
Abstract
Background
Psoriatic arthritis (PsA) is a distinct inflammatory arthritis occurring in 30% of psoriasis patients. There is a high prevalence of undiagnosed PsA in psoriasis patients; therefore, identifying soluble biomarkers for PsA could help in screening psoriasis patients for appropriate referral to a rheumatologist. Potential PsA biomarkers likely originate in sites of inflammation, such as the skin, and subsequently enter systemic circulation. Our goal was to identify candidate PsA biomarkers by comparing the proteome of skin biopsies obtained from patients with PsA to that from patients with psoriasis without PsA.
Methods
Skin biopsies were obtained from involved and uninvolved skin of 10 PsA and 10 age/gender-matched psoriasis patients without PsA (PsC). Using strong cation exchange chromatography, followed by label-free quantitative tandem mass spectrometry, we characterized the proteomes of pooled skin samples. Extracted ion current intensities were used to calculate protein abundance ratios, and these were utilized to identify differentially regulated proteins.
Results
Forty-seven proteins were elevated in PsA-derived skin compared to PsC-derived skin. Selected reaction monitoring assays were developed to quantify these potential PsA markers in individual skin samples, and 8 markers were confirmed in an independent sample set. ITGB5 and POSTN were measured in serum samples from 33 PsA and 15 PsC patients, using enzyme-linked immunosorbent assays. ITGB5 was significantly elevated in PsA serum (P < 0.01), and POSTN showed a trend. ITGB5 and POSTN correlated significantly in both patient groups (r = 0.472, P < 0.001).
Conclusion
Proteomic analysis of PsA and PsC skin identified eight new candidate biomarkers. These markers need to be validated with a larger and independent cohort, in order to delineate their clinical utility in PsA patients. These proteins may also uncover unknown aspects of PsA pathobiology.
Similar content being viewed by others


Background
Psoriatic arthritis (PsA) is a distinct inflammatory arthritis, which takes its name from its association with the cutaneous, autoimmune inflammatory disease, psoriasis. It occurs in 30% of psoriasis patients and has a predicted prevalence of up to 1% in the general population. PsA is a complex, potentially disabling musculoskeletal disorder often arising early in age. Patients with PsA have an increased risk for a spectrum of co-morbidities, such as obesity, metabolic syndrome, diabetes and cardiovascular disease [1–3]. The diagnosis of PsA presents a challenge, largely due to its heterogeneous clinical presentation [4, 5]; however, early diagnosis and prognosis of PsA is essential for prevention of joint damage and disability [6].
The key to early diagnosis is a better recognition of PsA in patients with psoriasis, since its presence indicates a high risk for current or future development of PsA [3]. Soluble biomarkers represent an ideal means for screening patients for PsA. With improvements in high-throughput genomic platforms, a number of putative markers, ranging from susceptibility genes to mRNA profiles have been proposed [7–11]; however there exists no single or panel of specific markers, or mediating factor(s). Much of the research, therefore, focuses on the discovery and validation of PsA biomarkers [7, 12–14]. Identifying these factors would not only facilitate diagnosis and prognosis of PsA, but also provide further insight into disease pathogenesis.
Mass spectrometry (MS)-based quantitative proteomic approaches are well-suited for the discovery of protein mediators and biomarkers of disease [15, 16]. Specifically, label-free quantification (LFQ) methods have been recently optimized, where quantification is based on the differential peak intensity [extracted ion current (XIC)] of the peptides in each MS scan [17, 18]. Such experiments often result in tens to hundreds of candidate biomarkers, therefore, in this context of biomarker discovery, high-throughput mass spectrometry-based proteomics is a powerful tool for obtaining disease-specific proteome profiles of biological materials [19]. While human plasma represents a diverse proteome and is an excellent source of potential disease markers, proteins secreted by tissues are diluted in blood, and are often undetectable by current MS methods [19]. Much interest has been given to the analysis of proximal fluids and tissues, such as synovial fluid and skin [19–21].
Cutaneous psoriasis develops simultaneously or precedes the onset of PsA in up to 90% of PsA patients, and in order to move forward in the search for PsA screening biomarkers, we must consider the preceding cutaneous psoriasis stage in PsA patients. Therefore, hypothesizing that there are differences in the skin proteome of patients with PsA compared to those with psoriasis but without PsA (PsC), comparative analysis of skin between these two patient groups represents a reasonable experimental workflow. In a pilot study, our group demonstrated that proteins elevated in the inflamed skin, are likewise upregulated at the serum level, and may serve as putative markers of psoriasis [22].
