
Abstract
Background
Both regulatory T cells (Tregs) and PD-1/PD-L1 pathway were critically involved in HCV viral persistence. However, the association between them was not well investigated. Herein, we aimed to investigate the distributional profiles of Tregs subsets and association between PD-1 expression on these subsets and development of HCV long-term persistence.
Methods
CD45RA and CD27 were employed to separate peripheral Tregs as naïve/central memory/effector memory/effector subsets. The phenotypic characteristics and PD-1 expression of Tregs were studied by flow cytometry.
Results
In the present study, the majority of Tregs was identified as central memory phenotype in chronic hepatitis C patients compared with nearly equal contribution of naïve and central memory subsets in healthy individuals. PD-1 expression was elevated in all CD4+ T cell subset in chronic HCV infected patients, including Tregs. Of note, higher level of PD-1 expression was found on TEM- and effector-Treg than naïve- and TCM-Tregs subsets. The ratio of TEM-Tregs/naive-Tregs and TEM-Tregs/TCM-Tregs regarding to PD-1 MFI were significantly lower in CHC patients compared to controls.
Conclusions
Our study indicated that distinctive characteristics of PD-1 expression on Tregs in HCV infection suggests associated with impaired adaptive immunity as well as viral long-term persistence. The cross talk between Treg cells and PD-1 induced inhibition in chronic HCV infection deserved further exploration for HCV infection associated immune pathogenesis.
Similar content being viewed by others


Introduction
Hepatitis C virus (HCV) infection presents a major global health challenge, with an estimation of more than 180 million infected patients worldwide and three to four million newly-infected cases annually [1, 2]. Although some breakthroughs were reached recently in the treatment of hepatitis C [3–5], HCV infection is still far away from effective prevention by vaccination and has become one of the most important causes of chronic hepatitis, cirrhosis, and hepatocellular carcinoma around the world.
It is well documented [6–8]that duration and intensity of HCV-specific CD4+ T cell responses played an important role in determining the clinical outcomes of acute self-recovery or long-term viremic persistence following acute HCV infection. Vigorous virus-specific CD4+ T cell responses led to virus clearance in HCV acute-resolved individuals. The failure of effective CD4+ T cell immune responses to clean virus was associated with chronic liver disease and HCV viral persistence. Tregs were defined as a subset of CD4+T lymphocytes, highly expressing CD25 (interleukin-2 receptor α-chain) on the cell surface and transcription factor Foxp3 intracellularly and occupied 5% of peripheral CD4+ T lymphocytes [9–11]. Tregs contributed critically to suppression of HCV-specific lymphocyte proliferation, differentiation and cytokine secretion [12–14], but their complicated mechanisms involved in HCV persistence were still obscure.
Another important negative regulating mechanism was the involvement of PD-1/PD-L1 pathway. PD-1 (programmed cell death-1) belongs to the CD28 family and is expressed on activated T, B, and myeloid cells. PD-1 and its ligand PD-L1 deliver inhibitory signals that regulate the balance between effector T cell activation and immune-mediated tissue damage [15, 16]. PD-1 knockout C57BL/6 or BALB/c mice were shown to develop lupus-like glomerulonephritis and dilated cardiomyopathy[17, 18]and treatment of activated T cells with anti-PD-L1 antibody in vitro resulted in reduction of T cell proliferation and IFN-γ secretion[19]. Several studies indicated that PD-1/PD-L1 pathway was critically involved in HCV long-term persistence and was regarded as a potential novel target for restoring function of exhausted HCV-specific T cell responses [20–22].
Since inhibition of the effector T cells by Tregs and the PD-1/PD-L1 pathway were described as mechanisms responsible for balancing of HCV adaptive T cell responses, the association between these two factors should be attractive and informative for the exploration of mechanisms associated with HCV long-term persistence. It was reported that Tregs proliferation and its suppressive function were negatively regulated by PD-1/PD-L1 pathway through a potentially novel mechanism involving the prevention of IL-2 driven STAT-5 phosphorylation during chronic HCV infection [23, 24]. However, detailed information with respect to PD-1 expression on different phenotypes (naïve/memory/effector) of Tregs was not well understood.
