
Abstract
Prostate cancer (PCa) is no exception to the multi-step process of metastasis. As PCa progresses, changes occur within the microenvironments of both the malignant cells and their targeted site of metastasis, enabling the necessary responses that result in successful translocation. The majority of patients with progressing prostate cancers develop skeletal metastases. Despite advancing efforts in early detection and management, there remains no effective, long-term cure for metastatic PCa. Therefore, the elucidation of the mechanism of PCa metastasis and preferential establishment of lesions in bone is an intensive area of investigation that promises to generate new targets for therapeutic intervention. This review will survey what is currently know concerning PCa interaction with the extracellular matrix (ECM) and the roles of factors within the tumor and ECM microenvironments that contribute to metastasis. These will be discussed within the context of changes in expression and functional heterodimerization patterns of integrins, changes in ECM expression and reorganization by proteases facilitating invasion. In this context we also provide a brief summary of how growth factors (GFs), cytokines and regulatory signaling pathways favor PCa metastasis to bone.
Similar content being viewed by others

Background
Prostate cancer (PCa) is the second most common malignancy in men worldwide [1]. In the United States for 2003 there are expected to be 220,900 new diagnoses and approximately 28,900 PCa deaths [2]. Notably, it has been determined that ~90% of patients with advanced PCa will develop osseous metastases [3, 4]. Similar studies on PCa patients at autopsy have found ≥ 80% of patients have established macrometastases involving bone, and of these bone lesions ≥ 90% will have an osteoblastic phenotype [5, 6]. The characteristics of clinical presentation with bone involvement include severe pain, pathologic fractures and spinal cord compression [7]. Typically these patients have a mean survival time of nine months to one year [8]. Thus, once PCa metastasizes to the bone it is difficult to eradicate.
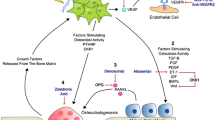
Metastasis requires the interaction of malignant cells with three distinct microenvironments 1) the primary organ, 2) the circulation, and 3) the target organ where a metastatic lesion will develop [9–11]. Both soluble and insoluble stromal elements within these microenvironments are involved in the metastatic cascade [11]. Successful metastasis requires that several well-documented steps be followed. Initially, angiogenesis must be induced to meet the nutrient needs of the growing cancer cells and facilitate removal of toxic waste products [12, 13]. The metastatic cells must then degrade or remodel basement membrane, detach from the primary tumor mass and intravasate. Tumor cells must next survive the stress of vascular transportation and evade host defense mechanisms [12]. Attachment to their preferred site of metastasis may be either targeted, via tissue-specific microvessel cell adhesion molecules (CAMs) [14, 15], or more general in nature simply allowing the tumor cells to reside in the target tissue long enough to respond to transendothelial soluble factors from the target organ (Sikes, unpublished data) [11]. Tumor cells then extravasate into the target organ parenchyma, where they proceed to proliferate in the new, supportive microenvironment as micrometastases. Finally, the micrometastases must induce angiogenesis to support growth of the new lesion [16].
Despite intensive research efforts very little is known about the specific mechanism(s) that contribute to the predominant pattern of PCa metastasis and establishment of bony lesions (Sikes, unpublished data). Various theories have been proposed, including venous drainage, lymphatic spread, and arterial emboli [5]. One of the oldest, for example, is based on anatomical juxtaposition of the veins draining into the lower vertebral column and pelvic girdle from the testes, penis and prostate gland called Batson's plexus [17, 18]. Data indicate that lymphatic channels are only capable of transporting metastatic cells to regional nodes, and further dissemination is carried out through the vascular system [17, 18].
Prior to reaching the circulation for transport to and subsequent re-colonization at a metastatic site, PCa cells must become motile and detach from the primary tumor and overcome the barrier of the extracellular matrix (ECM). Cell locomotion is a coordinated balance between adhesion and detachment of cells through CAMs that occurs simultaneously with tumor cell-induced remodeling of ECM [19–21]. Tumor cell adhesion may be modulated by cytokines and growth factors (GFs) that effect CAM expression and functional organization [22]. To date at least 50 CAMs have been identified and are divided into four major families based on protein structure: 1) the Ig superfamily, 2) the cadherin family, 3) integrins and 4) selectins [23]. Additionally, upregulation of matrix-metallo proteinases (MMPs) and other protease-receptor pairs, like protease-activated receptor 1 (PAR1), facilitate invasion through the basement membrane, providing necessary access to either the lymphatic or vascular circulations [24].
LNCaP and PC-3 (Table 1) models are the two principal culture-to-animal systems being used to delineate the specific mechanisms required for bone metastasis, and will be the focus of experimental data presented in this review, unless otherwise stated. Particularly, the LNCaP model of PCa progression [25–27] gives an opportunity to follow coordinated changes in integrin expression, usage, and behavior of PCa cells when exposed to different ECM substrata and stromally-derived soluble factors; and is unique in that all cell lines vary in metastatic potential but share a common genetic background. Previous phenotypic [27] and genotypic [28] characterizations of these cell lines also revealed their remarkable resemblance to the clinical progression of human prostate cancer. The goals of this review are to discuss the current data that point to tumor- and ECM-derived factors as major contributors of PCa metastasizing to bone, with specific attention to soluble and insoluble factors, CAMs and proteases that mediate PCa cell detachment from the primary tumor, migration and invasion to and through the ECM. At the end of each section, we will attempt to briefly integrate what the data demonstrates in light of key paracrine signaling mechanisms in both the tumor and ECM microenvironments, and explain how we believe these mechanisms may drive metastatic PCa progression. The various prostate cancer cell lines discussed in this review are briefly described in Table 1.
The extracellular matrix and prostate cancer progression
The ECM or basement membrane of most epithelial sheets is primarily composed of laminin and collagen type IV (coll IV), as well as other collagen subtypes. In addition, the ECM also consists of many non-collagenous molecules such as bone sialoprotein (BSP), osteopontin (OPN), osteonectin, osteocalcin (OC), fibronectin, vitronectin (VN), and thrombospondin [29]. As PCa progresses, the expression of many of these components is up-regulated, down-regulated, or lost all-together. These differential patterns of expression aid the tumor in ECM transmigration and ultimately metastasis. For example, when prostate cells are transformed with the neu oncogene, collagen IV is overexpressed in PCa cell lines PC-3 and NbE1.4 [30, 31]; while laminin alone is up-regulated in NbE1.4 [31] and the calcium-independent intercellular cell adhesion molecule-7 (ICAM-1) is down-regulated in PC-3 cells [30]. In the LNCaP progression model, BSP [32, 33], OPN [34] and OC [35] are overexpressed in the more aggressive C4-2 cells. Multiple studies have also demonstrated changes in expression of ECM molecules in advanced PCa tumor samples, such as increased expression of BSP and Cadherin 11 [36, 37], or decreased coll VII expression [38]. Xue et al., found an interesting pattern of expression in Tenascin-C, a molecule involved in stromal-epithelial interactions, where low and moderate-grade tumors showed high levels of the glycoprotein that diminished in high-grade tumors [39]. In the same study, they also documented a dramatic loss of laminin expression indicating a break in the basement membrane adjacent to the tumor cells. These findings were recently corroborated by Brar and colleagues [40].
Cadherins and prostate cancer progression
Cadherins include a multigene family of cell surface adhesion glycoproteins, that provide homotypic interactions between cells and are used to maintain cell:cell associations or mediate cell migration [41]. Classical cadherins (E-, N-, and P-cadherin) possess a highly conserved and characteristic cytoplasmic domain that interacts with their functional partners, β- or γ- and α-catenin [42]. Loss of expression of either pair in this functional complex has been associated with an invasive phenotype due to reduced cell:cell adhesion [43–45]. E-cadherin is an epithelial-specific, calcium dependent CAM that functions to maintain epithelial sheet integrity [46]. As carcinomas become more aggressive with propensity to metastasize, E-cadherin is often lost [46], making it a good candidate to be a metastasis suppressor. It may also serve as a useful prognostic marker for PCa because it is lost with increasing tumor grade and stage [47, 48], and down-regulated in PCa cell lines, PC-3, LNCaP and C4-2 [37, 49, 50]. The nature of the down-regulation is by both protein level and proteolysis resulting in loss of function. Conversely, the expression of N-cadherin in PC-3 cells [49] and advanced prostatic carcinomas [37] has been demonstrated to increase. This would be expected because N-cadherin expression in highly invasive breast cancers was found to replace E-cadherin in facilitating cell:cell contacts, and thought to mediate the interaction between mammary tumor and stromal cells [51]. It is therefore thought to contribute to metastasis via mammary stroma migration [52], and may function similarly in PCa.
In addition to classical cadherins, Bussemakers et al., found other cadherins, including -4, -6, and cadherin-11 were also expressed in a number of PCa cell lines such as PC-3 [37]. In particular, they found an increased expression of cadherin-11 and one of its splice-variants, previously associated with dominant-negative regulation of cell adhesion [53], suggesting it plays a role in PCa progression [37]. Interestingly, they also showed no detectable mRNA or protein expression of cadherin-11 in the less aggressive LNCaP cells.