In the current study, we performed label-free quantitation of skin proteins from PsA and PsC patients. Using selected reaction monitoring assays (SRM) we confirmed the elevation of some proteins in an independent set of samples from patients with PsA. Following a small-scale validation in serum using enzyme-linked immunosorbent assays (ELISA), we confirmed a significant elevation of β5 Integrin (ITGB5) in the serum of PsA patients. Periostin (POSTN) also showed a similar trend. These proteins may serve as potential PsA biomarkers and also shed new light into the pathogenesis of PsA.
Results
Delineating the PsA skin proteome
Our LC-MS/MS analysis yielded 1922 quantifiable proteins (Additional file 1: Table S1). Student’s t-tests, and a false discovery rate (FDR) threshold of 0.2, were utilized to identify upregulated proteins between the PsA and PsC lesional groups (PsA L, and PsC L, respectively). This comparative analysis generated a total of 62 proteins (Additional file 2: Table S2). Generally, these proteins exhibited a fold change ratio above 4.0. Comparison between PsA and PsC non-lesional skin (PsA N, and PsC N, respectively), at the same FDR, demonstrated elevated expression of 131 proteins (Additional file 2: Table S2). By comparing the proteins identified from the two independent analyses (PsA L vs PsC L, and PsA N vs. PsC N), only 7 of these proteins were common (Additional file 2: Table S2). These were PSME3, C1QC, RENBP, GRHPR, POLE, POSTN, and IGLV3-21.
Since our scope was to identify potential biomarkers present in PsA skin, we decided to focus on the 62 proteins overexpressed in the lesional PsA skin, which included the seven proteins elevated in non-lesional skin, since they are more likely to represent mediators and potential biomarkers of PsA. We further focused on proteins displaying strong expression in skin, bone, and immune regulatory cells, and excluded immunoglobulins, unknown proteins and high abundance serum proteins from further analysis. Additional file 3: Table S3 presents the final list of 47 proteins, corresponding fold changes (FC) between PsA and PsC samples, P-Values and FDR values. These were then assessed and quantified, using a multiplexed selected reaction monitoring assay, in individual skin samples included in the discovery set (Set I), as well as in an independent set of 10 samples (Set II).
SRM Verification of putative markers in individual skin samples
We developed a multiplexed SRM assay to verify the differences in protein expression between lesional PsA and PsC skin. Assays were developed for 47 peptides representative of the 47 proteins with increased expression in the skin of PsA patients, as well as 4 peptides representing 2 housekeeping proteins (ACTB, TUBB) (Additional file 4: Table S4). Consistent with our LC-MS/MS analysis of the pooled samples, overexpression of 12 out of the 47 proteins was also verified in the individual PsA lesional skin samples (Set I), when compared to lesional PsC samples. Each sample consisted of two technical replicates. The mean fold change of each confirmed protein in the lesional and non-lesional PsA and PsC skin samples, as well as the associated P-values are depicted in Table 1. The proteins included C16ORF62, SNCA, LZIC, SRP14, ITGB5, POSTN, SRPX, FHL1, PPP2R4, CPN2, GPS1, and PAFAH1B2. The distribution of these, and the housekeeping proteins across the Set I skin samples, are represented in Additional file 5: Figure S1.
Additionally, we further confirmed the elevation of 8 of the aforementioned 12 proteins, in an independent set of 5 PsA, and 5 PsC lesional skin samples (Set II). In this case, we utilized a heavy-labeled peptide in order to obtain the absolute concentration of these peptides in the skin. The proteins included SRP14, ITGB5, POSTN, SRPX, FHL1, PPP2R4, CPN2, and GPS1. The mean fold changes and P-values corresponding to each protein from set II are also depicted in Table 1. Additional file 6: Figure S2 shows the distribution of the 8 proteins across the Set II skin samples, as well as of the housekeeping proteins. Since only 8 proteins were confirmed in the independent sample set, we decided to consider for further analyses only these 8 potential markers.
Small-scale ELISA validation in serum
To validate the possible expression of markers in serum of PsA patients, we measured the levels of ITGB5 and POSTN in the serum of 15 PsC and 33 PsA patients. As shown in Figure 1, ITGB5 was significantly elevated in PsA, when compared to PsC patients (1.19 ± 0.5 compared to 0.77 ± 0.6; p < 0.01). The concentration of POSTN was not significantly different between the two groups (p > 0.05), but the levels in PsA serum showed an increasing trend, when compared to PsC serum (17.71 ± 6.4 compared to 14.53 ± 6.2). The mean FC and distribution of POSTN and ITGB5 in the PsA and PsC serum, are represented in Figure 1. We were unable to measure SRP14, SRPX, FHL1, PPP2R4, CPN2, and GPS1, due to unavailability of ELISA kits, antibodies, or protein standards.