Not only human CD8+ T cells, but also CD4+ T cells could be divided into four different subsets (naïve, central memory, effector memory and effector) according to expression of maturation marker CD45RA or CD45RO and another co-stimulating molecules (CD62L/CD27/CCR7)[25–29]. CD4+ Treg cells are a specialized subpopulation of CD4+ T cells that act to suppress activation of the immune system and thereby its phenotype and function could be studied through the similar classification principles as used by total CD4+ T cells. Additionally, some groups have already studied the phenotype and function of Treg cells based on expression of CD45RA and/or CD27 molecules[30–33]. In this study, we aimed to investigate the distributional profiles of Tregs subsets according to maturation markers CD45RA and CD27 expression. Association between PD-1 expression on these subsets and development of HCV long-term persistence was also examined. Our findings demonstrated that PD-1 expression on peripheral CD4+CD127loCD25hi FoxP3+ Tregs was elevated in HCV chronically infected patients while the extent of elevation was attenuated compared with other CD4+ subsets. Additionally, the dominant proportion of peripheral Tregs carried central memory phenotype in chronic hepatitis C patients compared with nearly equal contribution of naïve and central memory T cell in healthy individuals.
Materials and methods
Study population
A total of 19 patients with chronic HCV infection were enrolled in the study. All patients have more than 3-year history of commercial blood donations in early 1990 s. All these patients were positive for HCV antibodies for the past 5 years and previous HCV test data were collected from local CDC and hospital. HCV infection was confirmed by detectable plasma HCV RNA (≥1.48 log10 IU/ml) measured by Abbott Real Time™ HCV Amplification Kit (Abbott Molecular Inc. Des Plaines, IL, USA) and positive serum HCV antibodies measured by ARCHITECT Anti-HCV system (Abbott Molecular Inc. Des Plaines, IL, USA) according to the manufacture's instruction. HCV genotype was determined by amplifying and sequencing NS5b gene of HCV genome. CD4+/CD8+ T cell counts and liver function associated enzymes were measured based on clinical standard diagnostic criteria by the local Center for Disease Control and Prevention. None of these HCV infected patients received anti-HCV therapy. 17 non-HCV infected individuals were recruited as healthy controls. None of the participants showed evidence of neither concomitant HBV or HIV infection, or other non-viral causes of hepatic damage. The study was approved by institutional review authorities of Peking University Health Science Center and informed consent was obtained from all participants prior to enrollment.
Flow Cytometry Analysis
Peripheral blood mononuclear cells (PBMC) were isolated from heparinized blood by density gradient centrifugation using Ficoll-Hypaque (Sigma Chemical Co., St Louis, MO, USA). Expression of surface markers on CD4+ T cells were analyzed by multi-parameter flow cytometry on a FACSAria instruction (BD Biosciences, San Jose, CA) compensated with single fluorochromes and driven by FACSDiva software (BD Biosciences, San Jose, CA). The following antibodies were used in this study: anti-CD3-ECD, anti-CD4-APC-Cy7, anti-CD8-PE-Cy7, anti-CD45RA-PE-Cy5, anti-CD27-PE, anti-CD25-APC, anti-CD127-Pacific blue, anti-FoxP3-Alexa700, and anti-PD-1-FITC (BD Pharmingen, San Diego, CA, USA). Data analysis was performed by FlowJo software (Tree Star, Inc., San Carlos, CA).
Statistical analysis
Characteristics of the study population and the different immune or biochemical parameters were recorded as median and interquartile range. Comparisons between groups were analyzed using non-parametric T tests (Mann-Whitney test) or Wilcoxon match pairs test. Correlations between quantitative parameters were analyzed using Spearman rank correlation coefficient test. All statistical analysis was performed using GraphPad Prism V5.0 (GraphPad Software Inc., San Diego, CA). All P-values were two-tailed, and were considered significant when lower than 0.05.
Results
Characteristics of study subjects
Table 1 summarizes the main characteristics of 19 HCV-infected individuals. All data were described as median (IQR1-IQR3). There was no significant difference in age and gender between chronic HCV infection and healthy controls (data not shown).
