 23: 76–80.
Samper E, Goytisolo FA, Menissier-de Murcia J, Gonzalez-Suarez E, Cigudosa JC, de Murcia G, Blasco MA: Normal telomere length and chromosomal end capping in poly (ADP-ribose) polymerase-deficient mice and primary cells despite increased chromosomal instability. J Cell Biol 2001, 154: 49–60. 10.1083/jcb.200103049
Tarsounas M, Munoz P, Claas A, Smiraldo PG, Pittman DL, Blasco MA, West SC: Telomere Maintenance Requires the RAD51D Recombination/Repair Protein. Cell 2004, 117: 337–347. 10.1016/S0092-8674(04)00337-X
Crabbe L, Verdun RE, Haggblom CI, Karlseder J: Defective Telomere Lagging Strand Synthesis in Cells Lacking WRN Helicase Activity. Science 2004, 306: 1951–1953. 10.1126/science.1103619
Zhu XD, Kuster B, Mann M, Petrini JH, de Lange T: Cell-cycle-regulated association of RAD50/MRE11/NBS1 with TRF2 and human telomeres. Nat Genet 2000, 25: 347–352. 10.1038/77139
Ranganathan V, Heine WF, Ciccone DN, Rudolph KL, Wu X, Chang S, Hai H, Ahearn IM, Livingston DM, Resnick I, et al.: Rescue of a telomere length defect of Nijmegen breakage syndrome cells requires NBS and telomerase catalytic subunit. Curr Biol 2001, 11: 962–966. 10.1016/S0960-9822(01)00267-6
Metcalfe JA, Parkhill J, Campbell L, Stacey M, Biggs P, Byrd PJ, Taylor AM: Accelerated telomere shortening in ataxia telangiectasia. Nat Genet 1996, 13: 350–353. 10.1038/ng0796-350
Hande MP, Balajee AS, Tchirkov A, Wynshaw-Boris A, Lansdorp PM: Extra- chromosomal telomeric DNA in cells from Atm(-/-) mice and patients with ataxia- telangiectasia. Hum Mol Genet 2001, 10: 519–528. 10.1093/hmg/10.5.519
Bertuch AA, Lundblad V: The maintenance and masking of chromosome termini. Current Opinion in Cell Biology 2006, 18: 247–253. 10.1016/j.ceb.2006.04.005
Verdun RE, Crabbe L, Haggblom C, Karlseder J: Functional Human Telomeres Are Recognized as DNA Damage in G2 of the Cell Cycle. Molecular Cell 2005, 20: 551–561. 10.1016/j.molcel.2005.09.024
Francia S, Weiss RS, Hande MP, Freire R, d'Adda di Fagagna F: Telomere and Telomerase Modulation by the Mammalian Rad9/Rad1/Hus1 DNA-Damage- Checkpoint Complex. Curr Biol 2006, 16: 1551–1558. 10.1016/j.cub.2006.06.066
Kipling D, Cooke HJ: Hypervariable ultra-long telomeres in mice. Nature 1990, 347: 400–402. 10.1038/347400a0
Blasco MA, Lee HW, Hande MP, Samper E, Lansdorp PM, DePinho RA, Greider CW: Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell 1997, 91: 25–34. 10.1016/S0092-8674(01)80006-4
Hofmann ER, Milstein S, Boulton SJ, Ye M, Hofmann JJ, Stergiou L, Gartner A, Vidal M, Hengartner MO: Caenorhabditis elegans HUS-1 Is a DNA Damage Checkpoint Protein Required for Genome Stability and EGL-1-Mediated Apoptosis. Curr Biol 2002, 12: 1908–1918. 10.1016/S0960-9822(02)01262-9
Ahmed S, Hodgkin J: MRT-2 checkpoint protein is required for germline immortality and telomere replication in C. elegans. Nature 2000, 403: 159–164. 10.1038/35003120
Nakamura TM, Moser BA, Russell P: Telomere binding of checkpoint sensor and DNA repair proteins contributes to maintenance of functional fission yeast telomeres. Genetics 2002, 161: 1437–1452.
Corda Y, Schramke V, Longhese MP, Smokvina T, Paciotti V, Brevet V, Gilson E, Geli V: Interaction between Setlp and checkpoint protein Mec3p in DNA repair and telomere functions. Nat Genet 1999, 21: 204–208. 10.1038/5991
Longhese MP, Paciotti V, Neecke H, Lucchini G: Checkpoint proteins influence telomeric silencing and length maintenance in budding yeast. Genetics 2000, 155: 1577–1591.