Role of integrins during progression
The expression of CAMs on cancer cells, as well as on endothelial cells, is not static, but dynamic and strictly controlled by mediators such as GFs, cytokines/chemokines, and the composition of the ECM [22, 54–58]. Cell behavior decisions, such as decreasing cell-cell and cell-substrate attachment, and increasing cell motility are accompanied by changes in the expression and/or usage of adhesion receptors,especially those of the integrin family [59, 60]. Integrins are themselves heterodimeric molecules, consisting of one α and one β subunit, with at least 20 different combinations already described, many of which differ in their extra- and intracellular binding specificities [61, 62]. Integrin molecular structure, heterodimerization, and intra- and extracellular interactions with cytoplasmic regulatory proteins and ECM ligands provide tremendous potential for variation among cell types, well beyond that available through quantitative variation in integrin expression level alone [63].
Since integrins are intimately involved with cell adhesion and motility, experiments were performed to see if there were overt changes in the amount of cell surface integrins expressed in the LNCaP progression model and P69 cell lines. Compared to the cancer cell lines, P69 has much higher expression of integrin subunits (Sikes, unpublished data). These data correspond very well with the strong binding demonstrated in the adhesion assays and the low metastatic potential of this SV40-immortalized cell line (Sikes, unpublished data). Essentially, cells that adhere too well cannot move well. A direct comparison between the cell lines of the LNCaP lineage revealed very few changes in the absolute levels of integrin subunit expression. Only the α2 and β5 integrins increased appreciably. While α2β1 was shown to bind laminin (LN) and collagen [64], our previous work [63] indicates that this integrin pair is probably not utilized in the LNCaP progression model to bind LN. Integrin α2β1 also mediates PC-3 cell adhesion to collagen I, which is a major component of the bone matrix [57]; and was found to mediate cell adhesion to collagen type II, III and IV as well [65].
Despite the modest changes in the absolute levels of integrin subunits, we demonstrated that LNCaP and C4-2 cells have switched the functional pairing of integrin heterodimers [63]. As described by Edlund et al., LNCaP uses primarily α6β4 and not β1 pairs to bind LN, VN and OPN, while C4-2 uses a combination of α3β1 and αvβ3 integrins to bind these matrix components [63]. This switch in integrin heterodimer usage reflects a shift from a junctional integrin expression by LNCaP, α6β4, to a motility associated pair of integrin heterodimers in C4-2 cells, α3β1 and αvβ3. [63]. Additionally, the α6β1 pair is up regulated in C4-2 cells and has been associated with both an increase in metastatic behavior and enhanced cell spreading in many prostate cancer cell lines [31, 63]. In tissue, an increase in α6β1 expression was associated with invasion of the seminal vesicles by prostate cancer [66]. Cooper et al., demonstrated that the β1 subunit is involved in PC-3 cell adhesion to fibronectin, a soluble ECM component of the bone microenvironment [67]. These data would tend to rule out a direct effect of this integrin in the adhesion of either LNCaP or C4-2 cells, but does not rule out a role for this pair in cell motility or invasion, especially considering that the role of β1 in invasion and motility has been aptly demonstrated for endometrial cancer [68]. Direct evidence of this in LNCaP and C4-2 remains to be confirmed experimentally.
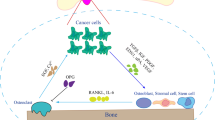
Refocusing on the LNCaP model, the more metastatic sublines were distinct in their use of αvβ3 and, when compared with parental cells, showed a shift in α6 heterodimerization, a subunit critical not only for interaction with prostate basal lamina but also for interaction with the bone matrix [63]. The involvement of the αvβ3 pair is unusual in epithelial cells since it is usually expressed in lymphocytes and other migratory cell types [69, 70]. It is clear, however, that C4-2 cells are using αvβ3 integrin heterodimers for both adhesion and migration on LN, OPN and VN, while LNCaP cells do not [63, 71, 72]. Interestingly, breast and lung cancer cell lines that were derived from bone marrow aspirates also expressed αvβ3, suggesting that it plays a role in overall metastasis of cancer cells to bone marrow [70]. PCa cell adhesion to and migration on components present in the bone matrix are also mediated, in part by αvβ3 [73]. For example, a number of studies have shown that both breast and PCa cells attach to bone-specific ECM components following transendothelial migration, including vitronectin and osteopontin [14, 22, 29, 74, 75]. The αvβ3 expressed on PCa cells, is also a natural receptor for many of the previously mentioned non-collagenous ECM proteins [22, 75–77]; and may give an indication as to why C4-2 cells, known to spontaneously relocate to bone [27], increase their use of this integrin pair as compared to α6β4. It is also relevant to note here that compared to a number of other PCa cell lines, αvβ3 expression was recently determined to be the greatest in PC-3 cells [11]. Thus, the data suggest that αVβ3, in part, facilitates PCa metastasis to bone by mediating PCa cell adhesion to and migration on OPN and VN, two dominant proteins in the bone microenvironment [59].
The mechanism involved in coordinating the heterodimer usage between these cell lines has not yet been determined. Curiously, our FACS analysis of live cells would indicate that the levels of surface α6 and β4 have not changed between LNCaP and C4-2. Even-so, α6β4 use declined in C4-2 cells, in conjunction with an increased use of α6β1, αvβ3, and α3β1 [63]. This was reiterated by the observation that the striking increase in the spreading of C4-2 cells on LN after treatment with stromal factors could be completely obliterated by the addition of function-blocking antibodies against α6 or β1, but not against α2, α3, β4, or αvβ3 [63]. Since α6β4 heterodimers participate in both the formation of hemidesmosomesas well as in the control of cell motility by unique properties of the β4 integrin [78], it is possible that the β4 subunit or α6β4 heterodimer are actively participating in the motility and invasive behavior of PCa cells in a manner that is different from that used by LNCaP cells to attach to a substrate. The function of the β4 integrin would then be determined in a context-dependent manner interpreting environmental cues. In primary prostate carcinomas and established PCa cell lines (DU-145), the α6 integrin subunit maintains a persistent expression during PCa progression, and shifts in β subunit heterodimerization partners were observed from α6β4 alone to also include α6β1 pairs [38, 79]. In other tumor cell types, these laminin-binding integrins (α6β4 and α6β1) have also been linked to acquisition of invasive behaviors [79, 80]. The shift in α6 usage concurs with previous studies, where α6β1 and α6β4 were both found in normal prostate cells, but β4 subunit expression was lost in carcinomas [38, 80, 81]. Taken together, these data indicate that functional changes in surface proteins that are involved in invasion are likely to occur with or without major changes in levels of the protein expressed and that these changes are dependent on the epithelial-ECM-stromal interactions within the tumor and subsequent bone microenvironments (Sikes, unpublished data).
The data also indicate that human prostate cancer has altered integrin expression when disseminated to the bone. When cancer tissue was compared to hyperplastic or benign tissues, the alterations in integrin usage in cancer were found in laminin-binding integrin expression in particular [74, 79, 82, 83]. For example, a number of studies showed decreases in the expression of the α6β4 integrin pair [81], β4 expression alone [84] loss of polarity [85]; as well as decreased α2, α4, and αv expression [86] in more advanced PCa tissues as compared to non-invasive samples. Additionally, Murant et al., found an increase in the β1 subunit as PCa progresses while Zheng et al., documented once more the trend of increased expression of the αvβ3 integrin pair in advanced carcinomas [74, 87].
Multiple pathways resulting from extracellular and intracellular signals regulate invasion of a carcinoma cell. Indeed, cell migration results from the merging of signaling pathways that employ GFs and their receptors, adhesion receptors (integrins) and cytoskeletal elements [88]. In one investigation, Aprikian et al. demonstrated that αVβ3 was involved in bombesin, a neurotransmitter and a cancer growth factor, stimulation of PCa cell motility [89]. Neuroendocrine cells in PCa express and secrete bombesin-like peptides, suggesting that these peptides are involved in PCa progression [89]. Bombesin increased PC-3 cell invasion through matrigel, but did not alter its adhesion to ECM proteins including VN [89]. Additionally, bombesin treatment was found to cause β1, β3, and β5 integrin subunits to coimmunoprecipitate with focal adhesion kinase (FAK) [89]. Functions of αvβ3in PCa cells are mediated by FAK, which activates the phosphatidylinositol 3-kinase (PI-3 kinase)/Akt pathway [74, 76]. The PI3K/Akt pathway may also be involved in androgen-independent growth of PCa [34, 90–94]. Once activated by an upstream kinase such as FAK, this pathway facilitates cell survival and proliferation by increasing expression of the cell cycle regulator E2F, which mediates progression through the cell cycle; as well as prevents the pro-apoptotic activity of BAD [95, 96]. FAK also activates NF-κB, which is known to regulate the transcription of anti-apoptotic proteins [96]. PC-3 cells adhered strongly to collagen type I, a major component of mineralized bone matrix, in the presence of TGF-β (10 ng/ml), a growth factor found in high levels in the bone matrix, and this interaction was mediated by integrins α3β1 and α2β1 [29, 57, 97, 98]. Kiefer and Farach-Carson [29] demonstrated that PC-3 cell adhesion to collagen type I stimulated an increase in cyclin D1 expression followed by an increase in cell division. This implicates the activation of PI3K, map kinase (ERK1/2) and p70S6 kinase in the collagen-mediated effect on PC3 cells.
