
Abstract
Clinical utilization of ovulation stimulation to facilitate the ability of a couple to conceive has not only provided a valuable therapeutic approach, but has also yielded extensive information on the physiology of ovarian follicular recruitment, endometrial receptivity and early embryo competency. One of the consequences of the use of fertility enhancing agents for ovarian stimulation has been the creation of a hyperestrogenic state, which may influence each of these parameters. Use of aromatase inhibitors reduces hyperestrogenism inevitably attained during ovarian stimulation. In addition, the adjunct use of aromatase inhibitors during ovarian stimulation reduces amount of gonadotropins required for optimum stimulation. The unique approach of reducing hyperestrogenism, as well as lowering amount of gonadotropins without affecting the number of mature ovarian follicles is an exciting strategy that could result in improvement in the treatment outcome by ameliorating the deleterious effects of the ovarian stimulation on follicular development, endometrial receptivity, as well as oocyte and embryo quality.
Similar content being viewed by others

PART ONE
1 Introduction
Current epidemiological evidence suggests that 15% of couples will experience infertility. Background prevalence rates now appear to be reasonably stable, but there is evidence of an increase in the rate of referrals for medical help [1, 2]. Farley and Belsey, 1988 [3], have reported estimates of the prevalence (percentage) of primary infertility by region and country. They estimated 6% for North America, 5.4% for Europe, 3% for the Middle East, 10.1% for Africa, 4.8% for Asia and Oceania, 3.1% for Latin America and 6.5% for the Caribbean. The American Society for Reproductive Medicine (ASRM) estimates that 5 million American heterosexual couples report difficulties in achieving a viable pregnancy, of which 1.3 million seek advice for the problem [4].
2 Ovarian stimulation and assisted reproduction for infertility management
After correcting the abnormalities detected during the diagnostic workup, ovulation induction is usually performed either for treatment of anovulation/oligo-ovulation, or empirically in regularly ovulating women. This approach results in a pregnancy rate of around 8%–15% per cycle depending on the agents used for ovulation induction and the characteristics of the couple, such as the woman's age and the presence or absence of a male factor. Couples who do not become pregnant with ovulation induction alone then undergo more sophisticated treatment modalities including intrauterine insemination (IUI) and in-vitro fertilization and embryo transfer (IVF-ET) as a treatment of last resort [5].
Since the birth of Louise Brown in 1978, IVF-ET has become the therapeutic mainstay for female infertility. It has become generally accepted as therapy for a wide array of fertility problems, and has been accompanied by the rapid expansion of IVF-ET clinics worldwide resulting in more than 1% of babies being conceived by IVF-ET in western countries [6].
2.1 Ovarian stimulation for assisted reproduction
In most assisted reproduction programs, gonadotropins are used alone or in combination to stimulate the growth and maturation of multiple follicles. This is essential because of the need to recruit a greater number of follicles, which provides the opportunity for retrieval of a large number of oocytes. This would improve the chance for fertilization of multiple oocytes and thereby allow an increased number of embryos for transfer in order to give acceptable success rates. Recent advances in the understanding of ovarian stimulation, the techniques of oocyte retrieval, the handling of gametes, the methods of assisted fertilization and improved conditions of culture media have steadily increased the fertilization rate. Fertilization rates of 60–70% can now be expected when conventional insemination, or even higher when intracytoplasmic sperm injection (ICSI) are carried out. However, there has not been a corresponding increase in implantation rates, which have remained steady at overall rates around 10%–15% [6].
2.2 Low implantation rates with assisted reproduction
Throughout the last five decades, a progressive series of revolutionary techniques have been developed to overcome infertility, starting with the successful fertilization of human oocytes in vitro [7] and followed nearly 10 years later by the birth of the first IVF-ET baby [8]. Several other new developments in assisted reproduction have emerged, including cryopreservation and storage of embryos for later transfer [9], fertilization of oocytes with a single injected spermatozoon to alleviate severe male infertility i.e. ICSI [10] and diagnosis of genetic defects from preimplantation embryos prior to intrauterine transfer [11]. However, although IVF-ET is now a standard, well-established treatment for infertility, success rates remain relatively low, with only about 33% of cycles resulting in pregnancy [12]. This is believed to be due to the low implantation rate that has not significantly increased as fertilization rates [13]. Efforts are being made to improve implantation rates after IVF-ET by improving culture conditions, optimizing gamete quality and developing new techniques of selecting viable embryos for transfer without significant success. For this reason, multiple embryos are generally transferred to improve pregnancy rates, but this has resulted in an unacceptably high rate of multiple-gestation pregnancies [14].
Although governed by multiple interactive events, embryo implantation depends mainly on the quality of embryos and the status of uterine receptivity. During the last two decades, several developments in controlled ovarian hyperstimulation [COH], fertilization, and embryo culture techniques have led to an optimization in the number and quality of embryos available for ET. In contrast, uterine receptivity has failed to benefit from parallel improvements, and its disarrangement is likely to represent an important cause of the sub-optimal embryo implantation rates observed in IVF-ET [15].
2.3 Poor outcome of infertility treatment associated with ovarian stimulation
In the following section we review in brief both animal and human evidence for the unfavorable outcome including impaired implantation and increased adverse outcomes in pregnancies achieved following ovarian stimulation when compared with spontaneous pregnancies.
2.3.1 Animal data
Increased pre- and post-implantation embryonic loss has been reported in mammals [16–21] including rats [16, 18], mice [17–19], murine [20] and hamsters [21], in association with ovarian hyperstimulation. These effects have been attributed to ovarian stimulation using standard doses of gonadotropins. At higher doses of gonadotropins, studies have found increased frequencies of oocyte aneuploidy, embryo mortality, fetal growth retardation and congenital abnormalities [22, 23].
The poor outcome after ovarian stimulation has been attributed to adverse effects on the maternal side and or the gametes and embryo side. On the maternal side: inadequate uterine synchrony or receptivity has been reported. On the gamete and embryo side, ovarian stimulation has been found associated with chromosomal defects in the oocyte leading to increased lethality during the preimplantation stages [22, 23]. However, because species-specific variations in implantation strategies exist, these differences preclude the formulation of a unifying theme for the molecular basis of this event.
2.3.2 Human data
Many studies have found higher pregnancy rates in donor oocyte recipients than patients undergoing standard IVF-ET [24, 25]. The higher success rates could be attributed to better quality oocytes from younger donors. However, in centers using a shared oocyte system, where the donor keeps half of the oocytes for herself, significantly higher pregnancy and implantation rates were found in the recipients [26].
Another evidence for adverse effects of ovarian stimulation on implantation is the higher implantation rate associated with IVF-ET in the natural or un-stimulated cycle. Although most studies were associated with a rather high proportion of cancelled cycles [25–75%] and a low clinical pregnancy rate per started cycle [range 0–23%], higher implantation rates have been reported (up to 30%) [27].
Adverse Obstetrical Outcome after ovarian stimulation
Induction of ovulation has been shown to raise the risk of miscarriage when compared with spontaneous pregnancies [28]. This was true even after controlling for advanced age, a known significant risk factor for miscarriage. Higher risks as compared with natural pregnancies are reported in pregnancies after IVF-ET, primarily owing to growth retardation and pre-term birth. Although this can be explained by the high multiple pregnancy rates in IVF-ET pregnancies [29], an increased rate of small for gestational age and pre-term birth children is reported in singleton IVF-ET pregnancies as compared with natural singleton pregnancies after adjustment for potential biases [30–32]. Moreover, twin pregnancies after assisted reproduction have a higher rate of perinatal mortality and lower birth weight as result of a higher rate of premature parturition when compared to spontaneously occurring twins [33, 34]. In addition, it was reported that women who conceived multiple gestations through assisted reproductive technologies have a 2.1-fold higher risk of preeclampsia than those who conceive spontaneously [35]. Pregnancies associated with severe ovarian hyperstimulation syndrome have been found to be complicated with increased miscarriage rates [36, 37]
While ovarian stimulation has been suggested to contribute, at least in part, for the adverse obstetric outcome other studies have shown that infertility itself is a factor that leads to increased obstetric risks and that sub-fertility is a predictor for low birth weight. Some believe that it is the cause of infertility itself, rather than the use of ovarian stimulation, that is the reason behind poor obstetric outcome after infertility treatment [38, 39].
3 Postulated mechanisms behind adverse effects of ovarian stimulation
Several causes and targets have been suggested to explain the poor outcome associated with ovarian stimulation (Table 1). These include (1) supraphysiological levels of estrogen, and other steroids and peptides, attained during ovarian hyperstimulation, (2) the use of exogenous gonadotropins as well as other medications applied during ovarian stimulation such as Gonadotropin releasing hormone (GnRH) analogues (agonists and antagonists), human chorionic gonadotropin (hCG) and clomiphene citrate (3) Other possible undetermined factors. These factors are believed to act through their effects on (i) the endometrium, (ii) the developing oocyte, (iii) the developing embryo, (iv) the ovaries and corpus luteum, (v) the pituitary gland, and (vi) possibly on other targets such as fallopian tubes, the coagulation system, as well as the early developing placenta.
3.1 Effect of supraphysiological levels of estrogen
It is believed that the supraphysiological levels of estrogen, attained during ovarian stimulation, may explain, at least in part, for the adverse effects of ovarian stimulation on the outcome of infertility treatment [40–42]. Different mechanisms have been suggested for the deleterious effects of the supraphysiological levels of estrogen attained during ovarian stimulation Table 2.
Considering all the patients together, significant decreases in pregnancy and implantation rates were observed when estradiol (E2) concentrations were > 2500 pg/ml [>9000 pmol/L.] compared with patients having lower E2 concentrations. High serum E2 concentrations on the day of hCG injection in high and normal responder patients, regardless of the number of oocytes retrieved and the serum progesterone concentration were found to be detrimental to uterine receptivity [42]. Later, it has been shown that a significant reduction in the implantation and pregnancy rates occurred in almost all women with a higher serum E2 concentration of ~5–6000 pg/ml [19–22,000 pmol/L] [43, 44]. Recently, we have presented data showing that high E2 levels are associated with less favorable treatment outcome in women undergoing controlled ovarian hyperstimulation and IVF-ET [45–51]. We studied the effect of E2 by looking at the area under the curve (AUC) for E2 levels along the stimulation cycle [45–51]. We believe this is more accurate than looking at a single measurement of E2 e.g. one day of hCG administration [46]. In addition to studying the AUC for E2 levels [45, 46], we looked at the E2 production per mature follicle and per gonadotropin dose administered during COH [48] and the effect of age on these parameters [47]. We found significant correlation between these parameters and the outcome of IVF-ET treatment showing that high E2 levels are associated with lower clinical pregnancy and implantation rates [45–49] in addition to increased adverse obstetric outcomes including higher miscarriage rate [49] and lower birth weight [50]. Successful IVF-ET treatment cycles were associated with lower AUC-E2 compared to unsuccessful cycles at the same patients [49].
3.1.1 Effect of supraphysiological estrogen levels on the endometrium
One of the causes for the reduced implantation rates associated with ovarian stimulation may be an impairment of endometrial receptivity, due to high concentrations of sex steroids. This suggestion is supported by higher implantation rates in hormonal replacement treatment cycles after ovum donation, as opposed to the standard IVF-ET cycles as explained earlier.
Effect on Implantation Window
It is generally believed that the embryo-uterine interactions leading to implantation can only succeed when embryonic development is synchronized with the preparation of the endometrium to the receptive state. Typically, this means that the embryos have reached the blastocyst stage and that the endometrium has undergone certain hormone-dependent changes during a specific time window in the preimplantation phase that prepare it to be receptive to the developing blastocyst [52].
The concept of endometrial receptivity introduced by Psychoyos [53, 54], has been shown to last only for several hours, thereby determining a narrow nidation window. The concept of an "implantation window" or "receptive endometrium" was initially established in rodents. In the rat, the fertilized embryo reaches the uterus on day 4 after fertilization, and implantation occurs in the afternoon of day 5 [53, 54]. In humans, the ovum is fertilized in the fallopian tube, arrives in the uterine cavity around day 17 (day 14 is taken as day of ovulation of a 28-day cycle), and remains there as a free-floating embryo until about day 19; implantation then occurs between days 19 to 22 [55, 56]. However, the precise timing and molecular basis of the receptive window in the human remain undefined. Unfortunately, endometrial receptivity knowledge in the human is limited due to the obvious experimental drawbacks and the lack of specific criteria to define a receptive endometrium.
In the literature, there is controversy regarding the effect of ovarian stimulation on endometrial development. Most of the investigators have reported adverse effects of high estrogen levels on endometrial development but there was no consensus on the actual effect. Some have shown endometrial advancement [57–61] while others showed endometrial retardation [62, 63]. However, all studies confirm direct deleterious effects on the endometrial development that jeopardize the chance of implantation due to the lack of synchronization between the development of the endometrium and the early embryo development. Implantation failure has been suggested to result from the disparity in maturation between the endometrial stroma and the epithelium observed in histology. Since a paracrine communication between the epithelium and the stroma may be important at the beginning of implantation, this disparity could compromise uterine receptivity or early trophoblastic invasion [64, 65].
The effect of high E2 concentrations (> 20,000 pmol/L) was found to be associated with gland-stromal dys-synchrony, which indicated a deficient secretory transformation of the endometrium that represents a sub-optimal endometrial environment for implantation. This finding substantiates clinical observation of significantly lower pregnancy rates in IVF-ET cycles of women with high E2 concentrations. In these patients, there was a marked stromal edema associated with a significantly greater number of stromal vessels, which suggested advanced stromal maturation [44].
In these studies, definitions of supraphysiological hormonal concentrations were variable. Also, the timing of the biopsies and the drug regimens used for ovarian stimulation were different. This, in addition to the variation in endometrial response to different E2 concentrations, may explain the disagreements reported in the literature [66]. Moreover, both the premature progesterone serum elevation, which occurs in 30% of stimulated IVF-ET cycles before hCG administration, and the advanced post-ovulatory rise in progesterone serum concentrations are believed to be responsible for the advanced endometrial development associated with ovarian stimulation [67, 68]. However, other studies failed to confirm these observations [69, 70].
Effect on endometrial pinopodes
At the time of implantation, the apical membranes of the epithelial cells lining the uterine cavity develop large and smooth membrane projections, named pinocytes due to their pinocytotic function [71]. Their development is progesterone-dependent with strict correlation with the implantation window in the rodents [72]. Similar structures have been seen in the human endometrium [73]. The number of pinopodes was found to have a strong correlation with implantation after embryo transfer [74, 75].
Hormonal treatment has been shown to be associated with changes in the timing of pinopode formation. During ovarian stimulation with clomiphene citrate followed by human menopausal gonadotropins (hMG)/hCG, fully developed pinopodes were found 2 to 3 days earlier [76]. In contrast, with E2 and progesterone treatment, fully developed pinopodes were found to be about two days later. Pinopodes were found to form as early as 4 days after hCG administration [59].
Nikas et al. [61], studied the temporal expression of pinopodes as a specific marker for receptivity in IVF-ET cycles induced by gonadotropins, compared to women with regular menstrual periods and proven fertility who served as controls. They did not find ovarian stimulation to affect endometrial pinopode formation in terms of quantity and life span. Instead, the cycle days when pinopodes formed were specific to the individual, being on average 1–2 days earlier in cycles with ovarian stimulation than in natural cycles. These changes in pinopode expression may reflect shifts in the window of receptivity, resulting in ovo-endometrial asynchrony and limiting implantation success in IVF-ET [61]. In IVF-ET, embryonic development is probably delayed while the uterus is advanced, resulting in an early closure of the nidation window, before the embryo eventually reaches a stage capable of initiating implantation. These findings support the theory that reduced implantation rates in IVF-ET cycles could result from impaired or premature endometrial maturation.
Effect on estrogen and progesterone receptors in the endometrium
A lack of estrogen receptors (ER) has been reported [77] during ovarian stimulation cycles that rendered the endometrium functionally hypoestrogenic or hypoprogestogenic. Also, it has been reported that the expression of progesterone receptors (PR) in the endometrium was decreased in the early part of luteal phase after ovarian stimulation [78]. This premature PR decrease was consistent with an early high progesterone level [79]. Papanikolaou et al. [80], investigated prospectively the effect of multi-follicular ovarian stimulation for IVF on the late follicular phase endometrium histology and the expression of ER and PR. Endometrial biopsies were taken in a natural cycle on the day of the onset of the surge of the LH, and in a subsequent stimulation cycle on the day of hCG administration for final oocyte maturation. Histological examination of biopsies both in natural and stimulated cycles showed no secretory changes. However, in stimulated cycles, PR expression was significantly up-regulated compared to natural cycles in both glands (1.67 versus 1.34, P < 0.05) and stroma (1.98 versus 1.62, P < 0.05), whereas ER was down-regulated in glands (1.15 versus 1.43, P < 0.05). In IVF cycles, the progesterone measurements, although within normal values (range 0.8–1.4 microg/l), were significantly higher than in natural cycles (0.99 vs 0.63 microg/l, respectively, P = 0.008). An ongoing pregnancy rate of 37.5% was achieved in the stimulated cycles. The authors concluded that although the current study found no early secretory transformation in stimulated endometria before hCG administration, the ER and PR expression in these endometria was similar to the one described during the first days of the luteal phase in natural cycles. Supraphysiological concentrations of estradiol and subtle progesterone rises in the late follicular phase might be responsible for this modulated steroid receptor profile. They added that this phenomenon indicated accentuated maturation of the endometrium in IVF cycles from the pre-ovulatory phase onwards. [80].
Ovarian stimulation with GnRH-agonist/hMG was found to induce precocious secretory endometrial transformation around the time of oocyte retrieval. Compared to natural cycles, there was an imbalance between endometrial steroid receptor content, proliferation index, and maturation in the peri- and postovulatory phases of stimulated cycles. The lower ER in stimulated cycles on the day of oocyte pick up as compared to the natural cycle controls on the day of ovulation was mainly observed in the stroma, but failed to reach a significant difference in the glands. As for PR, the staining intensity of the stromal ER in the stimulated cycles was higher than that of luteal phase day 2 of a natural cycle. These findings suggest a relative imbalance in ER and PR content of the endometrium in stimulated cycles compared to their natural cycle counterparts. [79].
In conclusion, excessive ovarian response was suggested to lead to insufficient secretory transformation of the endometrium, as well as discordant glandular and stromal development at a time that coincides with the period of maximum uterine receptivity.
Effect on endometrial vasculature and endometrial blood flow
Angiogenesis has a critical role in female reproductive physiology. Growth of the endometrium and placentation is also accompanied by extensive angiogenesis. Thus, an actively maintained blood supply is an essential requirement for reproductive functions, including normal implantation [81]. Endometrial vasculature has been shown to play a prominent role in the early endometrial response to the implanting blastocyst, and vascular changes may contribute to uterine receptivity [82].
The introduction of transvaginal Doppler ultrasound makes the measurement of uterine artery blood flow possible, and at one time it was hoped that uterine arterial resistance changes might reflect uterine receptivity [83]. Applebaum first introduced the concept of evaluating uterine receptivity by a uterine score including the endometrial blood flow [84]. Different studies have demonstrated significant changes in the Doppler indices of uterine and ovarian vessels during ovarian stimulation and spontaneous cycles [85, 86]. Basir et al., found evidence of impaired endometrial blood flow in association with significantly high estrogen levels in high responders to ovarian stimulation [87].
Although pregnancy outcome tended to be poor in patients with higher mean uterine arterial impedance indices, the predictive value of using a specific resistance index (RI) or pulsatility index [PI] variable in assessing endometrial receptivity seems to be limited [88]. One of the explanations is that the major uterine compartment is the myometrium and not the endometrium, and thus most of the blood passing through the uterine arteries never reaches the endometrium. A more logical approach would be to evaluate the vascularization around the endometrium directly in an attempt to assess endometrial receptivity.
Histological studies have confirmed that the sub-endometrial halo surrounding the endometrium represents the innermost layer of the myometrium, and compared with the outer myometrium, it consists of a distinct compartment of more tightly packed muscle cells with increased vascularity. Studies have shown that interactions between the junctional zone and the endometrium may play an important role in the implantation process [89, 90] and endometrial-sub-endometrial blood flow distribution pattern assessed by transvaginal color Doppler before ET was found to correlate with the implantation and pregnancy rate after IVF-ET [91]. With the absence of sub-endometrial blood flow, even in the presence of other favorable parameters, no conception was achieved. By using a similar approach, Salle et al., calculated a uterine score in the secretory phase of the menstrual cycle preceding IVF-ET [92]. Immunocytochemistry study revealed that the sub-endometrial myometrium, also called the junctional zone myometrium or archimyometrium, exhibits a cyclic pattern of estrogen and PR expression that parallels that of the endometrium [93]. Moreover, the responsiveness of the junctional zone has been shown to be associated with implantation success during IVF-ET treatment [90].
Many investigators have also noted the correlation of junctional zone contractions with pregnancy outcome in both natural [94] and assisted reproduction cycles [95]. Less-active junctional zone contractility is associated with higher pregnancy rates
Kupesic et al. compared the 2-D and 3-D ultrasonographic scoring systems by combining parameters including endometrial thickness, volume, echogenicity, and sub-endometrial blood flow. They found the two systems had similar efficiencies in predicting pregnancy outcome of IVF-ET procedures [96]. Several investigators noticed that when the endometrial and sub-endometrial flow parameters were combined, significant differences were found between pregnant and nonpregnant patients [97–99]. In contrast, there was no significant difference if attention was only focused on intraendometrial or sub-endometrial blood flow [100]. These results imply that the endometrial/sub-endometrial area must be considered as a whole in evaluating endometrial perfusion.
One hallmark of implantation is increased vascular permeability at the implantation site. Vasoactive agents, including histamine, platelet-activating factor, vascular endothelial growth factor, and eicosanoids, have been studied during implantation [101, 102]. Vaginal E2 administration improves endometrial proliferation and uterine perfusion, presumably because of combined local and systemic effects, but may interfere with P-induced uterine relaxation [103].
In a study by Basir et al., [104] the investigators compared the hemodynamic parameters of the utero-ovarian vasculature and the endometrial spiral arteries of women who showed a moderate response with women whose E2 concentrations were in excess of 20,000 pmol/l after ovarian stimulation. Despite low uterine PI and RI, the endometrial blood flow in high responders appears to be impaired. The authors concluded that this might contribute to the decline in implantation efficiency noted in high responders. The decreased endometrial blood flow despite the increased blood flow in the uterine arteries may indicate a shunt of blood flow from the endometrium into the myometrium.
The authors [104] found low pulsatility index and resistance index of the ovarian arteries indicating neovascularization and increased capillary permeability in the ovarian tissue of high responders. The authors suggested that the blood flow might be directed through the utero-ovarian collaterals to the ovaries. However, because the sample size in this study was small (19 patients with E2 > 20,000 pmol/L), further larger prospective studies are required to confirm the effect of excessively high concentrations of serum E2 on endometrial blood flow.
Moreover, the increase in hormonal concentrations in the peripheral plasma leads to a decrease in peripheral vascular resistance [105] and a decreased contractility of the uterine muscles. This results in relaxation and opening up of the small uterine vascular channels, which may also cause an increase in the capillary permeability. In a study on the endometrial morphological changes at high concentrations of E2, a significantly greater number of vessels and endometrial edema in women who responded excessively to ovarian stimulation was demonstrated. Therefore, it was postulated that the blood flow through these minute endometrial vessels may be very slow and the weak Doppler flow signals arising from them could not be picked up by the color Doppler despite low uterine PI and RI. The increase in capillary permeability and dilatation leads to extravasation of fluid from the intercellular to extracellular compartments, and hence endometrial edema [106]. In another study the investigators suggested that the blood flow per capillary might actually be reduced during edema [107].
Successful implantation and continuing development of implanted embryo depends on a complex series of cellular and molecular events between the blastocyst and the endometrium [108]. The decline in blood flow could therefore impede the exchange of essential nutrients, bioactive molecules and reactive compounds that are vital for implantation with absent endometrial and intra-endometrial vascularization appeared to be a useful predictor of failure of implantation in IVF-ET cycles [109].
Whether fertilization occurs in vivo or in vitro, most human embryos will not develop through gestation [110] with a very high proportion of developmental failure during the preimplantation stages associated with chromosomal defects of oocyte origin [111]. Van Blerkom suggested that the dissolved oxygen content of pre-ovulatory follicular fluid and the developmental competence of the corresponding oocyte were related [112]. A developmentally significant association between the chromosomal normality of the human oocyte and the level of intra-follicular oxygen and peri-follicular vascularity was reported suggesting that hypoxic intra-follicular conditions that result from the failure of an appropriate microvasculature to develop around the growing or pre-ovulatory follicle(s) could be a proximate cause of the maternal-age-related increase in the incidence of trisomic conditions [112–114].
Effect on Integrins
Integrins are a family of cell adhesion molecules. Previous work has shown the expression of integrins in the endometrium changes during the menstrual cycle [115]. Three integrins in particular (1β1, 4β1 and vβ3) are thought to play a vital role in implantation as all are expressed during the 'implantation window'. The β3 and β1 integrins have also been shown to be reduced in infertile patients using flow cytometry [116]. Aberrant patterns of integrin expression have also been associated with certain diagnoses in infertile patients, including luteal phase defects, endometriosis, hydrosalpinx and unexplained infertility [117].
The exact role of integrins remains controversial and results have not been duplicated in all studies. Creus and co-workers showed no difference in integrin expression in patients who became spontaneously pregnant compared with those that do not [118]. Thomas et al., demonstrated that integrin expression seems to be reduced in the glandular epithelium in the endometrium after ovulation induction, irrespective of the dating. The authors concluded that there might be an ideal E2 level that should be reached during IVF-ET treatment as low estrogen levels might reduce the yield of oocytes, but high levels might impair the receptivity of the endometrium reducing integrin expression and leading to lower implantation rates [119].
3.1.2 Effect of supraphysiological estrogen levels on the developing oocyte (follicular and oocyte development)
As the contributor of the bulk of the cytoplasm to the zygote and half of its nuclear DNA, the key importance of the oocyte is unquestionable. The principle, that embryogenesis is rooted in oogenesis, has been understood for nearly a century, and is now becoming better understood at the molecular level. During follicular growth, the oocyte is in close contact with granulosa cells through gap junctions and is therefore under the influence of the follicular environment. Since oocyte maturation is such a long process, any adverse events occurring during this period can damage vital structural molecules or whole organelles, resulting in oocytes with reduced developmental competence. These oocytes may in turn give rise to preimplantation embryos with a compromised viability, and hence, a reduced implantation potential [120]. The ER gene is expressed in the human cumulus-oocyte complexes and oocytes but not in granulosa/cumulus cells which might suggest a lack of receptor-mediated autocrine effect of estrogen during folliculogenesis. Conversely, estrogen secreted by granulosa/cumulus cells, may exert a paracrine effect to influence oocyte maturation and fertilization competence directly [121]. It was reported that retrieval of >10 oocytes during IVF-ET cycles was correlated with oocytes of lower quality, as manifested by a decrease in the fertilization rate [122].
Effect on the chromosomal structure and cytogenetics of the oocytes
Studies with animal systems have indicated that a single cycle of ovarian stimulation can have adverse effects on oocyte competence during early development, and may well have downstream effects on the normality of fetal growth and development. Whether developmental defects could result from chromosomal malsegregation during ovarian stimulation-induced meiotic maturation has been examined in the mouse system. Earlier studies compared oocytes obtained after spontaneous or hormonally induced ovulation and found no increase in the incidence of non-disjunction or oocyte aneuploidy after ovarian stimulation [123]. In contrast, cytogenetic analysis of pronuclear stage mouse eggs after single cycle of ovarian stimulation showed chromosomal aberrations largely confined to the female pronucleus, indicating developmental compromise prior to fertilization [124]. Sengoku and Dukelow found comparable frequencies of aneuploidy for cleavage-stage hamster embryos produced in natural and pregnant mare serum gonadotropins (PMSG)-stimulated cycles, indicating that peri-implantation mortality may have an epigenetic origin [125].
In IVF-ET programs, the high rate of oocyte recovery and successful fertilization in vitro contrasts with a relatively high rate of conception failures, as even the most advanced IVF-ET clinics can provide a 33% success rate per cycle at best [12], despite multiple embryo transfers. The possible reason for this conception failure is the high frequency of lethal chromosomal abnormalities that prevents embryonic development beyond the pre- and post-implantation stages. The high frequency of spontaneous abortions also indicates that the proportion of embryos with genetic defects is significant. Cytogenetic analysis of the early stages of cleavage indicates that chromosomal aberrations may be found in 35–44% of pre-implantation embryos produced in vitro [126]. It has to be emphasized that the majority of pre-implantation embryos carry numerical chromosomal defects. However, the high incidence of chromosomal anomalies is presumably biased by the fact that only embryos with a poor morphological score were analyzed (discarded embryos). It nevertheless indicates that the low implantation rates of human pre-implantation embryos in IVF-ET programs are the consequence of natural selection [127]. Aneuploidy is the most common abnormality found in normally developing embryos following ovarian stimulation and IVF-ET [128], but other aberrations were also revealed. Polyploidy and multinucleation were frequently described in arrested embryos
Estrogen is well known to induce chromosomal and cytogenetic damage. The natural hormone E2 has clearly been shown to induce alterations in chromosome number such as losses or gains of whole chromosomes [129, 130], chromosome translocations [131, 132], and gene amplifications [133, 134]. In addition, there is preliminary evidence of estrogen-induced gene mutations and gene deletions [135, 136].
The synthetic estrogen, diethylstilbestrol (DES) causes a severe yet reversible deterioration of meiotic spindle microtubule organization during maturation of the mouse oocytes [137]. High doses of E2 were found to induce numerical chromosome changes (both chromosome gains and losses) similar to the reported observations with the synthetic estrogen, diethylstilbestrol [129]. Estrogen is also known to cause direct DNA damage via its catecholestrogen metabolites [138, 139]. There are several types of free radical-mediated DNA damage, which are induced by estrogens and/or their metabolites [140–142].
As explained earlier, supraphysiological estrogen levels associated with ovarian stimulation impair the uterovarian blood flow resulting in follicular hypoxia. Gaulden proposed that follicular hypoxia might have a potent adverse influence on spindle organization and the normality of chromosomal segregation in the human oocyte [143].
Effect on the mitochondrial function of the oocytes and embryos
Van Blerkom has suggested that the developmental competence of mouse and human early embryos is related to the metabolic capacity of the mitochondria. It is thought that mitochondrial replication does not begin until after implantation, and that paternal contribution is minimal. Therefore the preimplantation embryo is completely reliant on maternally inherited mitochondria in the oocyte. Deletions and mutations in oocyte mitochondrial DNA may lead to mitochondrial dysfunction, influencing energy production and apoptosis in oocytes and early embryos, resulting in aberrant chromosomal segregation or developmental arrest [144].
