
Abstract
The subkingdom Bilateria encompasses the overwhelming majority of animals, including all but four early-branching phyla: Porifera, Ctenophora, Placozoa, and Cnidaria. On average, these early-branching phyla have fewer cell types, tissues, and organs, and are considered to be significantly less specialized along their primary body axis. As such, they present an attractive outgroup from which to investigate how evolutionary changes in the genetic toolkit may have contributed to the emergence of the complex animal body plans of the Bilateria. This review offers an up-to-date glimpse of genome-scale comparisons between bilaterians and these early-diverging taxa. Specifically, we examine these data in the context of how they may explain the evolutionary development of primary body axes and axial symmetry across the Metazoa. Next, we re-evaluate the validity and evolutionary genomic relevance of the zootype hypothesis, which defines an animal by a specific spatial pattern of gene expression. Finally, we extend the hypothesis that Wnt genes may be the earliest primary body axis patterning mechanism by suggesting that Hox genes were co-opted into this patterning network prior to the last common ancestor of cnidarians and bilaterians.
Open peer review
Reviewed by Pierre Pontarotti, Gáspár Jékely, and L Aravind. For the full reviews, please go to the Reviewers' comments section.
Similar content being viewed by others

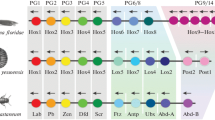

Background
The thirty-plus metazoan phyla are each characterized by a distinct "Bauplan" (or body plan; see [1, 2]). A major challenge facing evolutionary biologists lies in understanding the evolution of major animal features such as body symmetry, germ layers, body cavities, skeletal systems, and nervous systems that comprise these disparate metazoan body plans [3]. In approaching these questions, it is especially useful to consider the earliest-branching metazoan phyla – specifically, Porifera (sponges), Ctenophora (ctenophores), Placozoa (Trichoplax), and Cnidaria (e.g., sea anemones, corals, and jellyfish) (Figure 1). The origin of animals dates back to over 600 million years ago and perhaps much earlier [4]. Fossil evidence and molecular evidence suggest bilaterian animals radiated during the Cambrian period some 500–550 million years ago [5]. In the interval between these two events, the Symplasma and the Cellularia (both sub-phyla within Porifera), the Ctenophora, the Cnidaria, and the Placozoa emerged in succession as independent lineages (see Figure 1; see also Table 1 and references therein). Each of these early branching lineages offers a unique perspective into early animal evolution.

Evolutionary relationships of early-diverging metazoan lineages (Symplasma, Cellularia, Ctenophora, Placozoa, Anthozoa, and Medusozoa) with Bilateria and outgroups (Plantae, Fungi, and Choanoflagellata). Tree topology is based on [77]. Dotted line indicates uncertainty as to the placement of the placozoan branch. Arrows indicate the earliest known appearance of the specified group of genes.
One key morphological feature that has contributed to the traditional super-phyletic relationships of animals is symmetry about a primary body axis. A comprehensive analysis of animal anatomy produced by Beklemishev concluded that all animals are organized by a primary body axis (alternatively referred to as "anterior/posterior", "apical/basal", and "oral/aboral") [6]. This axis is especially evident during embryogenesis and larval stages. It is debated though whether this monaxonal symmetry is homologous throughout the Metazoa and at what point (or points) in animal evolution bilateral symmetry emerged.
Axial symmetry and the porifera
Sponges are often described as being asymmetric or containing imperfect radial symmetry [6]. While it is true that adult sponges are often asymmetrically shaped by their environment, sponge larvae are usually symmetrical about their primary body axis, especially in the Hexactinelida and Calcarea classes [6, 7] (Figure 2A). Moreover, many of the hallmark features of metazoan ontogeny (that is, cleavage, blastulation, and gastrulation) occur in sponges [8]. This fact is of critical importance when trying to reconstruct the body plan of the crown ancestor to metazoans. Recent reports have revealed that the phylum Porifera is genetically more complex than was anticipated [9–11]. They possess many of the signaling pathways and transcription factors that are involved in patterning the axes of bilaterians. There has yet to be extensive in situ hybridization assays involving sponges, but early results from the demosponge Amphimedon queenslandica suggest that transcription factors have the potential to specify distinct regions of the developing sponge [11]. Further insight into the processes responsible for patterning the sponge primary body axis will be essential in understanding the evolution of animal body plans.

Phylogenetic framework of primary body axis symmetry. (A) Radial symmetry of a sponge larva. (B) Top down view of Pleurobrachia (Ctenophora). The tentacles and anal pores of the ctenophore disrupt the octoradiate symmetry established by the 8 ciliary combs. (C) Bilateral symmetry of Actinia as seen through a cross-section of Actinia. The decemradiate symmetry defined by the septa is broken by a single siphonoglyph. (D) Bilateral symmetry defined by numerous structures as seen through the Cross-section of an earthworm. Abbreviations: ms = mesenteries, rm = retractor muscle, dbv = dorsal blood vessel, mwi = muscular wall of intestine, fiw = fold of intestinal wall, ct = cuticle, gc = gland cell, ep = epidermis, cm = circular muscles, lm = longitudinal muscles, sbv = subneural blood vessel, vnc = ventral nerve chord, vbv = ventral blood vessel, cf = ciliated funnel, exp = excretory pore, np = nephridium, br = bristle (only two of the 4 pairs of bristles are shown), pr = peritoneum, cc = chloragen cells. Illustrations in A, B, and D are after [78] with permission from University of Chicago Press. Illustration in C after [6].
Axial symmetry and the "radiata"
The superphyletic designation Radiata was originally proposed by Cuvier to encompass the so-called radially-symmetric animals (for example, jellyfish, polyps, starfish, sea urchins, and some Protozoa) [12]. As new evidence has surfaced, the composition of this group has been modified several times (c.f. [13, 14]). The term Radiata has also been frequently used to describe only the phyla Cnidaria and Ctenophora (for example, [15]). The rotating membership and the polyphyletic nature of these phyla (see Figure 1) contribute to the obvious confusion surrounding this term. However, the most confounding aspect of the term Radiata is that few species within Cnidaria and Ctenophora (the two consistent members of this group) actually exhibit true radial symmetry (that is, indefinite multiradiate symmetry), with many species within the group (particularly anthozoans) instead exhibiting bilateral symmetry.
