
Abstract
There has been considerable investigation of the potential for soyfoods to reduce risk of cancer, and in particular cancer of the breast. Most interest in this relationship is because soyfoods are essentially a unique dietary source of isoflavones, compounds which bind to estrogen receptors and exhibit weak estrogen-like effects under certain experimental conditions. In recent years the relationship between soyfoods and breast cancer has become controversial because of concerns – based mostly on in vitro and rodent data – that isoflavones may stimulate the growth of existing estrogen-sensitive breast tumors. This controversy carries considerable public health significance because of the increasing popularity of soyfoods and the commercial availability of isoflavone supplements. In this analysis and commentary we attempt to outline current concerns regarding the estrogen-like effects of isoflavones in the breast focusing primarily on the clinical trial data and place these concerns in the context of recent evidence regarding estrogen therapy use in postmenopausal women. Overall, there is little clinical evidence to suggest that isoflavones will increase breast cancer risk in healthy women or worsen the prognosis of breast cancer patients. Although relatively limited research has been conducted, and the clinical trials often involved small numbers of subjects, there is no evidence that isoflavone intake increases breast tissue density in pre- or postmenopausal women or increases breast cell proliferation in postmenopausal women with or without a history of breast cancer. The epidemiologic data are generally consistent with the clinical data, showing no indication of increased risk. Furthermore, these clinical and epidemiologic data are consistent with what appears to be a low overall breast cancer risk associated with pharmacologic unopposed estrogen exposure in postmenopausal women. While more research is required to definitively allay concerns, the existing data should provide some degree of assurance that isoflavone exposure at levels consistent with historical Asian soyfood intake does not result in adverse stimulatory effects on breast tissue.
Similar content being viewed by others


Background
In 1990, participants of a workshop sponsored by the U.S. National Cancer Institute concluded that soybeans contain several putative chemopreventive agents [1]. In the years since, there has been considerable investigation of the potential for soyfoods to reduce risk of cancer, and in particular cancer of the breast. The basis for the initial focus on breast cancer can be attributed to several things: the historically low breast cancer incidence rates in Asia, where soyfoods comprise an important dietary component [2]; research demonstrating the potential for isoflavones – one of the putative chemopreventive agents identified in soybeans – to exert antiestrogenic effects [3]; early epidemiologic data showing an inverse association between soy intake and breast cancer risk [4]; and rodent studies showing a protective effect of soy intake against carcinogen-induced mammary cancer [5].
In recent years, however, the relationship between soyfoods and breast cancer has become controversial because of concerns that soy-derived isoflavones, which exhibit estrogen-like properties under certain experimental conditions, may stimulate the growth of existing estrogen-sensitive breast tumors [6]. These concerns exist because of evidence showing that isoflavones bind and transactivate estrogen receptors (ERs) [7, 8], induce proliferation and estrogenic markers in MCF-7 cells, an ER positive (ER+) breast cancer cell line [9–14], and elicit estrogenic effects in rodent reproductive tissues [15, 16]. In contrast to these findings, epidemiologic evidence shows that among Asian women, higher soy intake is associated with a nearly one-third reduction in breast cancer risk [17] and that Japanese breast cancer patients, in comparison to Western women, exhibit better survival rates even after controlling for stage of diagnosis [18–22].
In 2006, the American Cancer Society concluded that breast cancer patients can safely consume up to three servings of traditional soyfoods per day, although the group advised against the use of more concentrated sources of isoflavones such as powders and supplements [23]. Other expert views are less supportive of the use of any isoflavone-containing products for breast cancer survivors and in some cases for women at high risk of this disease [24–28]. Many women are understandably confused about whether to incorporate soy into their diet. Thus, there is a need for health professionals to have a better understanding of the current evidence relating to soy and breast cancer so that they can better advise their patients and clients. In this analysis and commentary we attempt to outline current concerns regarding estrogen-like effects of isoflavones in the breast and place these concerns in context of recent evidence regarding estrogen therapy (ET) use in postmenopausal women.
Background on isoflavones
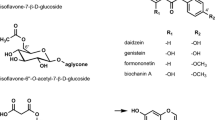
The three soybean isoflavones are genistein, daidzein, and glycitein. These non-steroidal compounds are naturally present in the soybean and non-fermented soyfoods primarily in their beta glycoside forms: genistin, daidzin, and glycitin. Throughout this paper isoflavone amounts refer to the aglycone weight, which is ~60% of the glycoside. In the soybean itself and in most soy products, genistin/genistein, daidzin/daidzein, and glycitin/glycitein account for approximately 50–55%, 40–45%, and 5–10% of total isoflavone content, respectively [29]. Older adults in Japan and Shanghai, China, typically consume between 25 and 50 mg/d isoflavones and probably no more than 5% of these populations consume ≥ 100 mg/d [30]. In contrast, people in the United States and Europe consume on average < 3 mg/d of isoflavones [31–33].