The frequency of different CD4+T cell subsets defined by CD127 and CD25, CD45RA and CD27 respectively
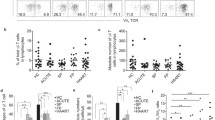
Three subsets of CD4+ T cells were defined by the expression of IL-2Rα (CD25) and IL-7Rα (CD127):CD127+CD25lo/-, CD127-CD25-, and CD127loCD25hi (Figure 1A). Given that the most definitive markers available to identify Tregs are the co-expression of CD25 and FoxP3, we examined the expression of FoxP3 in the CD127loCD25hi subsets which represented Tregs (Figure 1A). As shown in Figure 1A, more than 95% of CD127loCD25hi subsets were FoxP3 positive cells. The percentage of CD127loCD25hiFoxP3+Tregs was increased in chronic hepatitis C (CHC) patients compared with health controls (HCs) (p = 0.0338) (Figure 1B). On the contrary, decreased expression of CD127+CD25lo/- subsets was observed in CHC patients compared with HCs (p = 0.0032). No significant difference was found in CD127-CD25- subsets distribution (p = 0.0759) (Figure 1B). CD4+T cells can be divided into four different subsets depending on expression level of CD45RA and CD27: naïve cells (CD27+CD45RA+), central memory cells (CD27+CD45RA-), effector memory cells (CD27-CD45RA-), and effector cells (CD27-CD45RA+) (Figure 1A). The statistical results showed that the proportion of naïve CD4+T cells were declined in CHC patients compared with HCs (p = 0.0004), while the other three subsets (TCM, p = 0.0480; TEM, p = 0.0015; Effector T, p = 0.0016)were all dramatically increased in CHC patients(Figure 1C).

Increased frequency of Treg cells (CD4+CD127loCD25hiFoxP3+) and effector memory/effector CD4+ T cell subsets in peripheral blood of chronic HCV-infected patients. PBMCs were stained with T cell phenotyping markers (CD3, CD4, CD8, CD25, CD27, CD45RA, CD127 and FoxP3) and analyzed for T cells subsets by flow cytometric analysis. (A): Representative examples of the gating strategy that CD3+CD4+CD8- T cells were divided into three subsets -CD127-CD25-, CD127+CD25lo/- and CD127loCD25hi, based on expression of CD25 and CD127 or into four subsets- naïve, TCM, TEM and effector, based on expression of CD45RA and CD27. Additionally, more than 95% of CD127loCD25hi T cells were further gated as FoxP3+. (B): Comparison of CD4+ T cell subsets (CD127-CD25-, CD127+CD25lo/- and CD127loCD25hi FoxP3+) in chronic HCV infection (dark grey boxes) and healthy controls (open boxes). (C): Comparison of CD4+ T cell subsets (naïve, central memory, effector memory and effector) based on expressions of CD45RA and CD27 in chronic HCV infection (dark grey boxes) and healthy controls (open boxes). Error bars illustrate s.d. *P < 0.05; **P < 0.01.
Expression of PD-1 on different CD4+T cell subsets defined by CD27 and CD45RA
The PD-1 expression on different CD4+T cell subsets defined by CD27 and CD45RA was determined by percentage of PD-1 positive cells and mean fluorescent intensity (MFI) of PD-1. PD-1expression was significantly increased in total CD4+T cells and all four CD4+T cell subsets (naïve/TCM/TEM/effector) in CHC patients compared with HCs (Figure 2A, B, C). Of note, expression of PD-1 was higher on TEM/effector subsets than naive/TCM subsets in both CHC patients and HCs (Figure 2B and 2C).

Percentage of PD-1 positive cells on total CD4+ T cells and its subsets. (A): Representative dot plot analysis showing the expression of PD-1 on total CD4+ T cells and naïve, TCM, TEM, and effector subsets. PD-1 expressions on total CD4+ T cells and all four subsets were significantly higher in chronic HCV-infected patients (half black symbols) than in healthy controls (open symbols) presented by both percentage (B) and MFI (C). *P < 0.05; **P < 0.01; ***P < 0.001.