Changes in extracellular matrix proteases
The ECM is a barrier to a progressing cancer cell at both the primary and metastatic sites. To overcome the ECM barrier, cancer cells alter their production of specific proteases that degrade components of the ECM. Changes in several of these proteases have been associated with prostate cancer progression as described below.
Matrix-metalloproteinases (MMP) are a family of zinc-dependent endopeptidases with broad substrate specificities for a variety of ECM/BM components, such as collagen types I, II, III and IV, laminin and fibronectin [99]. As tumor cells grow and divide, they secrete MMPs that break down the stroma and basement membrane [11]. At the same time, there is down-regulation of tissue inhibitors of MMPs (TIMPs) that amplify the process [100]. In fact, some members of the MMP family of proteases may associate with cell membrane receptors able to drive an oriented degradation of ECM and display a disintegrin region that, by virtue of an RGD motif, play a role in cell-cell adhesion and cell migration (ADAM proteinases: A Disintegrin And Metallo proteinase) [101]. Previous studies have demonstrated the importance of MMPs associated with tumor and stroma as critical determinants for ECM deposition, remodeling and the establishment of PCa metastases in the bone [20, 102, 103]. Changes in expression of specific MMPs are reported to be associated with PCa progression. Lichtinghagen et al., recently demonstrated that MMP-9 protein was significantly higher in cancerous prostate tissue compared to normal prostate tissue [104]. There was no significant difference in MMP-2 expression between cancerous and normal tissues; however, there was a significant difference in the ratios of MMP-2 and MMP-9 to the tissue inhibitor of metalloproteinases 1 (TIMP-1), with cancerous tissue having a higher ratio. Although MMP-2 protein level is not altered during prostate cancer progression, an earlier study with prostate cancer cell lines demonstrated that the expression of membrane-type 1(MT1)-MMP, which activates proMMP-2 and is expressed on the surface of invasive cells, is up-regulated in PC-3 and DU-145 cells [105]. Since the main component of the basement membrane (BM) is collagen type IV, a substrate for both MMP-2 and MMP-9, it is conceivable that the higher expression of MMP-9 and MT1-MMP and the higher ratios of MMP-2 and MMP-9 to TIMP-1 play important roles in the destruction of the BM necessary for invasion and metastasis.
Neutral endopeptidase (NEP)-24.11 (neprilysin) is another cell surface metallopeptidase that may be involved in prostate cancer progression [106]. NEP degrades a variety of bioactive peptides including endothelin, which has been implicated in the growth of hormone refractory prostate cancer. Usmani et al., reported that NEP expression was down-regulated in advanced prostate cancer cell lines PC-3 and DU145 compared to LNCaP and normal immortalized prostate epithelial cells [106]. Also, NEP expression was down-regulated in cells derived from malignant tissues taken from radical prostatectomies compared to those cells derived from benign prostatic hyperplasia (BPH). We speculate that the down-regulation of NEP may contribute to an increase in bioactive peptides required for prostate cancer growth and metastasis
Cathepsins, a family of cysteine proteases capable of degrading several ECM components including coll IV, fibronectin and laminin [107–109]; are up-regulated during PCa progression. One study showed that the expression of Cathepsins (Cath) B and S was higher in prostate cancer tissue compared to BPH and normal prostate tissues; and, they were frequently co-expressed early in the development of prostate cancer [110]. Sinha et al., reported that CathB activity was elevated in prostate cancer tissue samples compared to BPH and normal tissue samples [111]. This study also showed, by biochemical and immunogold electron microscopic analysis, the association of CathB with the plasma membrane as well as in lysosomes [111]. Brubaker et al. recently reported that CathK was expressed in prostate cancer tissues and not in normal prostate tissues [112]. The expression was variable in primary prostate cancer samples and soft-tissue metastases, but was consistently elevated in bone metastases. Surprisingly, CathK expression in PC-3 cell line, which was derived from a bone metastasis, was low compared to DU145 (derived from a brain metastasis) and LNCaP (derived from a lymph node metastasis). Furthermore, more advanced sublines, C4 and C4-2, of the LNCaP progression model, demonstrated decreasing expression of CathK. Note here, the subline C4-2 has a strong tendency to metastasize to bone in murine hosts [113]. Together this information demonstrates that Cathepsins contribute to prostate cancer metastasis and can be up-regulated by the bone microenvironment, a preferred site of metastasis.
The activation of protease-activated receptor 1 (PAR1; thrombin receptor) by thrombin may stimulate prostate cancer cells to secrete MMPs [11, 24]. Chay et al., showed that PAR1 expression was up-regulated in PCa compared with normal prostate tissue. This overexpression was very pronounced in bone-derived PCa cell lines (VCaP and PC3) compared with soft tissue PCa cell lines (DUCaP, DU145, and LNCaP), suggesting the PAR1, like CathK, is up-regulated by stromal factors in the bone marrow [11, 24]. Currently, little is known about the role of PAR1 in prostate cancer progression and studies are underway to determine the effect of PAR1 stimulation on MMP-9 expression in prostate cancer cells. The exact function of PAR1 expression and thrombin in PCa metastasis has also not been delineated; however, these data suggest that PAR1 enhanced expression on bone-derived PCa cells may be important in targeting these cells to the bone [11].
The urokinase-like plasminogen activator receptor (u-PAR) is a membrane-associated serine protease receptor for urokinase or the urokinase plasminogen activator (u-PA). U-PAR is a three-domain molecule. Each domain is numbered 1 to 3 from the amino-terminus to the carboxy-terminus. Domain 1 is the only domain involved in (pro)-u-PA binding, domain 3 also participates in providing the u-PA binding site. The receptor (u-PAR/CD87) focuses the enzymatic activity of u-PA and allows activation of plasminogen (PG) at the cell surface, which in turn, is bound to the cell membrane by α-enolase receptors [114] or to other cell surface proteins endowed with C-terminal lysyl residues, and to plasmin (PL) [101]. PL, a serine proteinase, similar to trypsin, acts almost exclusively when associated with the plasma membrane, because only in that location is it resistant to its inhibitor, α 2-antiplasmin [101]. Plasmin is the main protease involved in (pro)-u-PA activation, which gives origin to the initiation of the classical protease cascade (plasmin, interstitial MMPs, MT1-MMP, Gelatinase A) leading to ECM degradation.
Three extracellular protein ligands involved in ECM degradation and cell adhesion have been identified for u-PAR, namely u-PA, vitronectin (VN), and kininogen. VN and the two-chain form of high molecular weight kininogen (HMWK) share overlapping and mutually exclusive binding sites for u-PAR domains 2 and 3. HMWK-bound kallikrein (kall) may activate the conversion of u-PAR-bound (pro)-u-PA to u-PA, thus providing an alternate pathway to the one triggered by plasmin to (pro)-u-PA activation. Many indications suggest that the u-PA/u-PAR system, together with specific inhibitors of plasminogen activators (PAIs), is an organizer of cell-ECM contacts and covers the full range of activities required to promote and disrupt cell attachment sites [115]. PC-3 and C4-2 secrete more u-PA than LNCaP cells [116, 117], which is most notably involved in the regulation of ECM-laminin degradation, thereby allowing for PC-3 and C4-2 cells to behave more aggressively. Indeed, in a later study conducted by Festuccia et al., the malignant phenotype of PCa cells (LNCaP, C4-2, PC-3 and DU-145) was correlated with both u-PA and u-PAR expression [118]. They found that differential production of u-PA corresponded with the ability of the more aggressive lines to bind and activate plasminogen; thus providing direct support that u-PA secretion and the levels of u-PA- u-PAR complexes characterize the invasive phenotype of these cells [118]. These blocking antibody experiments also provided evidence that this pattern of expression correlates with stage and grade in prostatic carcinomas, making u-PA or plasmin candidate target molecules for metastasis-inhibiting therapeutics.