Multiple examples of asymmetry, bilaterality, biradiality, and tetraradiality can be found collectively within the Cnidaria and Ctenophora (reviewed in [3, 15–17]). In particular, many species within the Anthozoa (representing all three subclasses) and several within the Hydrozoa show clear signs of bilateral symmetry [6, 18]. It is possible that this bilateral symmetry derived independently within these lineages, as suggested by Hyman and Beklemishev [6, 19]. However, these authors were basing their conclusions on an animal phylogeny that placed the biradial ctenophores sister to the Bilateria. Furthermore, their understanding of cnidarian evolution was substantially different. Evidence based on both genomic content [20] and mitochondrial structure [21] suggest that the anthozoan biphasic lifestyle may be more primitive than medusozoans; this is in contrast to the more popular view at that time, a view put forth by Hyman that advocated the reverse scenario [19]. This recent shift in phylogenetic perspective supports the turbellarian theory, developed by Hadzi [22] and advocated by both Hand [23] and Willmer [3]: that radial symmetry is a derived feature within the Cnidaria, and suggests that bilateral symmetry evolved prior to the cnidarian-bilaterian ancestor. Data from the sea anemone Nematostella vectensis show that similar molecular mechanisms, including Hox, Dpp and Wnt genes [24–28], are involved in patterning the primary and secondary axes of bilaterians and cnidarians further suggesting that bilateral symmetry is plesiomorphic with respect to the Bilateria.
Graded morphological complexity
It has been recognized previously that in terms of cell types and tissue types along the primary body axis, sponges are much simpler than bilaterians (i.e., sponges lack true tissues and organs) and that cnidarians occupy an intermediate level of axial complexity [29]. If the bilateral symmetry exhibited in anthozoans is considered the ancestral cnidarian condition, a pattern emerges of increasing morphological complexity along the primary body axis at each phyletic divergence point leading to the last common ancestor of bilaterians (Figure 2). Because of its uncertain phylogenetic position, Trichoplax is not considered in this model. Still, Placozoa is either the earliest diverging metazoan lineage (see [30]), in which case its asymmetry appropriately fits the model, or it is secondarily reduced (see Table 1 and references therein), in which case its asymmetry would not be appropriate to consider in this context. Sponges lack the complex organs and organ systems that are found in other Metazoa and are instead comprised of a series of asymmetric water current channels that define their (often imperfect) radial symmetry. The biradial nature of ctenophores is defined by the pharynx, the gastrovascular canals, and two sets of diametrically opposed structures: the anal pores and tentacles (Figure 2B). These structures break up the underlying multiradiate symmetry of these animals. Most bilaterally symmetrical anthozoans have underlying biradiate symmetry, defined by the pharynx and mesenteries; this symmetry is disrupted by a single structure, the siphonoglyph (and sometimes, the arrangement of the retractor muscles that attach to the mesenteries), which defines their bilateral symmetry (Figure 2C). In contrast to anthozoans, bilaterians have many structures that define their bilateriality. For example, bilaterial symmetry is defined in the earthworm by the dorsal and ventral blood vessels, the ventral nerve chord, and the intestine, to name a few (Figure 2D). Given that a limited set of developmental genes are responsible for patterning the body axis of most animals, a better understanding of the evolutionary history of these gene families will contribute greatly to our understanding of this tendency towards axial complexity in early metazoan history.
Developmental genes in early-diverging phyla
Studies from early-branching metazoans hold much promise towards furthering our understanding of the genetic and developmental basis of morphological change. The first step in this process involves establishing reliable phylogenetic classifications of gene families that highlight orthologous relationships, as well as identifying novel genes within each lineage. The next step entails identifying functional parallels (if they exist) between developmental genes in these early-branching taxa and their corresponding orthologs in Bilateria.
Hox and homeobox evolution near the base of the metazoa
Studying the homeobox superclass of transcription factors has yielded significant insight towards the goal of understanding basic developmental processes. Homeobox genes play critical roles in development in plants, fungi, and animals. A recent report showed that the common ancestor of bilaterians possessed at least 88 homeobox genes [31]. A subsequent study showed that at least 56 of these homeobox genes were likely present in the cnidarian-bilaterian ancestor [26]. This analysis demonstrates that many important genetic components in the bilaterian developmental toolkit were present in the last common ancestor of cnidarians and bilaterians.
The representation of Nematostella homeoboxes within the Hox families was conspicuously deficient when compared to the normal distribution of Nematostella to bilaterian genes within multi-family clades [26]. The Hox genes are essential for patterning the primary body axis of bilaterians, and mutations in these genes have homeotic effects on the developing embryo [32]. Evidence from the sequenced genome of the demosponge Amphimedon suggests that Hox genes emerged after the metazoan crown ancestor [33]. Furthermore, it seems unlikely that Ctenophores have Hox genes, given that there has been no report of the presence of Hox genes despite two EST sequencing projects [34, 35] and several degenerate PCR-based fishing projects [36–38].
Several previous studies have shown that Hox homologues existed in a wide range of cnidarians (reviewed in [39, 40]). However, the homeobox survey in Nematostella was the first report of a full repertoire of Hox genes from a cnidarian [26]. Subsequent studies have since demonstrated that Nematostella has at least seven Hox genes belonging to three distinct families [20, 41, 42]. In addition, genes from two of these families are organized in a cluster reminiscent of (but distinct in arrangement to) the Hox cluster of bilaterians [20, 42]. Furthermore, two additional genes from Nematostella were shown to be related to the three ParaHox genes of bilaterians [20, 41, 42]. These genes are also found in an independent cluster in many bilaterians, and this ParaHox cluster is believed to be a sister cluster to the Hox complex [43] (but see [42] for an alternate theory on the origin of the ParaHox cluster).