Expression of PD-1 on CD4+CD127loCD25hiFoxP3+ Tregs
We next examined and compared the expression profile of PD-1 on different CD4+T cell subsets defined by CD127 and CD25. PD-1 expression was remarkably higher on all three CD4+T cell subsets (CD127+CD25lo/-, CD127-CD25-, and CD127loCD25hiFoxP3+) in CHC patients compared to HCs (p < 0.0001) (Figure 3A, B and 3C). In addition, there was a significant decrease of PD-1 expression on CD127loCD25hiFoxP3+ Tregs compared with CD127+CD25lo/- (p < 0.0001 on percentage and MFI) or CD127-CD25-(p = 0.0003 on percentage and p = 0.0045 on MFI) T cell subsets in CHC patients (Figure 3B and 3C). The similar trend was found in CHC patients (Figure 3B and 3C).

Percentage of PD-1 positive cells on CD127-CD25-, CD127+CD25lo/- and CD127loCD25hi FoxP3+ CD4+ T cell subsets. (A): Representative dot plot analysis showing the expression of PD-1 on these three subsets. PD-1 expressions on these T cell subsets were significantly higher in chronic HCV-infected patients (half black symbols) than in healthy controls (open symbols) presented by both of percentage (B) and MFI (C). PD-1 expression on Treg cells (CD127loCD25hi FoxP3+) was lower than on paired CD127-CD25- and CD127+CD25lo/- subsets. ▵indicated that Wilcoxon match pairs test. *P < 0.05; **P < 0.01; ***P < 0.001.
The dominance of the central memory phenotype in CD4+CD127loCD25hiFoxP3+ Tregs in CHC patients
CD127loCD25hiFoxP3+ Tregs were divided into Naïve-Tregs, TCM-Tregs, TEM-Tregs, Effector-Tregs depending on their expressions of CD27 and CD45RA (Figure 4A). As shown in Figure 4B, the frequency of Naïve-Tregs was dramatically reduced in CHC patients compared with HC individuals (p < 0.0001), while expression of TEM/Effector-Tregs was dramatically enhanced in CHC patients compared with HCs (p < 0.0001). No statistical difference was observed regarding to the proportion of TCM-Tregs in total CD4+Tregs between CHC patients and HCs (Figure 4B). It should be pointed out that the highest proportion (nearly 50%) of Tregs carried central memory phenotype in CHC patients due to a decreased proportion of naïve-Tregs (Figure 4B).

The dominant proportion of the central memory phenotype was found in CD127loCD25hiFoxP3+ Tregs in CHC patients. (A) Representative dot plot analysis showing the phenotypic distribution of CD127loCD25hiFoxP3+ Tregs depending on expression of CD27 and CD45RA. (B) Frequency of Naïve -Tregs (□) was dramatically reduced in CHC patients compared with HCs while TEM (▥)/Effector (▦) -Tregs showed a higher levels in CHC compared with in HCs. (C) The percentage of naïve and effector subset decreased while the percentage of TCM subset increased in Tregs compared with in their total CD4+ T cell counterparts. *P < 0.05; **P < 0.01; ***P < 0.001.
In addition, the distributional pattern of different CD4+T cell subsets was evaluated and compared in total CD4+T cells and Tregs. The frequency of naïve cells displayed significantly declined trends from total CD4+T cells to Tregs (p < 0.0001) in both CHC and HCs. In contrast, a significantly increased frequency of TCM phenotype was found in Tregs compared with total CD4+T cells (p < 0.0001) (Figure 4C). The proportion of terminal differentiated effector cells was downregulated in Tregs (p = 0.0003 in CHC and p = 0.0063 in HCs) while no difference in TEM frequency was found between total CD4+T cells and Tregs (Figure 4C). Therefore, this results indicated that central memory CD4+T cells was the dominant subset in Tregs, instead of naïve CD4+T cells which contributed to the majority of subset in total CD4+T cells.
Expression of PD-1 on different CD4+CD127loCD25hi FoxP3+ Tregs subsets defined by CD27 and CD45RA
PD-1 expression on different Tregs subsets was studied. PD-1 expression was significantly higher on total Tregs (p < 0.0001) and all four Tregs subsets (Naïve, p < 0.0001; TCM, p < 0.0001; TEM, p < 0.0147; Effector, p < 0.0153) in CHC patients compared with HCs. Besides, PD-1 expression was higher in TEM/effector-Tregs compared with naïve/TCM-Tregs. TCM Tregs showed lower PD-1 expression than TEM subsets (CHC, p = 0.0125; HC, p < 0.0001) while higher than naïve subsets in both CHC patients and HCs (CHC, p < 0.0001; HC, p < 0.0001) (Figure 5A).