Several other molecules interact 'constitutively' with u-PAR domains 2 and/or 3, thereby functioning as 'coreceptors'. These molecules include the α2-macroglobulin receptor/low density lipoprotein receptor-related protein (α2MR/LRP), the mannose-6-phosphate receptor (Man6PR)/ IGFII-receptor, gp130, u-PAR-associated protein (u-PARAP) and integrins [101]. Integrin family members including β1, β2 and β3, may interact with u-PAR domains 2 and 3 as u-PAR co-receptors, which leads to an enfeeblement of integrin-ECM interactions [119]. If the cell expresses low or no caveolin, u-PAR-integrin complexes remain loose, floating on unspecialized areas of the cell membrane, integrin function will be impaired, and the cell-ECM interaction will rely solely on adhesive interactions mediated by u-PAR-VN [120]. This seems to be the case for leukocytes and transformed cells, where the motile properties of the cell must prevail on the cell-ECM moorings. On the contrary, if the cells express high levels of caveolin, u-PAR, as with many other GPI-anchored proteins, form clusters on caveolin-rich membrane rafts, together with the loose integrins. In this case, the complex "signalosome" of caveolae, rich in kinases of the src family, transduce signals leading to integrin overexpression, which reinforces cell-ECM interactions. This situation has been recognized in macrophages and metastatic tumor cells and results in enhanced adhesion and migration on ECM components [101]. The end result of the involvement of such a large range of signaling molecules is the activation of several groups of intracellular kinases such as Src, Src-like protein kinases (Hck and Fgf) and again FAK with convergence on the extracellular regulated kinase (ERK)1/2 pathway [121].
Summary and conclusions
The expression of CAMs on tumor cells is not static, but dynamic, and is regulated strictly by extracellular cues like soluble GFs, cytokines, and the insoluble proteins composing the ECM [54–58, 122, 123]. Although a number of integrin variations during PCa cell progression have been described [38, 74, 79–81, 124–126], neither modulation of these variations by external factors nor integrin heterodimer usage regulation is well understood [63]. "Inside-out" regulation of integrin heterodimer activity and subunit partner choices are thought to depend on unique cytoplasmic regulatory protein repertoires that differ among host cell types [127–130]. Likewise, "outside-in" regulation by integrins, in response to extracellular cues, has revealed shifts in integrin gene expression as well as changing integrin associations with numerous signaling molecules,including protein tyrosine kinases (FAK and pp60src), serine kinases (protein kinase C, extracellular signal-regulated kinase, c-Jun-NH2-terminal kinase, and integrin-linked kinase), and lipid intermediates (PI3K and phosphatidylinositol 4,5-kinase) [62, 131–133]. Hence, integrin activity within a given cell is tightly coordinated with its cell cycle, gene expression profiles, differentiation, and cell survival [61]. In our studies, few shifts in integrin expression were found to accompany PCa disease progression, while integrin heterodimer usage, changed significantly [63]. Changes in integrin expression or the functional reassortment of the heterodimers as a tumor progresses has been studied in both the PC-3 and LNCaP PCa cell line models but the mechanism has not been elucidated.
The final component to this story includes the protease-receptor complexes, which are also molecular organizers of cell-to-ECM interactions. They coordinate both adhesive and degradative activities necessary to facilitate metastatic progression [101]. Because traversing the ECM is a critical step in the invasive process, it is imperative that the mechanisms driving the conversion of stationary tumor cells to ones with the capacity to migrate be elucidated. Various cell-associated serine proteases and their respective receptors have been shown to up-regulate u-PAR, giving the u-PA/u-PAR/PAI-1 system prognostic significance in several tumor types [101], including PCa. As information about which u-PAR domains and u-PA sequences specifically mediate malignant invasion via proteolysis and adhesion increases, the likelihood of improved rational drug development to control the factors of the fibrinolytic system should also increase.
Integrin regulation of prostate epithelial proliferation is likely to involve interactions between these CAMs and GF receptors [63]. Such interactions are used by cells to interpret positive and negative GF and cytokine signals from surrounding stromal cells [134, 135], via common signaling cascade components (e.g., small GTPases), also important for integrin signaling and activation. Preferential associations between the GF receptors and the changing integrin heterodimers could have dramatic consequences on the responses of a cell to environmental cues [63]. In LNCaP cells, α6β4, is very important for attachment and would tend to restrict cell migration; while α6β1 and α3β1, both of which are involved in the formation of dynamic focal contacts cycled during migration, are important for cell locomotion [79]. In C4-2 cells, spreading in response to stromal factors appears to be mediated through α6β1 which, along with αvβ3 are responsible for migration [63]. Similarly, we found the α6β1 heterodimer to be more involved in cell spreading than static cell attachment in either neu-transformed cells or the LNCaP progression model [31, 63]. The ability of α3β1 to alter laminin chains and overall basement membrane architecture [136, 137] is particularly suggestive, given that proteolytic cleavage of laminin can drive cells from static adhesion to active migration [138, 139]. Although β1 was shown not to be responsible for PC-3 cell invasion, it may still play a role in C4-2 cell migration/invasion [140]. Most notably, the majority of these functions are directed by α6β4 in LNCaP, suggesting that a functional reassortment of integrins occurs coincident with the acquisition of additional metastatic traits by C4-2 [140]. Not frequently found in epithelial cells, αvβ3 is common to a number of bone metastases, including those of prostate and breast carcinomas [74, 126, 141]. Two possible consequences of αvβ3 heterodimer usage in the metastatic LNCaP sublines are (a) preferential relocation to the bone and (b) increased cell survival/suppressed cell death [63]. Outside of the role of αvβ3 in binding metastatic cells to the bone matrix, this integrin heterodimer is also a good candidate regulator of cell survival in the absence of cell adhesion [63], and thus will remain in the focus of targeted anti-invasive therapies. Although loss of appropriate adhesion is normally a cue for apoptosis, human breast cancer cells are able to use αvβ3 to inhibit p53 activity and suppress the bax death pathway [142]. Accordingly, αvβ3 has been shown to regulate cell proliferation in prostate epithelia [143]. The various patterns of differential expression of ECM molecules, proteases and integrins discussed in this review have been summarized in Table 1.
One of the most notable trends in the area of PCa metastasis is the move towards more complicated in vitro and in vivo systems. This has become necessary since it is now apparent that studying the individual components, prostate or bone cells, or other components of their microenvironments alone, is clearly deficient. These cells alter their gene expression and migratory behavior in response to co-culture or cross-feeding (personal communication Dr. Farach-Carson). Furthermore, the only way to effectively resolve the role of tumor versus ECM factors in the migration and downstream establishment of boney metastases will be to examine the behavior of these various cells types in structured mutlicellular assays. Candidate target proteins can be down regulated using antibodies, ribozymes or RNAi. Conversely critical proteins can be supplemented to systems of reduced complexity to test for the biological response or re-expressed in tumor cells to study the effects of their expression. The individual cells can be followed using fluorescent tags to visualize cell interactions. Once established, these models will allow for the functional analysis of ECM proteins, proteases and integrins involved in the dynamic journey of PCa cells from their site of origin to their interaction with factors and cells from the bone milieu that include ECM, stroma, osteoblasts and osteoclasts, to ultimately colonize the bone.
References
Parkin DM, Pisani P, Ferlay J: Global Cancer Statistics. CA Cancer Journal Clinicians. 1999, 49: 33-64.
Jemal A, Murray T, Samuels A, Ghafoor A, Ward E, Thun MJ: Cancer statistics, 2003. CA Cancer J Clin. 2003, 53: 5-26.
Rubin MA, Putzi M, Mucci N, Smith DC, Wojno K, Korenchuk S, Pienta KJ: Rapid ("warm") autopsy study for procurement of metastatic prostate cancer. Clin Cancer Res. 2000, 6: 1038-1045.
Bubendorf L, Schopfer A, Wagner U, Sauter G, Moch H, Willi N, Gasser TC, Mihatsch MJ: Metastatic patterns of prostate cancer: an autopsy study of 1,589 patients. Hum Pathol. 2000, 31: 578-583.
Jacobs SC: Spread of prostatic cancer to bone. Urology. 1983, 21: 337-344.
Koutsilieris M: Skeletal metastases in advanced prostate cancer: cell biology and therapy. Crit Rev Oncol Hematol. 1995, 18: 51-64.
Rubens RD: Bone metastases-the clinical problem. Eur J Cancer. 1998, 34: 210-213.
Cheville JC, Tindall D, Boelter C, Jenkins R, Lohse CM, Pankratz VS, Sebo TJ, Davis B, Blute ML: Metastatic prostate carcinoma to bone: clinical and pathologic features associated with cancer-specific survival. Cancer. 2002, 95: 1028-1036.
Radinsky R, Fidler IJ: Regulation of tumor cell growth at organ-specific metastases. In Vivo. 1992, 6: 325-331.
Mundy GR: Mechanisms of bone metastasis. Cancer. 1997, 80: 1546-1556.
Cooper CR,, Chay CH, Gendernalik JD, Lee H-L, Bhatia J, Taichman RS, McCauley LK, Keller ET, Pienta KJ: Stromal factors involved in prostate carcinoma metastasis to bone. Cancer. 2003, 97: 739-747.
Gopalkrishnan RV, Kang DC, Fisher PB: Molecular markers and determinants of prostate cancer metastasis. J Cell Physiol. 2001, 189: 245-256.
Ellis LM, Fidler IJ: Angiogenesis and metastasis. Eur J Cancer. 1996, 32A: 2451-2460.
Cooper CR, Pienta KJ: Cell adhesion and chemotaxis in prostate cancer metastasis to bone: a minireview. Prostate Cancer Prostatic Dis. 2000, 3: 6-12.