If we assume modest and uniform gene loss in the human, Drosophila, and Nematostella homeobox superfamilies, then the Hox subclass, more than any other subclass of homeobox genes besides Dux (which encode "double homeodomain" proteins), has underwent extensive independent radiation in the bilaterian lineage compared to the cnidarian lineage [26]. A conservative estimate is that the last common ancestor of cnidarians and bilaterians had three Hox and two ParaHox genes that likely radiated into seven Hox and three ParaHox genes in the crown bilaterian ancestor. The Hox gene radiation coincided with a significant increase in axial specificity (compare Figures 2C and 2D). This, combined with the fact that Hox genes likely arose after the divergence of Ctenophora from the rest of Metazoa, is consistent with the hypothesis that complexity and the number of Hox genes in a genome are positively correlated [44–48]. It has previously been observed that within the Bilateria, the level of complexity of a species does not necessarily correlate with the number of Hox genes in its genome [49]. Nevertheless, the pattern seen in these early branching lineages may offer insight into the early evolution of bilateral symmetry and its molecular underpinnings.
Hox, Wnt, and the evolution of the primary body axis
Does the Hox system define metazoans?
An influential review by Slack and coworkers hypothesized that Hox genes and their coordinated spatial expression along the primary body axis are metazoan synapomorphies [50]. Early reports of cnidarian Hox genes set off a debate as to whether or not this zootype hypothesis was valid, in light of these new findings [44, 51–53]. With the recent release of genomic data from Nematostella [54] and other species from early-diverging phyla, there is substantially more information from which to reevaluate the zootype hypothesis.
The evidence suggesting that the last common cnidarian-bilaterian ancestor had a Hox system is consistent with the zootype hypothesis. However, the apparent absence of Hox genes from Ctenophora and Porifera suggests that a Hox system was not in place in the earliest animals. In light of this evidence, the zootype hypothesis must be rejected sensu stricto. It is possible that the specification of regional identity along the primary body axis by a set of genes is a synapomorphy of the animal kingdom, and that the Hox genes were not the original set of genes responsible for this specification. If this is the case, then there must have been other genes responsible for patterning the primary body axis of the last common metazoan ancestor. It may be that this ancestral patterning system is still operational in sponges and ctenophores and that traces of the system in cnidarians and bilaterians might perhaps be detected.
Wnt pathway components in the early-diverging phyla
It has been postulated that the Wnt pathway may be the ancestral metazoan axial patterning system [55]. Several lines of evidence support this hypothesis. Firstly, components of the Wnt pathway have been found in two species of demosponge [9, 56] and, a recent study showed the Wnt expression pattern in the demosponge Amphimedon queenslandica to be consistent with a role in patterning the primary body axis during development[57]. Furthermore, Wnt signaling is involved in axial patterning in bilaterians. In vertebrates, hemichordates, and echinoderms, Wnt signaling is essential for posterior patterning (reviewed in [58]). In protostomes, there is evidence of expression in the posterior terminus of several insects and the posterior hindgut of polychaete annelids [59]. Similarly, the Dickkopf (Dkk) family of Wnt antagonists are required for formation of anterior structures in vertebrates (reviewed in [60]).
Lastly, phylogenetic and expression analyses from several cnidarians show that Wnt signaling plays a significant role in patterning the cnidarian primary body axis [27, 55, 61–66]. Genomic data show that there are at least 13 Wnt orthologs in Nematostella representing 11 of the 12 bilaterian Wnt subfamilies indicating that the Wnt family had extensively radiated prior to the last common cnidarian-bilaterian ancestor [27, 55]. Strikingly, in situ hybridization experiments reveal overlapping expression domains of these 13 Wnt genes at the oral end of the developing planula, encompassing roughly 75% of the animal in both germ layers. Furthermore, ectodermal expression of Dkk1/2/4, a gene whose vertebrate orthologs are major antagonists of Wnt ligands, is seen in the aboral region of the Nematostella planula, bringing the total Wnt-related signaling coverage of the developing planula close to 100% [27, 55].
Co-option of Hox genes
If a more primitive mechanism responsible for patterning the primary body axis of metazoans than the Hox system exists, then it is likely that the Hox genes were co-opted into this pathway sometime between their origin and the last common ancestor of cnidarians and bilaterians. Recent evidence from both protostomes and deuterostomes suggests that Wnt signaling partitions Hox and ParaHox domains to specify unique cell fates during development [67–71]. Additionally, Wnt genes are known to induce the expression of certain Hox genes under certain conditions in vertebrates [72]. Finally, evidence suggests that Wnt signaling works together with the ParaHox gene Cdx to regulate posterior Hox gene patterning in the mouse embryo (reviewed in [73]). These mounting data suggest that Wnt and Hox genes work together during axial specification.
It is possible, then, that the coordination of these two systems seen in modern-day bilaterians are traces of an ancient co-option event that occurred early in metazoan evolution. If the Hox genes were truly co-opted into the Wnt pathway, it is difficult to say whether the they were co-opted prior to the last common ancestor of cnidarians and bilaterians or subsequently, in the lineage leading to the last common ancestor of bilaterians. The expression patterns of Hox genes in the anthozoan Nematostella [42] and the hydrozoan Eleutheria [41] show that, while the cnidarian Hox genes are expressed in restricted regions along the primary body axis, the contiguous striped pattern that is seen in the bilaterian Hox genes is not observed. This could be an indication that the co-option process had began prior to the last common ancestor of bilaterians and cnidarians, but became more pronounced later, in the bilaterian lineage.
Speculation on the mechanisms underlying this co-option event might be premature. However, one possibility could be that a single Hox or ParaHox gene, which was high in the Hox/ParaHox cascade, was co-opted into the Wnt network. The Cdx genes (ParaHox) have been shown to be direct targets of Wnt signaling [74, 75] and thought to be direct regulators of Hox genes in vertebrates (reviewed in [73, 76]). At some point (perhaps prior to the cnidarian-bilaterian ancestor), the ortholog to Cdx could have come under the regulatory control of one or more Wnt genes. As the Hox/ParaHox family expanded through duplication, existing auto-regulatory mechanisms may have been transformed into an axial specifying network controlled initially by the Cdx ParaHox gene, which was under control of a smaller number of Wnt genes. This is only one of many other possible scenarios involving Wnt and/or other axial patterning networks. Nonetheless, these scenarios are testable and, on the surface, seem more likely than a scenario in which the Hox genes produced an independent primary body axis patterning network de novo. The initial wave of data from these early branching lineages is not yet at the stage where we can draw more definitive evolutionary scenarios, but they do provide a jumping-off point from which to generate testable hypotheses.