The classical model of E2 action has been described to be mediated by cytoplasmic/nuclear partitioning receptor proteins that stimulate gene transcription upon binding to specific DNA sequences [145]. However, there are increasing functional evidences for extra nuclear/cytoplasmic localization of steroid hormone receptors. Several studies showing rapid non-genomic actions of steroids have led to speculate about the existence of cell-surface resident receptor forms [146–148]. Reports have documented the presence of estrogen binding proteins localized at the plasma membrane [149, 150]. Independently, the known direct effects of various steroids on mitochondrial gene transcription support the idea of receptor attachment to the mitochondrial genome [151]. It was also identified early that estrogen specific binding sites were associated with mitochondrial and microsomal structures [152].
The mitochondrial-enriched subfraction represented an important source of E2 binding, where the steroid was recognized in a stereospecific and high affinity manner. The existence of mitochondrial and membrane estrogen binding sites correlated with the presence of ER but mainly with ER proteins. Using macromolecular E2 derivatives in Ligand Blot studies, both mitochondrial and membrane estrogen binding proteins were found in the uterus, and the ovary. This differential cellular partitioning of ER and forms may contribute to the known diversity of E2 effects in target organs [153].
Recently, in myocardial cell model, it was reported that at physiological concentrations, which do not inhibit mitochondrial functions, estrogens can protect heart mitochondria from the loss of cytochrome c induced by high calcium, and this might be one of the possible mechanisms by which estrogens preserve myocardial cell viability after ischemia/reperfusion [154]. High concentrations of estrogens (50–100 M) have been found to have a damaging effect on mitochondrial functions by strongly inhibiting mitochondrial respiration and membrane potential presumably due to decreased activity of the respiratory chain. The inhibition of the respiratory chain may be due to non-specific binding of estrogens to hydrophobic regions of the mitochondrial membranes, which may change protein/lipid interactions, disturb electron transport through the inner mitochondrial membrane and reduce membrane potential [155].
We postulate that supraphysiological levels of estrogen attained during ovarian stimulation may affect the mitochondrial function of the developing oocyte and embryo. This could be one of the mechanisms behind impaired developmental capacity of the oocytes and embryos obtained after ovarian stimulation.
3.1.3 Effect of supraphysiological estrogen levels on the spermatozoa and sperm/oocyte interaction
Sperm are exposed to estrogens within the male tract, and P450 aromatase has also been identified in human spermatozoa [156]. This raises the possibility that the spermatozoa can provide a continuing local source of estrogens in the epididymis, as well as while in the female tract on its journey to fertilize the oocyte. Inside the female genital tract, the spermatozoa would also be exposed to estrogens, particularly in tubal fluid following follicle rupture and when in close vicinity to released oocytes [157].
Estrogen receptors in the spermatozoa
Non-genomic effects of estrogens have also been reported in several cell types including the spermatozoa [158]. Immunohistochemical detection of ER has been reported for human spermatozoa, with distribution in both the head and flagellum [159].
Possible effect of estrogen on the spermatozoa function
After ejaculation, in vitro, the spermatozoa are initially unable to fertilize [160]. The spermatozoa acquire the capacity to fertilize, after a certain period of time that is species-specific when exposed to an appropriate environment either in vivo or in vitro. This process is called 'sperm capacitation' [161]. Capacitated spermatozoa are able to express hyperactivated motility, to undergo the acrosome reaction, and to fertilize an oocyte [160]. Estrogens are believed to have a possible effect on these events (capacitation and acrosome reaction), which would have important consequences on fertility in vivo.
Recently, Adeoya-Osiguwa provided evidence that E2 and environmental estrogens can significantly stimulate mammalian sperm capacitation, acrosome reactions. They found that in uncapacitated cells, E2 at 00.001 μmol/l, significantly stimulated capacitation and acrosome reactions while in capacitated cells, E2 had no effect. The authors concluded that whether these responses have effects on fertility in vivo remains to be determined, along with the mechanisms of action involved [157]. It is pertinent to mention here that the average E2 concentrations in follicular fluid from mature oocyte are in the micromolar range [162, 163].
The regulation of capacitation is very important in mammalian fertilization as evidence suggests that once capacitation has been initiated it will usually continue unchecked, frequently resulting in the spermatozoa undergoing spontaneous acrosome reactions and thus becoming non-fertilizing [164].
Adeoya-Osiguwa argued that E2 and the environmental estrogens appear primarily to stimulate the spermatozoa, accelerating the rate of capacitation and then promoting 'over-capacitation' in at least some of the cells, resulting in the acrosome reaction [157]. Since already acrosome-reacted spermatozoa are non-fertilizing [160], similar responses occurring in vivo could reduce the number of potentially fertilizing cells and so have an undesirable effect on fertility. As capacitation and fertilization occur in the female reproductive tract, it is likely that any effects of environmental estrogens on sperm function would be more pronounced in the female, but effects on mature spermatozoa awaiting ejaculation cannot be ruled out.
3.1.4 Effect of supraphysiological estrogen levels on the embryo
Mouse and human embryos, when cultured in vitro, undergo a delay in development compared with those grown in vivo. This delay can be caused by suboptimal culture conditions, but possible influences of ovarian stimulation cannot be excluded [165]. In order to determine if implantation failure associated with ovarian stimulation was due to abnormalities in the blastocysts or the endometrium, decidualization studies and embryo transfers to pseudopregnant recipients were performed [166, 167]. The uterus of a large proportion of superovulated animals was unable to undergo decidualization in time, whereas embryo transfers to pseudopregnant females resulted in normally developing fetuses, which indicated that hormonally treated oocytes themselves were not affected. Some studies found that the E2 concentrations in the fresh cycle were not related to the success of frozen-thawed embryo transfer cycles indicating that embryo quality seemed unaffected by the high estrogen levels [168, 169]. However, there is increasing evidence suggesting that ovarian stimulation and the associated high estrogen levels are detrimental on the embryo and associated with a decrease in the fertilization rate. When compared with blastocysts derived from naturally cycling mice, blastocysts that developed in vivo in superovulated mice were found to have fewer microvilli on their surface [170], a reduced [35S]-methionine uptake [171], and a lower cell number and mitotic index [172]. A reduced cell number and a two-fold decrease in viability post-transfer of embryos from gonadotropins-stimulated hamster females, was also observed [21]. Furthermore, it has been reported that the proportion of abnormal preimplantation embryos increases after superovulation, and that blastocysts have a smaller trophoblastic outgrowth in vitro [173]. Moreover, in mice as well as in humans, there is evidence for steroids being regulators of gene expression in the embryo and endometrium, and that embryo morphology and rate of development – both of which reflect embryo quality – have a genetic basis. Also, ovulation induction therapy has been found to be associated with an increased rate of mosaicism in the embryos, which fail to implant [52].
It has been proposed that high E2 levels after COH impair endometrial receptivity because oocyte quality, fertilization rate, and embryo cleavage (until day 2) were normal in patients with a high response [174] and the quality of embryos and the implantation rate seemed normal in subsequent frozen-thawed embryo transfer [43]. However, high E2 levels were found to be deleterious to embryo adhesion in vitro, mainly because they have a direct toxic effect on the embryo that may occur at the cleavage stage [175].
Mouse and human embryos, when cultured in vitro, undergo a delay in development compared with those grown in vivo. This delay can be caused by suboptimal culture conditions, but possible influences of ovarian stimulation cannot be excluded. In the mice, preimplantation embryonic development in vitro and in vivo was found to be negatively influenced by the ovarian stimulation itself, and results in an impaired blastocyst formation and fetal growth retardation at day 14 of gestation. The authors suggested that a similar negative effect of ovarian stimulation on oocyte and embryo quality seems likely in IVF-ET which might explain in part for the delay in embryonic development after IVF-ET, and for the low birth weight often observed after assisted reproductive technologies [165].
Effect of estrogen on blastocyst hatching
The successful hatching of the embryos is thought to be a key event in the implantation process. One reason for the low implantation rate that has been suggested is the limited ability of blastocysts to hatch from the zona pellucida. Suboptimal culture conditions might induce the hardening of zona pellucida, which could limit the hatching ability of embryos [176]. To help embryos hatching from their zona pellucida during blastocyst expansion, different types of assisted hatching have been developed, including mechanical partial zona dissection or zona drilling, chemical zona drilling with acidic Tyrode's solution, and the laser dissection technique [177]. However, there is controversy about the benefit of assisted hatching on the improvement of the implantation rate and pregnancy rate. Some agreed as to the benefit of assisted hatching in women with advanced age, in women with repeated IVF-ET failure [178].
The degree of zona pellucida thickness variation of the transferred embryos has been found to exhibit a strong correlation with clinical pregnancy outcome following IVF-ET treatment and to be important for embryo selection during clinical transfers [179, 180]. Zona pellucida thickness variation and character were found to correlate with implantation. Implantation rates were found to range from 10% for embryos with uniform thickness to 29% with thin or irregular zona pellucida [181, 182]. These reports suggested that patients transferred with embryos with thinner zonae had a better chance of successful implantation and pregnancy as compared to those transferred with embryos having thicker zonae. Other reports have proposed zona pellucida thickness variation as a reliable marker for selecting thawed as well as fresh human embryos for transfers [183, 184]. Cohen [185] conducted a retrospective analysis of zona pellucida thickness of transferred embryos through video recordings and concluded that the variations in zona pellucida thickness rather than the zona pellucida thickness per se of the transferred embryos was a stronger predictor of the IVF-ET.
A significant linear relationship was reported to exist between the mean zona pellucida thickness of each patient and the maximum E2 level and an increasing one with the hMG dose. The authors found that the zona pellucida thickness was basically an individual feature that influenced the fertilization rate [186].
3.1.5 Effect of supraphysiological estrogen levels on the ovaries [corpus luteum], pituitary, and hypothalamus
Abnormalities in the luteal phase have been shown in virtually all the stimulation protocols used in ovarian stimulation, on the hormonal, as well as on the endometrial level. All three aspects of a defective luteal phase, that is, a shortened luteal phase and/or low mid-luteal serum progesterone concentrations and/or abnormal endometrial histology, have been regularly observed in IVF-ET cycles. For that reason, luteal-phase supplementation with hCG or progesterone increases pregnancy rates, and its necessity has been well established, at least in GnRH-agonist cycles [187].
In the 'pre-agonists era', Edwards and Steptoe were the first to postulate luteal phase inadequacy resulting from ovarian stimulation as a cause of failure of IVF-ET cycles. With the introduction of the GnRH-agonist, used in ovarian stimulation cycles to avoid premature luteinizing hormone [LH] surge, luteal phase inadequacy was reported. A meta-analysis of different clinical trials demonstrated beneficial effects of luteal support when ovarian stimulation was carried out with human menopausal gonadotropins [hMG] in association with GnRHa [187].
Recently, GnRH antagonists have become available for clinical use. Whether GnRH antagonists induce down-regulation of pituitary GnRH receptors is still a subject of investigation [188]. It has been demonstrated that chronic administration of the GnRH antagonist Cetrorelix in rats causes an important down-regulation of pituitary GnRH receptors [189].
Direct effect on corpus luteum
Different estradiol receprors have been detected in human corpus luteum, indicating that estrogen might be a local regulator of corpus luteum function [190]. In the pre-agonist era, the alteration of the Estradiol/progesterone (E2/P) ratio was considered a main cause of luteal-phase inadequacy, possibly through the luteolytic action of E2 [5]
Estrogen is thought to exert a direct luteolytic action in primates as exogenous administration of E2 reduces progesterone concentrations during the luteal phase [191], probably via inhibiting the enzyme 3 beta-hydroxysteroid dehydrogenase, which is mandatory for progesterone synthesis [192]. Moreover, although the exact mechanism has not yet been established, estrogen may play a role in the regulation of proteins involved in the process of luteal-cell apoptosis [193].
Normal corpus luteum function is dependent on the proper function of the pituitary gland and hypothalamus. Adequate luteinizing hormone [LH] surge during ovulation and continuous tonic LH pulses during the luteal phase are necessary for the proper development of the corpus luteum. Corpus luteum dysfunction due to the effect of ovarian stimulation on the pituitary gland and hypothalamus will be discussed later.
Effect on the pituitary gland and hypothalamus
Normal corpus luteum function requires optimal follicular development in the follicular phase, especially follicle stimulating hormone (FSH) stimulation, adequate luteinizing hormone (LH) surge during ovulation and continuous tonic LH pulses during the luteal phase. In turn, the normal luteal phase is characterized by an optimal hormonal environment and adequate endometrial secretory transformation. As many factors contribute to a normal corpus luteum function, any alteration might exert a deleterious effect on the final target, the endometrium, leading to embryo/endometrial asynchrony [5]. Luteal phase insufficiency due to corpus luteum defects and LH suppression is known to be associated with failure to achieve and maintain pregnancy [194].
Effect on LH secretion
The lifespan and steroidogenic capacity of the human corpus luteum is dependent on continuous tonic luteinizing hormone secretion as well as healthy adequate gonadotropins surge. Feedback mechanisms from ovarian steroids and GnRH pulses regulate LH secretion during the luteal phase, but a number of autocrine and paracrine factors within the ovary might also play a role in controlling corpus luteum function [5].
Effect on LH surge
It is obvious that during COH for ART, the prevention of endogenous LH surge is mandatory to avoid the occurrence of premature LH surge as well as for timing of oocyte retrieval.
However, during ovarian stimulation cycles in which no pituitary suppression is used, there are data that suggest a defective endogenous gonadotropins surge. The gonadotropins surge is an event crucial for final oocyte maturation, ovulation, and subsequently for corpus luteum function. The duration of the LH peak seems to be more important than its amplitude for the induction of ovulation [195]. Ovulation induction by hCG is not physiological; the absence of an FSH surge, and the long duration of LH activity associated with hCG action, would contribute to some of the luteal phase abnormalities [196].
Messinis demonstrated that an attenuated LH surge is obtained in normally cycling women during superovulation induction with sequential clomiphene/hMG treatment. The peak values and the duration of the LH surge to have significant negative correlations with the plasma E2 levels, the number of follicles, and the total follicular fluid volume aspirated at laparoscopy. This suggests that during superovulation induction for IVF-ET, the endogenous LH surge is attenuated by factors, which are related to the degree of ovarian hyperstimulation [197].
As shown above, the induction of superovulation in women with human gonadotropins may result in blockage of the endogenous LH surge, but the reasons for this are not known. A high number of small follicles have been suggested to have a suppressive effect on both tonic and mid-cycle gonadotropins secretion [198].
Effect of abnormal LH surge on nuclear maturation of the egg
A timely LH surge of adequate amplitude and sufficient duration is important to bring about rapid and complex cellular differentiation, resulting in cascades of tightly coupled biochemical events, which initiate oocyte maturation, ovulation, and corpus luteum formation [199]. It is known that a midcycle LH surge of sufficiently high amplitude and duration is important for both nuclear and cytoplasmic maturation, which ensure the normal fertilization and developing potential of oocytes [199]. For PMSG-hyperstimulated rats, higher doses of hCG are required to completely ovulate the expanded cohort of preovulatory follicles [200].
Effect on LH tonic pulse during the luteal phase
Low luteal LH levels have been described after human menopausal gonadotropins treatment6 and after GnRH-agonist treatment or after GnRH-antagonist treatment. These low, almost undetectable, luteal LH levels may not be able to support corpus luteum. As a result, a shortened luteal phase and low mid-luteal progesterone concentrations have been described in cycles stimulated with the association either of a GnRH agonist or a GnRH antagonist [195, 196].
Supraphysiological progesterone serum concentrations may also interfere with the pituitary's luteinizing hormone secretion by disturbing the feedback control mechanisms and may result in a reduction of the LH serum levels [195, 196]. Progesterone modulates LH secretion during the luteal phase by influencing the LH pulse amplitude and pituitary release of LH [201]. A longer exposure to progesterone or the combined action of estrogen and progesterone decreases LH release [302]. As ovarian stimulation results in supraphysiological steroid serum concentrations, these high steroid levels may adversely affect LH secretion via a long-loop feedback mechanism. In turn, disturbed LH secretion may induce a luteal-phase defect with premature luteolysis, low progesterone levels and shortened luteal phase. It might therefore be hypothesized that deviation from the normal hormonal environment could be a prevalent effect of ovarian stimulation in the luteal phase; despite the use of different stimulation protocols [7]. This might be a possible explanation of the observation that in natural cycle, luteal-phase length was normal after GnRH-antagonist treatment [203]. However, in GnRH-antagonist cycles and after minimal ovarian stimulation, luteal-phase length was normal despite an abnormal endocrine profile [204].
3.1.6 Other probable effects of supraphysiological estrogen levels
There are other less defined probable mechanisms through which supraphysiological estrogen levels may cause adverse effects on the outcome of infertility treatment.
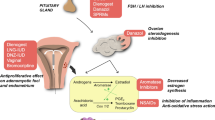
(A) Leptin-mediated effect
Recently, an important role for the leptin, the secretory product of adipocytes, in reproductive medicine has emerged and it is interesting to discuss in brief a possible link between leptin, induction of ovulation and aromatase inhibitors.
Soon after its discovery in the early 1990s, it was recognized that leptin played a significant role in reproduction, providing a critical link between metabolic state and fertility. It now appears that leptin may have an important role in both normal ovarian physiology and pathophysiology. Research has revealed that a minimum level of leptin stimulation is required for maintenance of fertility in animals and humans. Conversely, elevated leptin levels may impair fertility [205].
Leptin is a secretory product of adipocytes that correlates significantly with the body mass index with increased levels in obese women [206]. It can influence reproduction through central (on GnRH neural system) and peripheral (on the ovary directly) mechanisms [205]. A positive correlation has been found between leptin and BMI, as well as between leptin and testosterone in women with polycystic ovarian syndrome (PCOS) [207].
Numerous studies have shown that circulating leptin concentrations rise in parallel with E2 during ovarian stimulation [208–213]. These changes are not likely to be a direct action of FSH because FSH decreases as E2 and leptin rise during natural cycles, and high FSH levels experienced during ovarian stimulation simulate endogenous levels in postmenopausal women, who have lower serum leptin concentrations than premenopausal women [205]. Bützow [206] found that the larger the increase in serum leptin concentrations during FSH stimulation, the poorer the ovarian response in terms of number of follicles and retrieved oocytes. Moreover, higher serum leptin levels were found in oligo- and amenorrheic women who failed to respond to clomiphene therapy [214].
The effect of body weight on outcomes of assisted reproduction has been investigated. Fedorcsak [215] reported that among patients who conceived, overweight patients (BMI >25) had fewer oocytes retrieved, a higher miscarriage rate, and lower live birth rate. In a much larger retrospective study that included 8822 embryo transfer cycles, the cumulative pregnancy rate progressively decreased as BMI increased from <25 to >35 [216].
Higher follicular fluid leptin concentrations correlated with lower intrafollicular oxygen concentration [pO2], [217] a condition that negatively impacts oocyte developmental competence [218] with a direct evidence of a relationship between leptin levels and ART outcome reported by Mantzoros [219] who found significantly lower follicular fluid leptin concentrations in women who became pregnant within three cycles of IVF-ET or gamete intrafallopian transfer (GIFT). More recently, a significant negative correlation between non-fasting serum leptin levels measured at the beginning of FSH stimulation and pregnancy success in women undergoing first attempt IVF-ET cycles was reported [220]. Moreover, as leptin receptors are expressed in the human endometrium, [221] a role for leptin in endometrial receptivity cannot be excluded. These results imply that elevated leptin may be a key factor in obesity-related fertility problems, and conversely that elevated leptin may negatively impact fertility independently of body mass. Fewer good embryos and lower implantation rate suggest that elevated leptin impairs oocyte developmental competence and/or early cleavage stage embryo development, possibly via direct actions on the follicle [205].
E2 has been reported to increase leptin mRNA expression. In human adipose tissue culture, E2 stimulated leptin secretion in women but not in men [222]. In women, E2 was found to increase ob mRNA expression and leptin release. Moreover, in adipose tissue of women, the estrogen precursors; testosterone and dehydroepiandrosterone also induced an increase in leptin secretion, an effect that was prevented by the aromatase inhibitor letrozole. Moreover, the stimulatory effect of E2 observed in women was antagonized by the antiestrogen ICI182780 [223].
(B) Coagulation system
Estrogen has been pointed out as a pre-thrombotic factor. It has long been established that both pregnancy and oral contraceptive use have resulted in an increased level of many coagulation parameters. Generally, this has been thought to be a result of the estrogen component. Although it has been well established that long-term exposure to exogenous contraceptive steroids can have a promoting influence on the potential for thrombosis in women, it is less clear what role high levels of endogenous steroids might play. Undefined coagulation abnormalities were reported after hMG- and hCG-induced hyperstimulation of the ovary in several women [224–227]. Kim et al., [225] noted a large increase in fibrinogen after hMG treatment, accompanied by "significant increases" in the prothrombin time. However, the statistically significant activation of clotting factors occurring during controlled ovarian hyperstimulation is usually not accompanied by clinically significant coagulation disorders that may be explained by either of the following: (1) the increased levels of clotting activity were still "within normal limits"; (2) none of the patients had any conditions known to predispose to coagulopathies (history of coagulopathies, phlebitis, damaged or compromised endothelial cells, etc.); or (3) a combination of the two [226]. However, such subtle coagulation disturbances may exert an adverse effect on endometrial receptivity and the early development of the placenta by affecting the microcirculation.
Two case reports were published of activated protein C-resistant women who suffered a thrombotic event during IVF-ET treatment [227, 228]. Curvers [229] reported that the only coagulation parameter that changed considerably during IVF-ET treatment was the activated protein C (APC). In their study, the authors observed that hyperstimulation, i.e. high estrogen levels, induce APC resistance, and that under these conditions both the absolute values and the changes in the APC and the estrogen levels (hyperstimulation-baseline) correlate significantly. Prior to that study, it was reported that high estrogen levels were not associated with APC resistance [230, 231]. However, the APC resistance test used in these studies, which is based on quantification of the effect of APC on the clotting of plasma initiated via the intrinsic coagulation pathway, is not very sensitive to changes in sex hormones [232]. On the other hand, Curvers et al. [232], used an assay that quantifies down-regulation of extrinsic coagulation by APC that is particularly sensitive to hormonal changes in women [232].
We think it is possible that the reported effect of ovarian stimulation-associated supraphysiological levels of estrogen on the coagulation system, though being modest, may contribute at least partially to the reduced implantation rate observed after assisted reproduction. This could be due to an impact on the microcirculation in the endometrium that could affect the early stages of the implantation as well as the early development of the placenta. However, more studies are needed to support this hypothesis.
(C) Tubes and tubal transfer
Effect on the oviductal environment
Oviducts are biologically active, providing an environment that sustains and enhances fertilization during early embryonic development as the embryo travels toward the uterine cavity. Following superovulation, the fluid from the oviduct seems to impair embryo development. Furthermore, a stimulated oviductal environment has also been shown to have a negative influence on the implantation capacity of mouse embryos [233].
Superovulation in the mouse was described as a model for intra-uterine growth retardation
Superovulation is associated with a slower preimplantation embryo development, a later and impaired implantation and a prolonged gestation [234]. This suggests that the oviductal milieu rather than the embryo quality are responsible for the adverse effects observed after superovulation. The stimulated oviductal environment impairs the developmental capacity of embryos in comparison with untreated pseudopregnant females. In-vitro culture is also suboptimal but better than the stimulated oviductal environment. However, a detrimental effect of hormonal stimulation upon the oviductal environment has not yet been demonstrated in the human. A possible potential negative effect, however, is not contradicted by observations reporting a higher pregnancy rate after ovarian stimulation and IUI compared to insemination alone [235]. In the human, there are observations that GIFT results in higher pregnancy rates in comparison with IVF-ET [236, 237]. It is difficult to interpret these data since the embryos were exposed longer to the possible deleterious stimulated oviductal environment-using GIFT whereas in IVF-ET, possible "suboptimal" culture conditions as opposed to in-vivo conditions were used. Since the introduction of sequential culture media, incorporating amino acids, vitamins and growth factors, the in-vitro culture of embryos has been improved, resulting in implantation rates of >50% per transferred blastocyst [238]. Confirmation of these data in a trial would result in higher pregnancy rates than ever reported for GIFT [239].
Effect on the oviductal embryo transfer
Akira [240] has found that ovarian stimulation was associated with accelerated oviductal embryo transport. The authors have concluded that increased implantation failure in superovulated rats may result from the accelerated embryo transport resulting from elevated E2/P ratio. Accelerated oocyte/embryo transfer has been postulated to be the mechanism behind which high doses of estrogen work as postcoital emergency contraception as explained later.
Post-coital contraception with estrogen
The achievement of very high levels of estrogen by administering exogenous estrogen has been suggested for postcoital contraception as early as the 1960s, when high-dose estrogen was identified as a highly effective emergency contraceptive [241].
Greenwald [242] compared the response of the rabbit, rat, mouse, hamster, and guinea pig, to a single post-coital injection of E2 cyclopentylpropionate and showed that post-coital treatment with estrogens caused either tube locking of embryos or accelerated transport to the uterus. Although other effects were also detected, the alteration in oviductal transport accounted for the contraceptive effect. Embryos that entered the uterus prematurely were expelled whereas whose sojourn through the oviduct was prolonged, degenerated.
Other investigators found that a single injection of E2 was given at different times after coitus revealed that a wide range of effectiveness can be achieved and suggested different mechanisms can account for the contraceptive effect when the same steroid is given at different times post-coitus [243, 244]. Some believed the main target was the endometrium where they observed stromal edema, hemorrhage, and loss of decidua, all of which was considered unsuitable for implantation. Administering high dose of estrogen in the preovulatory phase was found to depress endometrial growth and angiogenesis through a negative influence on the vascular endothelial growth factor [245]. This suggested that estrogens might interfere with endometrial receptivity even if given before ovulation.
Other mechanisms for the contraceptive effect of post-coital estrogen were found to operate in monkeys. If given in the follicular phase, so as to advance the preovulatory increment in plasma estrogen, they evoke a premature LH surge that does not trigger ovulation and the formation of a functional corpus luteum, and the spontaneous LH surge is delayed or suppressed [246]. ER present in the granulosa cells of antral and preovulatory follicles and in luteal cells [247] allow for a diversity of effects in the ovary. In rhesus monkey, supraphysiological doses of estrogen given in the mid or late follicular phase induce atresia or luteinization without rupture of the dominant follicle, reduce the viability of granulosa cells, reduce the synthesis of E2 and progesterone and are detrimental to the oocyte [248].
Whatever the mechanism of action of high dose of estrogen for emergency contraception is, success in preventing pregnancy provides another evidence of the toxic effect of the supraphysiological estrogen levels fertilization, implantation and early development of the embryo which result in failure of achieving pregnancy.
[D] Effect on other factors involved in decidualization and early developing placenta
Paracrine factors
The importance of paracrine factors in mediating the cellular and biochemical changes involved in embryo implantation has been recognized. Many growth factors and cytokines, such as inhibins and activins, whose expression is generally limited to developmental and pathological states, are produced by actively remodeling endometrial cells, and play crucial roles in regulating endometrial cell function. Example of these factors includes the inhibin and activin family in the paracrine regulation of endometrial receptivity, decidualization and implantation. Estrogen is known to play an important role in regulation of these factors as discussed by Jones [249].
Role of Calcitonin
The peptide hormone calcitonin is currently being evaluated as a potential marker of the fertile human endometrium. In ovariectomized animals, it has been shown that administration of estrogen together with progesterone inhibits progesterone-mediated calcitonin gene induction.29 Such antagonistic interactions between estrogen and progesterone pathways have been documented previously in breast and uterine cells. It has been proposed that these phenomena reflect transcriptional cross talk occurring between estrogen and PR co-expressed in the same target tissue. A complex interplay of the two ovarian hormones, progesterone and estrogen, in the uterine milieu is believed to be critical for optimal calcitonin gene expression [250, 251].
3.2 Effect of the medications used for ovarian stimulation
3.2.1 Clomiphene citrate
In spite of the high ovulation rate with the use of clomiphene citrate (around 50–90%), the pregnancy rate is much lower (around 20–40%) [252–254]. Moreover, there is a higher than expected incidence of miscarriage in conception cycles following clomiphene citrate treatment [255]. Such discrepancy is believed to be due to the peripheral antiestrogenic effect of clomiphene citrate, particularly at the level of the cervical mucus [256, 257] and endometrium, [258, 259]. The persistence of the zu-isomer of clomiphene citrate in the body due to its long half-life (several weeks) and slow clearance adds to the accumulation of the antiestrogenic effects over subsequent cycles of administration [260, 261]. There is also evidence of a direct harmful effect of a high concentration of clomiphene citrate and its isomers on fertilization, and on early mouse [262] and rabbit [263] embryo development. Such effects, however, were not confirmed in other studies [264, 265]. There is still some controversy concerning a direct effect on the quality of oocytes associated with clomiphene citrate treatment [266, 267]. Decreased uterine blood flow during the early luteal phase and the peri-implantation stage is another explanation for the poor outcome of clomiphene citrate treatment [268]. Other investigators have suggested the presence of other unrecognized infertility factors [269, 270].
3.2.2 Gonadotropins
The existence of nongonadal gonadotropins (FSH, LH/hCG) receptors was first suggested by Ziecik [270] who conducted binding studies in the porcine uterus. Subsequently, nongonadal receptors were found in human tissues, including endometrium, myometrium, fallopian tube, umbilical cord, and brain, by using a variety of techniques, including immunohistochemistry; Northern, Western, and ligand blotting; and in situ hybridization [271–274]. This would suggest a possible direct action of the gonadotropins on the uterus and involvement in the endometrial development, implantation and establishment of pregnancy [275].
High concentrations of exogenous gonadotropins used for hyperstimulation of folliculogenesis were shown to be detrimental to oocyte and embryo development in many animal species. This is believed to be due to the associated supraphysiological E2 levels and other possible undetermined factors associated with ovarian stimulation. However, a direct effect of gonadotropins cannot be ruled out. It has been reported that early embryo loss due to superovulation could be rescued by an injection of goat antiserum against PMSG [276].