Distribution of markers across the PsA and PsC serum sets. Small-scale validation of ITGB5 (A) and POSTN (B) in PsA and PsC serum by ELISA. Dots represent serum samples from individual subjects; thin horizontal lines depict the mean, and vertical lines the standard deviation; FC represents the ratio of the mean concentration values corresponding to 33 PsA and 15 PsC serum samples. ** indicates P < 0.01; ns: non-significant. PsA, Psoriatic Arthritis; PsC, Cutaneous Psoriasis.
Correlation amongst biomarkers
Spearman’s rank correlation coefficient was used to assess the correlation amongst markers for the PsA, and PsC serum groups. ITGB5 correlated significantly with POSTN in PsC serum (Spearman r = 0.637, P = 0.013), and in PsA serum (Spearman r = 0.433, P = 0.012). Figure 2 displays the correlation between POSTN and ITGB5 in all samples (Spearman r = 0.472, P < 0.001).

Correlation between ITGB5 and POSTN across the PsA and PsC serum sets. Dots represent serum samples from individual subjects. r indicates the Spearman’s rank correlation coefficient.
Discussion
Recent advances in mass spectrometry-based proteomics facilitated the identification of putative novel biomarkers and mediators from a variety of tissues, and for various clinical applications [15, 16, 23]. In this study, we conducted high-throughput quantitative proteomics to identify differentially expressed proteins between skin derived from PsA and PsC patients. According to our filtering criteria, forty-seven proteins were elevated in the PsA group, when compared to the PsC group. Eight proteins were verified using a multiplexed selected reaction monitoring assay, in an independent skin sample set. None of these potential markers has been described before in the context of PsA or psoriasis. Since serum measurement of these proteins using SRMs is not efficient due to the high complexity of the fluid [18], we utilized available enzyme-linked immunosorbent assays to measure two biomarker candidates in serum (ITGB5, and POSTN).
Cumulative evidence strongly supports the involvement of Interleukin (IL)-23/IL-17 axis in the pathogenesis of PsA, and a number of compounds that target components of these pathways have been recently used in PsA clinical trials. IL-23 acts synergistically with IL-6 and TGF-β to promote rapid Th17 development and IL-17 release [24, 25], which, in turn, plays a central role in sustaining chronic inflammation [24]. Both POSTN and ITGB5 have been implicated with this pathway.
Periostin is a matricellular protein belonging to the fascilin family [26]. It interacts with several integrin molecules on cell surfaces, one of which is α5β5 Integrin, and provides signals for tissue development and remodeling [27]. More specifically, it is thought that POSTN interacts with α5β5 to induce pro-inflammatory cytokine production via the Akt/NF-κB signaling pathway [28]. In support of this, deficiency of POSTN or inhibition of α5 integrin (ITGA5) prevented development or progression of skin inflammation in an atopic dermatitis mouse model [28]. Additionally, POSTN has been shown to also act on immune cells, leading to their enhanced transmigration, chemotaxis, and adhesion [29], all of which further implicate this molecule in inflammatory processes relevant to PsA.
Apart from acting as a POSTN ligand (along with ITGA5), ITGB5 has also been implicated in rheumatoid arthritis, where it serves as a ligand for Cyr61 [30]. Cyr61 is a molecule secreted by fibroblast-like synoviocytes in the joint, and stimulates IL-6 production via ITGA5-ITGB5/Akt/NF-κB signaling pathway [29]. As described earlier, IL-6, along with IL-23 and TGB-β, trigger Th17 differentiation and IL-17 production [24, 25] in inflammatory processes, including PsA. Based on this data, we suggest that POSTN and ITGB5 are attractive molecules to investigate further as PsA biomarkers.
Following a small-scale serum validation of POSTN and ITGB5, we determined that only ITGB5 was significantly elevated in PsA serum, although POSTN also showed a trend. Additionally, Spearman correlation showed that serum ITGB5 correlates well with POSTN (r = 0.472, p < 0.001), which indicates that these two markers may, in the future, be used as part of a panel of markers to screen for PsA in PsC patients.
Although our findings are promising, the work reported here has some limitations. First, pooling of the skin samples in the discovery phase has advantages and disadvantages. While pooling allows for a more extensive coverage of disease heterogeneity, by increasing the likelihood of identifying proteins that are otherwise undetectable in some individual samples, it may also mask meaningful discrepancies among the different individual skin proteomes [19, 31]. To minimize this effect, samples were pooled to obtain two distinct pools in both, PsA and PsC groups. Additionally, all pool-derived candidates were further examined in all individual samples using a targeted and more sensitive mass spectrometry technique. As our results indicate, the correlation between the discovery data and SRM data is not ideal, whereby only 12 proteins were verified in the discovery set. This is due in part, to the “pooling” effects previously discussed. Our SRM data from individual samples identified several skin samples that had prominently higher protein concentrations. These samples most probably affected (increased) the final concentration of proteins in the pooled samples, which were used for our initial selection of candidates. Thus, analysis of individual samples via SRM assay allowed for elimination of such artifacts.