A better understanding of these evolutionary processes will come from more-strongly establishing the relationship between Hox, ParaHox, and Wnt genes in cnidarians, while refining those same relationships in bilaterians. A first step with respect to the Cnidaria would involve establishing the spatial and temporal relationships of the expression of these genes relative to each other. The next step would be to determine the effect of Wnt expression when Hox expression is perturbed and vice versa. If the co-option hypothesis holds, these data might help to reveal the progression of this co-option event relative to the cnidarian-bilaterian divergence as well a better understanding of the molecular mechanisms underlying bilaterian complexity and diversity.
Conclusion
It is becoming clear that combinatorial regulation plays a large part in cell fate specification [68]. A variety of transcription factors and signaling pathways active in a particular cell play a critical role in determining the fate of that cell. During embryogenesis, this combinatorial system is used to impart complexity into the developing embryo. It seems reasonable to postulate that an expansion of gene families involved in cell determination (akin to what is seen in the Wnt and Hox genes during the major transitions in metazoan history) would lead to an increase in cell types and tissue types available to embryogenesis; this would, in turn, lead to more complexity and variety in body plans. Characterizing the dynamics of gene family radiation, gene loss, and co-option is key to understanding these important metazoan transitions. By relating these data with morphological and developmental data in a phylogenetic context, we can begin to understand how changes in ontogeny have led to the burst of morphological diversity that occurred in the early Cambrian.
Reviewer's comments
Reviewer 1: Pierre Pontarotti, Université d'Aix Marseille, Marseille, France
Review of the article Hox, Wnt and the evolution of the primary body axis: insights from the early divergent phyla.
This article shed some light on the understanding and the evolution of the bilaterian animals using knowledge from non bilaterian phyla.
Two main important insights come out from this exegesis of the current literature: 1) the revaluation of the zootype hypothesis and 2) the putative role of the Wnt genes in the partitioning of the body axis in urbilateria.
I think that this review article will help experimental design for the "evo/devo" scientific community.
Specific comments:
"Early divergent phyla and the primary body axis" paragraph:
1) The single cell ancestor hypothesis
What really are the evidence that animals descent from several single cell ancestors? Multicellularity occurred several times independently in the history of life, see for example Fungi. I do not understand why the hypothesis of convergent multicellularity should not be considered in the case of animal.
Authors' response: The relationship of animals to single-celled eukaryotes is tangential to our thesis, so we have removed the reference to animal origins in the paper. Please also see the response to Comment 2.
2) The dating of the occurrence of animal multicellularity
I would be careful about the dating; the fact that we cannot witness older multicellular fossil than the one found in the Ediacaran fauna does not mean that they did not exist.
Authors' response: We reworded a couple of sentences to make it clear that these dates are only estimates based on published studies. The following sentences were changed:
"All animals descended from a single-celled ancestor (most likely a choanofagellate) over 600 million years ago[4]. Bilaterian animals radiated during the Cambrian explosion 544–525 million years ago."
The text now reads:
"The origin of animals dates back to over 600 million years ago and perhaps much earlier [4]. Fossil evidence and molecular evidence suggest bilaterian animals radiated during the Cambrian period some 500–550 million years ago [5]."
"Graded morphological complexity" paragraph:
The authors wrote: "sponges are much simpler than bilaterians and that cnidarians occupy an intermediate level of axial complexity". What is the objective parameter of complexity. The authors should explain it (even if this is evident for most of our colleague).
Authors' response: The following sentence was changed:
"It has been recognized previously that in terms of complexity along the primary body axis, sponges are much simpler than bilaterians and that cnidarians occupy an intermediate level of axial complexity [29]."
The text now reads:
"It has been recognized previously that in terms of cell types and tissue types along the primary body axis, sponges are much simpler than bilaterians (i.e., sponges lack true tissues and organs) and that cnidarians occupy an intermediate level of axial complexity [29]."
Concerning the phylogeny of Animal
Do the authors think that Bilaterian and Cnidarians form a monophylogentic group, do they think the hox where co-opted in the most common ancestor of these two phyla? It should be helpful if the authors discuss this possibility.
Authors' response: Figure 1shows the prevailing view that Cnidaria and Bilateria are sister taxa and form a monophyletic group. Most of the ribosomal RNA phylogenies support this relationship as shown in Table 1.
We've inserted the following into the text to address the second part of this comment:
"If the Hox genes were truly co-opted into the Wnt pathway, it is difficult to say whether the they were co-opted prior to the last common ancestor of cnidarians and bilaterians or subsequently, in the lineage leading to the last common ancestor of bilaterians. The expression patterns of Hox genes in the anthozoan Nematostella[42]and the hydrozoan Eleutheria[41]show that, while the cnidarian Hox genes are expressed in restricted regions along the primary body axis, the contiguous striped pattern that is seen in the bilaterian Hox genes is not observed. This could be an indication that the co-option process had began prior to the last common ancestor of bilaterians and cnidarians, but became more pronounced later, in the bilaterian lineage."
Reviewer 2: Gáspár Jékely, Max Planck Institute for Developmental Biology, Tübingen, Germany
Reviewed by: Gáspár Jékely, Max Planck Institute for Developmental Biology, Spemannstrasse 35, Tübingen, 72076 Germany, Tel: +49 7071 601 1310, gaspar.jekely@tuebingen.mpg.de
This paper gives a good overview of the early evolution of axial symmetry and the patterning along the AP axis in Metazoa. The paper is well written and I only have a few comments and suggestions for potential improvement.
The authors propose that the Hox system was probably co-opted into the more ancestral Wnt system for patterning along the AP axis. This is an interesting idea and it would be worth spelling it out in a bit more detail. How could it have happened? Did Hox genes intercalate between Wnt signalling and the target genes of Wnt etc.? Why did it allow more precise axial patterning? An extra figure showing the steps of the evolution of the Wnt-Hox axial patterning system in Metazoa would also help (e.g. schematic embryos with the expression domains).