Effect of gonadotropins on LH surge
It has been suggested that during ovarian stimulation, the supraphysiological FSH levels that persist into the late follicular phase, thereby overriding selection of the single dominant follicle of the natural cycle, secretion of an ovarian factor(s) blocks estrogen-induced LH surges [277]. Accumulated evidence has indicated that the ovaries produce another non-steroidal substance, named "gonadotropins surge-attenuating factor" (GnSAF), which may play a role in the control of the midcycle luteinizing hormone surge in women [278–281]. Although GnSAF activity is present during superovulation induction, it is still unclear whether this factor plays a physiological role during the normal menstrual cycle. Treatment with FSH initially attenuated the response of LH to GnRH via the production of GnSAF from the ovaries, while around the midfollicular phase, the rising concentrations of E2 were able to overcome the attenuating effect of GnSAF and increase pituitary sensitivity to GnRH. The increased pituitary sensitivity in the midfollicular phase of the FSH-treated cycles, however, was not further enhanced in the late follicular phase despite the continuous rise in E2 values [282]. It is suggested that eventually GnSAF was able to overcome the sensitizing effect of E2
Effect of gonadotropins on chromosomal development
A dose-response relationship between the PMSG dose and the incidence of polyploidy in the CD-1 mouse has been reported with the level of polyploidy rising from 2.9% with 10 IU PMSG to 10.5% with 15 IU PMSG, in the zygot stage. Both a disturbance at maturation division and an error at fertilization were the cause of polyploidy [22]. Whether this is the direct effect of PMSG or the resultant ovarian stimulation is not known.
3.2.3 GnRH analogues
GnRH plays a pivotal role in the control of female reproduction and is secreted by hypothalamic neurons in a pulsatile way. It binds to specific receptors on pituitary gonadotrophs, which is followed by the secretion of the gonadotropins, LH and FSH, which regulate steroidogenesis and gametogenesis in the ovary [283].
GnRH analogues are able to suppress gonadotropins release and, subsequently, gonadal function. This is the basis for their clinical application in ovarian stimulation. Several agonistic or antagonistic GnRH analogues have been developed for this purpose [284].
Effect of GnRH analogues on ovarian steroidogenesis and corpus luteum function
GnRH receptors and GnRH receptor mRNA were found to be expressed in human granulosa-luteal cells GnRH receptors have been found in the luteinized human granulosa cells, and a possible direct effect of GnRH on ovarian steroidogenesis has been suggested [485, 486].
Although there are many studies that investigated the possible direct effect of various GnRH agonists on ovarian steroidogenesis and hence the corpus luteum, there is no consensus of the effects in the human ovary. Also, there is little information concerning the direct effect of GnRH antagonists on ovarian steroidogenesis [487]. A few studies that looked at the effect of GnRH agonist treatments in vivo on in vitro steroidogenesis by human luteinized granulosa cells, found an impairment of progesterone production [287–290]. Minaretzis [291] have reported on the effect of the GnRH antagonist Nal-Glu compared with GnRH agonist leuprolide acetate treatment of patients undergoing controlled ovarian hyperstimulation. The progesterone production in human luteinized granulosa cells cultures was not different between the two groups. Both GnRH antagonists had no effect on basal or hCG-induced E2 or progesterone production by granulose lutein cells, independent of whether the cells were exposed to the compounds in vitro or in vivo. However, an in-vitro study demonstrated an inhibitory effect of GnRH antagonist on gonadal steroid secretion [292]. Pellicer and Miro who found that GnRH agonist exposure in vivo may affect human luteinized granulosa cells function in vitro. They compared the progesterone accumulation in human luteinized granulosa cells cultures between patients treated with clomiphene citrate/gonadotropins and GnRH agonist/gonadotropins. Granulosa cells obtained from patients treated with the GnRH agonist had lower progesterone production than cells isolated from women treated with clomiphene citrate/gonadotropins [293]. However, Minaretzis compared the human luteinized granulosa cells steroidogenesis in vitro from Nal-Glu and leuprolide acetate-treated women respectively. They reported that basal and gonadotropins-stimulated progesterone secretion was similar in the two treatment groups [291].
Weiss et al., found that in vivo treatment with triptorelin or the two GnRH antagonists, cetrorelix and ganirelix; did not have an effect on spontaneous or hCG-stimulated steroidogenesis. The authors also performed in vitro treatments with triptorelin, cetrorelix and ganirelix for up to 96 hours and did not find any effect of these treatments on basal or hCG-stimulated steroid production [287].
In all prior reports, which studied the effect of GnRH agonist treatment on ovarian steroidogenesis, only the effect of hCG on progesterone secretion by human luteinized granulose cells was investigated. Recently, we examined the effect of two known physiologic stimuli of ovarian steroidogenesis, hCG and insulin. We found that GnRH agonist treatment affected both the basal and hCG-stimulated progesterone production by human luteinized granulosa cells but the pattern of insulin-stimulated progesterone secretion was not affected. Because insulin and hCG may share common pathways beyond but not at the level of receptor activation, we hypothesize that in-vivo GnRH agonist might affect the expression and/or activation of LH receptors. In our study, we found that human luteinized granulosa cells from patients treated with GnRH antagonist responded to hCG in a fashion generally consistent with previously reported hCG-stimulated human luteinized granulosa cells progesterone production [294]. In addition, the basal progesterone production and absolute levels of insulin-stimulated progesterone production were both significantly higher than observed following GnRH agonist treatment. In contrast, human luteinized granulosa cells previously exposed in vivo to GnRH agonist had a blunted P response to hCG. Our results, therefore, support the hypothesis that GnRH agonists may have a direct negative effect on human ovarian steroidogenesis by the corpus luteum and suggest that luteal function may be less affected during IVF-ET cycles when GnRH antagonist is used. However, in another study, GnRH antagonist therapy in women undergoing ovarian stimulation was found to be associated with a significant effect on ovarian follicular steroidogenesis. The authors found the mean follicular fluid E2 concentration significantly lower in patients treated with GnRH antagonist than in those treated with GnRH agonist. However, no significant differences were found between groups in follicular fluid progesterone concentrations [295]. However, further studies are required to investigate the effect of GnRH agonist and antagonist on gonadotropins receptors and different enzymes involved in ovarian steroidogenesis by the corpus luteum.
Effect on LH secretion
Because pituitary LH secretion is dependent on GnRH stimulation and feedback mechanisms from ovarian steroids, any alteration may be deleterious. In cycles using GnRH agonists for ovarian stimulation, a significant drop in mid-luteal progesterone concentrations was observed, consistent with corpus luteum insufficiency. Long-term GnRH-agonist administration has been associated with a profound desensitization of the pituitary cells. In fact, studies on pituitary gonadotropins secretory capacity after GnRH-agonist treatment have indicated that this remains impaired for at least 14 days after the discontinuation of the GnRH-agonist and for the whole length of the luteal phase [296]. In a randomized trial, it was also demonstrated that, despite the early cessation of the GnRH agonist in the follicular phase, luteal-phase characteristics were abnormal [297].
GnRH-antagonist treatment has been shown to be effective in blocking the LH surge. GnRH antagonists bind competitively to pituitary GnRH receptors and cause an immediate inhibition of gonadotropins release. In contrast to GnRH agonists, it was suggested that treatment with GnRH antagonists might not adversely affect luteal LH secretion, since the pituitary maintains its responsiveness to the endogenous GnRH stimulus [298]. A normal luteal phase, in terms of duration and serum progesterone concentrations, was observed in natural cycles in which an antagonist was administered to prevent the LH surge [299]. However, data on the luteal phase from unsupplemented cycles after antagonist administration are limited as a result of the small number of patients involved and are rather controversial. Four out of six patients had either a shortened luteal phase or low progesterone concentrations in cycles stimulated with HMG and the GnRH antagonist Cetrorelix and receiving no luteal-phase supplementation. However, with or without luteal-phase supplementation, luteal LH levels were low, indicating that another mechanism might be involved [299, 300].
3.2.4 Human chorionic gonadotropins
Supraphysiological steroid serum concentrations may interfere with LH secretion via long-loop feedback, but, additionally, the exogenously administered hCG might amplify LH secretion arrest via a second short-loop negative feedback. Although in the monkeys such a negative feedback exists [301] there is a debate in the literature about its existence in humans, with some of the studies supporting this hypothesis [302] and others not [303, 304]. Nevertheless, findings from in-vitro studies further support this idea. GT1-7 neurons, which are morphologically and functionally similar to GnRH neurons, were found to contain LH/hCG receptors. In addition, exogenously administered hCG was found to decrease the expression of GnRH receptor gene in GT1-7 cells or GnRH secretion in immortalized GnRH neurons [305].
4 Measure to improve treatment outcome after ovarian stimulation and assisted reproduction
Even if the results of reproductive medicine have improved in terms of numbers of pregnancies, it is still striking that it is necessary to use stimulation which sometimes leads to hyperstimulation and multiple pregnancies, that embryo development in vitro is still limited, that implantation only occurs for 15–20% of transferred embryos and this ratio has not changed significantly along the last 25 years. We still need to improve techniques to gain pregnancy rates approaching 50% per embryo. During assisted reproduction technology interventions, controlled ovarian stimulation is used to obtain several oocytes in attempts to increase the likelihood of having at least one developmentally competent embryo available for transfer. However, current techniques for identifying the competent embryo[s] are by no means perfect. These limitations, coupled with pressures to maximize the chance of pregnancy, typically result in the transfer of multiple embryos. Not surprisingly, this practice has resulted in an unacceptably high rate of multiple pregnancies arising from IVF-ET. During the last few years, concerted efforts have focused on reducing these rates by restricting the number of embryos to transfer [306].
In order to fulfill this task, several approaches have been suggested to improve the outcome of treatment after ovarian stimulation and assisted reproduction. Table 3 summarizes the different approaches including the two main approaches tried so far i.e. optimizing ovarian stimulation protocols and improving embryology techniques. A third approach we suggest i.e. the use of aromatase inhibitors constitutes the third and most recent approach.
We suggest a THIRD novel approach that we believe it may carry a hope for improving the outcome of treatment after ovarian stimulation and assisted reproduction. This approach involves the use of aromatase inhibitors.
4.1 Modified ovarian stimulation protocols
4.1.1 Reducing the intensity of ovarian stimulation protocols
Different approaches have been suggested to improve the treatment outcome during assisted reproduction by reducing the intensity of ovarian stimulation that would reduce the high estrogen levels [307]. These approaches include no-stimulation (natural cycle IVF-ET), minimal stimulation IVF-ET cycles, step-down protocols, FSH coasting and in vitro maturation.
However, all these measures are associated with the major drawback of affecting the main goal of ovarian stimulation, which is the achievement of a suitable number of embryos when a good number of oocytes retrieved.
Natural cycle IVF-ET
The first IVF-ET treatment in the literature was a natural cycle IVF-ET. Since then natural cycle IVF-ET has been largely replaced by IVF-ET with ovarian stimulation. Natural cycle IVF-ET has several advantages including a close to zero multiple pregnancy rate, and a zero risk of ovarian hyperstimulation syndrome as well as being less time consuming, physically and emotionally less demanding for patients, and cheaper than stimulated IVF-ET. However, it seems to be less effective. Unfortunately, good quality randomized controlled trials and formal cost-effectiveness analyses are lacking. Pelinck reviewed 20 selected studies that comprised a total of 1800 cycles of natural cycle IVF-ET, resulting in 819 embryo transfers (45.5% per cycle) and 129 ongoing pregnancies (7.2% per cycle and 15.8% per embryo transfer). The authors concluded that: in spite of being a low-risk, low-cost and patient-friendly procedure, high cancellation rates because of premature LH rise and premature ovulations hamper efficacy of natural cycle IVF-ET and that a randomized controlled trial comparing natural cycle IVF-ET with current standard treatment strategies is warranted. [27].
The cumulative live birth rates after four cycles of treatment was reported to be 32% which is comparable with the value of 34% for women having conventional IVF-ET treatment [308]. However, there is enough data on cumulative live birth rates following conventional IVF-ET, which is likely that current cumulative rates for IVF-ET will be higher than previously reported.
In the implantation rate per embryo after natural cycle IVF-ET was found quite acceptable up to 50%. Also, the fertilization rate was found to be 100% for the oocytes retrieved from the mature follicles (as reviewed by [27] Pelinck). We believe that the significantly better oocyte quality (100% fertilization rate) as well as better implantation rate (about 50%), reflect clearly the value of avoiding the deleterious effects of ovarian stimulation on the treatment outcome. Moreover, there is an important point to mention regarding the success rates of natural IVF-ET cycles, which is the bias in selecting patients who underwent natural IVF-ET cycles. Many studies included patients with unfavorable outcome such as poor responders, old age and repeated IVF-ET failures.
Minimal stimulation IVF-ET
Minimal stimulation for IVF-ET that is less expensive than full stimulation and minimizes monitoring and patient discomfort has been described for almost 10 years and found to be associated with acceptable pregnancy rates and was suggested as an attractive alternative to select patients undergoing IVF-ET [309]. This protocol involves the use of CC plus FSH alone or in conjunction with GnRH antagonist to prevent premature LH surge [310]. It is expected that the minimal-stimulation regimens produce fewer oocytes, fewer embryos available after fertilization, and fewer excess embryos available for cryopreservation. This leads to the main drawback of the minimal-stimulation regimen, which is the lower total reproductive potential (pregnancies resulting from fresh and frozen-thawed embryos resulting from a single stimulation cycle) [311].
FSH step-down and coasting protocols
Other measures have been tried by decreasing the FSH dose (step down protocol) to improve endometrial receptivity. With the use of a step-down regimen with FSH in high responders, uterine receptivity was believed to improve secondary to lowering E2 levels during the preimplantation period [312].
Coasting or withholding FSH injection for a period of time has been suggested in patients at substantial risk for the development of severe hyperstimulation syndrome. Coasting in a studied subset of IVF-ET patients did not adversely affect cycle outcome parameters when it was not prolonged [313].
4.1.2 Application of new medications for ovarian stimulation
The ongoing development of more effective and safer pharmacological compounds is driving fundamental changes in medicine [314]. Along the last decade several new medications have been developed for application in ovarian stimulation and assisted reproduction including medications prepared by the new recombinant technology [recombinant FSH and LH] and GnRH antagonists as well as other medications.
Recombinant FSH
The introduction of recombinant FSH into the clinical management of patients suffering from infertility appears to be associated with several treatment benefits when compared with urinary human menopausal gonadotropins
The fact that rFSH preparations have batch-to-batch consistency, are free from urinary protein contaminants and have the potential to be produced in limitless quantities is advantageous. The question whether newer, more pure FSH products are beneficial from the clinical perspective, has not been settled beyond reasonable doubt. The price of rFSH has been reported to be three times as high as the price of the former FSH preparations [315].
Although both FSH and LH are required for normal follicular growth and maturation, the precise role of LH is at present still uncertain. Since it was shown in the late 1980s that too high a concentration of LH might have a negative effect on fertilization and embryo quality, the idea arose that pure FSH preparations might be superior to hMG preparations for patients with endogenous LH production [316]. This hypothesis was tested in several clinical trials comparing uFSH with hMG with respect to pregnancy rates per IVF-ET treatment cycle. Statistical significance was not reached in any of these individual studies [316, 317]. However, in 1995 a meta-analysis by Daya was published, which included eight studies and demonstrated a significant difference in favor of uFSH [316]. A few years later this finding was contradicted by a meta-analysis by Agrawal who argued that meta-analyses should take into account the different pituitary desensitization protocols used [318]. When pooling together 11 trials with the most commonly used GnRH agonist protocol [the long protocol], the overall odds ratio for comparing FSH and HMG was not significant. Although this meta-analysis was criticized on the issue of study selection bias, re-analysis following the inclusion and exclusion of selected studies did not change the overall results of the study [316, 318, 319].
The question whether rFSH also leads to more clinical pregnancies per IVF-ET cycle has been addressed by several clinical trials, none of which reached statistical significance. However, two meta-analyses pooling together the results of several trials did show significant treatment effects in favor of rFSH [320, 321]. The interpretation of these meta-analyses is still debated, since they compare two types of rFSH (the alpha and beta variant), as well as different types of urinary FSH (uFSH and uFSH-HP, and in Out's meta-analysis one study with HMG was also included). In Daya's analysis, which received Cochrane status, an absolute increase in pregnancy rates of 3.7% (95% confidence interval [CI] 0.5–6.9%) was demonstrated comparing rFSH with uFSH/uFSH-HP.
Data on the third possible comparison, between rFSH and hMG, are scarce. The studies that have been conducted so far found no statistically significant differences with respect to ongoing pregnancy rates [322–324]. The evidence available at this time, comparing the different FSH containing gonadotropins preparations, concerns their usage in IVF-ET. However, almost half of all gonadotropins are used in other settings, such as IUI. Very little data are available comparing the preparations for this indication. With respect to adverse effects of exogenous FSH administration, no significant differences have been found between products. The main risk associated with the use of FSH containing gonadotropins products is the development of the ovarian hyperstimulation syndrome (OHSS). The incidence of OHSS does not differ between products [321–324].
Recombinant LH
hCG has been used as a surrogate LH surge because of the degree of homology between the two hormones. The major differences between the two hormones include the sequence of the β-subunit, the regulation of the secretion of the two hormones, and the pharmacokinetics of clearance of hCG as opposed to LH [325, 326].
hCG has a slower plasma metabolic clearance, which consists of a rapid phase in the first 5–9 h following IM administration and a slower phase in the 1–1.3 days after administration. After 36 h, the calculated half-life of hCG was 2.32 days, as compared with LH, for which estimates have ranged from 1 h [326] to 3–5 h [327, 328]. By day 10 after administration, less than 10% of the originally administered hCG was measurable [329]. However, the long serum half-life of hCG is likely to be an undesirable characteristic in clinical practice.
Until recently, there was no other biologic preparation that was as effective as hCG in triggering the final stage of ovulation. Recent developments using genetic engineering technologies and posttranscriptional biosynthesis have led to the production of rhLH, which has been available for use in clinical trials for several years [330]. The rhLH produced in vitro is purified and further formulated to yield a pharmaceutical preparation with very high specific immunoactivity and bioactivity. A crossover pharmacokinetic study has been performed in nonhuman primates to assess and compare pituitary, urinary, and rhLH [331]. After intravenous administration of pituitary and urinary-derived LH, and rhLH, mean concentration-time curves were parallel. The mean areas under the curve (AUC) for concentration by time after dose curves were similar, after correction for immunologic differences in dose. The mean clearance estimates and volume of distribution at steady state, distribution, and terminal half-lives were similar for all three types of LH. Furthermore, the results of studies in human volunteers show that rhLH and urinary-derived hLH have similar pharmacokinetic characteristics; both preparations presented an iv distribution half-life of 1.2 h and a terminal half-life of 10 h [332, 333]. rhLH total body clearance was 2 L/h, with less than 5% of the dose being excreted by the kidneys. The steady state volume of distribution was approximately 8 L [332]. Furthermore, it has been demonstrated that co-administration of rhLH and rhFSH (follitropin α) does not modify their respective pharmacokinetic characteristics [332]. Finally, low doses of rhLH have been shown to promote ovarian steroidogenesis when follicular growth is induced with rhFSH in hypogonadotropic hypogonadal women [334].
The results show that a single dose of rhLH is effective in inducing final follicular maturation and early luteinization in IVF-ET and embryo transfer patients and is comparable with 5,000 IU u-hCG. A single dose of rhLH results in a highly significant reduction in OHSS compared with hCG. The dose of rhLH giving the highest efficacy to safety ratio was between 15,000 and 30,000 IU. This study has shown that the incidence of OHSS is significantly smaller when using a single dose of rhLH for induction of final maturation of follicles and oocytes in women treated for IVF-ET, compared with the use of 5,000 IU u-hCG or two doses of rhLH [335].
Chandrasekher [336] compared rhLH with u-hCG and pituitary hLH as an ovulatory stimulus in rhesus monkeys before IVF-ET. These investigators found that a single injection of 2,500 IU rhLH was as effective as 1,000 IU u-hCG in inducing oocyte maturation, oocyte fertilizability, and luteinization of granulosa cells. The data suggested that using a conversion factor of 2.5, a dose of 12,500 IU rhLH would be as effective as 5,000 IU u-hCG in humans. Another study (unpublished data) looked at the spontaneous LH surges in seven healthy female volunteers with normal menstrual cycles after a single injection of either 250 μg GnRH-agonist (buserelin) or 5,000 IU u-hCG administered when the dominant follicle reached a diameter of 17 mm. It was found that the mean Cmax of the spontaneous LH surge was 47 IU/L (95% CI, 28–65 IU/L) and the mean AUC was 1,019 IU h/L (95% CI, 718–1,320 IU h/L). Using rhLH pharmacokinetic characteristics defined in Phase I studies [332, 333], it was calculated that, to obtain a Cmax and an AUC larger than the corresponding mean values observed in spontaneous surges in 95% of cases, single sc injections of between 14,000 IU and 25,000 IU rhLH should be used.
GnRH antagonists
GnRH antagonists do not induce an initial stimulation of gonadotropins release, but cause an immediate and rapid, reversible suppression of gonadotropins secretion. The principal mechanism of action of GnRH antagonists is competitive receptor occupancy of GnRH-r [337].
GnRH antagonists have been introduced clinically to prevent premature endogenous LH surge to replace the GnRH agonists that require time for a state of desensitization to be reached and, to start with, LH secretion actually increases (flare-up). With GnRH antagonists, immediate blockade of pituitary gonadotropins secretion when premature luteinization during IVF-ET stimulation is imminent seemed an obvious approach. Several studies of dose and treatment schedules have been done, [338, 339] and two general approaches have emerged. The first is a single subcutaneous injection of a large dose on about the eighth day of stimulation with gonadotropins. The alternative is multiple (five or six daily) injections of a small dose from about day 6 of stimulation until the day that hCG for final oocyte maturation is given.
The next step in the development of GnRH antagonists use was to establish whether a GnRH antagonist is at least as effective as a GnRH agonist as the established reference medication testing the regimen of repeated antagonist injections [340, 341] or the regimen of a single-dose regimen [342].
With repeated injections, treatment with gonadotropins was found to be shortened by 1–2 days with slightly fewer follicles at the time of hCG injection so the number of recovered oocytes tends to be lower. No significant difference was found with respect to percentages of metaphase II oocytes, fertilization rates, and number of good quality embryos [340, 341].
Pregnancy rates were high in both groups in all four studies but in every one the rate was lower in the antagonist group. A meta-analysis of the five randomized trials, showed an overall significantly lower rate of pregnancy of 5%. This meta-analysis unfortunately included the study that compared a single-dose analogue regimen with different gonadotropins starting dose as an additional variable [342]. It has been suggested that the larger numbers of oocytes and embryos with agonists allow better selection [343], although the numbers of good quality embryos do not seem to be different. A direct adverse effect on the embryo cannot yet be ruled out but is not likely.
Antagonist blockade allows immediate reversal of pituitary gonadotropins secretion. This means that in IVF-ET ovulatory ripening can be triggered via endogenous gonadotropins by using a GnRH agonist [344] with the advantage of prevention of ovarian hyperstimulation syndrome, which is thought to result at least in part from the prolonged LH effect of hCG [345]. Finally, the use of GnRH antagonist blockade of premature luteinization can be used in IVF-ET with very low or without any hormonal stimulation, lower risk of overstimulation, and simplification of the procedure [346].
A possible disadvantage of an antagonist in IVF-ET is its narrow therapeutic range with the currently advised doses for repeated injections. Patient compliance needs to be high because there is a risk of premature LH secretion if an injection is missed [337].
Another disadvantage is the unpredictable timing of the start of the IVF-ET procedure, which begins with the administration of FSH on day 3 of the woman's cycle and so depends on how regular her menstruation is. This problem may be solved by pretreatment with an oral contraceptive [335].
4.1.3 Use of adjuvant medication during ovarian stimulation
Insulin sensitizers
The independent effect of insulin resistance on infertility treatment in PCOS is not well defined. Regardless of body weight, insulin-resistant PCOS women need higher gonadotropins doses during ovarian stimulation, and insulin resistance is also associated with a risk of multifollicular development and high cancellation rate [347]. Hyperinsulinemic PCOS women are more likely to produce lower quality oocytes that exhibit low fertilization rates after IVF-ET in association with lower implantation rates [348].
Improving insulin resistance with exercise, low-calorie diet and insulin-lowering drugs such as metformin, troglitazone and acarbose decrease insulin levels, correct the endocrine abnormalities induced by obesity and insulin resistance which may improve the outcome of infertility treatment [349–351]. This leads to the assumption that co-treatment with insulin-lowering drugs or weight reduction before and during down-regulation and ovarian stimulation.
Some studies did not find deleterious of insulin resistance on ovarian stimulation [352]. Metformin treatment has been reported to increase the number of mature oocytes retrieved from women with PCOS undergoing gonadotropins-stimulated IVF-ET and ICSI. A low dose of metformin of 500 mg twice daily was started on day 1 of the cycle prior to leuprolide acetate suppression, and continued to the day of the pregnancy test. Metformin treatment significantly increased the number of mature oocytes, fertilization rates, and number of embryos produced, but did not alter the total number of oocytes or peak E2 levels [353].
Later, the same authors reported that metformin therapy improves ovarian stimulation and IVF-ET outcomes in coasted patients with clomiphene-resistant PCOS [354]. No further published reports to date have examined this issue.
Corticosteroids
Successful use of corticosteroids in treatment of anovulatory infertility has been reported [355, 356]. Later corticosteroids were used as adjuvant therapy together with clomiphene citrate or gonadotropins in ovulation induction. The theoretical basis for this application has not been fully elucidated, but it has been postulated that corticosteroid treatment could improve ovulation through reduced influence of the adrenal androgens on follicular development [357]. There have been reports on improved ovulation and pregnancy rates with adjuvant corticosteroid treatment in anovulatory women with elevated androgens or androgens within the higher range [358, 359], as well as in normoandrogenic women [360]. Adding dexamethasone to gonadotropins in ovulation induction in women with normal serum concentrations of gonadotropins, androgens, and prolactin did not give an improved outcome in other studies [361].
Corticosteroids have also been used as adjuvant therapy in IVF-ET treatment. In 1986, Kemeter and Feichtinger [362] reported a significantly better pregnancy rate in a group of women with various infertility causes, except cycle abnormalities, undergoing IVF-ET with adjuvant prednisolone treatment, as compared to a group of women without adjuvant prednisolone. They stated that prednisolone would improve follicle maturation and thereby improve the pregnancy rate. In contrast, others [363] did not see any beneficial effects of adjuvant dexamethasone in a group of women with serum DHEA-S >2.5 μg/ml [6510 nmol/l] undergoing IVF-ET treatment, neither did Bider [361], in a group of women with tubal factor infertility after addition of dexamethasone.
In the above-mentioned study by Lobo [357], decreased serum concentrations of testosterone, unbound testosterone, and DHEA-S were noted after dexamethasone and clomiphene administration. In women who ovulated, testosterone and unbound testosterone increased again when clomiphene was added despite the continuation of dexamethasone. Decreased serum concentrations of DHEA-S and testosterone after ovarian stimulation with clomiphene citrate and human menopausal gonadotropins (HMG) and adjuvant prednisolone have been reported [362]. The other studies did not report data on androgen concentrations after glucocorticoid treatment. The present prospective, randomized, placebo-controlled study was performed to find out if adrenal suppression with prednisolone during ovarian stimulation before IVF-ET in a group of women with PCOS resulted in any changes in serum and follicular-fluid androgens. Clinical outcome variables, such as embryo quality, implantation rate, and clinical pregnancy rate were also noted.
Fridstrom [364] have reported that adjuvant glucocorticoids in ovarian stimulation before IVF-ET did not decrease the concentrations of adrenal androgens in serum and follicular fluid in PCOS. However, whether there were beneficial effects on ovum quality or implantation rate could not conclusively be determined in their study due to the small number of patients.
The addition of corticosteroids to ovarian stimulation protocols for assisted reproduction has been suggested in cases of recurrent pregnancy loss [365].
Progesterone antagonists: Receptor antagonist onapristone and mifeprisotone
Krusche found that after ovarian stimulation, the PR antagonist onapristone retarded endometrial transformation in the rabbit model. The authors concluded that since ovarian stimulation, used in human IVF-ET therapy, is frequently reported to cause an advancement of post-ovulatory endometrial development, a therapeutic application of PR antagonists to slow down such advanced endometrial transformation was suggested. Eventually, this modulation of advanced endometrial development may improve implantation rates [366].
4.2 Improving embryology techniques
Improving embryo techniques is a very important approach to improve the outcome of assisted reproduction treatment. This includes, improving culture conditions and extended culture till the blastocyst stage as well as improving the accuracy for selecting viable embryos for transfer. Another strategy employs pre-implantation genetic diagnosis (PGD). In addition there are other strategies to improve the embryo quality including oocyte donation and micromanipulation techniques (assisted hatching, and cytoplasmic and mitochondrial transfer) [367].
4.2.1 Improving embryo culture condition, selection and extended embryo culture (blastocyst transfer)
Over the past decade there has been considerable interest in optimizing culture media for supporting human embryos including reducing glucose concentrations [368], adding amino acids [369] and supplementing media with growth factors [370]. However, there have been no large prospective randomized studies on the effects of culture media on outcome after IVF-ET, apart from one, which reported that although glucose-free medium improved embryo quality, pregnancy rates were not increased [371]. All these improvements as well as the growing data on the composition of human female reproductive tract fluids and from embryo physiology studies has led to the proposition that extended embryo culture should take place till the blastocyst stage in more than a single medium formulation [372]. To this end, sequential media have been developed, tested extensively on animal models and subsequently used clinically [373].
One approach to increasing pregnancy rates is to improve the selection of viable embryos. Embryos are selected on the basis of morphology and rate of development, and the fastest developing embryos of the best morphology are selected for transfer. Although there is some correlation between morphology and blastocyst formation [374] and implantation [375], selection on the basis of morphology and rate of development remains an unsatisfactory and imprecise method of selecting viable embryos. It is impossible to identify visually with certainty, which embryos at early cleavage stages will subsequently arrest. One approach to improve the selection is the extended culture of the embryos to the blastocyst stage and transfer them on day 5 or 6, allowing the identification of developing embryos and the transfer of blastocysts, which are synchronous with the endometrium development, rather than cleavage stage embryos. It had been hoped also that, during extended culture, the chromosomally abnormal embryos so common at early cleavage stages [376] would arrest and fail to complete preimplantation development. However, although few human blastocysts have total chromosomal abnormality, a large number of blastocysts have various proportions of abnormal cells and are mosaic [377].
However, although many clinics have experienced increased success with extended culture, others have reported no benefit. So, it would be unwise to suggest that blastocyst culture and transfer represents a panacea for all clinics and all patients. On the contrary, transferring early stage embryos (zygot intrafallopian transfer) may help in repeated implantation failure rather than blastocyst transfer. [378].