Second, the identified markers were only tested on a small number of serum samples (n = 48), and only 20 of these were distinct from the Set I and Set II sample sets. Therefore, to assess their accuracy in serum, these markers still need to be tested in a large-scale verification study. In addition, as discussed previously, only two of the existing eight proposed markers have been investigated in serum due to the lack of ELISAs or antibodies for the rest. Hence, strategies to enable measurement of the remainder of the markers will be employed in the future.
Finally, the 47 proteins we investigated using SRM, represent few of the candidates we have presently identified. Proteins that were elevated in non-lesional skin also represent interesting candidates, and must be measured in the future.
Taken together, our current observations are consistent with the notion that label-free quantitative LC-MS/MS can be utilized to quantify proteins in tissues, which can then be shed into the circulation and serve as serum markers of PsA. Further studies to validate these findings are underway, in a larger and independent serum cohort. Investigating these proposed markers further, may not only result in the identification of PsA screening biomarkers, but may also uncover aspects of PsA pathobiology that are currently unknown.
Methods
Skin proteomic analysis
Clinical samples
Samples were collected with informed consent after research ethics board approval from the University Health Network, Toronto, Canada. For the discovery phase, skin biopsies were obtained from 10 cases with PsA (6 males, 4 females; age range 38-73 years) and 10 PsC patients (6 males, 4 females; age range 28-77 years) (Set I). PsA patients had psoriasis and satisfied the CASPAR classification criteria [32], while the PsC patients were assessed by a rheumatologist, to exclude PsA. Patients were not undergoing treatment with methotrexate (MTX) or anti-TNF agents. One 6mm punch biopsy was obtained from unaffected (non-lesional) skin, and one from affected (lesional) skin from each PsA and PsC patient, amounting to a total of 40 samples.
For the verification (quantification) phase, an independent set of skin samples was obtained from 5 PsA patients (all males; age range 49-63 years), and 5 PsC patients (3 males, 2 females; age range 49-67 years) (Set II). Inclusion criteria were the same as described above.
For the small-scale validation, serum samples were obtained from 33 PsA patients (22 males, 11 females; age range 21-76 years), and 15 PsC controls (9 males, 6 females, age range 28-77 years). Inclusion criteria were the same as described above.
Pre-analytical sample processing
Skin samples were snap-frozen in liquid nitrogen and stored at -80°C until use. Samples were suspended in 0.05% RapiGest (Waters, Milford, MA, USA) buffer, and the tissue was homogenized and sonicated for protein extraction. The tissue lysates were spun at 11,000g and total protein was measured by the Bradford assay (Pierce Biotechnology, Rockford, IL, USA), in the resulting supernatants. Equal protein amounts from each sample were pooled to create eight different pools of five samples each [2 PsA lesional (PsA L) vs. 2 PsA non-lesional (PsA N) vs. 2 PsC lesional (PsC L) vs. 2 PsC non-lesional (PsC N)]. Proteins in each pool were denatured and reduced with 5 mM dithiothreitol at 60°C for 45 minutes and alkylated with 15 mM iodoacetamide, in the dark, at room temperature, for 45 minutes. Sequencing grade trypsin (Promega, Madison, WI, USA) was added in a 1:50 (trypsin: protein) ratio, and allowed to digest for 18 hours at 37°C. Trifluoroacetic acid (1%) was added to cleave RapiGest and inhibit trypsin activity, and samples were centrifuged at 11,000g for 20 minutes prior to high performance liquid chromatography (HPLC) using strong cation exchange (SCX), to reduce peptide mixture complexity. During the initial stages of SCX, buffer blanks were run after each sample, and the resulting fractions were assessed for peptide carryover; apart from peptides corresponding to a few high abundance proteins, we found very few peptides (data not shown). The peptide fractions resulting from HPLC-SCX were collected, pooled, and subjected to liquid chromatography and tandem mass spectrometry (LC-MS/MS), and was followed by protein identification and quantification. Experimental details regarding HPLC-SCX, LC-MS/MS, and protein identification and quantification, were previously published [31], and are also outlined in Additional file 7.
Bioinformatic analysis
To detect upregulated proteins in the PsA group, one-sided Student’s t-tests were used between PsA L and PsC L, and PsA N and PsC N after log transformation of XIC values. Only proteins displaying significant differences [False discovery rate (FDR) <0.2] were used for further analysis [33]. The FDR threshold was chosen such that if we repeat the procedure many times, 20% of the significant differences are expected to be false, on average. Averages of the replicate XIC values were calculated for each PsA L, PsA N, PsC L, and PsC N, and ratios (fold change: FC) of PsA L/PsC L, and PsA N/PsC N were computed.