Authors' response: We've inserted the following paragraph into the text to address these questions:"Speculation on the mechanisms underlying this co-option event might be premature. However, one possibility could be that a single Hox or ParaHox gene, which was high in the Hox/ParaHox cascade, was co-opted into the Wnt network. The Cdx genes (ParaHox) have been shown to be direct targets of Wnt signaling[74, 75]and thought to be direct regulators of Hox genes in vertebrates (reviewed in[73, 76]). At some point (perhaps prior to the cnidarian-bilaterian ancestor), the ortholog to Cdx could have come under the regulatory control of one or more Wnt genes. As the Hox/ParaHox family expanded through duplication, existing auto-regulatory mechanisms may have been transformed into an axial specifying network controlled initially by the Cdx ParaHox gene, which was under control of a smaller number of Wnt genes. This is only one of many other possible scenarios involving Wnt and/or other axial patterning networks. Nonetheless, these scenarios are testable and, on the surface, seem more likely than a scenario in which the Hox genes produced an independent primary body axis patterning network de novo. The initial wave of data from these early branching lineages is not yet at the stage where we can draw more definitive evolutionary scenarios, but they do provide a jumping-off point from which to generate testable hypotheses."
Wnt expression was also analysed recently in the sponge Amphimedon queenslandica (PLoS ONE 2007 vol. 2 pp. e1031) and was found to be expressed at the posterior pole of larvae. This is in agreement with the idea that Wnt signalling plays an ancestral role in patterning the AP axis. These findings should also be discussed.
Authors' response: We inserted the following into the text:
"...a recent study showed the Wnt expression pattern in the demosponge Amphimedon queenslandica to be consistent with a role in patterning the primary body axis during development[57]."
Reviewer 3: L Aravind, Computational Biology Branch, NCBI, NLM, NIH, Bethesda, USA
This reviewer provided no comments for publication.
References
Nielsen C: Defining phyla: morphological and molecular clues to metazoan evolution. Evol Dev 2003, 5: 386-393. 10.1046/j.1525-142X.2003.03046.x
Valentine JW: On the Origin of Phyla. Chicago: The University of Chicago Press; 2004.
Willmer P: Invertebrate relationships : patterns in animal evolution. Cambridge; New York: Cambridge University Press; 1990.
King N: The Unicellular Ancestry of Animal Development. Developmental Cell 2004, 7: 313-325. 10.1016/j.devcel.2004.08.010
Valentine JW: Late Precambrain Bilaterians: Grades and Clades. Proceedings of the National Academy of Sciences 1994, 91: 6751-6757. 10.1073/pnas.91.15.6751
Beklemishev VN: Principles of comparative anatomy of invertebrates. Edinburgh,: Oliver & Boyd; 1969.
Manuel M, Borchiellini C, Alivon E, Le Parco Y, Vacelet J, Boury-Esnault N: Phylogeny and evolution of calcareous sponges: monophyly of calcinea and calcaronea, high level of morphological homoplasy, and the primitive nature of axial symmetry. Syst Biol 2003, 52: 311-333. 10.1080/10635150390196966
Leys SP, Degnan BM: Embryogenesis and metamorphosis in a haplosclerid demosponge: gastrulation and transdifferentiation of larval ciliated cells to choanocytes. Invertebr Biol 2002, 121: 171-189.
Nichols SA, Dirks W, Pearse JS, King N: Early evolution of animal cell signaling and adhesion genes. PNAS %R 101073/pnas0604065103 2006, 103: 12451-12456. 10.1073/pnas.0604065103
Muller WE, Wiens M, Adell T, Gamulin V, Schroder HC, Muller IM: Bauplan of urmetazoa: basis for genetic complexity of metazoa. Int Rev Cytol 2004, 235: 53-92.
Larroux C, Fahey B, Liubicich D, Hinman VF, Gauthier M, Gongora M, Green K, Worheide G, Leys SP, Degnan BM: Developmental expression of transcription factor genes in a demosponge: insights into the origin of metazoan multicellularity. Evolution & Development 2006, 8: 150-173. 10.1111/j.1525-142X.2006.00086.x
Cuvier G, M'Murtrie H: The animal kingdom, arranged in conformity with its organization. New York,: G. & C. & H. Carvill; 1832.
Cavalier-Smith T: A 6 kingdom classification and a unified phylogeny. In Endocytobiology II : intracellular space as oligogenetic ecosystem : proceedings/Second International Coloquium on Endocytobiology. Edited by: Schenk HEA, Schwemmler W. Berlin; New York: De Gruyter; 1983:1027-1034.
Cavalier-Smith T: A revised six-kingdom system of life. Biological reviews of the Cambridge Philosophical Society 1998, 73: 03-266.
Martindale MQ, Finnerty JR, Henry JQ: The Radiata and the evolutionary origins of the bilaterian body plan. Molecular Phylogenetics and Evolution 2002, 24: 358-365. 10.1016/S1055-7903(02)00208-7
Martindale MQ: The evolution of metazoan axial properties. Nat Rev Genet 2005, 6: 917-927. 10.1038/nrn1818
Finnerty JR: The origins of axial patterning in the metazoa: how old is bilateral symmetry? Int J Dev Biol 2003, 47: 523-529.
Stephenson TA: British Sea Anemones. London: The Ray Society; 1928.
Hyman LH: The invertebrates. 1st edition. New York,: McGraw-Hill; 1940.
Chourrout D, Delsuc F, Chourrout P, Edvardsen RB, Rentzsch F, Renfer E, Jensen MF, Zhu B, de Jong P, Steele RE, Technau U: Minimal ProtoHox cluster inferred from bilaterian and cnidarian Hox complements. Nature 2006, 442: 684-687. 10.1038/nature04863
Bridge D, Cunningham CW, Schierwater B, DeSalle R, Buss LW: Class-Level Relationships in the Phylum Cnidaria: Evidence from Mitochondrial Genome Structure. PNAS 1992, 89: 8750-8753. 10.1073/pnas.89.18.8750
Hadzi H: Turbellarijska teorija knidarijev (Turbellarien-Theorie der Knidarien). Ljubljana, Slovenia; 1944.