4.2.2 Preimplantation genetic diagnosis
Preimplantation genetic diagnosis (PGD) allows the diagnosis of a genetic disorder in an embryo before its implantation in the uterus. PGD involves the removal of one or two blastomeres from an eight-cell stage embryo after IVF-ET, analysis of the blastomeres using fluorescent in situ-hybridization (FISH) or PCR, and identifying affected embryos. As only unaffected embryos are transferred back to the uterus after PGD, termination can be avoided. PGD is of benefit to couples at risk of passing on a genetic disease to their offspring as well as for couples with repeated unexplained IVF-ET failures. The transfer of unaffected embryos is believed to enhance the success rate of IVF-ET. Unfortunately, a large number of oocytes need to be retrieved for a successful PGD cycle, as not all will fertilize, cleave, undergo successful biopsy and be identified as suitable for transfer. Although most embryos (96%) survive biopsy, the pregnancy rates after transfer of biopsied embryos (16% per treatment cycle) [377] are less than those after conventional IVF-ET (23% per cycle) [379]. However, PGD is still in its early developmental stages [14].
4.2.3 In-Vitro Maturation of Oocytes
Immature oocytes can be aspirated from small follicles of > 2 mm in diameter. Resumption of meiosis in fully-grown oocytes, germinal vesicle breakdown, extrusion of the first polar body and acquisition of the ability to be fertilized may all occur during in vitro maturation (IVM). Although the first human birth after IVM was achieved 20 years ago [380], a few cases have been reported subsequently, including births after the aspiration and IVM of oocytes retrieved from women who have natural or partially stimulated cycles [381] and women with PCOS [382]. When oocytes are removed from small antral follicles and placed in culture, approximately 60% will have undergone nuclear maturation within 48 h. After exposure to spermatozoa, about 40% of oocytes will undergo normal fertilization, exhibiting two pronuclei and extruding the second polar body. Between 20 and 25% of fertilized oocytes will undergo cleavage [383]. However, pregnancy rates after the transfer of such embryos have been extremely low (1–2%) and most embryos arrest between the four- and eight-cell stages with only few encouraging results [384]. Several factors may influence the fertilization, cleavage, blastocyst and pregnancy rates achieved after IVM, including the hormonal environment in vivo or in vitro and the composition of the culture medium [385]. Further research in these areas will lead to greater understanding of oocyte maturation and should result in improved implantation and pregnancy rates [14].
4.2.4 Micromanipulation techniques
Intracytoplasmic sperm injection (ICSI)
The use of small numbers of spermatozoa during IVF-ET results in fertilization failure. In these cases, it is possible to inject a single spermatozoon into the ooplasm of a mature metaphase II oocyte, a procedure known as ICSI. The first successful pregnancies were in the early 1990s [10]. Since then, the technique has become a widely accepted treatment for couples with severe male factor infertility. Extending the indication of ICSI to include non-male factor infertility has been suggested to improve the fertilization and pregnancy rates. However, several studies have not supported the benefit of ICSI over conventional IVF-ET in improving the treatment outcome for non-male factor indications with growing consensus against applying ICSI for all cases undergoing IVF-ET [386–389].
Assisted hatching
To help embryos hatching from their zona pellucida during blastocyst expansion, different types of assisted hatching have been developed, including mechanical partial zona dissection or zona drilling, chemical zona drilling with acidic Tyrode's solution, and the laser technique [176]. However, there is controversy about the benefit of assisted hatching on the improvement of the implantation rate and pregnancy rate. Some investigators agreed as to the benefit of assisted hatching in women with advanced age, in women with repeated IVF-ET failure, and in the general population [177, 178].
Mitochondrial injection and cytoplasm, and nuclear transfer
Van Blerkom suggested that the developmental competence of mouse and human early embryos is related to the metabolic capacity of the mitochondria [144]. Deletions and mutations in oocyte mtDNA may lead to mitochondrial dysfunction, influencing energy production and apoptosis in oocytes and early embryos, resulting in aberrant chromosomal segregation or developmental arrest [144]. In mice, cytoplasmic transfer from young to old oocytes improved older oocytes quality [390].
Cytoplasmic transfer from metaphase II donor oocytes to mature recipient oocytes from women with recurrent IVF-ET failure has been carried out in an attempt to restore normal growth in developmentally compromised oocytes and embryos [391].
Nuclear transfer has also been proposed for the treatment of mitochondrial disease [392] whereby a karyoblast containing metaphase II chromosomes from women with repeated failures of embryo development due to defective oocyte cytoplasm is fused to enucleated donor oocytes. The increased cost and complexity as well as the invasiveness of the procedure with the decreased implantation rates all make these procedures far from being applied routinely to improve the treatment outcome after assisted reproduction. Only selected cases might be expected to benefit from these newly developing techniques. It should be noted that currently in the United States, performance of cytoplasmic or nuclear transfer in humans can only be conducted following approval of the protocol by the FDA.
PART TWO
1. Aromatase inhibition to improve outcome of treatment after ovarian stimulation
1.1 Introduction
We hypothesize that aromatase inhibitors can be used to improve the treatment outcome after ovarian stimulation either alone, or in combination with IUI and assisted reproductive technology. The use of aromatase inhibitors during ovarian stimulation may have several benefits including: (1) The enhancement of implantation by lowering the supraphysiological levels of estrogen attained during ovarian hyperstimulation that is believed to adversely affect the development of the endometrium, oocytes and embryo, as well as other possible targets. (2) Reducing gonadotropins dose required for achievement of optimum ovarian stimulation. This would reduce possible deleterious direct effects of exogenous gonadotropins injection in addition to reducing the cost of treatment. (3) Other possible benefits such as improvement of ovarian response to FSH stimulation in poor responders, and prevention of premature endogenous gonadotropin surge, as well as lower risk of severe ovarian hyperstimulation syndrome.
The improvement in implantation, as well as the reduced cost of treatment by decreasing the gonadotropins dose required for ovarian hyperstimulation would encourage the policy of transferring one embryo to minimize the risk of multiple gestation. This would have a tremendous economic impact on the practice of assisted reproduction, as well as cost of health care for multiple gestation worldwide.
1.2 Outline of the use of aromatase inhibitors for ovarian stimulation
In the following section, we present brief outline on the aromatase enzyme, estrogen biosynthesis and the development of aromatase inhibitors followed by a summary of the available data concerning the use of aromatase inhibitors for ovarian stimulation.
The aromatase enzyme
Aromatase is a microsomal member of the cytochrome P450 hemoprotein-containing enzyme complex superfamily (P450 arom, the product of the CYP19 gene) that catalyzes the rate-limiting step in the production of estrogens, that is, the conversion of androstenedione and testosterone via three hydroxylation steps to estrone and E2 respectively [393]. Aromatase activity is present in many tissues, such as ovaries, brain, adipose tissue, muscle, liver, breast tissue, and in malignant breast tumors. The main sources of circulating estrogens are the ovaries in premenopausal women and adipose tissue in postmenopausal women [394, 395].
The development of aromatase inhibitors
Aromatase is a good target for selective inhibition because estrogen production is a terminal step in the biosynthetic sequence. Several aromatase inhibitors have been utilized in clinical studies over the last 20 years. The most successful, third generation aromatase inhibitors are licensed for breast cancer treatment [396]. Aromatase inhibitors have been classified in a number of different ways, including first-, second-, and third-generation; steroidal and non-steroidal; reversible (ionic binding), and irreversible (suicide inhibitor, covalent binding) [397]. Table 4 lists the different classes of aromatase inhibitors.
Steroidal aromatase inhibitors are androstenedione analogues that act as a false substrate and bind irreversibly to the androgen-binding site of the enzyme [398]. Non-steroidal aromatase inhibitors exert their function through binding to the heme moiety of the cytochrome P450 enzyme [399]. The first of these inhibitors to be used clinically was aminoglutethimide, which induces a medical adrenalectomy by inhibiting many other enzymes involved in steroid biosynthesis [400]. Although aminoglutethimide is an effective hormonal agent in postmenopausal breast cancer, its use is complicated by the need for concurrent corticosteroid replacement, in addition to side effects like lethargy, rashes, nausea and fever that result in 8–15% of patients stopping treatment [401, 402]. The lack of specificity and unfavorable toxicity profile of aminoglutethimide led to the search for more specific aromatase inhibitors. In addition, the above mentioned aromatase inhibitors were not able to completely inhibit aromatase activity in premenopausal patients.
Third generation aromatase inhibitors
The third-generation anti-aromatase agents commercially available include two non-steroidal preparations, anastrozole and letrozole, and a steroidal agent, exemestane. Anastrozole and letrozole are often referred to as aromatase inhibitors, whereas exemestane is called an aromatase inactivator [403, 404]. Anastrozole, ZN 1033, (Arimidex®) and letrozole, CGS 20267, (Femara®) are selective aromatase inhibitors, available for clinical use in North America, Europe and other parts of the world for treatment of postmenopausal breast cancer. These triazole (antifungal) derivatives are reversible, competitive aromatase inhibitors, which are highly potent and selective. At doses of 1–5 mg/day, they inhibit estrogen levels by 97% to >99% resulting in estrogen concentrations below detection by most sensitive immunoassays. Table 5 shows the relative potencies of different aromatase inhibitors. Aromatase inhibitors are completely absorbed after oral administration, with mean terminal half-life (t1/2) of approximately 45 hours (range, 30–60 hours). They are cleared from the systemic circulation mainly by the liver. Gastrointestinal disturbances account for most of the adverse events, although these have seldom limited therapy. Other adverse effects are asthenia, hot flashes, headache, and back pain [406]. The average wholesale cost for a month's supply (thirty tablets) of aromatase inhibitors is about $150 to $250 [407]. Table 6 lists the clinical advantages of the third generation aromatase inhibitors as regards potential use for ovarian stimulation.
Hypotheses behind the use of aromatase inhibitors for ovarian stimulation
In the late 1990s, we explored the hypothesis that it might be possible to mimic the action of clomiphene citrate, without depletion of ER, by administration of an aromatase inhibitor in the early part of the menstrual cycle. We hypothesized that the result of blocking estrogen production would be release of the hypothalamic/pituitary axis from estrogenic negative feedback, thereby increasing gonadotropin secretion and resulting in stimulation of ovarian follicular development. This first hypothesis is referred to as CENTRAL hypothesis. The selective non-steroidal aromatase inhibitors have a relatively short half-life (approximately 40 hours) compared to clomiphene citrate, and would be ideal for this purpose since they are eliminated from the body rapidly [408, 409]. In addition, no adverse effects are expected on estrogen target tissues, since no ER down-regulation occurs as observed in clomiphene citrate treatment cycles.
We subsequently developed a second hypothesis, which was referred to as the PERIPHERAL hypothesis to explain another mechanism of action of the aromatase inhibitors in ovarian stimulation. We believe that aromatase inhibitors also act locally in the ovary to increase follicular sensitivity to FSH stimulation. This may result from the temporary accumulation of intraovarian androgens, since conversion of androgen substrate to estrogen is reversibly blocked by aromatase inhibition. There are data supporting a stimulatory role for androgens in early follicular growth in primates [410]. Testosterone was found to augment follicular FSH receptor expression in primates suggesting that androgens promote follicular growth and estrogen biosynthesis indirectly by amplifying FSH effects [411, 412]. In addition, androgen accumulation in the follicle may stimulate insulin-like growth factor I (IGF-I), along with other endocrine and paracrine factors, which may synergize with FSH to promote folliculogenesis [413–416]. This hypothesis still waits for evidence to support it.
1.2.1 Use of aromatase inhibitors alone for ovarian stimulation
In the last few years, we have worked on the development of aromatase inhibitors for ovarian stimulation and infertility management and reported interesting success in different applications as follows:
Induction of ovulation in anovulatory women
Based on our CENTRAL hypothesis of using an aromatase inhibitor for induction of ovulation (as explained above), the success of an aromatase inhibitor in inducing ovulation in patients with PCOS was reported [417–420].
Augmentation of ovulation in ovulatory women
After demonstrating success in inducing ovulation in anovulatory women, we proceeded to test whether aromatase inhibition might enhance the release of endogenous gonadotropins enough to stimulate the development of multiple follicles in ovulatory women and result in augmentation of ovulation or even controlled ovarian hyperstimulation. The use of an aromatase inhibitor for augmenting ovulation in patients with ovulatory infertility was successful in women with unexplained infertility, endometriosis, and women undergoing therapeutic donor insemination, and in ovulating partners of infertile men [419, 420].
Induction and augmentation of ovulation after clomiphene citrate failure
For over 40 years, clomiphene citrate has been the most commonly used treatment for the induction and augmentation of ovulation, accounting for about two thirds of the fertility drugs prescribed in the United States [421, 422]. In spite of the high ovulation rate associated with the use of clomiphene citrate, the pregnancy rate is much lower than anticipated. This is particularly true when considering pregnancy rate per cycle after three cycles of CC treatment [421], with higher than expected incidence of miscarriage in conception cycles [255]. Such discrepancy is believed, as explained earlier, to be due to the peripheral antiestrogenic effect of clomiphene citrate particularly at the level of the cervical mucus and endometrium, and which manifest themselves even in the presence of gonadotropin treatment superovulation. The accumulation in the body of the isomers of clomiphene citrate due to the long half-life (several days to weeks) adds to the persistence of the antiestrogenic effect [261, 262].
In order to improve the outcome of clomiphene citrate treatment, various approaches have been suggested to overcome the antiestrogenic effect including concomitant estrogen administration. Some investigators reported increased endometrial thickness and improved pregnancy rates with this approach [423–425] while others have reported no benefit [426] or even a deleterious effect of estrogen administration [427]. Another approach was to administer clomiphene citrate earlier during the menstrual cycle [428], to allow the anti-estrogenic effect to wear off earlier before the critical period of fertilization and implantation. A third approach has been to combine another selective ER modulator like tamoxifen, which has more estrogen agonistic effect on the endometrium with clomiphene citrate [429] or to use it as an alternative to clomiphene citrate [430]. However, none of these strategies have proved to be completely successful in avoiding the peripheral antiestrogenic effects of clomiphene citrate. In addition to a discrepancy between ovulation and pregnancy rates with clomiphene citrate treatment, 20% to 25% of anovulatory women are resistant to clomiphene citrate and fail to ovulate at doses up to 200 mg daily.
The success of aromatase inhibition in inducing and augmenting ovulation encouraged trying aromatase inhibitors in cases of clomiphene citrate failure that was found to be successful in achieving ovulation and pregnancy [419, 420]. As explained above, the significantly shorter half-life of the third generation aromatase inhibitors compared to clomiphene citrate, allows rapid elimination from the body [408, 409]. In addition, since no ER down-regulation occurs, any adverse effects on estrogen target tissues, as observed in clomiphene citrate treated cycles, is expected.
1.2.2 Adjunct use of aromatase inhibitors for ovarian stimulation
We investigated the idea of combining an aromatase inhibitor with FSH injections to reduce the dose of FSH required to achieve optimum controlled ovarian hyperstimulation, without adverse antiestrogenic effects [431, 432]. A significant reduction in the FSH dose required (from 45% to 55% in women with PCOS and unexplained infertility) has been reported [433] without evidence of peripheral antiestrogenic effects [432, 433].
Improving ovarian response to FSH stimulation in poor responders
Reducing FSH dose required for optimum controlled ovarian hyperstimulation encouraged us to explore the use of an aromatase inhibitor in conjunction with FSH to improve response to ovarian stimulation in poor responders. In a selected group of women who had a poor response to FSH stimulation in at least two prior treatment cycles, adding an aromatase inhibitor resulted in improvement in the response to FSH stimulation. All patients developed a significantly greater number of mature ovarian follicles and almost a third of the treatment cycles resulted in pregnancy. In addition, the dose of FSH required to achieve such optimum response was significantly less than the dose used in the prior cycles in which FSH was used alone [434, 435].
Developing alternative regimens for administering aromatase inhibitors for ovarian stimulation
During all the above-mentioned studies, the aromatase inhibitor, letrozole, was administered orally as a daily dose of 2.5 mg from day 3 to day 7 of the menstrual cycle. Based on the pharmacokinetics of the new aromatase inhibitors (almost 100% bioavailability after oral administration, ~2 days half-life and no accumulation or significant metabolite accumulation), we thought of a potentially more convenient method of administering an aromatase inhibitor for ovulation induction. We hypothesized that significantly higher concentration of the aromatase inhibitor can be achieved early in the menstrual cycle with faster clearance later in the menstrual cycle with the administration of a high single dose of the aromatase inhibitor early in the menstrual cycle such as on day 3. A single dose regimen would satisfy two goals: first, achieving maximum estrogen suppression early in the menstrual cycle when it is desired, and second, to allow clearance of the aromatase inhibitor before the critical final stage of fertilization and embryogenesis, to maximize safety and avoid any possible undesirable effects of the aromatase inhibitors. Single dose administration of an aromatase inhibitor was found successful in inducing ovulation with ovulation and pregnancy rates comparable to the 5-day regimen [436].
Limitations of preliminary data on use of aromatase inhibitors for ovarian stimulation
In all our clinical trials described above, results were not obtained from randomized, prospective, placebo controlled studies, the optimum research design. However, due to the experimental nature of the use of the aromatase inhibitors for ovarian stimulation, which to our knowledge, has never been used before, we believed that the present observational trials were mandatory before proceeding into any definitive randomized studies.
The encouraging results of our data have led other investigators from different centers world-wide to study the use of aromatase inhibitors for ovarian stimulation and in general have reported findings similar to ours in terms of the success of aromatase inhibitors in infertility treatment [437–450].
In a randomized, prospective study, a superior uterine environment has been found in patients treated with an aromatase inhibitor compared with clomiphene citrate, reflecting the lack of the antiestrogenic effects with aromatase inhibitor treatment. Although a non-significant increase in pregnancy was observed in patients who received aromatase inhibitor treatment (16.7 versus 5.6% per patient, P = 0.55), this almost three-fold increase in pregnancy rate would have required about ten more patients per group to reach statistical significance. The superiority of a single dose administration of an aromatase inhibitor was also reported when compared to clomiphene citrate [439]. In other studies, the use of an aromatase inhibitor in conjunction with FSH was found to reduce the FSH dose [440] and improve response to ovarian stimulation in poor responders [441, 442], confirming our previous reports.
In another prospective randomized trial, Biljan [438] studied two doses (2.5 and 5 mg per day) of the aromatase inhibitor, letrozole for ovarian stimulation in patients with unexplained infertility. They found patients treated with a higher dose of letrozole developed more follicles without a detrimental effect on endometrial development. The potential for different doses and regimens of administration of aromatase inhibitors for ovarian stimulation still requires a lot of study, but the use of aromatase inhibitors at high doses should be cautiously considered.
In all reports and most of studies of other investigators, the aromatase inhibitor, letrozole was the one used. However, anastrozole, another third generation aromatase inhibitor similar to letrozole, was used in other studies [443, 444]. Due to the similar pharmacokinetics and pharmacodynamics, including similar potencies and specificity in inhibiting the aromatase enzyme, we believe there is likely to be no difference between the third generation aromatase inhibitors in their efficacy for ovarian stimulation. It does, however, need to be determined what dose of anastrozole is required to be equivalent to letrozole.
Future avenues for the use of aromatase inhibitors for infertility management
It seems that there are many interesting areas of research that need exploration as regards the development of aromatase inhibitors for infertility management. These directions for research would include: confirming the available preliminary data on the success of aromatase inhibition in induction and augmentation of ovulation, as well as reducing the dose of FSH needed for ovarian stimulation, improving response in poor responders, and finding the optimum regimen for administering aromatase inhibitors for infertility treatment
Moreover, the use of aromatase inhibitors for new applications including in-vitro maturation and prevention of severe ovarian hyperstimulation syndrome and endometriosis-related infertility are interesting future avenues for aromatase inhibitors potential use in infertility management.
In addition the use of aromatase inhibitors to improve the outcome of treatment after assisted reproduction as discussed earlier in this review is an exciting area of application.
1.3 Use of aromatase inhibitors to improve treatment outcome after ovarian stimulation and assisted reproduction
In addition to using aromatase inhibitors, ALONE, for ovarian stimulation, their use during assisted reproduction carries several potentials to improve the treatment outcome.
Improving implantation rates
We hypothesize that aromatase inhibition during assisted reproduction may improve the implantation rate mainly by reducing the estrogen levels attained during COH. In addition, two other mechanisms may apply including reducing the dose of FSH required for optimum COH as well as applying much simpler stimulation protocols that do not require the use of GnRH analogues, hence avoiding any possible direct deleterious effects of FSH and GnRH analogues on the endometrium.
1.3.1 Lowering supraphysiological estrogen levels
As discussed above, the undesirable effects of ovarian stimulation on the outcome of infertility treatment are believed to be due to the supraphysiological levels of estrogen irrespective to whatever mechanisms explain for that (various postulated mechanisms were discussed above). So, lowering estrogen levels may be associated with improved outcome of treatment in terms of improving the implantation and pregnancy rates in addition to lowering risk of sever ovarian hyperstimulation syndrome.
Reducing estrogen synthesis by aromatase inhibition seems to be an exciting idea to ameliorate the deleterious effects of the supraphysiological levels of estrogen on the endometrium, the developing oocyte and embryo as well as the luteal.
Until recently there was no suitable aromatase inhibitor that could be used clinically to reduce estrogen levels during ovarian stimulation. This is because the available aromatase inhibitors were not safe for clinical application during ovarian stimulation due to lack of specificity in inhibiting the aromatase enzyme without inhibiting other steroidogenesis enzymes (e.g. aminoglutethimide). The other aromatase inhibitors (steroidal androstenedione analogues) were irreversible in their effect on the aromatase enzyme besides being parentrally administered. Most important, these old aromatase inhibitors were not potent enough to inhibit the aromatase enzyme and lower estrogen levels in women of the reproductive age group. However, the third generation non-steroidal aromatase inhibitors group is very potent and specific in inhibiting the aromatase enzyme reversibly. These new aromatase inhibitors are orally administered with very high safety profile and well tolerability. Moreover, they are cheap with a relatively short half-life [~45 hours], and already approved for clinical use to reduce estrogen production in postmenopausal women with breast cancer.
They have not been used in women of the reproductive age group. However, we have found these aromatase inhibitors to be effective in inhibiting the aromatase enzyme and effectively decrease estrogen levels in women of the reproductive age group during their successful use for ovarian stimulation [417, 418, 431, 432].
In our experience with the use of an aromatase inhibitor for ovarian stimulation, estrogen levels were significantly lower (especially E2 level/mature follicle) when compared with conventional stimulation protocols (clomiphene citrate, FSH and clomiphene citrate plus FSH). Such low E2 levels may be beneficial and explains at least partially the improved outcome of treatment in terms of achieving promising high pregnancy rates during aromatase inhibitor treatment.
It may not be only the supraphysiological estrogen levels, which explain totally for the reduced implantation rate during ovarian stimulation cycles. Local paracrine factors, or possibly other undetermined factors may be responsible for the reduced implantation rate during COH. In this case the aromatase inhibitors may not offer a complete solution to overcome such drawbacks of ovarian stimulation. However, the idea is exciting and seems to be promising and warrant trials because high estrogen levels may explain, at least in part, for the deleterious effects of ovarian stimulation on treatment outcome.
In ddition, we think that, reducing estrogen levels achieved during induction of ovulation by using aromatase inhibitors may prevent the significant increase in leptin concentrations avoiding its possible deleterious effects on the outcome of treatment as explained above. Because elucidation of leptin's specific role in reproductive function has been challenging, with conflicting results reported by various investigators, it is still quite early and highly speculative to hypothesize a link between aromatase inhibitors use and the role of leptin during induction of ovulation. However, the available strong data about a possible role of leptin in mediating reproductive disorders especially in obese women and the firm findings of a regulatory effect of estrogen on leptin production make this hypothesis exciting and interesting enough to warrant future investigation.
1.3.2 Reducing gonadotropins requirements for optimum ovarian stimulation
As mentioned earlier, the use of an aromatase inhibitor significantly reduces the dose of FSH required for optimum COH. In addition to the significant economic benefit, we believe that reducing the dose of FSH may improve the treatment outcome of ovarian stimulation by reducing any possible deleterious effect of the exogenously administered FSH on the endometrium, developing oocyte or other targets.
Other possible benefits
It is believed that the endogenous LH surge arises once serum estrogen levels surpass a set threshold for a certain period of time [451–453].
The supraphysiological estrogen levels attained during ovarian stimulation are believed to cause premature release of the endogenous LH surge resulting in cycle cancellation during assisted reproduction. For that reason, GnRH analogues have been the standard of practice during most of the stimulation protocols for assisted reproduction to prevent the occurrence of endogenous LH surge either by direct inhibition of LH surge (GnRH antagonists) or by down-regulation of the GnRH receptors at the pituitary levels resulting in pituitary desensitization and prevention of the release of endogenous LH [454, 455].
Unfortunately, the use of GnRH analogues (which is a crucial part of the stimulation protocol to prevent cycle cancellation as explained above), is associated with several problems including increasing the dose of FSH required to achieve optimum COH (due to suppressing the endogenous gonadotropin secretion, and possible peripheral effect at the level of the ovaries), as well as the luteal phase defect due to a dysfunctional corpus luteum function secondary to persistent endogenous LH suppression as explained earlier. In addition, there is rising evidence of a possible direct deleterious effect of the GnRH analogues, especially the antagonist at the level of the endometrium [456].
The use of an aromatase inhibitor to reduce estrogen levels attained during COH may be effective in preventing the occurrence of premature ovarian surge. This would avoid the use of GnRH analogues during stimulation protocols for assisted reproduction which has several advantages including prevention of the possible deleterious effects of these agents as mentioned above in addition to reducing the cost of treatment as well allowing implementing much simpler stimulation protocols during assisted reproduction.
Aromatase inhibitors for endometriosis-related infertility
The expression of aromatase enzyme in endometriotic tissues with the significant role of locally produced estrogen in endometriosis progression [457] suggests a benefit of aromatase inhibitors in endometriosis-related infertility. The inhibition of local estrogen production in endometrial implants, and the lower peripheral estrogen levels associated with the use of aromatase inhibition for ovulation induction, is expected to protect, to some degree, against progression of endometriosis during infertility treatment.
1.4 Concerns regarding the use of aromatase inhibitors in infertility treatment
There are three main concerns that may arise regarding the use of the aromatase inhibitors to improve the outcome of treatment after ovarian stimulation and assisted reproduction. They include the possible deleterious effect of the low follicular estrogen milieu on the development of the oocytes, the possible direct effect of the aromatase inhibitors on the oocyte development, fertilization or embryogenesis, and the accumulation of the androgens that may result from inhibition of their conversion into estrogens.
1.4.1 Effect of low estrogen milieu on the developing oocyte
Palter et al. have reviewed the question whether an estrogen-free (or at least very low estrogen) intrafollicular environment is compatible with follicular development, ovulation, and corpus luteum formation [458]. The authors concluded that markedly reduced to nonexistent intrafollicular and circulating concentrations of estrogen are compatible with follicular "expansion", retrievable and fertilizable oocytes, as well as with cleavable and apparently transferable embryos. The authors drew their conclusion after discussing lessons from data in the literature including cases of deficiency of the 17-hydroxylase/17–20 lyase [459–466] and 3β-hydroxysteroid dehydrogenase [467, 468] and the aromatase enzyme [469–473] as well as cases of severe hypogonadotropism [474–478]. In addition, data on the use of aromatase inhibitors in animals were reviewed [479–482].
Cases of enzyme deficiency leading to a status of very low estrogen levels
17 alpha-hydroxylase/17–20 lyase deficiency
17alpha-hydroxylase/17–20 lyase deficiency is one form of congenital adrenal hyperplasia, which is associated with marked impairment of glucocorticoid, androgen, and estrogen biosynthesis [459]. Women suffering from this enzyme deficiency suffer from hypergonadotropic hypogonadism and sexual infantilism. However, early reports noted the presence of many primary and secondary follicles in ovarian material [460] and many of them had bilateral multicystic ovaries at the time of laparotomy [461–463].
It is interesting that Rabinovici reported on a patient afflicted with virtually complete 17-hydroxylase/17–20 lyase deficiency that, despite castrate levels of estrogens, underwent an apparently successful induction of ovulation associated with progressive follicular expansion and oocyte retrieval, IVF-ET, and early embryonic cleavage followed suit [464].
3β-hydroxysteroid dehydrogenase [3β-HSD] deficiency
The deficiency of 3β-HSD is associated with markedly reduced levels of estrogen. However, the existence of normal ovulatory function in a woman with late-onset of a mild form of 3β-HSD has been reported [483].
In a group of cycling female Rhesus monkeys exposed to ovulation induction with hFSH and hFSH + hLH in the absence or presence of the 3β-HSD inhibitor trilostane given on days 1–8 of the menstrual cycle [467], apparently healthy oocytes were obtained by follicular aspiration 34 h after hCG administration. Importantly, treatment with the 3β-HSD inhibitor, trilostane, led to a reduction in serum E2 levels to 7% of that of control animals throughout the follicular phase. Despite this dramatic reduction in E2 levels, neither the total number of large antral follicles per animal (17 ± 1 vs. 18 ± 2) nor their size distribution differed significantly from 3β-HSD inhibitor-untreated controls. Furthermore, treatment with the 3β-HSD inhibitor did not alter the overall maturation pattern of collected oocytes (atretic, prophase I, metaphase I, or metaphase II). However, the authors found a reduction in the percentage of metaphase II oocytes that were successfully fertilized (15 vs. 65%). Moreover, metaphase oocytes that required more than 8 h to complete meiosis in vitro failed to fertilize in three of four animals receiving 3β-HSD inhibitor relative to controls (31%). These observations suggest that follicular development and the completion of meiosis may be unaffected by the low estrogen levels but that cytoplasmic oocyte maturation and/or function could be unfavorably affected [467].
Aromatase enzyme deficiency and the use of aromatase inhibitors
Obviously, aromatase (estrogen synthase) enzyme deficiency is associated with marked decrease or almost absence of estrogen production.
Extreme examples of complete aromatase deficiency due to mutations in the aromatase gene, CYP19 gene, in adult human females, however, were reported [469, 470]. The affected patients suffered from ambiguous external genitalia, primary amenorrhea, sexual infantilism, and multicystic ovaries.
Morishima [471] reported on the aromatase deficiency in a 28-yr-old 46 XX proband followed since infancy. Null mutant mice for aromatase gene [ArKO] were generated [484], thereby affording the opportunity to examine the role of estrogen in the follicular development in the mouse ovary. Evaluation of the ovaries revealed the presence of many large follicles filled with granulosa cells and evidence of antrum formation, but no corpora lutea. The ovarian phenotype degenerated with age upon the appearance of hemorrhagic cystic follicles and the loss of secondary and antral follicles coincident with the infiltration of macrophages and with stromal hyperplasia [485–488]. Therefore, the ArKO females are infertile, due primarily to a complete lack of ovulation.