Lehr Jeffrey E., Pienta Kenneth J.: Preferential adhesion of prostate cancer cells to a human bone marrow endothelial cell line. Journal of the National Cancer Institute. 1998, 90: 118-123.
Chay Christopher H., Cooper Carlton R., Hellerstedt Beth A., Pienta Kenneth J.: Antimetastatic drugs in prostate cancer. Clinical Prostate Can. 2002, 1: 14-19.
Batson OV: The vertebral system of veins as a means for cancer dissemination. Prog Clin Cancer. 1967, 3: 1-18.
Batson OV: The function of the vertebral veins and their role in the spread of metastases. Clin Orthop. 1995, 1940: 4-9.
Holly SP, Larson MK, Parise LV: Multiple roles of integrins in cell motility. Exp Cell Res. 2000, 261: 69-74.
Hornebeck W, Emonard H, Monboisse JC, Bellon G: Matrix-directed regulation of pericellular proteolysis and tumor progression. Semin Cancer Biol. 2002, 12: 231-241.
Condeelis JS, Wyckoff JB, Bailly M, Pestell R, Lawrence D, Backer J, Segall JE: Lamellipodia in invasion. Semin Cancer Biol. 2001, 11: 119-128.
Cooper CR, Bhatia JK, Muenchen HJ, McLean L, Hayasaka S, Taylor J, Poncza PJ, Pienta KJ: The regulation of prostate cancer cell adhesion to human bone marrow endothelial cell monolayers by androgen dihydrotestosterone and cytokines. Clin Exp Metastasis. 2002, 19: 25-33.
Okegawa T, Li Y, Pong RC, Hsieh JT: Cell adhesion proteins as tumor suppressors. J Urol. 2002, 167: 1836-1843.
Chay Christopher H., Cooper Carlton R., Gendernalik James D., Dhanasekaran Saravana M., Chinnaiyan Arul M., Rubin Mark, Schmaier Alvin H., Pienta Kenneth J.: A functional thrombin receptor (PAR1) is expressed on bone-derived prostate cancer cell lines. Urology. 2002, 60: 760-765.
Chung LW, Kao C, Sikes RA, Zhau HE: Human prostate cancer progression models and therapeutic intervention. Hinyokika Kiyo. 1997, 43: 815-820.
Wu HC, Hsieh JT, Gleave ME, Brown NM, Pathak S, Chung LW: Derivation of androgen-independent human LNCaP prostatic cancer cell sublines: role of bone stromal cells. Int J Cancer. 1994, 57: 406-412.
Thalmann George N., Anezinis Ploutarchos E., Chang Shi-Ming, Zhau Haiyen E., Kim E. Edmund, Hopwood Vicki L., Pathak Sen, Eschenbach Andrew C. von, Chung Leland W. K.: Androgen-independent cancer progression and bone metastasis in the LNCaP model of human prostate cancer. Cancer Res. 1994, 54: 2577-2581.
Hyytinen ER, Thalmann GN, Zhau HE, Karhu R, Kallioniemi OP, Chung LW, Visakorpi T: Genetic changes associated with the acquisition of androgen-independent growth, tumorigenicity and metastatic potential in a prostate cancer model. Br J Cancer. 1997, 75: 190-195.
Kiefer JA, Farach-Carson MC: Type I collagen-mediated proliferation of PC3 prostate carcinoma cell line: implications for enhanced growth in the bone microenvironment. Matrix Biol. 2001, 20: 429-437.
Zhau HY, Zhou J, Symmans WF, Chen BQ, Chang SM, Sikes RA, Chung LW: Transfected neu oncogene induces human prostate cancer metastasis. Prostate. 1996, 28: 73-83.
Vafa A, Zhang Y, Sikes RA, Marengo SR: Overexpression of p185erbB2/neu in the NbE prostatic epithelial cell line increases cellular spreading and the expression of integrin alpha6beta1. Int J Oncol. 1998, 13: 1191-1197.
Koeneman KS, Yeung F, Chung LW: Osteomimetic properties of prostate cancer cells: a hypothesis supporting the predilection of prostate cancer metastasis and growth in the bone environment. Prostate. 1999, 39: 246-261.
Lin CC, Lu YP, Lou YR, Ho CT, Newmark HH, MacDonald C, Singletary KW, Huang MT: Inhibition by dietary dibenzoylmethane of mammary gland proliferation, formation of DMBA-DNA adducts in mammary glands, and mammary tumorigenesis in Sencar mice. Cancer Lett. 2001, 168: 125-132.
Thalmann GN, Sikes RA, Devoll RE, Kiefer JA, Markwalder R, Klima I, Farach-Carson CM, Studer UE, Chung LW: Osteopontin: possible role in prostate cancer progression. Clin Cancer Res. 1999, 5: 2271-2277.
Chung LW, Zhau HE, Wu TT: Development of human prostate cancer models for chemoprevention and experimental therapeutics studies. J Cell Biochem Suppl. 1997, 29: 174-181.
Waltregny D, Bellahcene A, Van Riet I, Fisher LW, Young M, Fernandez P, Dewe W, de Leval J, Castronovo V: Prognostic value of bone sialoprotein expression in clinically localized human prostate cancer. J Natl Cancer Inst. 1998, 90: 1000-1008.
Bussemakers MJ, Van Bokhoven A, Tomita K, Jansen CF, Schalken JA: Complex cadherin expression in human prostate cancer cells. Int J Cancer. 2000, 85: 446-450.
Nagle RB, Hao J, Knox JD, Dalkin BL, Clark V, Cress AE: Expression of hemidesmosomal and extracellular matrix proteins by normal and malignant human prostate tissue. Am J Pathol. 1995, 146: 1498-1507.
Xue Y, Li J, Latijnhouwers MA, Smedts F, Umbas R, Aalders TW, Debruyne FM, De La Rosette JJ, Schalken JA: Expression of periglandular tenascin-C and basement membrane laminin in normal prostate, benign prostatic hyperplasia and prostate carcinoma. Br J Urol. 1998, 81: 844-851.
Brar PK, Dalkin BL, Weyer C, Sallam K, Virtanen I, Nagle RB: Laminin alpha-1, alpha-3, and alpha-5 chain expression in human prepubertal [correction of prepubetal] benign prostate glands and adult benign and malignant prostate glands. Prostate. 2003, 55: 65-70.
Takeichi M: Cadherin cell adhesion receptors as a morphogenetic regulator. Science. 1991, 251: 1451-1455.
Kemler R: From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. 1993, 9: 317-321.
Behrens J: Cadherins as determinants of tissue morphology and suppressors of invasion. Acta Anat. 1994, 149: 165-169.
Behrens J: Cell contacts, differentiation, and invasiveness of epithelial cells. Invasion Metastasis. 1994, 14: 61-70.
Birchmeier W, Behrens J: Cadherin expression in carcinomas: role in the formation of cell junctions and the prevention of invasiveness. Biochim Biophys Acta. 1994, 1198: 11-26.
Takeichi M: Cadherins in cancer: implications for invasion and metastasis. Curr Opin Cell Biol. 1993, 5: 806-811.
Umbas R, Schalken JA, Aalders TW, al. et: Expression of the cellular adhesion molecule E-cadherin is reduced or absent in high-grade prostate cancer. Cancer Res. 1992, 52: 5104-5109.
Murant SJ, Rolley N, Phillips SM, Stower M, Maitland NJ: Allelic imbalance within the E-cadherin gene is an infrequent event in prostate carcinogenesis. Genes Chromosomes Cancer. 2000, 27: 104-109.
Tran NL, Nagle RB, Cress AE, Heimark RL: N-Cadherin expression in human prostate carcinoma cell lines. An epithelial-mesenchymal transformation mediating adhesion with Stromal cells. Am J Pathol. 1999, 155: 787-798.
Day ML, Zhao X, Vallorosi CJ, Putzi M, Powell CT, Lin C, Day KC: E-cadherin mediates aggregation-dependent survival of prostate and mammary epithelial cells through the retinoblastoma cell cycle control pathway. J Biol Chem. 1999, 274: 9656-9664.
Hazan RB, Kang L, Whooley BP, Borgen PI: N-cadherin promotes adhesion between invasive breast cancer cells and the stroma. Cell Adhes Commun. 1997, 4: 399-411.
Hagios C, Lochter A, Bissell MJ: Tissue architecture: the ultimate regulator of epithelial function?. Phil Trans R Soc London Series B: Biol Sci. 1998, 353: 857-
Okazaki M, Takeshita S, Kawai S, Kikuno R, Tsujimura A, Kudo A, Amann E: Molecular cloning and characterization of OB-cadherin, a new member of cadherin family expressed in osteoblasts. J Biol Chem. 1994, 269: 12092-12098.
Pauli Bendicht U., Lee Cho-Lea: Organ preference of metastasis: The role of organ-specifically modulated endothelial cells. Laboratory Investigation. 1988, 58: 379-387.
Augustin-Voss HG, Johnson RC, Pauli BU: Modulation of endothelial cell surface glycoconjugate expression by organ-derived biomatrices. Exp Cell Res. 1991, 192: 346-351.