Hand C: On the Origin and Phylogeny of the Coelenterates. Systematic Zoology 1959, 8: 191-202. 10.2307/2411883
Matus DQ, Pang K, Marlow H, Dunn CW, Thomsen GH, Martindale MQ: Molecular evidence for deep evolutionary roots of bilaterality in animal development. Proc Natl Acad Sci USA 2006, 103: 11195-11200. 10.1073/pnas.0601257103
Finnerty JR, Pang K, Burton P, Paulson D, Martindale MQ: Origins of bilateral symmetry: Hox and dpp expression in a sea anemone. Science 2004, 304: 1335-1337. 10.1126/science.1091946
Ryan JF, Burton PM, Mazza ME, Kwong GK, Mullikin JC, Finnerty JR: The cnidarian-bilaterian ancestor possessed at least 56 homeoboxes: evidence from the starlet sea anemone, Nematostella vectensis. Genome Biology 2006., 7:
Kusserow A, Pang K, Sturm C, Hrouda M, Lentfer J, Schmidt HA, Technau U, von Haeseler A, Hobmayer B, Martindale MQ, Holstein TW: Unexpected complexity of the Wnt gene family in a sea anemone. Nature 2005, 433: 156-160. 10.1038/nature03158
Rentzsch F, Anton R, Saina M, Hammerschmidt M, Holstein TW, Technau U: Asymmetric expression of the BMP antagonists chordin and gremlin in the sea anemone Nematostella vectensis: Implications for the evolution of axial patterning. Dev Biol 2006.
Finnerty JR: Cnidarians Reveal Intermediate Stages in the Evolution of Hox Clusters and Axial Complexity1. American Zoologist 2001, 41: 608-620. 10.1668/0003-1569(2001)041[0608:CRISIT]2.0.CO;2
Dellaporta SL, Xu A, Sagasser S, Jakob W, Moreno MA, Buss LW, Schierwater B: Mitochondrial genome of Trichoplax adhaerens supports placozoa as the basal lower metazoan phylum. Proc Natl Acad Sci USA 2006, 103: 8751-8756. 10.1073/pnas.0602076103
Nam J, Nei M: Evolutionary Change of the Numbers of Homeobox Genes in Bilateral Animals. Mol Biol Evol 2005.
Lewis EB: A gene complex controlling segmentation in Drosophila. Nature 1978, 276: 565-570. 10.1038/276565a0
Larroux CC, Fahey BB, Degnan SMSM, Adamski MM, Rokhsar DSDS, Degnan BMBM: The NK homeobox gene cluster predates the origin of Hox genes. Current biology 2007, 17: 706-710. 10.1016/j.cub.2007.03.008
Jager M, Queinnec E, Houliston E, Manuel M: Expansion of the SOX gene family predated the emergence of the Bilateria. Molecular Phylogenetics and Evolution 2006, 39: 477. 10.1016/j.ympev.2005.12.005
Blaxter ML, Aboobaker AA, Pang K, Martindale MQ: A survey of genes expressed in the ctenophore Mnemiopsis leidyi (GenBank EST submission). Book A survey of genes expressed in the ctenophore Mnemiopsis leidyi (GenBank EST submission) (Editor ed.^eds.). City 2003.
Finnerty JR: Homeobox genes in the Ctenophora: identification of paired-type and Hox homologues in the atentaculate ctenophore, Beroe ovata. Molecular marine biology and biotechnology 1996, 5: 249-258.
Martinelli C, Spring J: T-box and homeobox genes from the ctenophore Pleurobrachia pileus: comparison of Brachyury, Tbx2/3 and Tlx in basal metazoans and bilaterians. FEBS Lett 2005, 579: 5024-5028. 10.1016/j.febslet.2005.08.008
Bebenek IG, Gates RD, Morris J, Hartenstein V, Jacobs DK: sine oculis in basal Metazoa. Development Genes and Evolution 2004, 214: 342-351. 10.1007/s00427-004-0407-3
Ferrier DE, Minguillon C: Evolution of the Hox/ParaHox gene clusters. Int J Dev Biol 2003, 47: 605-611.
Galliot B: Conserved and divergent genes in apex and axis development of cnidarians. Curr Opin Genet Dev 2000, 10: 629-637. 10.1016/S0959-437X(00)00141-6
Kamm K, Schierwater B, Jakob W, Dellaporta SL, Miller DJ: Axial Patterning and Diversification in the Cnidaria Predate the Hox System. Curr Biol 2006.
Ryan JF, Mazza ME, Pang K, Matus DQ, Baxevanis AD, Martindale MQ, Finnerty JR: Pre-bilaterian Origins of the Hox Cluster and the Hox Code: Evidence from the Sea Anemone, Nematostella vectensis. PLoS One 2007, 2: e153. 10.1371/journal.pone.0000153
Brooke NM, Garcia-Fernandez J, Holland PW: The ParaHox gene cluster is an evolutionary sister of the Hox gene cluster. Nature 1998, 392: 920-922. 10.1038/31933
Finnerty JR: Homeoboxes in sea anemones and other nonbilaterian animals: implications for the evolution of the hox cluster and the zootype [In Process Citation]. Curr Top Dev Biol 1998, 40: 211-254.
Ruddle FH, Bartels JL, Bentley KL, Kappen C, Murtha MT, Pendleton JW: Evolution of Hox genes. Annu Rev Genet 1994, 28: 423-442. 10.1146/annurev.ge.28.120194.002231
Pendleton JW, Nagai BK, Murtha MT, Ruddle FH: Expansion of the Hox gene family and the evolution of chordates. Proc Natl Acad Sci USA 1993, 90: 6300-6304. 10.1073/pnas.90.13.6300
Kappen C, Schughart K, Ruddle FH: 2 Steps in the Evolution of Antennapedia-Class Vertebrate Homeobox Genes. Proceedings of the National Academy of Sciences of the United States of America 1989, 86: 5459-5463. 10.1073/pnas.86.14.5459
Akam M, Dawson I, Tear G: Homeotic Genes and the Control of Segment Diversity. Development 1988, 104: 123-133.