However, recently, Jones et al., reported that oocytes were harvested from the ovaries of 4- and 7-week old ArKO, wild type and heterozygote mice stimulated with 5 IU PMSG. The number of immature oocytes harvested from ArKO females did not differ from the number collected from wild type or heterozygote littermates of either age group. Oocyte in vitro maturation rates also did not differ between the three genotypes or two age groups, with almost 75% of the immature oocytes progressing to metaphase II. Chromatin staining confirmed the arrest of these oocytes at the second meiotic division with chromatin staining clearly present in the oocyte and polar body. Mature oocytes were inseminated and IVF-ET rates did not differ between the three genotypes or two age groups, with fertilization occurring in approximately 67% of oocytes. Fertilized oocytes were cultured to blastocysts. Again, blastocyst development rates did not differ between the groups, with approximately 65% of the zygotes developing into blastocysts. Blastocyst morphology was similar across all of the groups [489].
These results indicate that ArKO oocytes are competent to develop to at least the implantation stage. The authors concluded that estrogen might not be required for the production and maturation of developmentally competent oocytes. Rather, its role in folliculogenesis is probably via regulation of the hypophysial pituitary gonadal axis and thus gonadotropin secretion [489].
Regarding the effect of an estrogen-free/poor intrafollicular environment on gametogenic maturation, Palter et al. [458] concluded that the effect is a negative one. Their conclusion was based on a number of primate studies, which indicated that an estrogen-free/poor intrafollicular environment is associated with marked decrements in the rates of meiotic maturation and fertilization [467, 490]. In addition data derived from rodent models suggested that further suggested a compromise in early embryonic development [491].
In cycling female rhesus monkeys, the aromatase inhibitor, 1,4,6-androstatrien-3, 17-dione (ATD) was used to inhibit estrogen production during gonadotropin ovarian stimulation [479]. Animals treated with ATD displayed a drastic reduction in serum 17β-E2 levels to 37% of that of controls within 8 h of ATD treatment and to 16% of control by the day of hCG injection. In turn, the circulating levels of androstenedione rose. Despite the drastic reduction in the circulating levels of E2 and the increase in the circulating levels of androgens, the overall number of large antral follicles [16 ± 3 for controls and 20 ± 3 for ATD-treated] and their size distribution (as assessed by ultrasonography) proved comparable for control and ATD-treated animals. Similarly, no difference was noted in the number of oocytes collected or in the proportion of oocytes reinitiating meiosis (MI at the time of collection). In contrast, ATD-treated animals displayed a marked increase (31 vs. 11%) in the proportion of prophase I oocytes. Moreover, ATD-treated oocytes displayed retarded in vivo completion of maturation to MII (4% vs. 26%). Interestingly, the latter retardation was not observed in vitro. Furthermore, two of the four ATD-treated animals yielded oocytes that were morphologically abnormal. Finally, oocytes from ATD-treated animals displayed significantly reduced rates of fertilization (9% vs. 25%) as compared with controls. However, the cleavage rate after successful fertilization was similar for ATD-treated vs. ATD-untreated controls. In vitro cultures of granulosa cells collected at the time of oocyte aspiration revealed equivalent 24-h progesterone production in treated and control animals. The authors concluded that these findings suggest that the acute reduction in E2 levels during the terminal stage of gonadotropin stimulation had little effect upon follicular recruitment and expansion but an apparent detrimental effect upon gametogenic function may, in fact, exist. However, we cannot extrapolate from this study that the findings are simply due to the acute reduction of estrogen production as it is important to realize that there was a concomitant significant rise in androstenedione. Moreover, the time of administering an aromatase inhibitor during the latter part of the follicular phase as well as the irreversible nature of aromatase inhibition constitute important differences from our model of using a REVERSIBLE aromatase inhibitor, TEMPORARILY, EARLY in the menstrual cycle that has a SHORT half-life.
Selvaraj [480, 481] and Shetty [482] examine the effects of blocking estrogen biosynthesis during the follicular phase on follicular maturation in the adult female bonnet monkey. In their studies, they used one of the third-generation REVERSIBLE, aromatase inhibitors (CGS 16949A) starting EARLY on day 3 of the menstrual cycle. There were 53% and 70% reductions in the basal and surge levels of E2, respectively without obvious effect on follicular maturation, ovulation, and luteal function as assessed by serum hormone profiles as well as by laparotomy. Moreover, the concurrent administration of FSH and an aromatase inhibitor resulted in the suppression of the FSH-induced increase in the circulating levels of E2 (by 100%) with no effect noted on either the number of follicles developed or their size relative to control. In addition, granulosa and theca cells, removed on day 9 of the treatment cycle, were responsive to gonadotropins in vitro, disclosing no evidence of a deleterious effect on the cellular development and maturation of follicular cells.
Terry studied a possible role of high E2 levels in mediating the adverse effects of hyperstimulation with PMSG on early embryonic development in the rat [491]. They used the aromatase inhibitor, 4-hydroxyandrostenedione (4-OHA), to inhibit endogenous E2 production. The authors conducted three experiments. In the first, varying doses of 4-OHA were administered either concurrently with hCG to pro-estrus female rats hyperstimulated at early diestrus stage with 20 IU PMSG or alone into non-hyperstimulated pro-oestrus females. At high doses of 1000, 2000, or 5000 mg/rat, 4-OHA substantially improved the survival of embryos in hyperstimulated females with optimum protection at 2000 mg, while low doses of 100 and 500 mg/rat were ineffective. When administered alone, only the highest dose of 5000 mg/rat 4-OHA increased embryo count. In the second experiment, higher doses of PMSG were studied (30 or 40 IU), with or without 5000 mg/rat 4-OHA given at the time of hCG injection. PMSG proved to be more detrimental with increasing dose, and high doses of 4-OHA (5000 mg/rat) was needed to rescue embryos from death in the 30, but not 40, PMSG group. In the third experiment, the influence of the timing of 4-OHA treatment on its ability to improve the embryo count in hyperstimulated females was examined by introducing 4-OHA 24 h earlier, rather than at the time of hCG treatment. The results showed the importance of timing of 4-OHA administration, as 5000 mg/rat 4-OHA was able to restore embryo survival in the 40 PMSG hyperstimulated group only when it was administered 24 h before hCG injection. These results highlighted that 4-OHA, when administered at the appropriate time and dose, could reverse the negative effects of hyperstimulation from PMSG on early embryonic development. The authors concluded that this might be due to the suppression of estrogen production, thereby alleviating the supraphysiological level of E2, which is typically present in PMSG-treated females, which supports the hypothesis that excessive E2 is responsible for the negative effects of hyperstimulation with PMSG on early embryonic development.
The high success rates of ovulation and achievement of pregnancy in our reports on the use of aromatase inhibitor in ovarian stimulation, despite significantly lower estrogen levels could be due to several reasons. First: We used one of the third-generation "REVERSIBLE" aromatase inhibitors. Second: The aromatase inhibitor was administered EARLY in the follicular phase and for a limited period of time, which would allow the rapid clearance of the aromatase inhibitor from the body due to its SHORT half-life. Third: the reversible nature of the aromatase enzyme inhibition and the rising levels of FSH, which induce the expression of the aromatase enzyme, both would not allow for the estrogen levels to drop drastically below the low physiologic range. We believe that these low estrogen levels attained during the use of aromatase inhibitors for ovarian stimulation are compatible with healthy development of the oocytes, fertilization, embryo development, implantation and achievement of pregnancy. Fourth: the absence of significant rise in androgen levels would have prevented any possible unwanted effects on the terminal part of the oocyte maturation and ovulation.
1.4.2 Direct effect of the aromatase inhibitors on the developing oocyte, fertilization and embryogenesis
As explained above, the short half-life of the aromatase inhibitors and limiting the administration to the early part of the follicular phase would allow the rapid clearance of the medications from the body before the important stage of fertilization and embryogenesis. This in addition to the absence of accumulation of the aromatase inhibitors of any of their metabolites would make them safe for the ovarian stimulation. We have reported a preliminary data on the pregnancy outcome after the use of aromatase inhibitors for ovarian stimulation supporting the safety of using these medications for such indication [492].
Data have been accumulating regarding the absence of deleterious effects in association with aromatase inhibitor treatment on follicle/oocyte maturation and embryo development in mice [492, 493]. Treatment with the aromatase inhibitor, anastrozole was associated with similar number of total follicles found per animal (30.4 follicles/animal in the control group and 27 follicles/animal in the anastrozole-treated group) as well as comparable rates of development of embryos, morulae, blastocysts, and hatching blastocysts between the two groups (P = 0.20, 0.10, 0.44, and 0.38, respectively) [493].
1.4.3 Effect of the accumulated androgens
As mentioned before, the rapid clearance of the aromatase inhibitors from the body due to their short half-life and the reversible nature of the aromatase enzyme inhibition as well as the rising levels of FSH, which induce the expression of the aromatase enzyme, all would not allow the clearance of any accumulating androgens by converting them to estrogen. The conversion of the androgens into estrogens (the step catalyzed by the aromatase enzyme) is a terminal step in the cascade of steroidogenesis. So, substrate (androgens) accumulation is not expected to be very significant, as other alternative earlier steps in the steroidogenesis pathways will work to clear out the accumulating androgens. For these reasons, pharmacokinetic studies on the new aromatase inhibitors reported the absence of significant androgen elevations or abnormal changes in other steroids in patients receiving aromatase inhibitors for breast cancer. In a subset of our patients who received an aromatase inhibitor for ovarian stimulation, we did not find significant change in the androgen levels while receiving the medication when compared to stimulation with gonadotropins alone (unpublished data).
Most recently we published data on the favorable pregnancy outcome after the use of letrozole for ovarian stimulation alone or in conjunction with gonadotropins [494]. This study looked at outcome of pregnancies achieved after ovarian stimulation with the aromatase inhibitor, letrozole, alone or in conjunction with gonadotropins. The study included a cohort of 394 pregnancy cycles achieved after letrozole and other ovarian stimulation treatments in a ddition to a control group of pregnancies spontaneously conceived without ovarian stimulation. These 394 pregnancy cycles were achieved as follows: 63 pregnancies with 2.5 mg of letrozole alone or with gonadotropins, 70 pregnancies with 5.0 mg of letrozole, 113 pregnancies with clomiphene alone or with gonadotropins, 110 pregnancies with gonadotropins alone, and 38 pregnancies achieved without ovarian stimulation. The study found pregnancies conceived after letrozole treatments were associated with similar miscarriage and ectopic pregnancy rates compared with all other groups. In addition, letrozole use was associated with a significantly lower rate of multiple gestation compared with clomiphene citrate. The findings of the stuy support the favorable pregnancy outcome and low multiple gestation rate in association with the use of aromatase inhibitors for ovarian stimulation [494].
5 Summary and conclusion
Ovarian stimulation particularly in conjunction with assisted reproductive technologies aims at stimulating the recruitment of several mature ovarian follicles that would enhance the chance of successful treatment by obtaining several embryos readily available for transfer into the uterus. It is obvious that supraphysiological levels of estrogen are inevitably attained during ovarian stimulation due to the significant contribution of estrogen production from each one of the several mature ovarian follicles. There is growing evidence that such supraphysiological estrogen levels are deleterious on the development of the endometrium, oocytes, embryos as well as other targets through different mechanisms. This is believed to explain, at least in part, the low pregnancy rates associated with assisted reproduction, mainly as a result of the persistently low implantation rates that have not improved impressively along almost three decades of assisted reproduction experience. This led several investigators to adopt the concept of minimal ovarian stimulation for assisted reproduction as an alternative approach to reduce the deleterious effects of the supraphysiological estrogen levels attained during routine controlled ovarian hyperstimulation. Unfortunately such approach, despite being logical, is associated with the major drawback of achieving fewer oocytes than desired, which, expectedly, reduces the chances of treatment success. Recently, we reported the success of the new generation aromatase inhibitors in ovarian stimulation. We found these agents to be effective in reducing the amount of estrogen production by mature ovarian follicles significantly. Furthermore, we found the use of these agents to be associated with improved ovarian response to stimulation by gonadotropins, resulting in significant reduction in the dose of gonadotropins required for controlled ovarian hyperstimulation. The novel idea of using aromatase inhibitors during ovarian stimulation for assisted reproduction combines the benefit of a significant reduction in estrogen production as well as gonadotropins dose, with the exiting advantage of achieving a good number of mature oocytes as a result of enhanced ovarian response to gonadotropins. This is expected to improve the various aspects of treatment outcomes after assisted reproduction including increased safety, reduced cost, as well as enhanced implantation rate. If confirmed in well-designed clinical trials, this would facilitate acceptance of the concept of single embryo transfer by infertile couples and practitioners, thereby reducing the epidemic of multiple births after assisted reproduction with its significant deleterious health and economical effects. This is particularly true in the light of recent data supporting the success of the approach of single embryo transfer [495–497].
References
Templeton A: Infertility-epidemiology, aetiology and effective management. Health Bull Edinb. 1995, 535: 294-298.
Whiteford LM, Gonzalez L: Stigma: the hidden burden of infertility. Soc Sci Med. 1995, 27: 2013-2036.
Farley TMM, Belsey FH: The prevalence and etiology of infertility. Biological Components of Fertility. Proceedings of the African Population Conference, Dakar, Senegal, International Union for the Scientific Study of Population, Laege, Belgium l. 1988, 1: 2.1.15-2.1.30.
American Society for Reproductive Medicine: Guidelines for the provision of infertility service. 1996, ASRM, Birmingham AL
Speroff L, Glass RH, Kase NG: Female infertility. Clinical gynecologic endocrinology and infertility. 1994, Williams & Wilkins, Baltimore, 809-840.
Leung CK: Recent advances in clinical aspects of in vitro fertilisation. Hong Kong Med J. 2000, 62: 169-176.
Edwards RG, Bavister BD, Steptoe PC: Early stages of fertilization in vitroof human oocytes matured in vitro. Nature. 1969, 221: 632-635.
Steptoe PC, Edwards RG: Birth after the reimplantation of a human embryo. Lancet. 1978, 2: 366-
Mohr LR, Trounson AO: Cryopreservation of human embryos. Ann N Y Ac Sc. 1985, 442: 536-543.
Palermo G, Joris H, Devroey P, Van Steirteghem AC: Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet. 1992, 340: 17-18.
Handyside AH, Kontogianni EH, Hardy K, Winston RM: Pregnancies from biopsied human preimplantation embryos sexed by Y-specific DNA amplification. Nature. 1990, 344: 768-770.
CDC: Asstsed Reproductive Technology Success Rates. 1990, [http://www.cdc.gov]
Gardner DK, Lane M, Schoolcraft WB: Culture systems and blastocyst development. 2000, Serono Symposia USA, International Symposium on ART and Human Blastocyst, 2: 14-
Hardy K, Wright C, Rice S, Tachataki M, Roberts R, Morgan D, Spanos S, Taylor D: Future developments in assisted reproduction. Hum Reprod. 2002, 1232: 171-183.
Fanchin R: Assessing uterine receptivity in 2001: ultrasonographic glances at the new millennium. Ann N Y Acad Sci. 2001, 943: 185-202.
Vanderhyden B, Rouleau A, Walton E, Armstrong D: Increased mortality during early embryonic development after in-vitro fertilization of rat oocytes. J Reprod Fertil. 1986, 77: 401-409.
Fossum G, Davidson A, Paulson R: Ovarian hyperstimulation inhibits embryo implantation in the mouse. J In Vitro Fertil. Embryo Transfer. 1989, 6: 7-10.
De Boer P, van der Hoeven F, Wolters E, Mattheij J: Embryo loss, blastomere development and chromosomal constitution after human chorionic gonadotropin-induced ovulation in mice and rats with regular cycles. Gynecol Obstet Invest. 1991, 32: 200-205.
Ertzeid G, Storeng R, Lyberg T: Treatment with gonadotropins impaired implantation and fetal development in mice. J Assist Reprod Genet. 1993, 10: 286-291.
Vogel R, Spielmann H: Genotoxic and embryotoxic effects of gonadotropin-hyperstimulated ovulation of murine oocytes and preimplantation embryos, and term fetuses. Reprod Toxicol. 1992, 6: 329-333.
McKirnan SH, Bavister BD: Gonadotrophin stimulation of donor females decreases post-implantation viability of cultured one-cell hamster embryos. Hum Reprod. 1998, 13: 724-729.
Ma S, Kalousek DK, Yuen BH, Moon YS: Investigation of effects of pregnant mare serum gonadotropin PMSG on the chromosomal complement of CD-1 mouse embryos. J Assist Reprod Genet. 1997, 14: 162-169.
Spielmann H, Vogel R: Genotoxic and embryotoxic effects of gonadotropin hyperstimulated ovulation on murine oocytes, preimplantation embryos and term fetuses. Ann Ist Super Sanita. 1993, 29: 35-39.
Sauer MV, Paulson RJ: Oocyte and embryo donation. Curr Opin Obstet Gynecol. 1995, 7 (3): 193-198.
Klein J, Sauer MV: Oocyte donation. Best Pract Res Clin Obstet Gynaecol. 2002, 163: 277-291.
Tarlatzis BC, Pados G: Oocyte donation: clinical and practical aspects. Mol Cell Endocrinol. 2000, 1611-2: 99-102.
Check JH, O'Shaughnessy A, Lurie D, Fisher C, Adelson HG: Evaluation of the mechanism for higher pregnancy rates in donor oocyte recipients by comparison of fresh with frozen embryo transfer pregnancy rates in a shared oocyte programme. Hum Reprod. 1995, 10: 3022-3027.
Pelinck MJ, Hoek A, Simons AH, Heineman MJ: Efficacy of natural cycle IVF: a review of the literature. Hum Reprod Update. 2002, 82: 129-139.
Schieve LA, Tatham L, Peterson HB, Toner J, Jeng G: Spontaneous abortion among pregnancies conceived using assisted reproductive technology in the United States. Obstet Gynecol. 2003, 1015 (1): 959-967.
Buitendijk SE, Koudstaal J, Anthony S: Preterm birth in pregnancies following in vitro fertilization. Results of a Dutch multicentre prospective study. IVF Pregnancies: Outcome and Follow-up. 2000, Thesis, University of Leiden, The Netherlands, 35-58.
Gissler M, Silverio MM, Hemminki E: In-vitro fertilization pregnancies and perinatal health in Finland 1991–1993. Hum Reprod. 1995, 10: 1856-1861.
Pinborg A: IVF/ICSI twin pregnancies: risks and prevention. Hum Reprod Update. 2005 Aug 25,
Lambalk CB, van Hooff M: Natural versus induced twinning and pregnancy outcome: a Dutch nationwide survey of primiparous dizygotic twin deliveries. Eur J Obstet Gynecol Reprod Biol. 2001, 951: 68-72.
Ombelet W, Cadron I, Gerris J, De Sutter P, Bosmans E, Martens G, Ruyssinck G, Defoort P, Molenberghs G, Gyselaers W: Obstetric and perinatal outcome of 1655 ICSI and 3974 IVF singleton and 1102 ICSI and 2901 IVF twin births: a comparative analysis. Reprod Biomed Online. 2005, 11: 76-85.
Lynch A, McDuffie R, Murphy J, Faber K, Orleans M: Preeclampsia in multiple gestation: the role of assisted reproductive technologies. Obstet Gynecol. 2002, 99 (3): 445-451.
Elchalal U, Schenker JG: Obstetric outcome of in vitro fertilized pregnancies complicated by severe ovarian hyperstimulation syndrome: a multicenter study. Fertil Steril. 1998, 706: 1070-1076.
Mordechai E, Ron-El R, Raziel A, Friedler S, Schachter M, Strassburger D: Increased early pregnancy loss in IVF patients with severe ovarian hyperstimulation syndrome. Hum Reprod. 2002, 171: 107-110.
Ghazi HA, Spielberger C, Källén B: Delivery outcome after infertility – a registry study. Fertil Steril. 1991, 55: 726-732.
Williams MA, Goldman MB, Mittendorf R, Monson RR: Subfertility and the risk of low birth weight. Fertil Steril. 1991, 56: 668-671.
Hadi FH, Chantler E, Anderson E, Nicholson R, McClelland RA, Seif MW: Ovulation induction and endometrial steroid receptors. Hum Reprod. 1994, 9: 2405-2410.
Paulson RJ, Sauer MV, Lobo RA: Embryo implantation after human in vitro fertilization: importance of endometrial receptivity. Fertil Steril. 1990, 53: 870-874.
Simón C, Cano F, Valbueña D, Remohí J, Pellicer A: Clinical evidence for a detrimental effect on uterine receptivity of high serum E2 concentrations in high and normal responder patients. Hum Reprod. 1995, 10: 2432-2437.
Ng EHY, Yeung WSB, Lau EY, Ho PC: High serum oestradiol concentration in fresh IVF cycles do not impair implantation and pregnancy rates in subsequent frozen-thawed embryo transfer cycles. Hum Reprod. 2000, 15: 250-255.
Ghazala Sikandar Basir GS, Wai-sum O, Ng EH, Ho PC: Morphometric analysis of peri-implantation endometrium in patients having excessively high oestradiol concentrations after ovarian stimulation. Hum Reprod. 2001, 163: 435-440.
Mitwally MFM, Bhakoo HS, Crickard K, Sullivan MW, Batt RE, Yeh J: AUC Area under the Curve for estradiol (E2) Predicts Treatment Outcome in Women Undergoing IVF-ET In-Vitro Fertilization and Embryo Transfer. 59th Annual Meeting of the American Society for Reproductive Medicine October 11–15. 2003, , San Antonio, Texas, abstract No. 55
Mitwally MFM, Bhakoo HS, Crickard K, Sullivan MW, Batt RE, Yeh J: Area Under The Curve for estradiol (E2) Levels Do Not Reflect E2 Levels on the Day of hCG Administration among Individual Patients Undergoing COH Controlled Ovarian Hyperstimulation forIVF-ET In-Vitro Fertilization and Embryo Transfer. 59th Annual Meeting of the American Society for Reproductive Medicine October 11–15. 2003, , San Antonio, Texas, abstract No. 97
Mitwally MFM, Bhakoo HS, Crickard K, Sullivan MW, Batt RE, Yeh J, Casper RF: Advancing Age is Associated with Increased AUC Area Under the Curve for estradiol (E2) Production per Mature Follicle and Decreased AUC Per FSH Stimulation Irrespective to the Protocol of Ovarian Stimulation. 59th Annual Meeting of the American Society for Reproductive Medicine October 11–15. 2003, [Abstract], , San Antonio, Texas, abstract No. 240
Mitwally MFM, Bhakoo HS, Valenti AM, O'Gavahan H, Crickard K, Sullivan MW, Batt RE, Yeh J: Estradiol production per administered FSH and per mature follicle are good predictors for outcome of IVF-ET treatment. 51st Annual meeting of Society for Gynecologic Investigation SGI, Houston, TX. 2004
Mitwally MFM, Bakou G, Crickart K, Suillivan M, Batt R, Yeh J: The effect of estrogen levels on the outcome of treatment in repeated IVF cycles. 50th Annual meeting of Society for Gynecologic Investigation SGI, Washington DC. 2003, [Abstract]
Mitwally MFM, Abdel-Razeq SS, O'Gavaghan H, Crickard K, Suilivan M: Estradiol production during controlled ovarian hyperstimulation correlates with the risk of pregnancy loss in pregnancies achieved after IVF-ET. 60th Annual Meeting of the American Society for Reproductive Medicine October 2004, Philadelphia, PA. 2004
Mitwally MFM, Abdel-Razeq SS, Suilivan M, Crickard K: Estradiol production during controlled ovarian hyperstimulation correlates with the birth weight in pregnancies achieved after in vitro fertilization. 60th Annual Meeting of the American Society for Reproductive Medicine October 2004, Philadelphia, PA. 2004
Bagchi IC, Li Q, Cheon YP: Role of steroid hormone regulated genes in implantation. Ann N Y Acad Sci. 2001, 943: 68-76.
Psychoyos A: Hormonal control of ovoimplantation. Vitam Horm. 1973, 31: 205-255.
Psychoyos A: Uterine receptivity for nidation. Ann NY Acad Sci. 1986, 476: 36-42.
Hertig AT, Rock J, Adams EC: A description of 34 human ova within the first 17 days of development. Am J Anat. 1956, 98: 435-493.
Navot DM, Scott RT, Droesch K: The window of embryo transfer and the efficiency of human conception in vitro. Fertil Steril. 1991, 55: 114-117.
Forman R, Fries N, Testart J, Belaisch-Allart J, Hazout A, Frydman R: Evidence for an adverse effect of elevated serum estradiol concentration on embryo implantation. Fertil Steri. 1988, 49: 118-112.
Garcia JE, Acosta AA, Hsiu JG, Jones HWJ: Advanced endometrial maturation after ovulation induction with human menopausal gonadotropin/human chorionic gonadotropin for in vitro fertilization. Fertil Steril. 1984, 41: 31-35.
Kolb AB, Najmabadi S, Paulson RJ: Ultrastructural characteristics of the luteal phase endometrium in donors undergoing controlled ovarian hyperstimulation. Fertil Steril. 1997, 67: 625-630.
Fanchin R, Righini C, Olivennes F, Taieb J, de Ziegler D, Frydman R: Computerized assessment of endometrial echogenicity: clues to the endometrial effects of premature progesterone elevation. Fertil Steril. 1999, 71: 174-181.
Nikas G, Develioglu OH, Toner JP, Jones HW: Endometrial pinopodes indicate a shift in the window of receptivity in IVF cycles. Hum Reprod. 1999, 14: 787-792.
Graf MJ, Reyniak JV, Battle MP, Laufer N: Histologic evaluation of the luteal phase in women following follicle aspiration for oocyte retrieval. Fertil Steril. 1988, 49: 616-619.
Sterzik K, Dallenbach C, Schneider V, Sasse V, Dallenbach-Hellweg G: In vitro fertilization: the degree of endometrial insufficiency varies with the type of ovarian stimulation. Fertil Steril. 1988, 50: 457-462.
Toner JP, Singer GA, Jones HW: Uterine receptivity after ovarian stimulation for assisted reproduction. Implantation in Mammals. Edited by: Gianaroli L, Campana A, Trounson AO. 1993, Raven Press, New York, 231-238.
Noci I, Borri P, Coccia ME, Criscuoli L, Scarselli G, Messeri G, Paglierani M, Moncini D, Taddei G: Hormonal patterns, steroid receptors and morphological pictures of endometrium in hyperstimulated IVF cycles. Eur J Obstet. Gynecol Reprod Biol. 1997, 75: 215-220.
Macrow PJ, Li TC, Seif MW, Buckley CH, Elstein M: Endometrial structure after superovulation: a prospective controlled study. Fertil Steril. 1994, 61: 696-699.
Schoolcraft W, Sinton E, Schlenker T, Huynh D, Hamilton F, Meldrum DR: Lower pregnancy rates with premature luteinization during pituitary suppression with leuprolide acetate. Fertil Steril. 1991, 55: 563-566.
Develioglu OH, Hsiu JG, Nikas G, Toner JP, Oehninger S, Jones HW: Endometrial estrogen and progesterone receptor and pinopod expression in stimulated cycles of oocyte donors. Fertil Steril. 1999, 71: 1040-1047.
Bustillo M, Stern JJ, Coulam CB: Serum progesterone at the time of human chorionic gonadotrophin does not predict pregnancy in in-vitro fertilization and embryo transfer. Hum Reprod. 1995, 10: 2862-2867.
Ubaldi F, Camus M, Smitz J, Bennink HC, Van Steirteghem A, Devroey P: Premature luteinization in in vitro fertilization cycles using gonadotropin-releasing hormone agonist GnRH-a and recombinant follicle-stimulating hormone FSH and GnRH-a and urinary FSH. Fertil Steril. 1996, 66: 275-280.
Enders AD, Nelson DM: Pinocytotic activity of the uterus of the rat. Am J Anat. 1973, 138: 277-300.
Martel D, Monier MN, Roche D, Psychoyos A: Hormonal dependence of pinopode formation at the uterine luminal surface. Hum Reprod. 1991, 6: 597-603.
Martel D, Malet C, Gautray JP, Psychoyos A: Surface changes of the luminal uterine epithelium during the human menstrual cycle: a scanning electron microscopic study. The Endometrium: Hormonal Impacts. Edited by: de Brux J, Mortel R, Gautray JP. 1981, Plenum Press, New York, NY, USA, 15-
Nikas G, Drakakis P, Loutradis D, Mara-Skoufari C, Koumantakis E, Michalas S, Psychoyos A: Uterine pinopodes as markers of the 'Nidation Window' in cycling women receiving exogenous oestradiol and progesterone. Hum Reprod. 1995, 10: 1208-1213.
Nikas G, Reddy N, Winston RML: Implantation correlates highly with the expression of uterine pinopodes in ovum recipients under HRT: A preliminary study. Abstr. FR21 at the IX World Congress in Human Reproduction, Philadelphia PA. May 29-June, 1, 1996. [Abstract]
Martel D, Frydman R, Glissant M, Maggioni C, Roche D, Psychoyos A: Scanning electron microscopy of postovulatory human endometrium in spontaneous cycles and cycles stimulated by hormone treatment. J Endocrinol. 1987, 114: 319-324.
Thornburgh I, Anderson MC: The endometrial deficient secretory phase. Histopathology. 1997, 30: 11-15.
Seppala M, Tiitinen A: Endometrial response to corpus luteum products in cycles with induced ovulation: theoretical and practical considerations. Hum Reprod. 1995, 10S2: 67-76.
Gregory CW, Wilson EM, Apparao KB, Lininger RA, Meyer WR, Kowalik A, Fritz MA, Lessey BA: Steroid receptor coactivator expression throughout the menstrual cycle in normal and abnormal endometrium. J Clin Endocrinol Metab. 2002, 876: 2960-2966.
Papanikolaou EG, Bourgain C, Kolibianakis E, Tournaye H, Devroey P: Steroid receptor expression in late follicular phase endometrium in GnRH antagonist IVF cycles is already altered, indicating initiation of early luteal phase transformation in the absence of secretory changes. Hum Reprod. 2005, 20: 1541-7.
Gordon JD, Shifren JL, Foulk RA, Taylor RN, Jaffe RB: Angiogenesis in the human female reproductive tract. Obstet Gynecol Surv. 1995, 50: 688-697.
Rogers PAW: Structure and function of endometrial blood vessels. Hum Reprod Update. 1996, 2: 57-62.
Sterzik K, Hutter W, Grab D, Rosenbusch B, Sasse V, Terinde R: Doppler sonographic findings and their correlation with implantation in an in vitro fertilization program. Fertil Steril. 1991, 52: 825-828.
Applebaum M: The uterine biophysical profile. Ultrasound Obstet Gynecol. 1995, 51: 67-68.
Baber RJ, McSweeney MB, Gill RW, Porter RN, Picker RH, Warren PS, Kossoff G, Saunders DM: Transvaginal pulsed Doppler ultrasound assessment of blood flow to the corpus luteum in IVF patients following embryo transfer. Br J Obstet Gynaecol. 1988, 95: 1226-1230.