Haraldsen G, Kvale D, Lien B, Farstad IN, Brandtzaeg P: Cytokine-regulated expression of E-selectin, intercellular adhesion molecule-1 (ICAM-1), and vascular cell adhesion molecule-1 (VCAM-1) in human microvascular endothelial cells. Journal of Immunology. 1996, 156: 2558-2565.
Kostenuik PJ, Singh G, Orr FW: Transforming growth factor beta upregulates the integrin-mediated adhesion of human prostatic carcinoma cells to type I collagen. Clin Exp Metastasis. 1997, 15: 41-52.
Khatib AM, Kontogiannea M, Fallavollita L, Jamison B, Meterissian S, Brodt P: Rapid induction of cytokine and E-selectin expression in the liver in response to metastatic tumor cells. Cancer Res. 1999, 59: 1356-1361.
Varner JA, Cheresh DA: Tumor angiogenesis and the role of vascular cell integrin alphav beta3. Important Advances in Oncology. Edited by: VT DeVita, S Hellman and SA Rosenberg. 1996, Philadelphia, Lippincott-Raven Publishers, 66-87.
Albelda SM: Role of integrins and other cell adhesion molecules in tumor progression and metastasis. Lab Invest. 1993, 68: 4-17.
Aplin AE, Howe A, Alahari SK, Juliano RL: Signal transduction and signal modulation by cell adhesion receptors: the role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins. Pharmacol Rev. 1998, 50: 197-263.
Schwartz MA, Schaller MD, Ginsberg MH: Integrins: emerging paradigms of signal transduction. Annu Rev Cell Dev Biol. 1995, 11: 549-599.
Edlund Magnus, Miyamoto Tadayuki, Sikes Robert A., Ogle Roy, Laurie Gordon W., Farach-Carson Mary C., Otey Carol A., Zhau Haiyen E., Chung Leland W. K.: Integrin expression and usage by prostate cancer cell lines on laminin substrata. Cell Growth Differ. 2001, 12: 99-107.
Hemler ME, Elices MJ, Chan BM, Zetter B, Matsuura N, Takada Y: Multiple ligand binding functions for VLA-2 (alpha 2 beta 1) and VLA-3 (alpha 3 beta 1) in the integrin family. Cell Differ Dev. 1990, 32: 229-238.
Rokhlin OW, Cohen MB: Expression of cellular adhesion molecules on human prostate tumor cell lines. Prostate. 1995, 26: 205-212.
Schmelz M, Cress AE, Scott KM, Burger F, Cui H, Sallam K, McDaniel KM, Dalkin BL, Nagle RB: Different phenotypes in human prostate cancer: alpha6 or alpha3 integrin in cell-extracellular adhesion sites. Neoplasia. 2002, 4: 243-254.
Cooper Carlton R., McLean Lisa, Mucci Neil R., Poncza Paul, Pienta Kenneth J.: Prostate cancer cell adhesion to quiescent endothelial cells is not mediated by beta-1 integrin subunit. Anticancer Research. 2000, 20: 4159-4162.
Prifti S, Zourab Y, Koumouridis A, Bohlmann M, Strowitzki T, Rabe T: Role of integrins in invasion of endometrial cancer cell lines. Gynecol Oncol. 2002, 84: 12-20.
Eliceiri BP, Cheresh DA: Role of alpha v integrins during angiogenesis. Cancer J. 2000, 6 Suppl 3: S245-9.
Cooper CR, Chay CH, Pienta KJ: The role of alpha(v)beta(3) in prostate cancer progression. Neoplasia. 2002, 4: 191-194.
Seiffert D: Detection of vitronectin in mineralized bone matrix. J Histochem Cytochem. 1996, 44: 275-280.
Liaw L, Skinner MP, Raines EW, Ross R, Cheresh DA, Schwartz SM, Giachelli CM: The adhesive and migratory effects of osteopontin are mediated via distinct cell surface integrins. Role of alpha v beta 3 in smooth muscle cell migration to osteopontin in vitro. J Clin Invest. 1995, 95: 713-724.
Hullinger TG, McCauley LK, Dejoode ML, Somerman MJ: Effect of bone proteins on human prostate cancer cell lines in vitro. Prostate. 1998, 36: 14-22.
Zheng DQ, Woodard AS, Fornaro M, Tallini G, Languino LR: Prostatic carcinoma cell migration via alpha(v)beta3 integrin is modulated by a focal adhesion kinase pathway. Cancer Res. 1999, 59: 1655-1664.
Byzova Tatiana V., Kim Wes, Midura Ronald J., Plow Edward F.: Activation of integrin avB3 regulates cell adhesion and migration on bone sialoprotein. Exp Cell Res. 2000, 254: 299-308.
Zheng Duo-Qi, Woodard Amy S., Tallini Giovanni, Languino Lucia R.: Substrate specificity of avb3 integrin-mediated cell migration and phosphatidylinositol 3-kinase/AKT pathway activation. Journal of Biological Chemistry. 2000, 275: 24565-24574.
Pecheur I, Peyruchaud O, Serre CM, Guglielmi J, Voland C, Bourre F, Margue C, Cohen-Solal M, Buffet A, Kieffer N, Clezardin P: Integrin alpha(v)beta3 expression confers on tumor cells a greater propensity to metastasize to bone. FASEB. 2002, 16: 1266-1268.
Mercurio AM, Rabinovitz I: Towards a mechanistic understanding of tumor invasion--lessons from the alpha6beta 4 integrin. Semin Cancer Biol. 2001, 11: 129-141.
Rabinovitz I, Nagle RB, Cress AE: Integrin alpha 6 expression in human prostate carcinoma cells is associated with a migratory and invasive phenotype in vitro and in vivo. Clin Exp Metastasis. 1995, 13: 481-491.
Bonkhoff H, Stein U, Remberger K: Differential expression of alpha 6 and alpha 2 very late antigen integrins in the normal, hyperplastic, and neoplastic prostate: simultaneous demonstration of cell surface receptors and their extracellular ligands. Hum Pathol. 1993, 24: 243-248.
Knox JD, Cress AE, Clark V, Manriquez L, Affinito KS, Dalkin BL, Nagle RB: Differential expression of extracellular matrix molecules and the alpha 6-integrins in the normal and neoplastic prostate. Am J Pathol. 1994, 145: 167-174.
Rabinovitz I, Mercurio AM: The integrin alpha6beta4 functions in carcinoma cell migration on laminin-1 by mediating the formation and stabilization of actin- containing motility structures. J Cell Biol. 1997, 139: 1873-1884.
Kitazawa S, Maeda S: Development of skeletal metastases. Clin Orthop. 1995, 45-50.
von Bredow DC, Nagle RB, Bowden GT, Cress AE: Cleavage of beta4 Integrin by Matrilysin. Experimental Cell Res. 1997, 236: 341-345.
Dans M, Gagnoux-Palacios L, Blaikie P, Klein S, Mariotti A, Giancotti FG: Tyrosine phosphorylation of the beta 4 integrin cytoplasmic domain mediates Shc signaling to extracellular signal-regulated kinase and antagonizes formation of hemidesmosomes. J Biol Chem. 2001, 276: 1494-1502.
Cress AE, Rabinovitz I, Zhu W, Nagle RB: The alpha 6 beta 1 and alpha 6 beta 4 integrins in human prostate cancer progression. Cancer Metastasis Rev. 1995, 14: 219-228.
Murant SJ, Handley J, Stower M, Reid N, Cussenot O, Maitland NJ: Co-ordinated changes in expression of cell adhesion molecules in prostate cancer. Eur J Cancer. 1997, 33: 263-271.
Bonaccorsi L, Muratori M, Carloni V, Zecchi S, Formigli L, Forti G, Baldi E: Androgen receptor and prostate cancer invasion. Int J Androl. 2003, 26: 21-25.
Aprikian AG, Tremblay L, Han K, Chevalier S: Bombesin stimulates the motility of human prostate-carcinoma cells through tyrosine phosphorylation of focal adhesion kinase and of integrin-associated proteins. Int J Cancer. 1997, 72: 498-504.
Miyake H, Nelson C, Rennie PS, Gleave ME: Overexpression of insulin-like growth factor binding protein-5 helps accelerate progression to androgen-independence in the human prostate LNCaP tumor model through activation of phosphatidylinositol 3'-kinase pathway. Endocrinology. 2000, 141: 2257-2265.
Lin J, Adam RM, Santiestevan E, Freeman MR: The phosphatidylinositol 3'-kinase pathway is a dominant growth factor- activated cell survival pathway in LNCaP human prostate carcinoma cells. Cancer Res. 1999, 59: 2891-2897.
Murillo H, Huang H, Schmidt LJ, Smith DI, Tindall DJ: Role of PI3K signaling in survival and progression of LNCaP prostate cancer cells to the androgen refractory state. Endocrinology. 2001, 142: 4795-4805.