(A) Increased MFI of TEM/effector Treg cells compared with naïve/TCM Treg cells in CHC patients and HC controls; (B) Ratios of TEM-Tregs to naive-Tregs and TEM-Tregs to TCM-Tregs regarding to PD-1 MFI were significantly lower in CHC patients than in HCs. ▵indicated that Wilcoxon match pairs test. *P < 0.05; **P < 0.01; ***P < 0.001.
Since PD-1 expression was enhanced on all CD4+T cell subsets on both chronic HCV infected patients and healthy controls, it is necessary and informative to compare the ratio of TEM-Tregs to naive-Tregs and of TEM-Tregs to TCM-Tregs regarding to PD-1 MFI between CHC patients and HCs. The results shown in Figure 5B indicated both TEM-Tregs/naïve-Tregs ratio and TEM-Tregs/TCM-Tregs ratio were significantly lower (p < 0.0001) in CHC patients than health controls. Unfortunately, no correlation was found between these ratios and peripheral HCV RNA level (data not shown).
Discussion
Both CD4+CD127loCD25hi and CD4+FoxP3+were classic phenotypes of regulatory CD4+ T cells, which has been described by accumulating literatures [34–36]. In the present study, we employed CD4+CD127loCD25hiFoxP3+ cells as circulating natural Tregs to investigate distributional characteristics of naïve, memory and effector subsets of Tregs. PD-1 expression on Tregs and its subsets was also studied in chronic HCV infected patients and healthy controls. Previous studies demonstrated human Tregs could be phenotypically and functionally divided into resting Tregs (rTregs) and activated Tregs (aTregs) according to the expression of surface CD45RA. Both rTregs and aTregs were shown to suppress proliferation of CD25- CD4+ responder T cells in vitro and CD45RA- FoxP3low/- subsets were non-Treg cells since they had no suppressive function [30]. Herein, our study investigated the classification of Tregs based on the expression of CD45RA and CD27 (a TNFR family member) since total CD4+ T cells could be subdivided into four different subsets (naïve/central memory/effector memory/effector) according to the expression of CD45RA and CD27. Our results (Figure 4) indicated that the proportion of Tregs in peripheral bulk CD4+ T cells was enhanced in CHC patients. The phenotypic characteristics of Tregs were switched from nearly equal contribution of naïve and central memory T cell of peripheral Tregs in HCs to central memory T cell contributing to be the dominant one in CHC patients, suggesting that peripheral Tregs of CHC patients were changed quantitatively and functionally.
PD-1/PD-L1 pathway was proved to play a critical role in maintaining the balance between protective T cell responses and immunopathology induced by hyper-activated effector T cells [15, 16, 20, 37–40]. It has been reported that CD4+ T-cell responses, including virus-specific IFN-γ production, were severely suppressed in chronically HCV-infected subjects through PD-1/PD-L1 pathway [20]. Though PD-1 was demonstrated to be expressed on Tregs and negatively regulated CD4+CD25+Foxp3+ Tregs function by preventing STAT-5 phosphorylation in CHC patients [23], it was still far from well understanding the molecular mechanisms of PD-1/PD-L1 pathway involving in inhibition of CD4+ T cell function, including Foxp3- effector T cells, Foxp3- memory T cells and Tregs. Our data showed that PD-1 expression on total Tregs as well as its subsets was significantly higher in HCV infected patients than healthy controls (Figure 5A). This results were not consistent with the hypothesis that chronic HCV infection was associated with higher inhibitory effect of Tregs since proliferation of Tregs was inhibited by PD-1 and subsequent IL-2-driven STAT-5 phosphorylation. With further investigation, we found (Figure 5B) that the TEM-Tregs/naive-Tregs ratio and TEM-Tregs/TCM-Tregs ratio of PD-1 MFI were significantly lower in CHC patients than healthy controls, indicating that PD-1 expression on TEM-Tregs or effector-Tregs were inhibited in relative to its correspondent naïve- or TCM-Tregs in CHC patients compared with healthy controls. Theoretically, the relatively reduced PD-1 expression on TEM-Tregs and effector-Tregs could enhance the inhibitory capacity of activated Tregs on hyper-activated effector T cells, attributing to impaired T cell immune response and subsequently inducing long-term persistence of HCV infection.