Scholtes MC, Wladimiroff JW, Van RH, Hop WC: Uterine and ovarian flow velocity waveforms in the normal menstrual cycle: a transvaginal Doppler study. Fertil Steril. 1989, 52: 981-985.
Basir GS, Lam TP, Chau MT, Ng EH, O WS, Ho PC: Colour Doppler analysis of peri-implantation utero-ovarian haemodynamics in women with excessively high oestradiol concentrations after ovarian stimulation. Hum Reprod. 2001, 610: 2114-2117.
Friedler S, Schenker JG, Herman A, Lewin A: The role of ultrasonography in the evaluation of endometrial receptivity following assisted reproductive treatments: a critical review. Hum Reprod Update. 1996, 2: 323-335.
Turnbull LW, Manton JD, Horsman A, Killick SR: Magnetic resonance imaging changes in uterine zonal anatomy during a conception cycle. Br J Obstet Gynaecol. 1995, 102: 330-331.
Lesny P, Killick SR, Tetlow RL, Manton DJ, Robinson J, Maguiness SD: Ultrasound evaluation of the uterine zonal anatomy during in-vitro fertilization and embryo transfer. Hum Reprod. 1999, 14: 1593-1598.
Chien LW, Au HK, Chen PL, Xiao J, Tzeng CR: Assessment of uterine receptivity by the endometrial-subendometrial blood flow distribution pattern in women undergoing in vitro fertilization-embryo transfer. Fertil Steril. 2002, 782: 245-251.
Salle BV, Bied-Damon , Benchaib M, Desperes S, Gaucherand P, Rudigoz RC: Preliminary report of an ultrasonography and colour Doppler uterine score to predict uterine receptivity in an in-vitro fertilization programme. Hum Reprod. 1998, 13: 1669-1673.
Noe M, Kunz G, Herbertz M, Mall G, Leyendecker G: The cyclic pattern of the immunocytochemical expression of oestrogen and progesterone receptors in human myometrial and endometrial layers: characterization of the endometrial-subendometrial unit. Hum Reprod. 1999, 141: 190-197.
IJland MM, Evers JL, Dunselman GA, Volovics L, Hoogland HJ: Relation between endometrial wavelike activity and fecundability in spontaneous cycles. Fertil Steril. 1997, 67: 492-495.
Fanchin Righini RC, Olivennes F, Taylor S, de Ziegler D, Frydman R: Uterine contractions at the time of embryo transfer alter pregnancy rates after in-vitro fertilization. Hum Reprod. 1998, 13: 1968-1974.
Kupesic S, Bekavac I, Bjelos D, Kurjak A: Assessment of endometrial receptivity by ransvaginal color Doppler and three-dimensional power Doppler ultrasonography in patients undergoing in vitro fertilization procedures. J Ultrasound Med. 2001, 20: 125-134.
Zaidi J, Campbell S, Pittrof R, Tan SL: Endometrial thickness, morphology, vascular penetration and velocimetry in predicting implantation in an in vitro fertilization program. Ultrasound Obstet Gynecol. 1995, 6: 191-198.
Yang JH, Wu MY, Chen CD, Jiang MC, Ho HN, Yang YS: Association of endometrial blood flow as determined by a modified color Doppler technique with subsequent outcome of in-vitro fertilization. Hum Reprod. 1999, 14: 1606-1610.
Yuval Y, Lipitz S, Dor J, Achiron R: The relationships between endometrial thickness, and blood flow and pregnancy rates in in-vitro fertilization. Hum Reprod. 1999, 14: 1067-1071.
Schild RL, Knobloch C, Dorn C, Fimmers R, van der Ven H, Hansmann H: Endometrial receptivity in an in vitro fertilization program as assessed by spiral artery blood flow, endometrial thickness, endometrial volume, and uterine artery blood flow. Fertil Steril. 2001, 5: 361-366.
Lim H, Song H, Paria BC, Reese J, Das SK, Dey SK: Molecules in blastocyst implantation: uterine and embryonic perspectives. Vitam Horm. 2002, 64: 43-76.
Paria BC, Lim H, Das SK, Reese J, Dey SK: Molecular signaling in uterine receptivity for implantation. Semin Cell Dev Biol. 2000, 112: 67-76.
Fanchin R, Righini C, Schonauer LM, Olivennes F, Filho JS, Frydman R: Vaginal versus oral estradiol administration: effects on endometrial thickness, uterine perfusion, and contractility. Fertil Steril. 2001, 765: 994-998.
Basir GS, Lam TP, Chau MT, Ng EH, O WS, Ho PC: Colour Doppler analysis of peri-implantation utero-ovarian haemodynamics in women with excessively high oestradiol concentrations after ovarian stimulation. Hum Reprod. 2001, 1610: 2114-2117.
Long MG, Boultbee JE, Hanson ME, Begent RH: Doppler time velocity waveform studies of the uterine artery and uterus. Br J Obstet Gynaecol. 1989, 96: 588-593.
Basir GS, Ng EHY, Ho PC: Morphometric analysis of peri-implantation endometrium in patients having excessively high oestradiol concentrations after ovarian stimulation. Hum Reprod. 2001, 16: 435-440.
Gannon BJ, Carati CJ, Verco CJ: Endometrial perfusion across the normal human menstrual cycle assessed by laser Doppler fluxmetry. Hum Reprod. 1997, 12: 132-139.
Cross JC, Werb Z, Fisher SJ: Implantation and the placenta: key pieces of the development puzzle Implantation and the placenta: key pieces of the development puzzle. Science. 1994, 266: 1508-1518.
Zaidi J, Jurkovic D, Campbell S, Okokon E, Tan SL: Circadian variation in uterine artery blood flow indices during the follicular phase of the menstrual cycle. Ultrasound Obstet Gynecol. 1995, 5: 406-410.
Edwards R: Causes of early embryonic loss in human pregnancy. Hum Reprod. 1986, 1: 185-198.
Van Blerkom J: Developmental failure in human reproduction associated with chromosomal abnormalities and cytoplasmic pathologies in meiotically-mature human oocytes. The Biological Basis of Early Reproductive Failure in the Human: Applications to Medically Assisted Conception. Edited by: Van Blerkom J. 1994, Oxford University Press, Oxford, 283-325.
Van Blerkom J: The influence of intrinsic and extrinsic factors on the developmental potential of chromosomal normality of the human oocyte. J Soc Gyn Invest. 1996, 3: 3-11.
Van Blerkom J: Epigenetic influences on oocyte developmental competence: perifollicular vascularity and intrafollicular oxygen. J Assist Reprod Genet. 1998, 155: 226-234.
Van Blerkom J, Antczak M, Schrader R: The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum Reprod. 1997, 125: 1047-1055.
Tabibzadeh S: Patterns of expression of integrin molecules in human endometrium throughout the menstrual cycle. Hum Reprod. 1992, 7: 876-882.
Gonzalez RR, Palomino A, Boric A, Vega M, Devoto L: A quantitative evaluation of alpha1, alpha4, alphaV and beta3 endometrial integrins of fertile and unexplained infertile women during the menstrual cycle. A flow cytometric appraisal. Hum Reprod. 1999, 14: 2485-2492.
Klentzeris LD: Adhesion molecules in reproduction. Br J Obstet Gynaecol. 1997, 104: 401-409.
Creus M, Balasch J, Ordi J, Fabregues F, Casamitjana R, Quinto L, Coutifaris C, Vanrell JA: Integrin expression in normal and out of phase endometria. Hum Reprod. 1998, 13: 3460-3468.
Thomas K, Thomson AJ, Sephton V, Cowan C, Wood S, Vince S, Kingsland CR, Lewis-Jones DI: The effect of gonadotrophic stimulation on integrin expression in the endometrium. Hum Reprod. 2002, 17: 63-68.
Gosden RG: Oogenesis as a foundation for embryogenesis. Mol Cell Endocrinol. 2002, 25: 149-53.
Wu TC, Wang L, Wan YJ: Detection of estrogen receptor messenger ribonucleic acid in human oocytes and cumulus-oocyte complexes using reverse transcriptase-polymerase chain reaction. Fertil Steril. 1993, 591: 54-59.
Pellicer A, Ruiz A, Castellvi RM, Calatayud C, Ruiz M, Tarin JJ, Miro F, Bonilla-Musoles F: Is the retrieval of high numbers of oocytes desirable in patients treated with gonadotrophin-releasing hormone analogues GnRHa and gonadotrophins?. Hum Reprod. 1989, 4: 536-540.
Dailey T, Dale B, Cohen J: Association between nondisjunction and maternal age in meiosis-II human oocytes. Am J Hum Genet. 1996, 9: 2013-2084.
Golbus M: The influence of strain, maternal age, and method of maturation on mouse oocyte aneuploidy. Cytogenet Cell Genet. 1981, 31: 84-90.
Sengoku K, Dukelow R: Gonadotropin effects on chromosomal normality of hamster preimplantation embryos. Biol Reprod. 1988, 38: 150-155.
Van Blerkom J: Origin and detection of chromosomal defects in human oocytes and embryos. Presented on International Symposium on Genetics of Gametes and Embryos, 2–5 June, 1994, New York. Book of Abstracts. Genetics of Gametes and Embryos. Serono Symposia, New York
Laverge H, De-Sutter P, Verschraegen-Spae MR, De-Paepe A, Dhont M: Triple colour fluorescence in situ hybridization for chromosomes X, Y, and 1 on spare human embryos. Hum Reprod. 1997, 4: 809-814.
Munne S, Alikani M, Tomkin G, Grifo J, Cohen J: Embryo morphology, developmental rates, and maternal age are correlated with chromosome abnormalities. Fertil Steril. 1995, 2: 382-391.
Tsutsui T, Suzuki N, Fukuda S, Sato M, Maisumi H, McLachlan JA, Barrett JC: 17β-estradiol-induced cell transformation and aneuploidy of Syrian hamster embryo cells in culture. Carcinogenesis. 1987, 8: 1715-1719.
Tsutsui T, Barrett JC: Neoplastic transformation of cultured mammalian cells by estrogens and estrogen-like chemicals. Environ Health Perspect. 1997, 105: 619-324.
Jones LA, Hajek RA: Effects of estrogenic chemicals on development. Environ Health Perspect. 1995, 103: 63-67.
Hajek RA, Pathak S, Boddie AK, Jones LA: Aneuploidy of mouse cervicovaginal epithelium induced by perinatal estrogen treatment. Proc Am Assoc Cancer Res. 1989, 30: 299-
Klein CB: Are diethylstilbestrol and estradiol mutagenic?. Proc Am Assoc Cancer Res. 1995, 36: 259 Abstract-
Su L, Zinaman R, Klein CB: Mechanisms of gene inactivation by estrogens. Proc Am Assoc Cancer Res. 1998, 39: 93 Abstract-
Liehr JG: Is estradiol a genotoxic mutagenic carcinogen?. Endocr Rev. 2000, 211: 40-54.
Lengauer C, Kinzler KW, Vogelstein B: Genetic instabilities in human cancers. Nature. 1998, 396: 643-649.
Can A, Semiz O: Diethylstilbestrol DES-induced cell cycle delay and meiotic spindle disruption in mouse oocytes during in-vitro maturation. Mol Hum Reprod. 2000, 62: 154-162.
Liehr JG, Fang WF, Sirbasku DA, Ari-Ulubelen A: Carcinogenicity of catechol estrogens in Syrian hamsters. J Steroid Biochem. 1986, 24: 353-356.
Liehr JG, Sirbasku DA: Estrogen-dependent kidney tumors. In: Taub M ed Tissue Culture of Epithelial Cells. 1985, Plenum Press, New York, 205-234.
Ho S-M, Roy D: Sex hormone-induced nuclear DNA damage and lipid peroxidation in the dorsolateral prostates of Noble rats. Cancer Lett. 1994, 84: 155-162.
Han X, Liehr JG: Microsome-mediated 8-hydroxylation of guanine bases of DNA by steroid estrogens: correlation of DNA damage by free radicals with metabolic activation to quinones. Carcinogenesis. 1994, 16: 2571-2574.
Rosier JA, Van Peteghem CH: Peroxidative in vitro metabolism of diethylstilbestrol induces formation of 8-hydroxy-2'-deoxyguanosine. Carcinogenesis. 1989, 10: 405-406.
Gaulden M: Maternal age effect: the enigma of Down syndrome and other trisomic conditions. Mutat Res. 1992, 296: 69-88.
Van Blerkom J, Sinclair J, Davis P: Mitochondrial transfer between oocytes: potential applications of mitochondrial donation and the issue of heteroplasmy. Hum Reprod. 1998, 3: 2857-2868.
Evans RM: The steroid and thyroid hormone receptor superfamily. Science. 1988, 240: 889-895.
Wehling M: Specific, nongenomic actions of steroid hormones. Ann Rev Physiol. 1997, 59: 365-393.
Nemere I, Farach-Carson MC: Membrane receptors for steroid hormones: a case for specific cell surface binding sites for vitamin D metabolites and estrogens. Biochem Biophys Res Commun. 1998, 248: 443-449.
Revelli A, Massobrio M, Tesarik J: Nongenomic actions of steroid hormones in reproductive tissues. Endocr Rev. 1998, 19: 3-17.
Pietras RJ, Szego CM: Estrogen receptors in uterine plasma membranes. J Steroid Biochem. 1979, 11: 1471-1483.
Pappas T, Gametchu B, Watson C: Membrane estrogen receptors identified by multiple antibody labelling and impeded-ligand binding. FASEB J. 1995, 9: 404-410.
Demonacos CV, Karayanni N, Hatzoglou E, Tsiriyotis C, Spandidos DA, Sekeris CE: Mitochondrial genes as sites of primary action of steroid hormones. Steroids. 1996, 61: 226-232.
Noteboom WD, Gorski J: Stereospecific binding of estrogens in the rat uterus. Arch Biochem Biophys. 1965, 111: 559-568.
Monje P, Boland R: Subcellular distribution of native estrogen receptor alpha and beta isoforms in rabbit uterus and ovary. J Cell Biochem. 2001, 823: 467-479.
Morkuniene R, Jekabsone A, Borutaite V: Estrogens prevent calcium-induced release of cytochrome c from heart mitochondria. FEBS Lett. 2002, 19: 5211-5213.
Leshchenko MV, Sergeev PV: Nature of steroid hormone binding to mitochondria. Pharmacol Toxicol. 1987, 50: 60-63.
Aquila S, Sisci D, Gentile M, Middea E, Siciliano L, Ando S: Human ejaculated spermatozoa contain active P450 aromatase. J Clin Endocrinol Metab. 2002, 87: 3385-3390.
Adeoya-Osiguwa SA, Markoulaki S, Pocock V, Milligan SR, Fraser LR: 17beta-estradiol and environmental estrogens significantly affect mammalian sperm function. Hum Reprod. 2003, 181: 100-107.
Luconi M, Muratori M, Forti G, Baldi E: Identification and characterization of a novel functional estrogen receptor on human sperm membrane that interferes with progesterone effects. J Clin Endocrinol Metab. 1999, 84 (5): 1670-167.
Durkee TJ, Mueller M, Zinaman M: Identification of estrogen receptor protein and messenger ribonucleic acid in human spermatozoa. Am J Obstet Gynecol. 1998, 178: 1288-1297.
Yanagimachi R: Mammalian fertilization. The Physiology of Reproduction. Edited by: Knobil E, Neill JD. 1994, Raven Press, New York, 189-317. 2
Austin CR: The "capacitation" of the mammalian sperm. Nature. 1952, 170: 326-
Lobo RA, diZerega GS, Marrs RP: Follicular fluid steroid levels in dysmature and mature follicles from spontaneous and hyperstimulated cycles in normal and anovulatory women. J Clin Endocrinol Metab. 1985, 60: 81-87.
Andersen CY: Characteristics of human follicular fluid associated with successful conception after in vitro fertilization. J Clin Endocrinol Metab. 1993, 77: 1227-1234.
Fraser LR, Adeoya-Osiguwa SA: Fertilization promoting peptide – A possible regulator of sperm function in vivo. Vit Horm. 2001, 63: 1-28.
Van der Auwera , D'Hooghe T: Superovulation of female mice delays embryonic and fetal development. Hum Reprod. 2001, 166: 1237-1243.
Walton EA, Armstrong DT: Oocyte normality after superovulation in immature rats. J Reprod Fertil. 1983, 67: 309-314.
Walton EA, Huntley S, Kennedy TG, Armstrong DT: Possible causes of implantation failure in superovulated immature rats. Biol Reprod. 1982, 27: 847-852.
Ng EHY, Yeung WSB, Lau EYL, So WWK, Ho PC: High serum oestradiol concentrations in fresh IVF cycles do not impair implantation and pregnancy rates in subsequent frozen-thawed embryo transfer cycles. Hum Reprod. 2000, 15: 250-255.
Schalkoff ME, Oskowitz SP, Powers RD: A multifactorial analysis of the pregnancy outcome in a successful cryopreservation program. Fertil Steril. 1993, 59: 1070-1074.
Champlin AK, Kuzia SJ, Rice BA, Mobraaten LE: Cell surface characteristics of blastocysts from spontaneously ovulating and gonadotropin-treated mice. Biol Reprod. 1987, 36: 439-444.
Wetzels AM, Artz MT, Goverde HJ, Bastiaans BA, Hamilton CJ, Rolland R: Gonadotropin hyperstimulation influences the 35S-methionine metabolism of mouse preimplantation embryos. J Assist Reprod Genet. 1995, 12: 744-746.
Elmazar M, Vogel R, Spielmann H: Maternal factors influencing development of embryos from mice superovulated with gonadotropins. Reprod Toxicol. 1989, 3: 135-138.
Ertzeid G, Storeng R: Adverse effects of gonadotrophin treatment on pre- and postimplantation development in mice. J Reprod Fertil. 1992, 96: 649-655.
Pellicer A, Valbuena D, Cano F, Remohí J, Simón C: Lower implantation rates in high responders: evidence for an altered endocrine milieu during the preimplantation period. Fertil Steril. 1996, 65: 1190-1195.
Valbuena D, Martin J, de Pablo JL, Remohi J, Pellicer A, Simon C: Increasing levels of estradiol are deleterious to embryonic implantation because they directly affect the embryo. Fertil Steril. 2001, 765: 962-968.
Khalifa EA, Tucker MJ, Hunt P: Cruciate thinning of the zona pellucida for more successful enhancement of blastocyst hatching in the mouse. Hum Reprod. 1992, 7: 532-536.
Germond M, Nocera D, Senn A, Rink K, Delacretaz G, Pedrazzini T, Hornung JP: Improved fertilization and implantation rates after non-touch zona pellucida microdrilling of mouse oocytes with a 1.48 micrometer diode laser beam. Hum Reprod. 1996, 11: 1043-1048.
Montag M, van der Ven H: Laser-assisted hatching in assisted reproduction. Croat Med J. 1999, 40: 398-403.
Bertrand E, Van den Beergh M, Englert Y: Does zona pellucida thickness influence the fertilization rate?. Hum Reprod. 1995, 105: 1189-1193.
Garside WT, Loret De Mola JR, Bucci JA, Tureck RW, Heyner : Sequential analysis of zona thickness during in vitro culture of human zygotes: correlation with embryo quality, age, and implantation. Mol Reprod Dev. 1997, 47 (1): 99-104.
Cohen J, Alikani M, Trowbridge J, Rosenwaks Z: Implantation enhancement by selective assisted hatching using zona drilling. Hum Reprod. 1992, 21: 565-570.
Cohen J, Feldberg D: Effects of the site and number of zona pellucida openings on hatching and trophoblast outgrowth in the mouse embryo. Mol Reprod Dev. 1991, 30: 70-78.
Chan PJ: Developmental potential of human oocytes according to zona pellucida thickness. J In Vitro Fert Embryo Transfer. 1987, 4 (4): 237-241.
Cohen J, Wiemer KE, Wright G, Prognostic value of morpho-7, Loret De Mola JR, Garside WT, Bucci J, Tureck RW, Heyner S: Analysis of the human zona pellucida during culture: correla-logic characteristics of cryopreserved embryos: A study using videocinematography. Fertil Steril. 1988, 49: 827-834.
Cohen J, Inge KL, Suzman M, Wiker SR, Wright G: Videocinematography of fresh and cryopreserved embryos: a retrospective analysis of embryonic morphology and implantation. Fertil Steril. 1989, 51 (5): 820-827.
Bertrand E, Van den Bergh M, Englert Y: Clinical parameters influencing human zona pellucida thickness. Fertil Steril. 1996, 66: 408-411.
Soliman S, Daya S, Collins J, Hughes EG: The role of luteal phase support in infertility treatment: a meta-analysis of randomized trials. Fertil Steril. 1994, 61: 1068-1076.
Schally AV, Halmos G: Terminology for luteinizing hormone-releasing hormone antagonists letter. Fertil Steril. 1995, 64: 226-
Srkalovic G, Bokser L, Radulovic S, Korkut E, Schally AV: Receptors for luteinizing hormone-releasing hormone LHRH in Dunning R3327 prostate cancer and rat anterior pituitaries after treatment with a sustained delivery system of LHRH antagonist SB-75. Endocrinology. 1990, 12: 3052-3060.
Hosokawa K, Ottander U, Wahlberg P, Ny T, Cajander S, Olofsson IJ: Dominant expression and distribution of oestrogen receptor b over oestrogen receptor a in the human corpus luteum. Mol Hum Reprod. 2001, 7: 137-145.
Gore BZ, Caldwell BV, Speroff L: Estrogen-induced human luteolysis. J Clin Endocrinol Metab. 1973, 36: 615-617.
Vega M, Devoto L, Castro O, Kohen P: Progesterone synthesis by human luteal cells: modulation by estradiol. J Clin Endocrinol Metab. 1994, 79: 466-469.
Khan-Dawood FS, Yang J, Dawood MY: Expression of gap junction protein connexin-43 in the human and baboon Papio anubis corpus luteum. J Clin Endocrinol Metab. 1996, 81: 835-842.
Westergaard : Increased risk of early pregnancy loss by profound suppression of luteinizing hormone during ovarian stimulation in normogonadotrophic women undergoing assisted reproduction. Hum Reprod. 2000, 155: 1003-1008.
Goh HH, Ratnam SS: The gonadotrophin surge in humans: its mechanism and role in ovulatory function – a review. Ann Acad Med Singapore. 1990, 194: 524-529.
Gandar R, Collin D: Pre-ovulatory peaks of gonadotropins. J Gynecol Obstet Biol Reprod Paris. 1997, 266: 567-575.
Messinis IE, Templeton A, Baird DT: Relationships between the characteristics of endogenous luteinizing hormone surge and the degree of ovarian hyperstimulation during superovulation induction in women. Clin Endocrinol Oxf. 1986, 254: 393-400.
Messinis IE, Templeton AA: Endocrine and follicle characteristics of cycles with and without endogenous luteinizing hormone surges during superovulation induction with pulsatile follicle-stimulating hormone. Hum Reprod. 1998, 21: 11-16.
Goh HH: The gonadotrophin surge in human reproduction: endocrine and biochemical mechanisms. Adv Clin Chem. 1994, 31: 135-176.
Tain CF, Goh VH, Ng SC: Effects of hyperstimulation with gonadotrophins and age of females on oocytes and their metaphase II status in rats. Mol Reprod Dev. 2000, 551: 104-108.
Soules M, Steiner R, Clifton D, Cohen N, Aksel S, Bremner W: Progesterone modulation of pulsatile luteinizing hormone secretion in normal women. J Clin Enocrinol Metab. 1984, 58: 378-382.
Nippoldt TB, Reame N, Kelch RP, Marshall J: The roles of estradiol and progesterone in decreasing luteinizing hormone pulse frequency in the luteal phase of the menstrual cycle. J Clin Endocrinol Metab. 1989, 69: 67-76.
Ditkoff EC, Cassidenti DL, Paulson RJ, Sauer MV, Paul WL, Rivier J, Yen SS, Lobo RA: The gonadotropin-releasing hormone antagonist Nal-Glu acutely blocks the luteinizing hormone surge but allows for resumption of folliculogenesis in normal women. Am J Obstet Gynecol. 1991, 165: 1811-1817.
de Jong D, Macklon MD, Fauser BCJM: A pilot study involving minimal ovarian stimulation for in-vitro fertilization: extending the 'follicle-stimulating hormone window' combined with the gonadotropin releasing hormone antagonist Cetrorelix. Fertil Steril. 73: 1051-1054.
Brannian JD, Hansen KA: Leptin and ovarian folliculogenesis: implications for ovulation induction and ART outcomes. Semin Reprod Med. 2002, 202: 103-112.
Bützow TL, Moilanen JM, Lehtovirta M: Serum and follicular fluid leptin during in vitro fertilization: relationship among leptin increase, body fat mass, and reduced ovarian response. J Clin Endocr Metab. 1999, 84: 3135-3139.
Brzechffa PR, Jakimiuk AJ, Agarwal SK, Weitsman SR, Buyalos RP, Magoffin DA: Serum immunoreactive leptin concentrations in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 1996, 81: 4166-4169.
Yamada M, Irahara M, Tezuka M, Murakami T, Shima K, Aono T: Serum leptin profiles in the normal menstrual cycles and gonadotropin treatment cycles. Gynecol Obstet Invest. 2000, 49: 119-123.
Mannucci E, Ognibene A, Becorpi A: Relationship between leptin and estrogens in healthy women. Eur J Endocr. 1998, 139: 198-201.
Strowitzki T, Kellerer M, Capp E, Haring HU: Increase in serum leptin concentrations in women undergoing controlled ovarian hyperstimulation for assisted reproduction. Gynecol Endocr. 1998, 12: 167-169.
Stock SM, Sande EM, Bremme KA: Leptin levels vary significantly during the menstrual cycle, pregnancy, and in vitro fertilization treatment: possible relation to estradiol. Fertil Steril. 1999, 72: 657-662.
Zhao Y, Kreger DO, Brannian JD: Serum leptin concentrations in women during gonadotropin stimulation cycles. J Reprod Med. 2000, 45: 121-125.
Lindheim SR, Sauer MV, Carmina E, Chang PL, Zimmerman R, Lobo RA: Circulating leptin levels during ovulation induction: relation to adiposity and ovarian morphology. Fertil Steril. 2000, 73: 493-498.
Unkila-Kallio L, Anderson S, Koistinen HA, Karonen SL, Ylikorkala O, Tiitinen A: Leptin during assisted reproductive cycles: the effect of ovarian stimulation and of very early pregnancy. Hum Reprod. 2001, 16: 657-662.
Fedorcsak P, Storeng R, Dale PO, Tanbo T, Abyholm T: Obesity is a risk factor for early pregnancy loss after IVF or ICSI. Acta Obstet Gyn Scand. 2000, 79: 43-48.
Wittemer C, Ohl J, Bailly M, Bettahar-Lebugle K, Nisand I: Does body mass index of infertile women have an impact on IVF procedure and outcome?. J Asst Reprod Genet. 2000, 17: 547-542.
Barroso G, Barrionuevo M, Rao P: Vascular endothelial growth factor, nitric oxide, and leptin follicular fluid levels correlate negatively with embryo quality in IVF patients. Fertil Steril. 1999, 72: 1024-1026.
Van Blerkom J, Antczak M, Schrader R: The developmental potential of the human oocyte is related to the dissolved oxygen content of follicular fluid: association with vascular endothelial growth factor levels and perifollicular blood flow characteristics. Hum Reprod. 1997, 12: 1047-1055.
Manzoros CS, Cramer DW, Liberman RF, Barbieri RL: Predictive value of serum and follicular fluid leptin concentrations during assisted reproductive cycles in normal women and in women with the polycystic ovarian syndrome. Hum Reprod. 2000, 15: 539-544.
Zhao Y, Kreger DO, Brannian JD: Leptin concentrations in serum and follicular fluid during gonadotropin stimulation cycles. J Soc Gynecol Invest. 1998, 5Sl: 50A-
Kitawaki J, Koshiba H, Ishihara H, Kusuki I, Tsukamoto K, Honjo H: Expression of leptin receptor in human endometrium and fluctuation during the menstrual cycle. J Clin Endocr Metab. 2000, 85: 1946-1950.
Casabiell X, Pineiro V, Peino R: Gender differences in both spontaneous and stimulated leptin secretion by human omental adipose tissue in vitro: dexamethasone and estradiol stimulate leptin release in women, but not in men. J Clin Endocr Metab. 1998, 83: 2149-2155.
Wu-Peng S, Rosenbaum M, Nicholson M, Chua SC, Leibel RL: Effects of exogenous gonadal steroids on leptin homeostasis in rats. Obes Res. 1999, 7: 586-592.
Sher G, Salem R, Feinman M, Dodge S, Zouves C, Knutzen V: Eliminating the risk of life-threatening complications following overstimulation with menotropin fertility agents: a report on women undergoing in vitro fertilization and embryo transfer. Obstet Gynecol. 1993, 81: 1009-1011.
Kim HC, Kemmann E, Shelden RM, Saidi P: Response of blood coagulation parameters to elevated endogenous 17 beta-estradiol levels induced by human menopausal gonadotropins. Am J Obstet Gynecol. 1981, 140: 807-810.
Lox C, Canez M, DeLeon F, Dorsett J, Prien S: Hyperestrogenism induced by menotropins alone or in conjunction with luprolide acetate in in vitro fertilization cycles: the impact on hemostasis. Fertil Steril. 1995, 633: 566-570.
Brechmann J, Unterberg C: Superior vena cava thrombosis after in vitro fertilization. Deutsche Medicine Wochenschr. 2000, 125: 1429-1432.
Ludwig M, Felberbaum RE, Diedrich K: Deep vein thrombosis during administration of HMG for ovarian stimulation. Archives of Gynecological Obstetrics. 2000, 263: 139-141.
Curvers J, Nap AW, Thomassen MC, Nienhuis SJ, Hamulyak K, Evers JL, Tans G, Rosing J: Effect of in vitro fertilization treatment and subsequent pregnancy on the protein C pathway. Br J Haematol. 2001, 1152: 400-4007.
Biron C, Galtier-Dereure F, Rabesandratana H, Bernard I, Aguilar-Martinez P, Schved JF, Hedon B: Hemostasis parameters during ovarian stimulation for in vitro fertilization: results of a prospective study. Fertil Steril. 1997, 67: 104-109.
Wramsby ML, Bokarewa MI, Blomback M, Bremme AK: Response to activated protein C during normal menstrual cycle and ovarian stimulation. Hum Reprod. 2000, 15: 795-797.
Curvers J, Thomassen MC, Nicolaes GA, Van Oerle R, Hamulyak K, Hemker HC, Tans G, Rosing J: Acquired APC resistance and oral contraceptives: differences between two functional tests. Br J Haemat. 1999, 105: 88-94.
Chegini N: Oviductal-derived growth factors and cytokines: implications in preimplantation. Semin Reprod Endocrinol. 1996, 14: 219-229.