Yang L, Wang L, Lin HK, Kan PY, Xie S, Tsai MY, Wang PH, Chen YT, Chang C: Interleukin-6 differentially regulates androgen receptor transactivation via PI3K-Akt, STAT3, and MAPK, three distinct signal pathways in prostate cancer cells. Biochem Biophys Res Commun. 2003, 305: 462-469.
Beresford SA, Davies MA, Gallick GE, Donato NJ: Differential effects of phosphatidylinositol-3/Akt-kinase inhibition on apoptotic sensitization to cytokines in LNCaP and PCc-3 prostate cancer cells. J Interferon Cytokine Res. 2001, 21: 313-322.
Downward J: Mechanisms and consequences of activation of protein kinase B/Akt. Curr Opin Cell Biol. 1998, 10: 262-267.
Sonoda Yoshiko, Watanbe Shiro, Matsumoto Yaeko, Aizu-Yokota Eriko, Kasahara Tadashi: FAK is the upstream signal protein of the phosphatidylinositol 3-kinase-Akt survival pathway in hydrogen peroxide-induced apoptosis of a human glioblastoma cell line. Journal of Biological Chemistry. 1999, 274: 10566-10570.
Kostenuik PJ, Sanchez-Sweatman O, Orr FW, Singh G: Bone cell matrix promotes the adhesion of human prostatic carcinoma cells via the alpha 2 beta 1 integrin. Clin Exp Metastasis. 1996, 14: 19-26.
Festuccia C, Bologna M, Gravina GL, Guerra F, Angelucci A, Villanova I, Millimaggi D, Teti A: Osteoblast conditioned media contain TGF-beta1 and modulate the migration of prostate tumor cells and their interactions with extracellular matrix components. Int J Cancer. 1999, 81: 395-403.
Stamenkovic I: Matrix metalloproteinases in tumor invasion and metastasis. Semin Cancer Biol. 2000, 10: 415-433.
Rigg AS, Lemoine NR: Adenoviral delivery of TIMP1 or TIMP2 can modify the invasive behavior of pancreatic cancer and can have a significant antitumor effect in vivo. Cancer Gene Ther. 2001, 8: 869-878.
Del Rosso M, Fibbi G, Pucci M, D'Alessio S, Del Rosso A, Magnelli L, Chiarugi V: Multiple pathways of cell invasion are regulated by multiple families of serine proteases. Clin Exp Metastasis. 2002, 19: 193-207.
Nemeth JA, Harb JF, Barroso U., Jr., He Z, Grignon DJ, Cher ML: Severe combined immunodeficient-hu model of human prostate cancer metastasis to human bone. Cancer Res. 1999, 59: 1987-1993.
Nemeth JA, Yousif R, Herzog M, Che M, Upadhyay J, Shekarriz B, Bhagat S, Mullins C, Fridman R, Cher ML: Matrix metalloproteinase activity, bone matrix turnover, and tumor cell proliferation in prostate cancer bone metastasis. J Natl Cancer Inst. 2002, 94: 17-25.
Lichtinghagen R, Musholt PB, Lein M, Romer A, Rudolph B, Kristiansen G, Hauptmann S, Schnorr D, Loening SA, Jung K: Different mRNA and protein expression of matrix metalloproteinases 2 and 9 and tissue inhibitor of metalloproteinases 1 in benign and malignant prostate tissue. Eur Urol. 2002, 42: 398-406.
Nagakawa O, Murakami K, Yamaura T, Fujiuchi Y, Murata J, Fuse H, Saiki I: Expression of membrane-type 1 matrix metalloproteinase (MT1-MMP) on prostate cancer cell lines. Cancer Lett. 2000, 155: 173-179.
Usmani BA, Shen R, Janeczko M, Papandreou CN, Lee WH, Nelson WG, Nelson JB, Nanus DM: Methylation of the neutral endopeptidase gene promoter in human prostate cancers. Clin Cancer Res. 2000, 6: 1664-1670.
Li Z, Hou WS, Bromme D: Collagenolytic activity of cathepsin K is specifically modulated by cartilage-resident chondroitin sulfates. Biochemistry. 2000, 39: 529-536.
Bonnefoy A, Legrand C: Proteolysis of subendothelial adhesive glycoproteins (fibronectin, thrombospondin, and von Willebrand factor) by plasmin, leukocyte cathepsin G, and elastase. Thromb Res. 2000, 98: 323-332.
Ioachim E, Kamina S, Kontostolis M, Agnantis NJ: Immunohistochemical expression of cathepsin D in correlation with extracellular matrix component, steroid receptor status and proliferative indices in breast cancer. Virchows Arch. 1997, 431: 311-316.
Fernandez PL, Farre X, Nadal A, Fernandez E, Peiro N, Sloane BF, Shi GP, Chapman HA, Campo E, Cardesa A: Expression of cathepsins B and S in the progression of prostate carcinoma. Int J Cancer. 2001, 95: 51-55.
Sinha AA, Quast BJ, Wilson MJ, Fernandes ET, Reddy PK, Ewing SL, Sloane BF, Gleason DF: Ratio of cathepsin B to stefin A identifies heterogeneity within Gleason histologic scores for human prostate cancer. Prostate. 2001, 48: 274-284.
Brubaker KD, Vessella RL, True LD, Thomas R, Corey E: Cathepsin K mRNA and protein expression in prostate cancer progression. J Bone Miner Res. 2003, 18: 222-230.
Thalmann GN, Sikes RA, Wu TT, Degeorges A, Chang SM, Ozen M, Pathak S, Chung LW: LNCaP progression model of human prostate cancer: androgen-independence and osseous metastasis. Prostate. 2000, 44: 91-103 Jul 1;44(2)..
Miles LA, Dahlberg CM, Plescia J, al. et: Role of cell-surface lysines in plasminogen binding to cells: Identification of alpha enolase as a candidate plasminogen receptor.
Andreasen PA, Egelund R, Petersen HH: The plasminogen activation system in tumor growth, invasion, and metastasis. Cell Mol Life Sci. 2000, 57: 25-40.
Hoosein NM, Boyd DD, Hollas WJ, Mazar A, Henkin J, Chung LW: Involvement of urokinase and its receptor in the invasiveness of human prostatic carcinoma cell lines. Cancer Commun. 1991, 3: 255-264.
Xing RH, Rabbani SA: Regulation of urokinase production by androgens in human prostate cancer cells: effect on tumor growth and metastases in vivo. Endocrinology. 1999, 140: 4056-4064.
Festuccia C, Dolo V, Guerra F, Violini S, Muzi P, Pavan A, Bologna M: Plasminogen activator system modulates invasive capacity and proliferation in prostatic tumor cells. Clin Exp Metastasis. 1998, 16: 513-528.
Chapman HA: Plasminogen activators, integrins, and the coordinated regulation of cell adhesion and migration. Curr Opin Cell Biol. 1997, 9: 714-724.
Chapman HA, Wei Y, Simon DI, Waltz DA: Role of urokinase receptor and caveolin in regulation of integrin signaling. Thromb Haemost. 1999, 82: 291-297.
Ossowski L, Aguirre-Ghiso JA: Urokinase receptor and integrin partnership: coordination of signaling for cell adhesion, migration and growth. Curr Opin Cell Biol. 2000, 12: 613-620.
Mainiero F, Pepe A, Yeon M, Ren Y, Giancotti FG: The intracellular functions of alpha6beta4 integrin are regulated by EGF. J Cell Biol. 1996, 134: 241-253.
Ridley AJ, Hall A: The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell. 1992, 70: 389-399.
Nagle RB, Knox JD, Wolf C, Bowden GT, Cress AE: Adhesion molecules, extracellular matrix, and proteases in prostate carcinoma. J Cell Biochem Suppl. 1994, 19: 232-237.
Dedhar S, Saulnier R, Nagle R, Overall CM: Specific alterations in the expression of alpha 3 beta 1 and alpha 6 beta 4 integrins in highly invasive and metastatic variants of human prostate carcinoma cells selected by in vitro invasion through reconstituted basement membrane. Clin Exp Metastasis. 1993, 11: 391-400.
Witkowski Colette M., Rabinovitz Isaac, Nagle Raymmond B., Affinito Kit-Sahn D., Cress Anne E.: Characterization of integrin subunits, cellular adhesion and tumorgenicity of four human prostate cell lines. J Cancer Res Clin Oncol. 1993, 119: 637-644.
Hughes PE, Pfaff M: Integrin affinity modulation. Trends Cell Biol. 1998, 8: 359-364.
Elices MJ, Hemler ME: The human integrin VLA-2 is a collagen receptor on some cells and a collagen/laminin receptor on others. Proc Natl Acad Sci U S A. 1989, 86: 9906-9910.
O'Toole TE, Mandelman D, Forsyth J, Shattil SJ, Plow EF, Ginsberg MH: Modulation of the affinity of integrin alpha IIb beta 3 (GPIIb-IIIa) by the cytoplasmic domain of alpha IIb. Science. 1991, 254: 845-847.
Sims PJ, Ginsberg MH, Plow EF, Shattil SJ: Effect of platelet activation on the conformation of the plasma membrane glycoprotein IIb-IIIa complex. J Biol Chem. 1991, 266: 7345-7352.