Gene names of the upregulated proteins were checked against gene [BioGPS (http://biogps.org/#goto=welcome) [34]], and protein [Human Protein Atlas (http://www.proteinatlas.org/) [35] databases, to identify proteins with strong expression in PsA-associated tissues and cell types (skin, bone, immune cells). The Plasma Proteome Database (http://www.plasmaproteomedatabase.org/) [36] was employed to identify proteins present in high-abundance in serum, which could represent potential contaminants. Selected reaction monitoring (SRM) assays were developed for the top upregulated proteins in the lesional PsA group and housekeeping (ACTB, TUBB) proteins, and relative protein quantification was performed in individual skin samples to confirm their elevation in PsA skin.
Verification of identified proteins using SRM
SRM methods were developed for verification of protein ratios in skin, following our previously published protocols [23, 31]. Details regarding the SRM assay development, skin sample preparation, and subsequent protein quantification are also outlined in Additional file 7.
Validation of verified proteins
Enzyme-linked immunosorbent assays (ELISA)
The concentration of ITGB5 in serum was measured using an in-house developed protocol, the details of which are outlined in Additional file 7. Serum was diluted 10-fold.
The concentration of POSTN in serum was measured using a multiplexed Luminex Screening Assay (R&D Systems), according to manufacturer’s instructions. Serum was diluted 100-fold.
Statistical analysis
The Student’s t-tests were performed to analyze the LC-MS/MS data using R statistical software (http://www.r-project.org/). Other statistical analyses were performed using Graph Pad Prism v.6.0 for Mac (GraphPad Software, CA, USA). Results were analyzed using the nonparametric Mann-Whitney U test. P-values less than 0.05 were considered statistically significant. Spearman's rank correlation coefficient was used to assess the correlations between serum levels of POSTN, and ITGB5. Concentrations are reported in the text as mean ± standard deviation (SD).
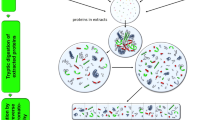
The overall experimental design is provided in Figure 3.

Summary of the experimental design. See text and abbreviations for more details.
Abbreviations
- ACE:
-
Angiotensin-converting enzyme
- ACN:
-
Acetonitrile
- ACTB:
-
Beta-actin
- ACTH:
-
Adrenocorticotropic hormone
- C16ORF62:
-
Chromosome 16 open reading frame 62
- C1QC:
-
Complement C1q
- CASPAR:
-
Classification criteria for Psoriatic Arthritis
- CPN2:
-
Carboxypeptidase N, polypeptide 2
- ELISA:
-
Enzyme-linked immunosorbent assay
- FA:
-
Formic Acid
- FC:
-
Fold change
- FDR:
-
False discovery rate
- FHL1:
-
Four and a half LIM domain1 1
- GPS1:
-
G-protein pathway suppressor 1
- GRHPR:
-
Glyoxylate reductase/hydroxypyruvate reductase
- HPLC:
-
High-performance liquid chromatography
- IGLV3-21:
-
Immunoglobulin Lambda Variable 3-21
- IL:
-
Interleukin
- ITGA5:
-
Integrin alpha 5
- ITGB5:
-
Integrin-beta 5
- LC-MS/MS:
-
Liquid chromatography and tandem mass spectrometry
- LFQ:
-
Label-free quantification
- LZIC:
-
Leucine Zipper and CTNNBIP1 Domain-Containing Protein
- MS:
-
Mass-Spectrometry
- MTX:
-
Methotrexate
- PAFAH1B2:
-
Platelet-activating factor acetylhydrolase 1b, catalytic subunit 2
- POLE:
-
DNA Polymerase II Subunit A
- POSTN:
-
Periostin
- PPP2R4:
-
Protein phosphatase 2A activator, regulatory subunit 4
- PsA L:
-
Lesional Psoriatic Arthritis
- PsA N:
-
Non-lesional Psoriatic Arthritis
- PsA:
-
Psoriatic Arthritis
- PsC L:
-
Lesional Cutaneous Psoriasis
- PsC N:
-
Non-lesional Cutaneous Psoriasis
- PsC:
-
Cutaneous Psoriasis
- PSME3:
-
Activator of Multicatalytic Protease Subunit 3
- SCX:
-
Strong cation exchange
- SD:
-
Standard deviation
- SNCA:
-
Alpha synuclein
- SRM:
-
Selected reaction monitoring
- SRP14:
-
Signal recognition particle 14kDa
- SRPX:
-
Sushi-repeat containing protein
- TFA:
-
Trifluoroacetic acid
- TGF:
-
Transforming growth factor
- TNF:
-
Tumor necrosis factor
- TUBB:
-
Beta-tubulin
- XIC:
-
Extracted Ion Current.