Evans MI, Schulman JD, Golden L, Mukherjee AB: Superovulation-induced intrauterine growth retardation in mice. Am J Obstet Gynecol. 1981, 141: 433-435.
Guzick DS, Carson SA, Coutifaris C, Overstreet JW, Factor-Litvak P, Steinkampf MP, Hill JA, Mastroianni L, Buster JE, Nakajima ST, Vogel DL, Canfield RE: Efficacy of superovulation and intrauterine insemination in the treatment of infertility. National Cooperative Reproductive Medicine Network. N Engl J Med. 1999, 340: 177-183.
Chang YS, Kim SH, Moon SY, Lee JY: Current status of assisted reproductive technology in Asia and Oceania. J Obstet Gynaecol Res. 1996, 22: 305-330.
ASRM/SART: Assisted reproductive technology in the United Stated and Canada: 1994 results generated from the American Society for Reproductive Medicine/Society for Assisted Reproductive Technology Registry. Fertil Steril. 1996, 66: 697-705.
Gardner DK, Lane M: Culture and selection of viable blastocysts: a feasible proposition for human IVF?. Hum Reprod Update. 1997, 3: 367-382.
Van der Auwera I, Pijnenborg R, Koninckx PR: The influence of in vitro culture versus stimulated and untreated oviductal environment on mouse embryo development and implantation. Human Reprod. 1999, 14: 2570-2574.
Akira S, Sanbuissho A, Lin YC, Araki T: Acceleration of embryo transport in superovulated adult rats. Life Sci. 1993, 5315: 1243-1251.
Haspels AA: Post-coital oestrogen in large doses. IPPF Med Bull. 1972, 62: 3-4.
Greenwald GS: Species differences in egg transport in response to exogenous estrogen. Anat Rec. 1967, 157: 163-172.
Harper MJK: Interference with early pregnancy in rats by estrogen: mechanism of action of dienestrol. Anat Rec. 1968, 162: 433-452.
Ortiz ME, Bastías G, Darrigrande O, Croxatto HB: Importance of uterine expulsion of embryos in the interceptive mechanism of postcoital oestradiol in rats. Reprod Fertil Dev. 1991, 3: 333-3337.
Greb RR, Heikinheimo O, Williams RF, Hodgen GD, Goodman AL: Vascular endothelial growth factor in primate endometrium is regulated by estrogen-receptor and progesterone-receptor ligands in vivo. Hum Reprod. 1997, 12: 120-129.
Yamaji T, Dierschke DJ, Hotchkiss J, Bhattacharya AN, Surve AH, Knobil E: Estrogen induction of LH release in the Rhesus monkey. Endocrinology. 1971, 89: 1034-1041.
Chaffin ChL, Heimler I, Rawlins RG, Wimpee BAB, Sommer C, Hutz RJ: Estrogen receptor and aromatic hydrocarbon receptor in the primate ovary. Endocrine. 1996, 315: 2013-2021.
Hutz RJ, Dierschke DJ, Wolf RC: Estradiol-induced follicular atresia in rhesus monkey is not prevented by exogenous gonadotropins. Am J Primatol. 1991, 23: 247-255.
Jones RL, Salamonsen LA, Findlay JK: Potential roles for endometrial inhibins, activins and follistatin during human embryo implantation and early pregnancy. Trends Endocrinol Metab. 2002, 134: 144-150.
Ding YQ, Zhu LJ, Bagchi MK, Bagchi IC: Progesterone stimulates calcitonin gene expression in the uterus during implantation. Endocrinology. 1994, 135: 2265-2274.
Bagchi IC, Li Q, Cheon YP: Role of steroid hormone-regulated genes in implantation. Ann N Y Acad Sci. 2001, 943: 68-76.
Franks S, Adams J, Mason H, Polson D: Ovulatory disorders in women with polycystic ovary syndrome. Clin Obstet Gynaecol. 1985, 12: 605-632.
Kistner R: Induction of ovulation with clomiphene citrate clomid. Obstet Gynecol Surv. 1965, 20: 873-900.
Garcia J, Jones GS, Wentz AC: The use of clomiphene citrate. Fertil Steril. 1977, 28: 707-717.
Goldfarb AF, Morales A, Rakoff AE, Protos P: Critical review of 160 clomiphene-related pregnancies. Obstet Gynecol. 1968, 31: 342-345.
Randall JM, Templeton A: Cervical mucus score and in vitro sperm mucus interaction in spontaneous and clomiphene citrate cycles. Fertil Steril. 1991, 56: 465-468.
Gysler M, March C, Mishell DJ, Bailey EJ: A decade's experience with an individualized clomiphene treatment regimen including its effects on the postcoital test. Fertil Steril. 1982, 37: 161-167.
Gonen Y, Casper RF: Sonographic determination of an adverse effect of clomiphene citrate on endometrial growth. Hum Reprod. 1990, 5: 670-674.
Nelson LM, Hershlag A, Kurl RS, Hall JL, Stillman RJ: Clomiphene citrate directly impairs endometrial receptivity in the mouse. Fertil Steril. 1990, 53: 727-731.
Young SL, Opsahl MS, Fritz MA: Serum concentrations of enclomiphene and zuclomiphene across consecutive cycles of clomiphene citrate therapy in anovulatory infertile women. Fertil Steril. 1999, 71: 639-644.
Mikkelson TJ, Kroboth PD, Cameron WJ, Dittert LW, Chungi V, Manberg PJ: Single-dose pharmacokinetics of clomiphene citrate in normal volunteers. Fertil Steril. 1986, 46: 392-396.
Schmidt GE, Kim MH, Mansour R, Torello L, Friedman CI: The effects of enclomiphene and zuclomiphene citrates on mouse embryos fertilized in vitro and in vivo. Am J Obstet Gynecol. 1986, 154: 727-736.
Yoshimura Y, Hosoi Y, Atlas SJ, Wallach EE: Effect of clomiphene citrate on in vitro ovulated ova. Fertil Steril. 1986, 45: 800-804.
Branigan EF, Estes MA: Minimal stimulation IVF using clomiphene citrate and oral contraceptive pill pretreatment for LH suppression. Fertil Steril. 2000, 73: 587-590.
Zayed F: Outcome of stimulated in vitro fertilisation SIVF using clomiphene citrate and human menopausal gonadotropin in different infertility groups. Clin Exp Obstet Gynecol. 1999, 26: 227-279.
London SN, Young D, Caldito G, Mailhes JB: Clomiphene citrate-induced perturbations during meiotic maturation and cytogenetic abnormalities in mouse oocytes in vivo and in vitro. Fertil Steril. 2000, 73: 620-626.
Oktay K, Berkowitz P, Berkus M, Schenken RS, Brzyski RG: The re-incarnation of an old question clomid effect on oocyte and embryo?. Fertil Steril. 2000, 74: 422-
Hsu CC, Kuo HC, Wang ST, Huang KE: Interference with uterine blood flow by clomiphene citrate in women with unexplained infertility. Obstet Gynecol. 1995, 86: 917-921.
Hammond M, Halme J, Talbert L: Factors affecting the pregnancy rate in clomiphene citrate induction of ovulation. Obstet Gynecol. 1983, 62: 196-202.
Ziecik AJ, Derecka-Reszka K, Rzucidlo SJ: Extragonadal gonadotropin receptors, their distribution and function. J Physiol Pharmacol. 1992, 434 (S1): 33-49.
Reshef E, Lei ZM, Rao CV, Pridham DD, Chegini N, Luborsky JL: The presence of gonadotropin receptors in nonpregnant human uterus, human placenta, fetal membranes, and decidua. J Clin Endocrinol Metab. 1990, 70: 421-430.
Lei ZM, Rao CV, Lincoln SR, Ackermann DM: Increased expression of human chorionic gonadotropin/human luteinizing hormone receptors in adenomyosis. J Clin Endocrinol Metab. 1993, 76: 763-768.
Toth P, Li X, Rao CV, Lincoln SR, Sanfilippo JS, Spinnato JA, Yussman MA: Expression of functional human chorionic gonadotropin/human luteinizing hormone receptor gene in human uterine arteries. J Clin Endocrinol Metab. 1994, 79: 307-315.
Lincoln SR, Lei ZM, Rao CV, Yussman MA: The expression of human chorionic gonadotropin/human luteinizing hormone receptors in ectopic human endometrial implants. J Clin Endocrinol Metab. 1992, 75: 1140-1144.
Stewart EA: Gonadotropins and the uterus: is there a gonad-independent pathway?. J Soc Gynecol Investig. 2001, 86: 319-326.
Casslén BG, Siler-Khodr TM, Harper MJK: Progesterone regulation of prolactin release from human endometrial stromal cells in culture: potential bioassay for progestational activity. Acta Endocrinol Copenhagen. 1990, 122: 137-144.
Schenken RS, Hodgen GD: Follicle-stimulating hormone induced ovarian hyperstimulation in monkeys: blockade of the luteinizing hormone surge. J Clin Endocrinol Metab. 1983, 571: 50-55.
Messinis IE, Templeton AA: Pituitary response to exogenous LHRH in superovulated women. J Reprod Fertil. 1989, 87: 633-639.
Messinis IE, Templeton AA: Evidence that gonadotrophin surge-attenuating factor exists in man. J Reprod Fertil. 1991, 92: 217-223.
Messinis IE, Lolis D, Papadopoulos L, Tsahalina T, Papanikolaou N, Seferiadis K, Templeton AA: Effect of varying concentrations of follicle stimulating hormone on the production of gonadotrophin surge attenuating factor GnSAF in women. Clin Endocrinol. 1993, 39: 45-50.
Messinis IE, Lolis D, Zikopoulos K, Milingos S, Kollios G, Seferiadis K, Templeton AA: Effect of follicle stimulating hormone or human chorionic gonadotrophin treatment on the production of gonadotrophin surge attenuating factor GnSAF during the luteal phase of the human menstrual cycle. Clin Endocrinol. 1996, 44: 169-175.
Messinis IE, Milingos S, Zikopoulos K, Hasiotis G, Seferiadis K, Lolis D: Luteinizing hormone response to gonadotrophin-releasing hormone in normal women undergoing ovulation induction with urinary or recombinant follicle stimulating hormone. Hum Reprod. 1998, 13: 2415-2420.
Knobil E: The neuroendocrine control of the menstrual cycle. Recent Prog Horm Res. 1980, 36: 53-88.
Ortmann O, Weiss JM, Diedrich K: Embryo implantation and GnRH antagonists: ovarian actions of GnRH antagonists. Hum Reprod. 2001, 164: 608-6011.
Peng C, Fan NC, Ligier M, Vaananen J, Leung PC: Expression and regulation of gonadotropin-releasing hormone GnRH and GnRH receptor messenger ribonucleic acids in human granulosa-luteal cells. Endocrinology. 1994, 135: 1740-1746.
Minaretzis D, Jakubowski M, Mortola JF, Pavlon SN: Gonadotropin-releasing hormone receptor gene expression in human ovary and granulosa lutein cells. J Cl Endocrinol Metab. 1995, 80: 430-434.
Weiss JM, Oltmanns K, GuÈrke EM, Polack S, Eick F, Felberbaum R, Diedrich K, Ortmann O: Actions of gonadotropin-releasing hormone antagonists on steroidogenesis in human granulosa lutein cells. Europ J Endocrinol. 2001, 144: 677-685.
Pellicer A, Tarin JJ, Miro F, Sampaio M, Delos Santos MJ, Remohi J: The use of gonadotropin-releasing hormone analogues GnRHa, in in vitro fertilization: some clinical and experimental investigations of a direct effect on the human ovary. Hum Reprod. 1992, 7: 39-47.
Hamori M, Torok A, Zwirner M, Batteux C, Schinkmann W, Bodis J: In vitro progesterone production of human granulose luteal cells: the impact of different stimulation protocols, poor response and polycystic ovarian syndrome. Hum Reprod. 1992, 7: 592-596.
Dirnfeld M, Goldman S, Gonen Y, Koifman M, Lissak A, Kraiem Z, Abramovici H: Functional differentiation in progesterone secretion by granulosa versus cumulus cells in the human preovulatory follicle and the effect of different induction of ovulation protocols. Fertil Steril. 1993, 60: 1025-1030.
Minaretzis D, Alper MM, Oskowitz SP, Lobel SM, Mortola JF, Pavlon SN: Gonadotropin-releasing hormone antagonist versus agonist administration in women undergoing controlled ovarian hyperstimulation: cycle performance and in vitro steroidogenesis of granulosa lutein cells. Am J Obstet Gynecol. 1995, 172: 1518-1525.
Spona J, Coy DH, Zatlasch E, Wakolbinger C: LH-RH antagonist inhibits gonadal steroid secretion in vitro. Peptides. 1985, 6: 379-382.
Pellicer A, Miro F: Steroidogenesis in vitro of human granulosa-luteal cells pretreated in vivo with gonadotropin-releasing hormone analogs. Fertil Steril. 1990, 54: 590-596.
Mitwally MFM, Casper RF: Effect of in vivo GnRH agonist and GnRH antagonist on hCG and insulin-stimulated progesterone production by human granulosa-lutein cells in vitro. J Assist Reprod Genet. 2002, 198: 384-289.
Garcia-Velasco JA, Isaza V, Vidal C, Landazabal A, Remohi J, Simon C, Pellicer A: Human ovarian steroid secretion in vivo: effects of GnRH agonist versus antagonist cetrorelix. Hum Reprod. 2001, 16: 2533-2539.
Smitz J, Erard P, Camus M, Devroey P, Tournaye H, Wisanto A, Van Steirteghem AC: Pituitary gonadotrophin secretory capacity during the luteal phase in superovulation using GnRH-agonists and HMG in a desensitization or flare-up protocol. Hum Reprod. 1992, 7: 1225-1229.
Beckers NG, Laven JS, Eijkemans MJ, Fauser BC: Follicular and luteal phase characteristics following early cessation of gonadotrophin-releasing hormone agonist during ovarian stimulation for in vitro fertilization. Hum Reprod. 2000, 15: 43-49.
Albano C, Grimbizis G, Smitz J, Riethmuller-Winzen H, Reissmann T, Van Steirteghem A, Devroey P: The luteal phase of non supplemented cycles after ovarian superovulation with human menopausal gonadotropin and the gonadotropin-releasing hormone antagonist Cetrorelix. Fertil Steril. 1998, 70: 357-359.
Felberbaum RE, Reissmann T, Kupker W, Bauer O, al Hasani S, Diedrich C, Diedrich K: Preserved pituitary response under ovarian stimulation with HMG and GnRH antagonists Cetrorelix in women with tubal infertility. Eur J Obstet Gynecol Reprod Biol. 1995, 61: 151-155.
Jones GS: Luteal phase defect: a review of pathophysiology. Curr Opin Obstet Gynecol. 1991, 3: 641-668.
Silverman AY, Smith CG, Siler-Khodr TM, Asch RH: Human chorionic gonadotropin blocks the estrogen-induced luteinizing hormone release in long-term castrated rhesus monkeys: evidence for an ultrashort-loop negative feedback. Fertil Steril. 1981, 35: 74-78.
Miyake A, Aono T, Kinugasa T, Tanizawa O, Kurachi K: The time course change after castration in short-loop negative feedback control of LH by HCG in women. Acta Endocrinol Copenh. 1978, 88: 1-6.
Kyle CV, Griffin J, Jarrett A, Odell WD: Inability to demonstrate an ultrashort loop feedback mechanism for luteinizing hormone in humans. J Clin Endocrinol Metab. 1989, 69: 170-176.
Nader S, Berkowitz AS: Endogenous luteinizing hormone surges following administration of human chorionic gonadotropin: further evidence for lack of loop feedback in humans. J Assist Reprod Genet. 1992, 9: 124-127.
Mores N, Krsmanovic LZ, Catt KJ: Activation of LH receptors expressed in GnRH neurons stimulates cyclic AMP production and inhibits pulsatile neuropeptidase release. Endocrinology. 1996, 137: 5731-5734.
Sengoku K: Present state and future in reproductive medicine. Hokkaido Igaku Zasshi. 2000, 75 (4): 237-242.
Nargund G, Waterstone J, Bland JM, Philips Z, Parsons J, Campbell S: Cumulative conception and live birth rates in natural unstimulated IVF cycles. Hum Reprod. 2001, 16: 259-262.
Tan SL, Royston P, Campbell S, Jacobs HS, Betts J, Mason B, Edwards RG: Cumulative conception and livebirth rates after in-vitro fertilisation. The Lancet. 1992, 339: 1390-1394.
Corfman RS, Milad MP, Bellavance TL, Ory SJ, Erickson LD, Ball GD: A novel ovarian stimulation protocol for use with the assisted reproductive technologies. Fertil Steril. 1993, 60: 864-870.
Williams SC, Gibbons WE, Muasher SJ, Oehninger S: Minimal ovarian hyperstimulation for in vitro fertilization using sequential clomiphene citrate and gonadotropin with or without the addition of a gonadotropin-releasing hormone antagonist. Fertil Steril. 2002, 785: 1068-1072.
Toner JP, Brzyski RG, Oehninger S, Veeck LL, Simonetti S, Muasher SJ: Combined impact of the number of pre-ovulatory oocytes and cryopreservation on IVF outcome. Hum Reprod. 1991, 6: 284-289.
Simon C, Garcia Velasco JJ, Valbuena D, Peinado JA, Moreno C, Remohi J, Pellicer A: Increasing uterine receptivity by decreasing estradiol levels during the preimplantation period in high responders with the use of a follicle-stimulating hormone step-down regimen. Fertil Steril. 1998, 70: 234-239.
Tortoriello DV, McGovern PG, Colon JM, Skurnick JH, Lipetz K, Santoro N: Coasting" does not adversely affect cycle outcome in a subset of highly responsive in vitro fertilization patients. Fertil Steril. 1998, 693: 454-460.
Anonymous: Making medicines affordable: the prize factor position paper. Am J Public Health. 2001, 91: 486-490.
Zwart-van Rijkom JE, Broekmans FJ, Leufkens HG: From HMG through purified urinary FSH preparations to recombinant FSH: a substitution study. Hum Reprod. 2002, 174: 857-865.
Daya S, Gunby J, Hughes EG, Collins JA, Sagle MA: Follicle-stimulating hormone versus human menopausal gonadotropin for in vitro fertilization cycles: a meta-analysis. Fertil Steril. 1995, 64: 347-354.
Agrawal R, Holmes J, Jacobs HS: Follicle-stimulating hormone or human menopausal gonadotrophin for ovarian stimulation in in-vitro fertilization cycles: a meta-analysis. Fertil Steril. 2000, 73: 338-343.
Agrawal R, Jacobs HS: The challenge of meta-analysis and the need to register clinical trials. Fertil Steril. 2000, 74: 420-422.
Balasch J: The challenge of meta-analysis and the need to register clinical trials. Fertil Steril. 2000, 74: 420-
Out HJ, Driessen SG, Mannaerts BM, Coelingh Bennink HJ: Recombinant follicle-stimulating hormone follitropin beta, Puregon yields higher pregnancy rates in in vitro fertilization than urinary gonadotropins. Fertil Steril. 1997, 68: 138-142.
Daya S, Gunby J: Recombinant versus urinary follicle stimulating hormone for ovarian stimulation in assisted reproduction. Hum Reprod. 1999, 4: 2207-2215.
Out HJ, Mannaerts BM, Driessen SG, Coelingh Bennink HJ: Recombinant follicle stimulating hormone rFSH; Puregon in assisted reproduction: more oocytes, more pregnancies. Results from five comparative studies. Hum Reprod Update. 1996, 2: 162-171.
Gordon UD, Harrison RF, Fawzy M, Hennelly B, Gordon AC: A randomized prospective assessor-blind evaluation of luteinizing hormone dosage and in vitro fertilization outcome. Fertil Steril. 2001, 75: 324-331.
Strehler E, Abt M, El-Danasouri I, De Santo M, Sterzik K: Impact of recombinant follicle-stimulating hormone and human menopausal gonadotropins on in vitro fertilization outcome. Fertil Steril. 2001, 75: 332-336.
Weissman A, Lurie S, Zalel Y, Goldchmit R, Shoham Z: Human chorionic gonadotropin: pharmacokinetics of subcutaneous administration. Gynecol Endocrinol. 1996, 10: 273-276.
Yen SSC, Llenera G, Little B, Pearson O: Disappearance rate of endogenous luteinizing hormone and chorionic gonadotropin in man. J Clin Endocrinol Metab. 1968, 28: 1763-1767.
Fritz MA, McLachlan RI, Cohen NL, Dahl KD, Bremner WJ, Soules MR: Onset and characteristics of the midcycle surge in bioactive and immunoactive luteinizing hormone secretion in normal women: influence of physiological variations in periovulatory ovarian steroid hormone secretion. J Clin Endocrinol Metab. 1992, 75: 489-493.
Diczfalusy E, Harlin J: Clinical-pharmacological studies on human menopausal gonadotropin. Hum Reprod. 1988, 3: 21-27.
Damewoad MD, Shen W, Zacur HA, Schlaff WD, Rock JA, Wallach EE: Disappearance of exogenously administered human chorionic gonadotropin. Fertil Steril. 1989, 52: 398-400.
Loumaye E, Campbell R, Salat-Baroux J: Human follicle stimulating hormone produced by recombinant DNA technology: a review for clinicians. Hum Reprod Update. 1995, 1: 188-199.
Porchet HC, Lo Cotonnec JY, Nouteboom B, Canali S, Zanolo G: Pharmacokinetics of recombinant human luteinizing hormone after intravenous, intramuscular, and subcutaneous administration in monkeys and comparison with intravenous administration of pituitary human luteinizing hormone. J Clin Endocrinol Metab. 1995, 80: 667-673.
Le Cotonnec JY, Porchet HC, Beltrami V, Munafo A: Clinical pharmacology of recombinant human luteinizing hormone. Part I. Pharmacokinetics after intravenous administration to healthy female volunteers and comparison with urinary human luteinizing hormone. Fertil Steril. 1998, 69: 189-194.
Le Cotonnec JY, Loumaye E, Porchet HC, Beltrami V, Munafo A: Pharmacokinetic and pharmacodynamic interactions between recombinant human luteinizing hormone and recombinant human follicle stimulating hormone. Fertil Steril. 1998, 69: 201-209.
The European Recombinant Human LH Study Group : Recombinant human luteinizing hormone LH to support recombinant human follicle stimulating hormone FSH-induced follicular development in LH- and FSH-deficient anovulatory women: a dose-finding study. J Clin Endocrinol Metab. 1998, 83: 1507-1514.
European Recombinant LH Study Group: Human Recombinant Luteinizing Hormone Is as Effective as, But Safer Than, Urinary Human Chorionic Gonadotropin in Inducing Final Follicular Maturation and Ovulation in in Vitro Fertilization Procedures: Results of a Multicenter Double-Blind Study. J C Endocrinol Metab. 2001, 86: 2607-2618.
Chandrasekher YA, Hutchison JS, Zelinski Wooten MB, Hess DL, Wolf DP, Stouffer RL: Initiation of periovulatory events in primate follicles using recombinant and native human luteinizing hormone to mimic the midcycle gonadotropin surge. J Clin Endocrinol Metab. 1994, 79: 298-306.
Huirne JA, Lambalk CB: Gonadotropin-releasing-hormone-receptor antagonists. Lancet. 2001, 3589: 1793-1803.
Olivennes F, Alvarez S, Bouchard P, Fanchin R, Salat-Baroux J, Frydman R: The use of a GnRH antagonist cetrorelix in a single dose protocol in IVF-embryo transfer: a dose finding study of 3 versus 2 mg. Hum Reprod. 1998, 13: 2411-2414.
Albano C, Smitz J, Camus M, Riethmuller-Winzen H, Van Steirteghem A, Devroey P: Comparison of different doses of gonadotropin-releasing hormone antagonist cetrorelix during controlled ovarian hyperstimulation. Fertil Steril. 1997, 67: 917-922.
Albano C, Felberbaum RE, Smitz J, Riethmuller-Winzen H, Engel J, Diedrich K, Devroey P: Ovarian stimulation with HMG: results of a prospective randomized phase III European study comparing the luteinizing hormone-releasing hormone LHRH-antagonist cetrorelix and the LHRH-agonist buserelin. Hum Reprod. 2000, 15: 526-531.
The European Orgalutran Study Group, Borm G, Mannaerts B: Treatment with the gonadotrophin-releasing hormone antagonist ganirelix in women undergoing ovarian stimulation with recombinant follicle stimulating hormone is effective, safe and convenient: results of a controlled, randomized, multicentre trial. Hum Reprod. 2000, 15: 1490-1498.
Olivennes F, Belaisch-Allart J, Emperaire JC, Dechaud H, Alvarez S, Moreau L, Nicollet B, Zorn JR, Bouchard P, Frydman R: Prospective, randomized, controlled study of in vitro fertilization-embryo transfer with a single dose of a luteinizing hormone-releasing hormone LH-RH antagonist cetrorelix or a depot formula of an LH-RH agonist triptorelin. Fertil Steril. 2000, 73: 314-320.
Al-Inany H, Aboulghar M: GnRH antagonist in assisted reproduction. Cochrane review. The Cochrane Library. 2001, 4
Christin-Maitre S, Olivennes F, Dubourdieu S, Chabbert-Buffet N, Charbonnel B, Frydman R, Bouchard P: Effect of gonadotrophin-releasing hormone GnRH antagonist during the LH surge in normal women and during controlled ovarian hyperstimulation. Clin Endocrinol Oxf. 2000, 52: 721-726.
Itskovitz-Eldor J, Kol S, Mannaerts B: Use of a single bolus of GnRH agonist triptorelin to trigger ovulation after GnRH antagonist ganirelix treatment in women undergoing ovarian stimulation for assisted reproduction, with special reference to the prevention of ovarian hyperstimulation syndrome: preliminary report: short communication. Hum Reprod. 2000, 15: 1965-1968.
Olivenne F, Ayoubi JM, Fanchin R, Rongieres-Bertrand C, Hamamah S, Bouchard P, Frydman R: GnRH antagonist in single-dose applications. Hum Reprod Update. 2000, 6: 313-317.
Fulghesu AM, Villa P, Pavone V, Guido M, Apa R, Caruso A, Lanzone A, Rossodivita A, Mancuso S: The impact of insulin secretion on the ovarian response to exogenous gonadotropins in polycystic ovary syndrome. J Clin Endocrinol Metab. 1997, 82: 644-648.
Cano F, Garcia-Velasco JA, Millet A, Remohi J, Simon C, Pellicer A: Oocyte quality in polycystic ovaries revisited: identification of a particular subgroup of women. J Assist Reprod Genet. 1997, 14: 254-261.
Clark AM, Thornley B, Tomlinson L, Galletley C, Norman RJ: Weight loss in obese infertile women results in improvement in reproductive outcome for all forms of fertility treatment. Hum Reprod. 1998, 13: 1502-1505.
Ehrmann DA: Insulin-lowering therapeutic modalities for polycystic ovary syndrome. Endocrinol. Metab Clin North Am. 1999, 28: 423-438.
Mitwally MFM, Kuscu NK, Yalcinkaya TM: High ovulatory rates with use of troglitazone in clomiphene-resistant women with polycystic ovary syndrome. Hum. Reprod. 1999, 14: 2700-2703.
Fedorcsák P, Dale PO, Storeng R, Tanbo T, Åbyholm T: The impact of obesity and insulin resistance on the outcome of IVF or ICSI in women with polycystic ovarian syndrome. Hum Reprod. 2001, 16: 1086-1091.
Stadtmauer LA, Riehl RM, Toma SK, Huang S, Barker S, Talbert LM: Metformin treatment of patients with polycystic ovarian syndrome undergoing IVF increases the number of mature oocytes, the fertilization rate and the number of embryos with changes in the levels of insulin-like growth factors. Abstracts of the American Society for Reproductive Medicine. 1999, 72: S12-
Stadtmauer LA, Toma SK, Riehl RM, Talbert LM: Impact of metformin therapy on ovarian stimulation and outcome in 'coasted' patients with polycystic ovary syndrome undergoing in-vitro fertilization. Reprod Biomed Online. 2002, 52: 112-116.
Jones GES, Howard JE, Langford H: The use of cortisone in follicular phase disturbances. Fertil Steril. 1953, 4: 49-62.
Greenblatt RB, Barfield W, Lampros CP: Cortisone in the treatment of infertility. Fertil Steril. 1956, 7: 203-212.
Lobo RA, Paul W, March CM, Granger L, Kletzky OA: Clomiphene and dexamethasone in women unresponsive to clomiphene alone. Obstet Gynecol. 1982, 60: 497-501.
Evron S, Navot D, Laufer N, Diamant YZ: Induction of ovulation with combined human gonadotropins and dexamethasone in women with polycystic ovarian disease. Fertil Steril. 1983, 40: 183-186.
Daly DC, Walters CA, Soto-Albors CE, Tohan N, Riddick DH: A randomized study of dexamethasone in ovulation induction with clomiphene citrate. Fertil Steril. 1984, 41: 844-848.
Singh KB, Dunnihoo DR, Mahajan DK, Bairnsfather LE: Clomiphene-dexamethasone treatment of clomiphene-resistant women with and without the polycystic ovary syndrome. J Reprod Med. 1992, 37: 215-218.
Bider D, Amoday I, Tur-Kaspa I, Livshits A, Dor J: The addition of glucocorticoid to the protocol of programmed oocyte retrieval for in-vitro fertilization – a randomized study. Hum Reprod. 1996, 11: 1606-1608.
Kemeter P, Feichtinger W: Prednisolone supplementation to clomid and/or gonadotrophin stimulation for in-vitro fertilization – a prospective randomized trial. Hum Reprod. 1986, 1: 441-444.
Rein MS, Jackson KV, Sable DB, Thomas PP, Hornstein MD: Dexamethasone during ovulation induction for in-vitro fertilization: a pilot study. Hum Reprod. 1996, 11: 253-255.
Fridstrom M, Carlstrom K, Sjoblom P, Hillensjo T: Effect of prednisolone on serum and follicular fluid androgen concentrations in women with polycystic ovary syndrome undergoing in-vitro fertilization. Hum Reprod. 1999, 146: 1440-1444.
Kutteh WH: Autoimmune factors in assisted reproduction. Minerva Ginecol. 2002, 543: 217-224.
Krusche CA, Herrler A, Classen-Linke I, Beier HM: The progesterone antagonist onapristone retards the advanced endometrial transformation after gonadotropin stimulation in rabbits. Steroids. 2000, 11: 773-782.
Grochowski D: Assisted reproductive medicine in the next age. Ginekol Pol. 2001, 721: 36-43.
Conaghan J, Handyside AH, Winston RM, Leese HJ: Effects of pyruvate and glucose on the development of human preimplantation embryos in vitro. J Reprod Fertil. 1993, 99: 87-95.