Finnerty JR, Martindale MQ: The evolution of the Hox cluster: insights from outgroups. Current Opinion in Genetics & Development 1998, 8: 681-687. 10.1016/S0959-437X(98)80037-3
Slack JM, Holland PW, Graham CF: The zootype and the phylotypic stage. Nature 1993, 361: 490-492. 10.1038/361490a0
Martinez DE, Bridge D, Masuda-Nakagawa LM, Cartwright P: Cnidarian homeoboxes and the zootype. Nature 1998, 393: 748-749. 10.1038/31616
Schierwater B, Desalle R: Current problems with the zootype and the early evolution of Hox genes. J Exp Zool 2001, 291: 169-174. 10.1002/jez.1066
Schierwater B, Kuhn K: Homology of Hox genes and the zootype concept in early metazoan evolution. Mol Phylogenet Evol 1998, 9: 375-381. 10.1006/mpev.1998.0489
Putnam NH, Srivastava M, Hellsten U, Dirks B, Chapman J, Salamov A, Terry A, Shapiro H, Lindquist E, Kapitonov VV, et al.: Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317: 86-94. 10.1126/science.1139158
Lee PN, Pang K, Matus DQ, Martindale MQ: A WNT of things to come: Evolution of Wnt signaling and polarity in cnidarians. Seminars in Cell & Developmental Biology 2006, 17: 157-167. 10.1016/j.semcdb.2006.05.002
Adell T, Nefkens I, Muller WEG: Polarity factor 'Frizzled' in the demosponge Suberites domuncula: identification, expression and localization of the receptor in the epithelium/pinacoderm. FEBS Letters 2003, 554: 363-368. 10.1016/S0014-5793(03)01190-6
Adamska M, Degnan SM, Green KM, Adamski M, Craigie A, Larroux C, Degnan BM: Wnt and TGF-beta Expression in the Sponge Amphimedon queenslandica and the Origin of Metazoan Embryonic Patterning. PLoS ONE 2007, 2: e1031. 10.1371/journal.pone.0001031
Holland LZ: Heads or Tails? Amphioxus and the Evolution of Anterior-Posterior Patterning in Deuterostomes. Developmental Biology 2002, 241: 209-228. 10.1006/dbio.2001.0503
Seaver EC, Kaneshige LM: Expression of 'segmentation' genes during larval and juvenile development in the polychaetes Capitella sp. I and H. elegans. Developmental Biology 2006, 289: 179-194. 10.1016/j.ydbio.2005.10.025
Niehrs C: Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 25: 7469-7481. 10.1038/sj.onc.1210054
Augustin R, Franke A, Khalturin K, Kiko R, Siebert S, Hemmrich G, Bosch TCG: Dickkopf related genes are components of the positional value gradient in Hydra. Developmental Biology 2006, 296: 62-70. 10.1016/j.ydbio.2006.04.003
Broun M, Gee L, Reinhardt B, Bode HR: Formation of the head organizer in hydra involves the canonical Wnt pathway. Development 2005, 132: 2907-2916. 10.1242/dev.01848
Guder C, Pinho S, Nacak TG, Schmidt HA, Hobmayer B, Niehrs C, Holstein TW: An ancient Wnt-Dickkopf antagonism in Hydra. Development 2006, 133: 901-911. 10.1242/dev.02265
Hobmayer B, Rentzsch F, Kuhn K, Happel CM, von Laue CC, Snyder P, Rothbacher U, Holstein TW: WNT signalling molecules act in axis formation in the diploblastic metazoan Hydra. Nature 2000, 407: 186-189. 10.1038/35025063
Wikramanayake AH, Hong M, Lee PN, Pang K, Byrum CA, Bince JM, Xu R, Martindale MQ: An ancient role for nuclear [beta]-catenin in the evolution of axial polarity and germ layer segregation. Nature 2003, 426: 446-450. 10.1038/nature02113
Plickert G, Jacoby V, Frank U, Muller WA, Mokady O: Wnt signaling in hydroid development: Formation of the primary body axis in embryogenesis and its subsequent patterning. Developmental Biology 2006, 298: 368-378. 10.1016/j.ydbio.2006.06.043
Arata Y, Kouike H, Zhang Y, Herman MA, Okano H, Sawa H: Wnt signaling and a Hox protein cooperatively regulate psa-3/Meis to determine daughter cell fate after asymmetric cell division in C. elegans. Dev Cell 2006, 11: 105-115. 10.1016/j.devcel.2006.04.020
Bondos S: Variations on a theme: Hox and Wnt combinatorial regulation during animal development. Sci STKE 2006, 2006: pe38. 10.1126/stke.3552006pe38
Merabet S, Hombria JC, Hu N, Pradel J, Graba Y: Hox-controlled reorganisation of intrasegmental patterning cues underlies Drosophila posterior spiracle organogenesis. Development 2005, 132: 3093-3102. 10.1242/dev.01889
Bondos SE, Tan X-X, Matthews KS: Physical and Genetic Interactions Link Hox Function with Diverse Transcription Factors and Cell Signaling Proteins. Molecular Cellular Proteomics 2006, 5: 824-834. 10.1074/mcp.M500256-MCP200
Bilder D, Graba Y, Scott MP: Wnt and TGFbeta signals subdivide the AbdA Hox domain during Drosophila mesoderm patterning. Development 1998, 125: 1781-1790.
Altmann CR, Brivanlou AH, Lawrence DEa KWJ: Neural patterning in the vertebrate embryo. In International Review of Cytology. Volume 203. Academic Press; 2001:447-482.
Lohnes D: The Cdx1 homeodomain protein: an integrator of posterior signaling in the mouse. BioEssays 2003, 25: 971-980. 10.1002/bies.10340
Pilon N, Oh K, Sylvestre JR, Bouchard N, Savory J, Lohnes D: Cdx4 is a direct target of the canonical Wnt pathway. Dev Biol 2006, 289: 55-63. 10.1016/j.ydbio.2005.10.005
Pilon N, Oh K, Sylvestre JR, Savory JG, Lohnes D: Wnt signaling is a key mediator of Cdx1 expression in vivo. Development 2007, 134: 2315-2323. 10.1242/dev.001206
Deschamps J, van Nes J: Developmental regulation of the Hox genes during axial morphogenesis in the mouse. Development 2005, 132: 2931-2942. 10.1242/dev.01897
Collins AG, Valentine JW: Defining phyla: evolutionary pathways to metazoan body plans. Evol Dev 2001, 3: 432-442. 10.1046/j.1525-142X.2001.01048.x
Buchsbaum R: Animals without backbones. Chicago, Illinois: University of Chicago Press; 1938.