Grandin N, Damon C, Charbonneau M: Cdcl3 prevents telomere uncapping and Rad50-dependent homologous recombination. Embo J 2001, 20: 6127–6139. 10.1093/emboj/20.21.6127
Meier B, Clejan I, Liu Y, Lowden M, Gartner A, Hodgkin J, Ahmed S: trt-1 Is the Caenorhabditis elegans Catalytic Subunit of Telomerase. PLoS Genetics 2006, 2: el8. 10.1371/journal.pgen.0020018
Kelman Z, Hurwitz J: Protein-PCNA interactions: a DNA-scanning mechanism? Trends Biochem Sci 1998, 23: 236–238. 10.1016/S0968-0004(98)01223-7
Garg P, Stith CM, Majka J, Burgers PM: Proliferating Cell Nuclear Antigen Promotes Translesion Synthesis by DNA Polymerase {zeta}. J Biol Chem 2005, 280: 23446–23450. 10.1074/jbc.C500173200
Ellison V, Stillman B: Biochemical Characterization of DNA Damage Checkpoint Complexes: Clamp Loader and Clamp Complexes with Specificity for 5' Recessed DNA. PLoS Biol 2003, 1: E33. 10.1371/journal.pbio.0000033
Majka J, Binz SK, Wold MS, Burgers PM: Replication protein A directs loading of the DNA damage checkpoint clamp to 5'-DNA junctions. J Biol Chem 2006, 281: 27855–27861. 10.1074/jbc.M605176200
Williamson JR: G-quartet structures in telomeric DNA. Annu Rev Biophys Biomol Struct 1994, 23: 703–730. 10.1146/annurev.bb.23.060194.003415
Zahler AM, Williamson JR, Cech TR, Prescott DM: Inhibition of telomerase by G- quartet DNA structures. Nature 1991, 350: 718–720. 10.1038/350718a0
Verdun RE, Karlseder J: The DNA Damage Machinery and Homologous Recombination Pathway Act Consecutively to Protect Human Telomeres. Cell 2006, 127: 709–720. 10.1016/j.cell.2006.09.034
Henson JD, Neumann AA, Yeager TR, Reddel RR: Alternative lengthening of telomeres in mammalian cells. Oncogene 2002, 21: 598–610. 10.1038/sj.onc.1205058
Nabetani A, Yokoyama O, Ishikawa F: Localization of hRad9, hHus1, hRad1, and hRad17 and Caffeine-sensitive DNA Replication at the Alternative Lengthening of Telomeres-associated Promyelocytic Leukemia Body. J Biol Chem 2004, 279: 25849–25857. 10.1074/jbc.M312652200
Sogo JM, Lopes M, Foiani M: Fork Reversal and ssDNA Accumulation at Stalled Replication Forks Owing to Checkpoint Defects. Science 2002, 297: 599–602. 10.1126/science.1074023
Lemoine FJ, Degtyareva NP, Lobachev K, Petes TD: Chromosomal translocations in yeast induced by low levels of DNA polymerase a model for chromosome fragile sites. Cell 2005, 120: 587–598. 10.1016/j.cell.2004.12.039
Zou L, Elledge SJ: Sensing DNA Damage Through ATRIP Recognition of RPA- ssDNA Complexes. Science 2003, 300: 1542–1548. 10.1126/science.1083430
Tercero JA, Longhese MP, Diffley JF: A central role for DNA replication forks in checkpoint activation and response. Mol Cell 2003, 11: 1323–1336. 10.1016/S1097-2765(03)00169-2
Liberi G, Maffioletti G, Lucca C, Chiolo I, Baryshnikova A, Cotta-Ramusino C, Lopes M, Pellicioli A, Haber JE, Foiani M: Rad51-dependent DNA structures accumulate at damaged replication forks in sgsl mutants defective in the yeast ortholog of BLM RecQ helicase. Genes Dev 2005, 19: 339–350. 10.1101/gad.322605
Acknowledgements
We thank members of the d'Adda di Fagagna and Weiss labs for discussions. F.d'A.d.F. is supported by AIRC, S.F. is a SEMM student (European School of Molecular Medicine) and R.S.W. is supported by National Institutes of Health grant RO1 CA108773.
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Francia, S., Weiss, R.S. & d'Adda di Fagagna, F. Need telomere maintenance? Call 911. Cell Div 2, 3 (2007). https://doi.org/10.1186/1747-1028-2-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1747-1028-2-3