Devreker F, Winston RM, Hardy K: Glutamine improves human preimplantation development in vitro. Fertil Steril. 1998, 69: 293-299.
Spanos S, Becker DL, Winston RM, Hardy K: Anti-apoptotic action of insulin-like growth factor I during human preimplantation embryo development. Biol Reprod. 2000, 63: 1413-1420.
Coates A, Rutherford AJ, Hunter H, Leese HJ: Glucose-free medium in human in vitro fertilization and embryo transfer: a large-scale, prospective, randomized clinical trial. Fertil Steril. 1999, 72: 229-232.
Gardner D, Leese HJ: Concentrations of nutrients in mouse oviduct fluid and their effects on embryo development and metabolism in vitro. J Reprod Fert. 1990, 88: 361-368.
Gardner D, Lane M: Culture of viable human blastocysts in defined sequential serum-free media. Hum Reprod. 1998, 13S3: 148-159.
Hardy K: Apoptosis in the human embryo. Rev Reprod. 1999, 4: 125-134.
Giorgetti C, Terriou P, Auquier P, Hans E, Spach JL, Salzmann J, Roulier R: Embryo score to predict implantation after in vitrofertilization: based on 957 single embryo transfers. Hum Reprod. 1995, 10: 2427-2431.
Jamieson ME, Coutts JR, Connor JM: The chromosome constitution of human preimplantation embryos fertilized in vitro. Hum Reprod. 1994, 9: 709-715.
ESHRE PGD Consortium: ESHRE preimplantation genetic diagnosis PGD consortium: data collection II May 2000. Hum Reprod. 2000, 15: 2673-2683.
Levran D, Farhi J, Nahum H, Royburt M, Glezerman M, Weissman A: Prospective evaluation of blastocyst stage transfer vs. zygote intrafallopian tube transfer in patients with repeated implantation failure. Fertil Steril. 2002, 77 (5): 971-977.
Human Fertilisation and Embryology Authority: 2000, Ninth Annual Report HFEA Paxton House, London
Veeck LL, Wortham JW, Witmyer J, Sandow BA, Acosta AA, Garcia JE, Jones GS, Jones HW: Maturation and fertilization of morphologically immature human oocytes in a program of in vitro fertilization. Fertil Steril. 1983, 39: 594-602.
Mikkelson AL, Smith SD, Lindenberg S: In vitro maturation of human oocytes from regularly menstruating women may be successful without follicle stimulating hormone priming. Hum Reprod. 1999, 14: 1847-1851.
Trounson A, Wood C, Kausche A: In vitro maturation and the fertilization and developmental competence of oocytes recovered from untreated polycystic ovarian patients. Fertil Steril. 1994, 62: 353-362.
Cha K-Y, Chian RC: Maturation in vitro of immature human oocytes for clinical use. Hum Reprod Update. 1998, 4: 103-120.
Chian RC, Gulekli B, Buckett WM, Tan SL: Priming with human chorionic gonadotropin before retrieval of immature oocytes in women with infertility due to the polycystic ovary syndrome. N Eng J Med. 1999, 341: 1624-1626.
van de Sandt JJ, Schroeder AC, Eppig JJ: Culture media for mouse oocyte maturation affect subsequent embryonic development. Mol Reprod Dev. 1990, 25: 164-171.
Staessen C, Camus M, Clasen K, De Vos A, Van Steirteghem A: Conventional in vitro fertilization versus intracytoplasmic sperm injection in sibling oocytes from couples with tubal infertility and normozoospermic semen. Hum Reprod. 1999, 14: 2474-2479.
Aboulghar MA, Mansour RT, Serour GI, Amin YM, Kamal A: Prospective controlled randomised study of in vitro fertilization versus intracytoplasmic sperm injection in the treatment of tubal factor infertility with normal semen parameters. Fertil Steril. 1996, 66: 753-756.
Oehninger S, Gosden RG: Should ICSI be the treatment of choice for all cases of in-vitro conception?: No, not in light of the scientific data. Hum Reprod. 2002, 179: 2237-2242.
Ola B, Afnan B, Sharif K, Papaioannou S, Hammadieh N, Barratt CLR: Should ICSI be the treatment of choice for all cases of in-vitro conception?: Considerations of fertilization and embryo development, cost ffectiveness and safety. Hum Reprod. 2001, 1612: 2485-2490.
Nagai S, Kasai T, Hirata S, Hoshi K, Yanagimachi R, Huang T: Cytoplasmic transfer in the mouse in conjunction with intracytoplasmic sperm injection. Reprod Biomed Online. 2004, 81: 75-80.
Cohen J, Scott R, Alikani M, Schimmel T, Munne S, Levron J, Wu L, Brenner C, Warner C, Willadsen S: Ooplasmic transfer in mature human oocytes. Mol Hum Reprod. 1998, 4: 269-280.
Tesarík J, Nagy ZP, Mendoza C, Greco E: Chemically and mechanically induced membrane fusion: non-activating methods for nuclear transfer in mature human oocytes. Hum Reprod. 2000, 15: 1149-1154.
Bulun SE, Sebastian S, Takayama K, Suzuki T, Sasano H, Shozu M: The human CYP19 (aromatase P450) gene: update on physiologic roles and genomic organization of promoters. J Steroid Biochem Mol Biol. 2003, 86 (3–5): 219-24.
Santen RJ, Manni A, Harvey H, Redmond C: Endocrine treatment of breast cancer in women. Endocrine Rev. 1990, 11: 1-45.
Lonning PE: Aromatase inhibition for breast cancer treatment. Acta Oncolog. 1996, 5: 38-43.
Buzdar A, Howell A: Advances in aromatase inhibition: clinical efficacy and tolerability in the treatment of breast cancer. Clin Cancer Res. 2001, 7: 2620-2635.
Brueggemeier RW: Aromatase inhibitors: Mechanisms of steroidal inhibitors. Br Cancer Res Treat. 1994, 30: 31-42.
Coombes RC, Goss P, Dowsett M, Gazet JC, Brodie AMH: 4-hydroxyandrostenedione treatment of postmenopausal patients with advanced breast cancer. The Lancet. 1984, 2: 1237-1239.
Brodie AM, Njar VC: Aromatase inhibitors and breast cancer. Semin Oncol. 1996, 23: 10-20.
Santen RJ, Lipton A, Kendall J: Successful medical adrenalectomy with aminoglutethimide: role of altered drug metabolism. J Am Med Assoc. 1974, 230: 1661-
Lipton A, Santen RJ: Medical adrenalectomy using aminoglutethimide and dexamethasone in advanced breast cancer. Cancer. 1974, 33: 503-512.
Newsome HH, Brown PW, Terz JJ, Lawrence W: Medical and surgical adrenalectomy in patients with advanced breast carcinoma. Cancer. 1977, 39: 542-546.
Winer EP, Hudis C, Burstein HJ, Chlebowski RT, Ingle JN, Edge SB, Mamounas EP, Gralow J, Goldstein LJ, Pritchard KI, Braun S, Cobleigh MA, Langer AS, Perotti J, Powles TJ, Whelan TJ, Browman GP: American Society of Clinical Oncology Technology Assessment on the Use of Aromatase Inhibitors as Adjuvant Therapy for Women With Hormone Receptor-Positive Breast Cancer: Status Report. J Clin Oncol. 2002, 2015: 3317-3327.
Buzdar A, Jonat W, Howell A, Jones SE, Blomqvist C, Vogel CL, Eiermann W, Wolter JM, Azab M, Webster A, Plourde PV: Anastrozole, a potent and selective aromatase inhibitor, versus megestrol acetate in postmenopausal women with advanced breast cancer: results of overview analysis of two phase III trials. J Clin Oncol. 1996, 14: 2000-2011.
Marty M, Gershanovich M, Campos B, Romien G, Lurie H, Bonaventura T: Aromatase imhibitors: a new potent, selective aromatase inhibitor superior to aminoglutethimide AG in postmenopausal women with advanced breast cancer previously treated with antioestrogens. Proc Am Soc Clin Oncol. 1997, 16: 156-
Sing Ranger G: Current concepts in the endocrine therapy of breast cancer: tamoxifen and aromatase inhibitors. J Clin Pharm Ther. 2005, 30 (4): 313-317.
Dranitsaris G, Leung P, Mather J, Oza A: Cost-utility analysis of second-line hormonal therapy in advanced breast cancer: a comparison of two aromatase inhibitors to megestrol acetate. Anticancer Drugs. 2000, 117: 591-560.
Sioufi A, Gauducheau N, Pineau V, Marfil F, Jaouen A, Cardot JM, Godbillon J, Czendlik C, Howald H, Pfister C, Vreeland F: Absolute bioavailability of letrozole in healthy post-menopausal women. Biopharm. Drug Dispos. 1997, 18: 779-789.
Sioufi A, Sandrenan N, Godbillon J, Trunet P, Czendlik C, Howald H, Pfister C, Ezzet F: Comparative bioavailability of letrozole under fed and fasting conditions in 12 healthy subjects after a 2.5 mg single oral administration. Biopharm Drug Dispos. 1997, 186: 489-497.
Weil SJ, Vendola K, Zhou J, Adesanya OO, Wang J, Okafor J, Bondy CA: Androgen receptor gene expression in the primate ovary: cellular localization, regulation, and functional correlations. J Clin Endocrinol Metab. 1998, 837: 2479-2485.
Weil S, Vendola K, Zhou J, Bondy CA: Androgen and follicle-stimulating hormone interactions in primate ovarian follicle development. J Clin Endocrinol Metab. 1999, 848: 2951-2956.
Vendola KA, Zhou J, Adesanya OO, Weil SJ, Bondy CA: Androgens stimulate early stages of follicular growth in the primate ovary. J Clin Invest. 1998, 12: 2622-2629.
Vendola K, Zhou J, Wang J, Famuyiwa OA, Bievre M, Bondy CA: Androgens promote oocyte insulin-like growth factor I expression and initiation of follicle development in the primate ovary. Biol Reprod. 1999, 612: 353-357.
Adashi E: Intraovarian regulation: the proposed role of insulin-like growth factors. Ann NY Acad Sci. 1993, 687: 10-12.
Giudice LC: Insulin-like growth factors and ovarian follicular development. Endocr Rev. 1992, 13: 641-669.
Yen SSC, Laughlin GA, Morales AJ: Interface between extra-and intra-ovarian factors in polycystic ovary syndrome PCOS. Ann NY Acad Science. 1993, 687: 98-111.
Mitwally MFM, Casper RF: The use of an aromatase inhibitor for induction of ovulation in cases of clomiphene citrate failure [abstract]. Program and abstracts of the 16th Annual Meeting of the European Society for Human Reproduction and Embryology ESHRE; Bologna, Italy. 2000, 15: 71-
Mitwally MFM, Casper RF: Aromatase Inhibition: a novel method of ovulation induction in women with polycystic ovarian syndrome. Reprod Technol. 2000, 10: 244-247.
Mitwally MFM, Casper RF: The Aromatase Inhibitor, Letrozole: a Promising Alternative for Clomiphene Citrate for Induction of Ovulation. [Abstract]. Fertil Steril. 2000, 743 (Supplement 1): S35-
Mitwally MFM, Casper RF: Use of an aromatase inhibitor for induction of ovulation in patients with an inadequate response to clomiphene citrate. Fertil Steril. 2001, 75: 305-309.
Hull MGR: The causes of infertility and relative effectiveness of treatment. Infertility. Edited by: Templeton AA, Drife JO. 1992, London: Springer-Verlag, 33-62.
Wysowski DE: Use of fertility drugs in the United States, 1979 through 1991. Fertil Steril. 1991, 60: 1096-1098.
Yagel S, Ben-Chetrit A, Anteby E, Zacut D, Hochner-Celnikier D, Ron M: The effect of ethinyl estradiol on endometrial thickness and uterine volume during ovulation induction by clomiphene citrate. Fertil Steril. 1992, 57: 33-36.
Shimoya K, Tomiyama T, Hashimoto K, Moriyama A, Kawamoto A, Tokugawa Y, Ohashi K, Saji F, Murata Y: Endometrial development was improved by transdermal estradiol in patients treated with clomiphene citrate. Gynecol Obstet Invest. 1999, 474: 251-254.
Gerli S, Gholami H, Manna A, Di Frega D, Vitiello C, Unfer V: Use of ethinyl estradiol to reverse the antiestrogenic effects of clomiphene citrate in patients undergoing intrauterine insemination: a comparative, randomized study. Fertil Steril. 2000, 73: 85-89.
Ben-Ami M, Geslevich Y, Matilsky M, Battino S, Weiner E, Shalev E: Exogenous estrogen therapy concurrent with clomiphene citrate – lack of effect on serum sex hormone levels and endometrial thickness. Gynecol Obstet Invest. 1994, 373: 180-182.
Bateman BG, Nunley WC, Kolp LA: Exogenous estrogen therapy for treatment of clomiphene citrate-induced cervical mucus abnormalities: is it effective?. Fertil Steril. 1990, 54: 577-579.
CH WU, Winkel CA: The effect of therapy initiation day on clomiphene citrate therapy. Fertil Steril. 1989, 52: 564-568.
Saleh A, Biljan MM, Tan SSSL, Tulandi T: Effects of Tamoxifen Tx on Endometrial Thickness and Pregnancy Rates in Women Undergoing Superovulation with Clomiphene Citrate CC and Intrauterine Insemination IUI. Fertil Steril. 2000, 74S1: S90-
Boostanfar R, Jain JK, Paulson RJ, Mishell DR: A Prospective Randomized Trial Comparing Clomiphene Citrate with Tamoxifen for Ovulation Induction in Anovulatory Women. Fertil Steril. 2000, 74S1: S62-
Mitwally MFM, Casper RF: The aromatase inhibitor, letrozole, decreases FSH dose required for ovarian superovulation. Proceedings of the 46th Annual Meeting of the Canadian Fertility and Andrology Society;. 2000, [Abstract], September Newfoundland, Canada
Mitwally MFM, Casper RF: Aromatase inhibition reduces gonadotrophin dose required for controlled ovarian stimulation in women with unexplained infertility. Hum Reprod. 2003, 18 (8): 1588-1597.
Mitwally MFM, Casper RF: Aromatase inhibition reduces the dose of gonadotropin required for controlled ovarian hyperstimulation. J Soc Gynecol Investig. 2004, 11 (6): 406-415.
Mitwally MFM, Casper RF: Aromatase inhibition improves ovarian response to FSH: a potential option for low responders during ovarian stimulation. Fertil Steril. 2001, 754S1: S8-S9.
Mitwally MFM, Casper RF: Aromatase inhibition improves ovarian response to follicle-stimulating hormone in poor responders. Fertil Steril. 2002, 774: 776-780.
Mitwally MFM, Casper RF: Single-dose administration of an aromatase inhibitor for ovarian stimulation. Fertil Steril. 2005, 83 (1): 229-231.
Sammour A, Biljan MM, Tan SL, Tulandi T: Prospective randomized trial comparing the effects of letrazole LE and clomiphene citrate CC on follicular development, endometrial thickness and pregnancy rate in patients undergoing super-ovulation prior to intrauterine insemination IUI. Fertil Steril. 2001, 76 (S1): S110-
Biljan MM, Tan SL, Tulandi T: Prospective randomized trial comparing the effects of 2.5 and 5.0 mg of letrozole LE on follicular development, endometrial thickness and pregnancy rate in patients undergoing super-ovulation. Fertil Steril. 2002, 78S1: S55-
El Helw B, El Sadek M, Matar H, Fouad S, El Nomrosy K, Abbas H: Single dose Letrozole versus Clomiphene citrate for superovulation prior to intrauterine insemination: a prospective randomised study. [abstract]. Program and abstracts of the 18th Annual Meeting of the European Society for Human Reproduction and Embryology ESHRE; Vienna, Austria. 2002, 17 (75): O-209-
Healey S, Sylvestre M, Tan SL, Tulandi S, Biljan M: A comparison between superovulation with FSH and a combination of FSH and letrozole. Fertil Steril. 2002, 78 (Supplement 1): S55-S56.
Prapas G, Pistofidis , Tika M, Pelekanos M: Experience with the use of an aromatase inhibitor letrozolein patients with a history of poor ovarian responce in previous assisted reproduction cycles. [abstract]. Program and abstracts of the 18th Annual Meeting of the European Society for Human Reproduction and Embryology ESHRE; Vienna, Austria. 2002, 17 (75): O-098-[Abstract]
Schoolcraft W, Surrey W, Minjarez D, Gardner D: Antagonist/letrozole protocol for patients failing microdose agonist flare stimulation. Fertil Steril. 2002, 78S1: S234-
Al-Omari WR, Al-Hadithi N, Sulaiman WR, Izat B: Comparison of two aromatase inhibitors in clomiphene resistant PCOS. [abstract]. Program and abstracts of the 18th Annual Meeting of the European Society for Human Reproduction and Embryology ESHRE; Vienna, Austria. 2002, 17 (75): O-243-
Krasnopolskaya K, Kaluina A: Application of aromatase inhibitors Anastrosol in IVF program for the treatment of infertility associated with severe endometriosis. [abstract]. Program and abstracts of the 18th Annual Meeting of the European Society for Human Reproduction and Embryology ESHRE; Vienna, Austria. 2002, 17 (75): P-438-
Al-Fozan H, Al-Khadouri M, Tan SL, Tulandi T: A randomized trial of letrozole versus clomiphene citrate in women undergoing superovulation. Fertil Steril. 2004, 82 (6): 1561-3.
Goswami SK, Das T, Chattopadhyay R, Sawhney V, Kumar J, Chaudhury K, Chakravarty BN, Kabir SN: A randomized single-blind controlled trial of letrozole as a low-cost IVF protocol in women with poor ovarian response: a preliminary report. Hum Reprod. 2004, 19 (9): 2031-2035.
Al-Omari WR, Sulaiman WR, Al-Hadithi N: Comparison of two aromatase inhibitors in women with clomiphene-resistant polycystic ovary syndrome. Int J Gynaecol Obstet. 2004, 85 (3): 289-291.
Fatemi HM, Kolibianakis E, Tournaye H, Camus M, Van Steirteghem AC, Devroey P: Clomiphene citrate versus letrozole for ovarian stimulation: a pilot study. Reprod Biomed Online. 2003, 7 (5): 543-546.
Healey S, Tan SL, Tulandi T, Biljan MM: Effects of letrozole on superovulation with gonadotropins in women undergoing intrauterine insemination. Fertil Steril. 2003, 80 (6): 1325-1329.
Tredway DR, Buraglio M, Hemsey G, Denton G: A phase I study of the pharmacokinetics, pharmacodynamics, and safety of single- and multiple-dose anastrozole in healthy, premenopausal female volunteers. Fertil Steril. 2004, 82 (6): 1587-1593.
Hoff JD, Quigley ME, Yen SSC: Hormonal dynamics at midcycle: a reevalution. J Clin Endocrinol Metab. 1983, 57: 792-796.
Park SJ, Goldsmith LT, Weiss G: Age-related changes in the regulation of luteinizing hormone secretion by estrogen in women. Exp Biol Med Maywood. 2002, 2277: 455-464.
Knobil E: On the control of gonadotropin secretion in the rhesus monkey. Rec Prog Horm Res. 1974, 30: 1-46.
Hughes EG, Fedorkow DM, Daya S, Sagle MA, van de Koppel P, Collins JA: The routine use of gonadotropin-releasing hormone agonists prior to in vitro fertilization and gamete intrafallopian transfer: a meta-analysis of randomized controlled trials. Fertil Steril. 1992, 58: 888-896.
Diedrich K, Diedrich C, Santos E, Zoll C, al-Hasani S, Reissmann T, Krebs D, Klingmuller D: Suppression of the endogenous luteinizing hormone surge by the gonadotrophin-releasing hormone antagonist cetrorelix during ovarian stimulation. Hum Reprod. 1994, 9: 788-791.
Felberbaum R, Diedrich K: Ovarian stimulation for in-vitro fertilization/intracytoplasmic sperm injection with gonadotrophins and gonadotrophin-releasing hormone analogues: agonists and antagonists. Hum Reprod. 1999, 14 (S1): 207-221.
Bulun SE, Fang Z, Imir G, Gurates B, Tamura M, Yilmaz B, Langoi D, Amin S, Yang S, Deb S: Aromatase and endometriosis. Semin Reprod Med. 2004, 22 (1): 45-50.
Palter S, Tavares AB, Hourvitz A, Veldhuis JD, Adashi EY: Are Estrogens of Import to Primate/Human Ovarian Folliculogenesis?. Endocr Rev. 2001, 22: 389-424.
Yanase T, Simpson ER, Waterman M: 17-hydroxylase/17,20-lyase deficiency: from clinical investigation to molecular definition. Endocr Rev. 1991, 12: 91-108.
Mallin SR: Congenital adrenal hyperplasia secondary to 17-hydroxylase deficiency: two sisters with amenorrhea, hypokalemia, hypertension and cystic ovaries. Ann Intern Med. 1968, 70: 69-73.
Araki S, Chikazawa K, Sekiguchi I, Yamauchi H, Motoyama M, Tamada T: Arrest of follicular development in a patient with 17-hydroxylase deficiency: folliculogenesis in association with a lack of estrogen synthesis in the ovaries. Fertil Steril. 1987, 47: 169-172.
Nagamani M, Dinh TV: 17-Hydroxylase deficiency in genetic females. A report of two cases. J Reprod Med. 1986, 31: 734-738.
Roger M, Merceron RE, Girard F, Canlorbe P, Dehennin L, Konopka P, Seneze J, Toublanc JE: Dexamethasone-suppressible hypercorticosteronism in 46XX subjects with ambiguous genitalia and ovarian cysts. Partial defect of 17-hydroxyase or 17,20-desmolase. Horm Res. 1982, 16: 23-
Rabinovici J, Blankstein J, Goldman B, Rudak E, Dor Y, Pariente C, Geir A, Lunenfeld B, Mashiac S: IVF and primary embryonic cleavage are possible in 17-hydroxylase deficiency despite extremely low intrafollicular 17β estradiol. J Clin Encrinol Metab. 1989, 68: 693-697.
Geier A, Lunenfeld B, Pariente C, Kotev-Emeth S, Shadmi A, Kokia E, Blankstein J: Estrogen receptor binding material in blood of patients following clomiphene citrate administration: determination by radioreceptor assay. Fertil Steril. 1987, 47: 778-784.
Pariente C, Rabinovici J, Lunenfeld B, Rudak E, Dor J, Mashiach S, Levran D, Blankstein J, Geier A: Steroid secretion by granulosa cells isolated from a woman with 17-hydroxylase deficiency. J Clin Endocrinol Metab. 1990, 71: 984-987.
Zelinski-Wooten MB, Hess DL, Wolf DP, Stouffer RL: Steroid reduction during ovarian stimulation impairs oocyte fertilization, but not folliculogenesis, in rhesus monkeys. Fertil Steril. 1994, 61: 1147-1155.
Mannaerts B, Uilenbroek J, Schot P, DeLeeuw R: Folliculogenesis in hypophysectomized rats after treatment with recombinant human follicle-stimulating hormone. Biol Reprod. 1994, 51: 72-81.
Ito Y, Fisher CR, Conte FA, Grambach MM, Simpson ER: Molecular basis of aromatase deficiency in an adult female with sexual infantilism and polycystic ovaries. Proc Natl Acad Sci. 1993, 90: 11673-11677.
Conte FA, Grumbach MM, Ito Y, Fisher CR, Simpson ER: A syndrome of female pseudohermaphrodism, hypergonadotropic hypogonadism, and multicystic ovaries associated with missense mutations in the gene encoding aromatase P450 arom. J Clin Endocrinol Metab. 1994, 78: 1287-1292.
Morishima A, Grumbach MM, Simpson ER, Fisher C, Quin K: Aromatase deficiency in male and female siblings caused by a novel mutation and the physiological role of estrogens. J Clin Endocrinol Metab. 1995, 80: 3689-3698.
Mullis PE, Yoshimura N, Kuhlmann B, Lippuner K, Jaeger P, Harada H: Aromatase deficiency in a female who is compound heterozygote for two new point mutations in the P450 arom gene: impact of estrogens on hypergonadotropic hypogonadism, multicystic ovaries, and bone densitometry in childhood. J Clin Endocrinol Metab. 1997, 82: 1739-1745.
Bulun SE: Aromatase deficiency in women and men: would you have predicted the phenotypes?. J Clin Endocrinol Metab. 1996, 81: 867-871.
Couzinet B, Lestrat N, Brailly S, Forest M, Schaison G: Stimulation of ovarian follicular maturation with pure follicle-stimulating hormone in women with gonadotropin deficiency. J Clin Endocrinol Metab. 1988, 66: 552-556.
Schoot DCJM, Herjan JT, Bennink C, Mannaerts B, Lamberts SWJ, Bouchard P, Fauser BC: Human recombinant FSH induced growth of preovulatory follicles without concomitant increase in androgen and estrogen biosynthesis in a woman with isolated GT deficiency. J Clin Endocrinol Metab. 1992, 74: 1471-1473.
Mannaerts B, Leeuw R, VanRavestein JG, Wezenbeek P, Schuurs A, Kloosterboer H: Comparative in vitro and in vivo studies on the biological characteristics of human recombinant follicle-stimulating hormone. Endocrinology. 1991, 129: 2623-2630.
Shoham Z, Mannaerts B, Insler V, Coelingh HB: Induction of follicular growth using recombinant human follicle-stimulating hormone in two volunteer women with hypogonadotropic hypogonadism. Fertil Steril. 1993, 59: 738-742.
Shoham Z, Balen A, Patel A, Jacobs HS: Results of ovulation induction using human menopausal gonadotropins or purified follicle-stimulating hormone in hypogonadotropic hypogonadism patients. Fertil Steril. 1991, 56: 1048-1053.
Zelinski-Wooten MB, Hess DL, Baughman WO, Molskness TA, Wolf DP, Stouffer RL: Administration of an aromatase inhibitor during the late follicular phase of gonadotropin-treated cycles in Rhesus monkeys: effects on follicle development, oocyte maturation, and subsequent luteal function. J Clin Endocrinol Metab. 1993, 76: 988-995.
Selvaraj N, Shetty G, Vijayalakshmi K, Bhatnagar AS, Moudgal NR: Effect of blocking oegrogen synthesis with a new generation aromatase inhibitor CGS 16949A on follicular maturation induced by pregnant mare serum gonadotrophin in the immature rat. J Endocrinol. 1994, 142: 563-570.
Selvaraj J, Bhatnagar AS, Moudgal NR: Aromatase inhibition during follicular phase in Bonnet monkeys: is there a role for estrogen in follicular maturation in the primate?. Endocrine. 1995, 3: 245-249.
Shetty G, Krishnamurthy H, Krishnamurthy HN, Bhatnagar S, Moudgal RN: Effect of estrogen deprivation on the reproductive physiology of male and female primates. J Steroid Biochem Mol Biol. 1997, 613-6: 157-166.
Medina M, Herrera J, Flores M, Martin O, Bermudez JA, Zarate A: Normal ovarian function in a mild form of late-onset 3 beta-hydroxysteroid dehydrogenase deficiency. Fertil Steril. 1986, 466: 1021-1025.
Fisher CR, Graves KH, Parlow AF, Simpson ER: Characterization of mice deficient in aromatase ArKO because of targeted disruption of the cyp19 gene. Proc Natl Acad Sci. 1998, 95: 6965-6970.
Britt KL, Drummond AE, Cox VA, Dyson M, Wreford NG, Jones ME, Simpson ER, Findlay JK: An age-related ovarian phenotype in mice with targeted disruption of the Cyp 19 aromatase gene. Endocrinology. 2000, 141: 2614-2623.
Miyaura C, Toda K, Inada M, Ohshiba T, Matsumoto C, Okada T, Ito M, Shizuta Y, Ito A: Sex- and age-related response to aromatase deficiency in bone. Biochem Biophys Res Commun. 2001, 280: 1062-1068.
Toda K, Saibara T, Okada T, Onishi S, Shizuta Y: A loss of aggressive behaviour and its reinstatement by oestrogen in mice lacking the aromatase gene Cyp19. J Endocrinol. 2001, 168: 217-220.
Simpson ER: Genetic mutations resulting in loss of aromatase activity in humans and mice. J Soc Gynecol Invest. 2000, 7: S18-21.
Jones MEE, Kim Huynh , Jones Gayle, Thouas George, Trounson Alan, Evan R: Oocytes Do Not Require Estrogen To Mature or To Retain Developmental Competence. ENDO. 2002, San Francisco, California, 84th Annual Meeting June 19 – 22, 2002
Paesi FJA: The effect of small doses of oestrogen on the ovary of the immature rat. Acta Endocrinol Copenh. 1952, 11: 251-268.
Terry YY, Tong , Victor HH, Goh : Effects of 4-hydroxyandrostenedione and hyperstimulation with pregnant mare serum gonadotrophin on early embryonic development in rats. Can J Physiol Pharmacol. 2001, 79: 744-753.
Hu Y, Cortvrindt R, Smitz J: Effects of aromatase inhibition on in vitro follicle and oocyte development analyzed by early preantral mouse follicle culture. Mol Reprod Dev. 2002, 614: 549-59.
Guo Y, Guo KJ, Huang L, Tong XG, Li X: Effect of estrogen deprivation on follicle/oocyte maturation and embryo development in mice. Chin Med J Engl. 2004, 1174: 498-502.
Mitwally MF, Biljan MM, Casper RF: Pregnancy outcome after the use of an aromatase inhibitor for ovarian stimulation. Am J Obstet Gynecol. 2005, 192 (2): 381-386.
De Neubourg D, Gerris J, Mangelschots K, Van Royen E, Steylemans A, Vercruyssen M, Elseviers M, Gerris J: The obstetrical and neonatal outcome of babies born after single embryo transfer in IVF/ICSI does not compare unfavorably to spontaneous conceived babies. Abstract O-151, ESHRE. 2005, i56-
De Neubourg D, Gerris J, Mangelschots K, Van Royen E, Vercruyssen M, Elseviers M: Single top quality embryo transfer as a model for prediction of early pregnancy outcome. Hum Reprod. 2004, 19: 1476-1479.
De Sutter P, Van der Elst J, Coetsier T, Dhont M: Single embryo transfer and multiple pregnancy rate reduction in IVF/ICSI: a 5-year appraisal. Reprod Biomed Online. 2003, 6: 464-469.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
MFMM has written the review article, RFC and MPD have reviewed the manuscript
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Mitwally, M.F., Casper, R.F. & Diamond, M.P. The role of aromatase inhibitors in ameliorating deleterious effects of ovarian stimulation on outcome of infertility treatment. Reprod Biol Endocrinol 3, 54 (2005). https://doi.org/10.1186/1477-7827-3-54
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1477-7827-3-54