References
Gladman DD, Antoni C, Mease P, Clegg DO, Nash P: Psoriatic arthritis: epidemiology, clinical features, course, and outcome. Ann Rheum Dis 2005,64(Suppl 2):ii14–7.
Chandran V, Raychaudhuri SP: Geoepidemiology and environmental factors of psoriasis and psoriatic arthritis. J Autoimmun 2010, 34:J314–21. 10.1016/j.jaut.2009.12.001
Zachariae H: Prevalence of joint disease in patients with psoriasis. Am J Clin Dermatol 2003, 4:441–4. 10.2165/00128071-200304070-00001
Chandran V, Schentag CT, Gladman DD: Sensitivity and specificity of the CASPAR criteria for psoriatic arthritis in a family medicine clinic setting. J Rheumatol 2008, 35:2069–70.
Angelo D, Mennillo S, Cutro GA, Leccese MS, Nigro P, Padula A, Olivieri A: Sensitivity of the classification of psoriatic arthritis criteria in early psoriatic arthritis. J Rheumatol 2009, 36:368–70. 10.3899/jrheum.080596
Haroon M, Ghallagher P, FitzGerald O: Diagnostic delay of more than 6 months contributes to poor radiographic and functional outcome in psoriatic arthritis. Ann Rheum Dis Advance Online Publ doi:10.1136/annrheumdis-2013–204858
FitzGerald O, Mease PJ: Biomarkers: project update from the GRAPPA 2012 annual meeting. J Rheumatol 2013, 40:1453–4. 10.3899/jrheum.130462
Dominguez P, Gladman DD, Helliwell P, Mease PJ, Husni ME, Qureshi AA: Development of screening tools to identify psoriatic arthritis. Curr Rheumatol Rep 2010, 12:295–9. 10.1007/s11926-010-0113-2
Rahman P, Roslin NM, Pellett FJ, Lemire M, Greenwood CM, Beyene J, Pope A, Peddle L, Paterson AD, Uddin M, Gladman DD: High resolution mapping in the major histocompatibility complex region identifies multiple independent novel loci for psoriatic arthritis. Ann Rheum Dis 2011, 70:690–4. 10.1136/ard.2010.133561
Roberson ED, Bowcock AM: Psoriasis genetics: breaking the barrier. Trends Genet 2010, 26:415–23. 10.1016/j.tig.2010.06.006
Chandran V: The genetics of psoriasis and psoriatic arthritis. Clin Rev Allergy Immunol 2013, 44:149–56. 10.1007/s12016-012-8303-5
Bresnihan B: Synovial biomarkers in the spondylarthropathies. Curr Opin Rheumatol 2006, 18:359–63. 10.1097/01.bor.0000231903.27419.2d
Chen CH, Lin KC, Yu DTY, Yang C, Huang F, Chen HA, Liang HT, Tsai CY, Wei JC, Chou CT: Serum matrix metalloproteinases and tissue inhibitors of metalloproteinases in ankylosing spondylitis: MMP-3 is a reproducibly sensitive and specific biomarker of disease activity. Rheumatology 2006, 45:414–20. 10.1093/rheumatology/kei208
Yang C, Gu J, Rihl M, Baeten D, Huang F, Zhao M, Zhang H, Maksymowych WP, De Keyser F, Veys EM, Yu DT: Serum levels of matrix metalloproteinase 3 and macrophage colony-stimulating factor 1 correlate with disease activity in ankylosing spondylitis. Arthritis Rheum 2004, 51:691–9. 10.1002/art.20696
Drabovich AP, Dimitromanolakis A, Saraon P, Soosaipillai A, Batruch I, Mullen B, Jarvi K, Diamandis EP: Differential diagnosis of azoospermia with proteomic biomarkers ECM1 and TEX101 quantified in seminal plasma. Sci Transl Med 2013, 5:212ra160. 10.1126/scitranslmed.3006260
Martínez-Morillo E, García Hernández P, Begcevic I, Kosanam H, Prieto García B, Alvarez Menendez FV, Diamandis EP: Identification of novel biomarkers of brain damage in patients with hemorrhagic stroke by integrating bioinformatics and mass spectrometry-based proteomics. J Proteome Res 2014, 13:969–81. 10.1021/pr401111h
Megger DA, Bracht T, Meyer HE, Sitek B: Label-free quantification in clinical proteomics. Biochim Biophys Acta 1834, 2013:1581–90.