Wainright PO, Hinkle G, Sogin ML, Stickel SK: Monophyletic origins of the metazoa: an evolutionary link with fungi. Science 1993, 260: 340-342. 10.1126/science.8469985
Katayama T, Wada H, Furuya H, Satoh N, Yamamoto M: Phylogenetic position of the dicyemid mesozoa inferred from 18S rDNA sequences. Biological Bulletin 1995, 189: 81-90. 10.2307/1542458
Hanelt B, Van Schyndel D, Adema CM, Lewis LA, Loker ES: The phylogenetic position of Rhopalura ophiocomae (Orthonectida) based on 18S ribosomal DNA sequence analysis. Molecular Biology and Evolution 1996, 13: 1187-1191.
Van De Peer Y, De Wachter R: Evolutionary relationships among the eukaryotic crown taxa taking into account site-to-site rate variation in 18S rRNA. Journal of Molecular Evolution 1997, 45: 619-630. 10.1007/PL00006266
Abouheif E, Zardoya R, Meyer A: Limitations of metazoan 18s rRNA sequence data: Implications for reconstructing a phylogeny of the animal kingdom and inferring the reality of the cambrian explosion. Journal of Molecular Evolution 1998, 47: 394-405. 10.1007/PL00006397
Collins AG: Evaluating multiple alternative hypotheses for the origin of Bilateria: an analysis of 18S rRNA molecular evidence. Proc Natl Acad Sci USA 1998, 95: 15458-15463. 10.1073/pnas.95.26.15458
Halanych KM: Considerations for Reconstructing Metazoan History: Signal, Resolution, and Hypothesis Testing. Integrative and Comparative Biology 1998, 38: 929-941. 10.1093/icb/38.6.929
Lipscomb DL, Farris JS, Ka?llersjo M, Tehler A: Support, ribosomal sequences and the phylogeny of the eukaryotes. Cladistics 1998, 14: 303-338. 10.1111/j.1096-0031.1998.tb00341.x
Littlewood DTJ, Telford MJ, Clough KA, Rohde K: Gnathostomulida – An Enigmatic Metazoan Phylum from both Morphological and Molecular Perspectives. Molecular Phylogenetics and Evolution 1998, 9: 72-79. 10.1006/mpev.1997.0448
Winnepenninckx BMH, Van Peer YDE, Backeljau T: Metazoan Relationships on the Basis of 18S rRNA Sequences: A Few Years Later. Integrative and Comparative Biology 1998, 38: 888-906. 10.1093/icb/38.6.888
Zrzavy J, Mihulka S, Kepka P, Bezde?k A, Tietz D: Phylogeny of the Metazoa Based on Morphological and 18S Ribosomal DNA Evidence. Cladistics 1998, 14: 249-285.
Kim J, Kim W, Cunningham CW: A new perspective on lower metazoan relationships from 18S rDNA sequences [2]. Molecular Biology and Evolution 1999, 16: 423-427.
Giribet G, Wheeler WC: The Position of Arthropods in the Animal Kingdom: Ecdysozoa, Islands, Trees, and the "Parsimony Ratchet". Molecular Phylogenetics and Evolution 1999, 13: 619-623. 10.1006/mpev.1999.0679
Siddall ME, Whiting MF: Long-branch abstractions. Cladistics 1999, 15: 9-24. 10.1111/j.1096-0031.1999.tb00391.x
Peterson KJ, Eernisse DJ: Animal phylogeny and the ancestry of bilaterians: Inferences from morphology and 18S rDNA gene sequences. Evolution and Development 2001, 3: 170-205. 10.1046/j.1525-142x.2001.003003170.x
Podar M, Haddock SH, Sogin ML, Harbison GR: A molecular phylogenetic framework for the phylum Ctenophora using 18S rRNA genes. Mol Phylogenet Evol 2001, 21: 218-230. 10.1006/mpev.2001.1036
Collins AG: Phylogeny of Medusozoa and the evolution of cnidarian life cycles. Journal of Evolutionary Biology 2002, 15: 418-432. 10.1046/j.1420-9101.2002.00403.x
Jondelius U, Ruiz-Trillo I, Bagun?a J, Riutort M: The Nemertodermatida are basal bilaterians and not members of the Platyhelminthes. Zoologica Scripta 2002, 31: 201-215. 10.1046/j.1463-6409.2002.00090.x
Martinelli C, Spring J: Distinct expression patterns of the two T-box homologues Brachyury and Tbx2/3 in the placozoan Trichoplax adhaerens. Dev Genes Evol 2003, 213: 492-499. 10.1007/s00427-003-0353-5
Zrzavy J, Hypsa V: Myxozoa, Polypodium, and the origin of the Bilateria: The phylogenetic position of "Endocnidozoa" in light of the rediscovery of Buddenbrockia [1]. Cladistics 2003, 19: 164-169. 10.1016/S0748-3007(03)00007-0
Wallberg A, Thollesson M, Farris JS, Jondelius U: The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling. Cladistics 2004, 20: 558-578. 10.1111/j.1096-0031.2004.00041.x
Acknowledgements
We are very grateful to Gaspar Jekely and Pierre Pontarotti for their insightful reviews. We thank John Finnerty, Mark Martindale, Chris Schneider, Jim Mullikin, Daniel Segrè, and Adam Reitzel for comments on various versions of this manuscript. We also thank Allen Collins, Shawn Burgess, Yingzi Yang, and the members of the Finnerty Lab at Boston University for helpful discussion of these topics. This research was funded by the Intramural Research Program of the National Human Genome Research Institute, National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Ryan, J.F., Baxevanis, A.D. Hox, Wnt, and the evolution of the primary body axis: insights from the early-divergent phyla. Biol Direct 2, 37 (2007). https://doi.org/10.1186/1745-6150-2-37
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1745-6150-2-37