Taken together, our findings provided insight into the negative regulatory mechanism of Tregs and PD-1 on CD4+T cell responses during chronic HCV infection. We demonstrated that most Tregs displayed a central memory phenotype and PD-1 expression had a trend of relatively lower upregulated expression on effector Tregs and effector memory Tregs in relative to their naïve- or TCM-Tregs counterparts in chronic HCV infected patients compared with healthy individuals. The cross talk between Treg cells and PD-1 induced inhibition in chronic HCV infection and its implication in dysfunction or impairment of CD4+ T cell responses should be interesting and suggestive for further exploration of persistent HCV infection associated immune pathogenesis.
References
Craxi A, Laffi G, Zignego AL: Hepatitis C virus (HCV) infection: a systemic disease. Mol Aspects Med 2008, 29: 85-95. 10.1016/j.mam.2007.09.017
Munir S, Saleem S, Idrees M, Tariq A, Butt S, Rauff B, Hussain A, Badar S, Naudhani M, Fatima Z, Ali M, Ali L, Akram M, Aftab M, Khubaib B, Awan Z: Hepatitis C treatment: current and future perspectives. Virol J 2010, 7: 296. 10.1186/1743-422X-7-296
Clark PJ, Thompson AJ, McHutchison JG: IL28B genomic-based treatment paradigms for patients with chronic hepatitis C infection: the future of personalized HCV therapies. Am J Gastroenterol 2010, 106: 38-45.
Schinazi RF, Bassit L, Gavegnano C: HCV drug discovery aimed at viral eradication. J Viral Hepat 2010, 17: 77-90. 10.1111/j.1365-2893.2009.01246.x
Aghemo A, Rumi MG, Colombo M: Pegylated interferons alpha2a and alpha2b in the treatment of chronic hepatitis C. Nat Rev Gastroenterol Hepatol 2010, 7: 485-494. 10.1038/nrgastro.2010.101
Bowen DG, Walker CM: Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature 2005, 436: 946-952. 10.1038/nature04079
Grakoui A, Shoukry NH, Woollard DJ, Han JH, Hanson HL, Ghrayeb J, Murthy KK, Rice CM, Walker CM: HCV persistence and immune evasion in the absence of memory T cell help. Science 2003, 302: 659-662. 10.1126/science.1088774
Lechner F, Wong DK, Dunbar PR, Chapman R, Chung RT, Dohrenwend P, Robbins G, Phillips R, Klenerman P, Walker BD: Analysis of successful immune responses in persons infected with hepatitis C virus. J Exp Med 2000, 191: 1499-1512. 10.1084/jem.191.9.1499
Fontenot JD, Gavin MA, Rudensky AY: Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol 2003, 4: 330-336.
Khattri R, Cox T, Yasayko SA, Ramsdell F: An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nat Immunol 2003, 4: 337-342.