Yates JR, Ruse CI, Nakorchevsky A: Proteomics by mass spectrometry: approaches, advances, and applications. Annu Rev Biomed Eng 2009, 11:49–79. 10.1146/annurev-bioeng-061008-124934
Cretu D, Diamandis EP, Chandran V: Delineating the synovial fluid proteome: recent advancements and ongoing challenges in biomarker research. Crit Rev Clin Lab Sci 2013, 50:51–63. 10.3109/10408363.2013.802408
Eming SA, Koch M, Krieger A, Brachvogel B, Kreft S, Bruckner-Tuderman L, Krieg T, Shannon JD, Fox JW: Differential proteomic analysis distinguishes tissue repair biomarker signatures in wound exudates obtained from normal healing and chronic wounds. J Proteome Res 2010, 9:4758–66. 10.1021/pr100456d
Tilleman K, Van Beneden K, Dhondt A, Hoffman I, De Keyser F, Veys E, Elewaut D, Defore D: Chronically inflamed synovium from spondyloarthropathy and rheumatoid arthritis investigated by protein expression profiling followed by tandem mass spectrometry. Proteomics 2005, 5:2247–57. 10.1002/pmic.200401109
Eissa A, Cretu D, Soosaipillai A, Thavaneswaran A, Pellett F, Diamandis A, Cevikbas F, Steinhoff M, Diamandis EP, Gladman D, Chandran V: Serum kallikrein-8 correlates with skin activity, but not psoriatic arthritis, in patients with psoriatic disease. Clin Chem Lab Med 2013, 51:317–25.
Martínez-Morillo E, Cho C-KJ, Drabovich AP, Shaw JL, Soosaipillai A, Diamandis EP: Development of a multiplex selected reaction monitoring assay for quantification of biochemical markers of down syndrome in amniotic fluid samples. J Proteome Res 2010, 11:3880–7.
Weaver CT, Hatton RD, Mangan PR, Harrington LE: IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu Rev Immunol 2007, 25:821–52. 10.1146/annurev.immunol.25.022106.141557
Mangan PR, Harrington LE, O'Quinn DB, Helms WS, Bullard DC, Elson CO, Hatton RD, Wahl SM, Schoeb TR, Weaver CT: Transforming growth factor-β induces development of the TH17 lineage. Nature 2006, 441:231–4. 10.1038/nature04754
Hamilton DW: Functional role of periostin in development and wound repair: implications for connective tissue disease. J Cell Commun Signal 2008, 2:9–17. 10.1007/s12079-008-0023-5
Ruan K, Bao S, Ouyang G: The multifaceted role of periostin in tumorigenesis. Cell Mol Life Sci 2009, 66:2219–30. 10.1007/s00018-009-0013-7
Masuoka M, Shiraishi H, Ohta S, Suzuki S, Arima K, Aoki S, Toda S, Inagaki N, Kurihara Y, Hayashida S, Takeuchi S, Koike K, Ono I, Noshiro H, Furue M, Conway SJ, Narisawa Y, Izuhara K: Periostin promotes chronic allergic inflammation in response to Th2 cytokines. J Clin Invest 2012, 122:2590–600. 10.1172/JCI58978
Blanchard C, Mingler MK, McBride M, Putnam PE, Collins MH, Chang G, Stringer K, Abonia JP, Molkentin JD, Rothenberg ME: Periostin facilitates eosinophil tissue infiltration in allergic lung and esophageal responses. Mucosal Immunol 2008, 1:289–96. 10.1038/mi.2008.15
Lin J, Zhou Z, Huo R, Xiao L, Ouyang G, Wang L, Sun Y, Shen B, Li D, Li N: Cyr61 induces IL-6 production by fibroblast-like synoviocytes promoting Th17 differentiation in rheumatoid arthritis. J Immunol 2012, 188:5776–84. 10.4049/jimmunol.1103201
Cretu D, Prassas I, Saraon P, Batruch I, Gandhi R, Diamandis EP, Chandran V: Identification of psoriatic arthritis mediators in synovial fluid by quantitative mass spectrometry. Clin Proteomics 2014, 11:27. 10.1186/1559-0275-11-27
Taylor W, Gladman D, Helliwell P, Marchesoni A, Mease P, Mielants H: Classification criteria for psoriatic arthritis: development of new criteria from a large international study. Arthritis Rheum 2006, 54:2665–73. 10.1002/art.21972
Benjamini Y, Hochberg Y: Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc Series B Stat Methodol 1995, 57:289–300.