Clark EA, Brugge JS: Integrins and signal transduction pathways: the road taken. Science. 1995, 268: 233-239.
Burridge K, Chrzanowska-Wodnicka M: Focal adhesions, contractility, and signaling. Annu Rev Cell Dev Biol. 1996, 12: 463-518.
Schoenwaelder SM, Burridge K: Bidirectional signaling between the cytoskeleton and integrins. Curr Opin Cell Biol. 1999, 11: 274-286.
Chung LW, Davies R: Prostate epithelial differentiation is dictated by its surrounding stroma. Mol Biol Rep. 1996, 23: 13-19.
Giancotti FG, Ruoslahti E: Integrin signaling. Science. 1999, 285: 1028-1032.
DiPersio CM, Hodivala-Dilke KM, Jaenisch R, Kreidberg JA, Hynes RO: alpha3beta1 Integrin is required for normal development of the epidermal basement membrane. J Cell Biol. 1997, 137: 729-742.
Rabinovitz I, Cress AE, Nagle RB: Biosynthesis and secretion of laminin and S-laminin by human prostate carcinoma cell lines. Prostate. 1994, 25: 97-107.
Calof AL, Campanero MR, O'Rear JJ, Yurchenco PD, Lander AD: Domain-specific activation of neuronal migration and neurite outgrowth- promoting activities of laminin. Neuron. 1994, 13: 117-130.
Giannelli G, Falk-Marzillier J, Schiraldi O, Stetler-Stevenson WG, Quaranta V: Induction of cell migration by matrix metalloprotease-2 cleavage of laminin-5. Science. 1997, 277: 225-228.
Cooper Carlton R., Sikes Robert A., Nicholson Brian E., Sun Yan-Xi, Pienta Kenneth J., Taichman Russell S.: Cancer cells homing to bone: the significance of chemotaxis and cell adhesion. The biology of skeletal metastases. Edited by: Evan Keller and Leland WChung. 2003, Kluwer Academic Publishers-in press
Liapis H, Adler LM, Wick MR, Rader JS: Expression of alpha(v)beta3 integrin is less frequent in ovarian epithelial tumors of low malignant potential in contrast to ovarian carcinomas. Hum Pathol. 1997, 28: 443-449.
Stromblad S, Becker JC, Yebra M, Brooks PC, Cheresh DA: Suppression of p53 activity and p21WAF1/CIP1 expression by vascular cell integrin alphaVbeta3 during angiogenesis. J Clin Invest. 1996, 98: 426-433.
Elgavish A, Prince C, Chang PL, Lloyd K, Lindsey R, Reed R: Osteopontin stimulates a subpopulation of quiescent human prostate epithelial cells with high proliferative potential to divide in vitro. Prostate. 1998, 35: 83-94.
Marengo SR, Sikes RA, Anezinis P, Chang SM, Chung LW: Metastasis induced by overexpression of p185neu-T after orthotopic injection into a prostatic epithelial cell line (NbE). Mol Carcinog. 1997, 19: 165-175.
Sikes RA, Chung LW: Acquisition of a tumorigenic phenotype by a rat ventral prostate epithelial cell line expressing a transfected activated neu oncogene. Cancer Res. 1992, 52: 3174-3181.
Chung LW, Chang SM, Bell C, Zhau H, Ro JY, von Eschenbach AC: Prostatic carcinogenesis evoked by cellular interaction. Environ Health Perspect. 1988, 77: 23-28.
Chung LW, Chang SM, Bell C, Zhau HE, Ro JY, von Eschenbach AC: Co-inoculation of tumorigenic rat prostate mesenchymal cells with non-tumorigenic epithelial cells results in the development of carcinosarcoma in syngeneic and athymic animals. Int J Cancer. 1989, 43: 1179-1187.
Bae VL, Jackson-Cook CK, Brothman AR, Maygarden SJ, Ware JL: Tumorigenicity of SV40 T antigen immortalized human prostate epithelial cells: association with decreased epidermal growth factor receptor (EGFR) expression. Int J Cancer. 1994, 58: 721-729.
Bae VL, Jackson-Cook CK, Maygarden SJ, Plymate SR, Chen J, Ware JL: Metastatic sublines of an SV40 large T antigen immortalized human prostate epithelial cell line. Prostate. 1998, 34: 275-282.
Stone KR, Mickey DD, Wunderli H, Mickey GH, Paulson DF: Isolation of a human prostate carcinoma cell line (DU 145). International Journal of Cancer. 1978, 21: 274-281.
Horoszewicz JS, Leong SS, Chu TM, Wajsman ZL, Friedman M, Papsidero L, Kim U, Chai LS, Kakati S, Arya SK, Sandberg AA: The LNCaP cell line-a new model for studies on human prostatic carcinoma. Progress in Clinical & Biological Research. 1980, 37: 115-132.
Korenchuk S, Lehr JE, L M. Clean, Lee YG, Whitney S, Vessella R, Lin DL, Pienta KJ: VCaP, a cell-based model system of human prostate cancer. In Vivo. 2001, 15: 163-168.
Kaighn ME, Narayan KS, Ohnuki Y, Lechner JF, Jones LW: Establishment and characterization of a human prostatic carcinoma cell line (PC-3). Investigative Urology. 1979, 17: 16-23.
Rios-Doria Jonathan, Day Kathleen C., Kuefer Rainer, Rashid Michael G., Chinnaiyan Arul M., Rubin Mark A., Day Mark L.: The Role of Calpain in the Proteolytic Cleavage of E-cadherin in Prostate and Mammary Epithelial Cells. The Journal Of Biological Chemistry. 2003, 278: 1372-1379.
Usmani BA, Harden B, Maitland NJ, Turner AJ: Differential expression of neutral endopeptidase-24.11 (neprilysin) and endothelin-converting enzyme in human prostate cancer cell lines. Clin Sci (Lond). 2002, 103 Suppl 48: 314S-317S..
Conover CA, Perry JE, Tindall DJ: Endogenous cathepsin D-mediated hydrolysis of insulin-like growth factor-binding proteins in cultured human prostatic carcinoma cells. J Clin Endocrinol Metab. 1995, 80: 987-993.
Nunn SE, Peehl DM, Cohen P: Acid-activated insulin-like growth factor binding protein protease activity of cathepsin D in normal and malignant prostatic epithelial cells and seminal plasma. J Cell Physiol. 1997, 171: 196-204.
Manes S, Llorente M, Lacalle RA, Gomez-Mouton C, Kremer L, Mira E, Martinez AC: The matrix metalloproteinase-9 regulates the insulin-like growth factor-triggered autocrine response in DU-145 carcinoma cell. J Biol Chem. 1999, 274: 6935-6945.
Waltregny D, Bellahcene A, de Leval X, Florkin B, Weidle U, Castronovo V: Increased expression of bone sialoprotein in bone metastases compared with visceral metastases in human breast and prostate cancers. J Bone Miner Res. 2000, 15: 834-843.
Ylikoski A, Pettersson K, Nurmi J, Irjala K, Karp M, Lilja H, Lovgren T, Nurmi M: Simultaneous quantification of prostate-specific antigen and human glandular kallikrein 2 mRNA in blood samples from patients with prostate cancer and benign disease. Clin Chem. 2002, 48: 1265-1271.
Kanoh Y, Akahoshi T, Ohara T, Ohtani N, Mashiko T, Ohtani S, Egawa S, Baba S: Expression of matrix metalloproteinase-2 and prostate-specific antigen in localized and metastatic prostate cancer. Anticancer Res. 2002, 22: 1813-1817.
Hashimoto K, Kihira Y, Matuo Y, Usui T: Expression of matrix metalloproteinase-7 and tissue inhibitor of metalloproteinase-1 in human prostate. J Urol. 1998, 160: 1872-1876.
Chambon M, Rebillard X, Rochefort H, Brouillet JP, Baldet P, Guiter J, Maudelonde T: Cathepsin D cytosolic assay and immunohistochemical quantification in human prostate tumors. Prostate. 1994, 24: 320-325.
Uria JA, Lopez-Otin C: Matrilysin-2, a new matrix metalloproteinase expressed in human tumors and showing the minimal domain organization required for secretion, latency, and activity. Cancer Res. 2000, 60: 4745-4751.
Acknowledgments
The authors gratefully acknowledge Dr. Mary C. Farach-Carson for her invaluable discussions on prostate: bone interactions and for critical reading of this manuscript. This work was supported by the prostate cancer research program grant DAMD17-00-1-0049 (RAS), NIH DK/ES59146 (RAS), NIH 1K22CA97117-01 (CRC) and start-up funds from the University of Delaware (RAS/CRC).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Stewart, D.A., Cooper, C.R. & Sikes, R.A. Changes in extracellular matrix (ECM) and ECM-associated proteins in the metastatic progression of prostate cancer. Reprod Biol Endocrinol 2, 2 (2004). https://doi.org/10.1186/1477-7827-2-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1477-7827-2-2