von Boehmer H: Mechanisms of suppression by suppressor T cells. Nat Immunol 2005, 6: 338-344. 10.1038/ni1180
Ebinuma H, Nakamoto N, Li Y, Price DA, Gostick E, Levine BL, Tobias J, Kwok W, Chang KM: Identification and in vitro expansion of functional antigen-specific CD25+ FoxP3+ regulatory T cells in hepatitis C virus infection. J Virol 2008, 82: 5043-5053. 10.1128/JVI.01548-07
Boettler T, Spangenberg HC, Neumann-Haefelin C, Panther E, Urbani S, Ferrari C, Blum HE, von Weizsäcker F, Thimme R: T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J Virol 2005, 79: 7860-7867. 10.1128/JVI.79.12.7860-7867.2005
Cabrera R, Tu Z, Xu Y, Firpi RJ, Rosen HR, Liu C, Nelson DR: An immunomodulatory role for CD4(+)CD25(+) regulatory T lymphocytes in hepatitis C virus infection. Hepatology 2004, 40: 1062-1071. 10.1002/hep.20454
Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ: The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol 2007, 8: 239-245. 10.1038/ni1443
Keir ME, Francisco LM, Sharpe AH: PD-1 and its ligands in T-cell immunity. Curr Opin Immunol 2007, 19: 309-314. 10.1016/j.coi.2007.04.012
Nishimura H, Okazaki T, Tanaka Y, Nakatani K, Hara M, Matsumori A, Sasayama S, Mizoguchi A, Hiai H, Minato N, Honjo T: Autoimmune dilated cardiomyopathy in PD-1 receptor-deficient mice. Science 2001, 291: 319-322. 10.1126/science.291.5502.319
Nishimura H, Nose M, Hiai H, Minato N, Honjo T: Development of lupus-like autoimmune diseases by disruption of the PD-1 gene encoding an ITIM motif-carrying immunoreceptor. Immunity 1999, 11: 141-151. 10.1016/S1074-7613(00)80089-8
Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne MC, Horton HF, Fouser L, Carter L, Ling V, Bowman MR, Carreno BM, Collins M, Wood CR, Honjo T: Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med 2000, 192: 1027-1034. 10.1084/jem.192.7.1027
Urbani S, Amadei B, Tola D, Massari M, Schivazappa S, Missale G, Ferrari C: PD-1 expression in acute hepatitis C virus (HCV) infection is associated with HCV-specific CD8 exhaustion. J Virol 2006, 80: 11398-11403. 10.1128/JVI.01177-06
Golden-Mason L, Palmer B, Klarquist J, Mengshol JA, Castelblanco N, Rosen HR: Upregulation of PD-1 expression on circulating and intrahepatic hepatitis C virus-specific CD8+ T cells associated with reversible immune dysfunction. J Virol 2007, 81: 9249-9258. 10.1128/JVI.00409-07
Radziewicz H, Ibegbu CC, Fernandez ML, Workowski KA, Obideen K, Wehbi M, Hanson HL, Steinberg JP, Masopust D, Wherry EJ, Altman JD, Rouse BT, Freeman GJ, Ahmed R, Grakoui A: Liver-infiltrating lymphocytes in chronic human hepatitis C virus infection display an exhausted phenotype with high levels of PD-1 and low levels of CD127 expression. J Virol 2007, 81: 2545-2553. 10.1128/JVI.02021-06
Franceschini D, Paroli M, Francavilla V, Videtta M, Morrone S, Labbadia G, Cerino A, Mondelli MU, Barnaba V: PD-L1 negatively regulates CD4+CD25+Foxp3+ Tregs by limiting STAT-5 phosphorylation in patients chronically infected with HCV. J Clin Invest 2009, 119: 551-564. 10.1172/JCI36604
Radziewicz H, Dunham RM, Grakoui A: PD-1 tempers Tregs in chronic HCV infection. J Clin Invest 2009, 119: 450-453. 10.1172/JCI38661
Sallusto F, Geginat J, Lanzavecchia A: Central memory and effector memory T cell subsets: function, generation, and maintenance. Annu Rev Immunol 2004, 22: 745-763. 10.1146/annurev.immunol.22.012703.104702
Humphreys TL, Baldridge LA, Billings SD, Campbell JJ, Spinola SM: Trafficking pathways and characterization of CD4 and CD8 cells recruited to the skin of humans experimentally infected with Haemophilus ducreyi. Infect Immun 2005, 73: 3896-3902. 10.1128/IAI.73.7.3896-3902.2005
Schiott A, Lindstedt M, Johansson-Lindbom B, Borrebaeck C: CD27- CD4+ memory T cells define a differentiated memory population at both the functional and transcriptional levels. Immunology 2004, 113: 363-370. 10.1111/j.1365-2567.2004.01974.x
De Jong R, Brouwer M, Hooibrink B, Van der Pouw-Kraan T, Miedema F, Van Lier RA: The CD27- subset of peripheral blood memory CD4+ lymphocytes contains functionally differentiated T lymphocytes that develop by persistent antigenic stimulation in vivo. Eur J Immunol 1992, 22: 993-999. 10.1002/eji.1830220418
Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A: Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401: 708-712. 10.1038/44385
Miyara M, Yoshioka Y, Kitoh A, Shima T, Wing K, Niwa A, Parizot C, Taflin C, Heike T, Valeyre D, Mathian A, Nakahata T, Yamaguchi T, Nomura T, Ono M, Amoura Z, Gorochov G, Sakaguchi S: Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009, 30: 899-911. 10.1016/j.immuni.2009.03.019
Sardella G, De Luca L, Francavilla V, Accapezzato D, Mancone M, Sirinian MI, Fedele F, Paroli M: Frequency of naturally-occurring regulatory T cells is reduced in patients with ST-segment elevation myocardial infarction. Thromb Res 2007, 120: 631-634. 10.1016/j.thromres.2006.12.005
Coenen JJ, Koenen HJ, van Rijssen E, Hilbrands LB, Joosten I: Rapamycin, and not cyclosporin A preserves the highly suppressive CD27+ subset of human CD4+CD25+ regulatory T cells. Blood 2006, 107: 1018-1023.