Wu C, Orozco C, Boyer J, Leglise M, Goodale J, Batalov S, Hodge CL, Haase J, Janes J, Huss JW 3rd, Su A: BioGPS: an extensible and customizable portal for querying and organizing gene annotation resources. Genome Biol 2009, 10:R130. 10.1186/gb-2009-10-11-r130
Berglund L, Björling E, Oksvold P, Fagerberg L, Asplund A, Szigyarto CA, Persson A, Ottosson J, Wernerus H, Nilsson P, Lundberg E, Sivertsson A, Navani S, Wester K, Kampf C, Hober S, Ponten F, Uhlen M: A genecentric human protein Atlas for expression profiles based on antibodies. Mol Cell Proteomics 2008, 7:2019–27. 10.1074/mcp.R800013-MCP200
Muthusamy B, Hanumanthu G, Suresh S, Rekha B, Srinivas D, Karthick L, Vrushabendra BM, Sharma S, Mishra G, Chatterjee P, Mangala KS, Shivashankar HN, Chandrika KN, Deshpande N, Suresh M, Kannabiran N, Niranjan V, Nalli A, Prasad TS, Arun KS, Reddy R, Chandran S, Jadhav T, Julie D, Mahesh M, John SL, Palvankar K, Sudhir D, Bala P, Rashmi NS, Vishnupriya G, Dhar K, Reshma S, Chaerkady R, Gandhi TK, Harsha HC, Mohan SS, Deshpande KS, Sarker M, Pandey A: Plasma proteome database as a resource for proteomics research. Proteomics 2005, 13:3531–6.
Acknowledgments
We would like to thank Dr. Dafna Gladman for her support and helpful suggestions, and Daniel Pereira and Fatima Abji for their ongoing assistance in sample collection.
Funding sources
This work was supported by grants from the Canadian Institutes of Health Research (CIHR) [Funding Reference Number 115719], Abbvie Canada, The Krembil Foundation, and the Laboratory Medicine and Pathobiology program at University of Toronto.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
DC, VC, and EPD participated in the conception and design of the study. KL performed statistical analysis. VC provided PsA and PsC samples. DC, PS, and IB performed proteomic analysis, and developed SRM assays. DC and IB analyzed data and interpreted proteomic results. DC developed and performed ELISAs, and interpreted data. DC drafted the manuscript. DC, KL, EPD and VC prepared the final version of the manuscript. All authors read and approved the final manuscript.
Electronic supplementary material
12014_2014_86_MOESM1_ESM.xlsx
Additional file 1: Table S1: RAW data listing 1922 proteins identified by LC-MS/MS, in lesional and non-lesional PsA and PsC skin. (XLSX 450 KB)
12014_2014_86_MOESM2_ESM.xlsx
Additional file 2: Table S2: Unfiltered list of elevated proteins in lesional and non-lesional PsA skin, as identified by LC-MS/MS. The red font represents data and calculations ycorresponding to PsA lesional skin, while the blue font indicates the PsA non-lesional skin data points and calculations. Proteins commonly elevated in the lesional and non-lesional PsA groups are depicted in the last column. (XLSX 44 KB)
12014_2014_86_MOESM3_ESM.xlsx
Additional file 3: Table S3: Fold change (FC) of 47 elevated proteins in PsA skin compared to PsC skin, as identified by LC-MS/MS. *FC represents the ratio of the mean XIC values of 10 skin samples per group (PsA L vs. PsC L); **N/A indicates that a ratio could not be compiled since the protein was absent in PsC skin; ***P-Values were calculated using the student's t-tests; ****FDR represents the false discovery rate of each protein. (XLSX 11 KB)
12014_2014_86_MOESM4_ESM.xlsx
Additional file 4: Table S4: List of 47 filtered and 2 housekeeping proteins, and the corresponding peptide sequences and transitions that were monitored in the multiplexed SRM assay. The sequence and transitions of the spiked-in heavy peptide are also depicted in the last three rows. (XLSX 13 KB)
12014_2014_86_MOESM5_ESM.pdf
Additional file 5: Figure S1: Distribution of markers across the PsA and PsC skin Set I. Dots represent skin samples from individual subjects; thin horizontal lines depict the mean, and vertical lines the SD. **** indicates P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns:non-significant. (PDF 122 KB)
12014_2014_86_MOESM6_ESM.pdf
Additional file 6: Figure S2: Distribution of markers across the PsA and PsC skin Set II. Dots represent skin samples from individual subjects; thin horizontal lines depict the mean, and vertical lines the SD. **** indicates P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns:non-significant. (PDF 70 KB)
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Cretu, D., Liang, K., Saraon, P. et al. Quantitative tandem mass-spectrometry of skin tissue reveals putative psoriatic arthritis biomarkers. Clin Proteom 12, 1 (2015). https://doi.org/10.1186/1559-0275-12-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1559-0275-12-1