Koenen HJ, Fasse E, Joosten I: CD27/CFSE-based ex vivo selection of highly suppressive alloantigen-specific human regulatory T cells. J Immunol 2005, 174: 7573-7583.
Sakaguchi S, Yamaguchi T, Nomura T, Ono M: Regulatory T cells and immune tolerance. Cell 2008, 133: 775-787. 10.1016/j.cell.2008.05.009
Dunham RM, Cervasi B, Brenchley JM, Albrecht H, Weintrob A, Sumpter B, Engram J, Gordon S, Klatt NR, Frank I, Sodora DL, Douek DC, Paiardini M, Silvestri G: CD127 and CD25 expression defines CD4+ T cell subsets that are differentially depleted during HIV infection. J Immunol 2008, 180: 5582-5592.
Seddiki N, Santner-Nanan B, Martinson J, Zaunders J, Sasson S, Landay A, Solomon M, Selby W, Alexander SI, Nanan R, Kelleher A, Fazekas de St Groth B: Expression of interleukin (IL)-2 and IL-7 receptors discriminates between human regulatory and activated T cells. J Exp Med 2006, 203: 1693-1700. 10.1084/jem.20060468
Ha SJ, Mueller SN, Wherry EJ, Barber DL, Aubert RD, Sharpe AH, Freeman GJ, Ahmed R: Enhancing therapeutic vaccination by blocking PD-1-mediated inhibitory signals during chronic infection. J Exp Med 2008, 205: 543-555. 10.1084/jem.20071949
Day CL, Kaufmann DE, Kiepiela P, Brown JA, Moodley ES, Reddy S, Mackey EW, Miller JD, Leslie AJ, DePierres C, Mncube Z, Duraiswamy J, Zhu B, Eichbaum Q, Altfeld M, Wherry EJ, Coovadia HM, Goulder PJ, Klenerman P, Ahmed R, Freeman GJ, Walker BD: PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 2006, 443: 350-354. 10.1038/nature05115
Boni C, Fisicaro P, Valdatta C, Amadei B, Di Vincenzo P, Giuberti T, Laccabue D, Zerbini A, Cavalli A, Missale G, Bertoletti A, Ferrari C: Characterization of hepatitis B virus (HBV)-specific T-cell dysfunction in chronic HBV infection. J Virol 2007, 81: 4215-4225. 10.1128/JVI.02844-06
Penna A, Pilli M, Zerbini A, Orlandini A, Mezzadri S, Sacchelli L, Missale G, Ferrari C: Dysfunction and functional restoration of HCV-specific CD8 responses in chronic hepatitis C virus infection. Hepatology 2007, 45: 588-601. 10.1002/hep.21541
Acknowledgements
We thanks all participants involved in this study. This work was supported by grants from the National S&T Major Project for Infectious Diseases (2008ZX10002-012, 2008ZX10002-013, and 2009ZX10004-903).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Authors' contributions
TS, HL and FL design the study. TS and JZ performed the statistical analysis and interpretation of the data. TS and HL drafted the manuscript. TS, JZ, CX, TZ, and QX collected samples and performed benchwork. All suthors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Tao Shen, Jiajia Zheng contributed equally to this work.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Shen, T., Zheng, J., Liang, H. et al. Characteristics and PD-1 expression of peripheral CD4+CD127loCD25hiFoxP3+ Treg cells in chronic HCV infected-patients. Virol J 8, 279 (2011). https://doi.org/10.1186/1743-422X-8-279
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1743-422X-8-279