Isoflavones are diphenolic compounds with a chemical structure similar to estrogen that bind to both estrogen receptors alpha (ERα) and beta (ERβ) and, for this reason, are commonly referred to as phytoestrogens [34, 35]. Isoflavones exhibit estrogen-like properties but bind more weakly to ERs than 17β-estradiol (E2), which is the primary physiologic estrogen. Genistein, which is the main circulating and best-studied isoflavone, transactivates ERα and induces estrogenic effects with ~103-104 less potency than E2 [7, 8]. However, serum isoflavone concentrations after a high-soy meal can reach low micromolar levels [36, 37], thereby exceeding postmenopausal total estrogen concentrations by ~103 [38]. This evidence has contributed to the idea that isoflavones may potentially elicit estrogen-like effects and thus serve as a natural alternative to ET in postmenopausal women. Isoflavones also preferentially bind to and transactivate ERβ in comparison to ERα [9, 39, 40] and induce distinct changes in ER conformation [41], leading to speculation that they may function as selective estrogen receptor modulators (SERMs) [42–44]. Despite this designation, unlike different forms of estrogen [45–55], there is scant evidence for any clear estrogen-like or antiestrogenic-like effects of soyfood or isoflavone intake on the human breast or a number of other parameters [44, 55–64].
Effects of isoflavones on mammary/breast cell proliferation
Animal studies
Concern over the possible tumor-stimulatory effects of isoflavones is based largely on the proliferative effect of genistein on MCF-7 cells in vitro and in studies of mammary cancer in rodents. A variety of studies have shown that isoflavones stimulate ER+ human breast cancer cell xenoplants in ovariectomized athymic mice [13, 65–68], estrogen-dependent mammary tumors in rats [69], and reproductive tissues in adult female mice [70, 71]. Other research using rodent models has also demonstrated that genistein is the primary isoflavone responsible for tumor stimulation [72]; that more processed soy products result in faster tumor growth than less processed soy products [68]; and that genistein inhibits the efficacy of tamoxifen, a SERM used in the treatment and prevention of breast cancer [73].
Even in rodent models, however, isoflavones are generally weak estrogen agonists relative to E2. Most rodent studies use scaled doses at least 5 times the amount found in traditional Asian diets [30], and many studies have used direct injection of purified isoflavones, which results in substantially higher levels of unconjugated isoflavones than dietary administration [70]. Importantly, the isoflavone dose required for estrogen-like effects in women has yet to be identified despite three decades of study. So although isoflavones clearly act as estrogens in rodent models, relevant dose effects for human consumption are still very unclear.
There are several noteworthy limitations/weaknesses of the ovariectomized athymic mouse models used in many of the experiments noted above. First, the lack of immune function, which is a necessary element of these models, may eliminate a potential mechanism by which genistein reduces tumor development. Recent research in B6C3F1 mice shows that enhanced immune function resulting from pretreatment with genistein (20 ppm) is correlated with protection against chemically-induced mammary tumors [74]. Second, unlike postmenopausal women, ovariectomized mice do not produce sufficient endogenous estrogen to promote development and growth of estrogen-dependent tumors. Thus, the effects of isoflavones are occurring in an estrogen-depleted environment that does not accurately reflect conditions in either premenopausal or postmenopausal women. It has been argued that estrogenic and tumor-stimulatory effects of isoflavones may be evident only in this type of hypoestrogenic environment. However, this criticism has been addressed by two different models in which isoflavones still lead to tumor stimulation. In one, mice are implanted with MCF-7Ca cells transfected with the enzyme aromatase, enabling the cells to synthesize estrogen; in the other model, mice are continually given small amounts of estrogen [75].
A third critique relates to isoflavone dose. In many studies showing estrogenic effects, mice are exposed to an amount of genistein (750 ppm) that greatly exceeds typical dietary intake. In Japan for example, adults consume about 15–20 mg genistein daily (total mean isoflavone intake is approximately 40 mg), which equates to a dietary concentration of about 30–40 ppm. When expressed on a caloric basis to adjust for differences in metabolism, the difference between human and rodent isoflavone exposure is ~8–16 times higher than the 25 – 50 mg per 1800 Kcal in a traditional Asian diet. (A 30 gm mouse consuming 3 gm of food/d with 750 ppm genistein will consume ~2.25 mg/d of isoflavones, which equates to ~405 mg per 1800 Kcal.) Exposure to purified genistein levels as low as 150 ppm has also been shown to stimulate MCF-7 cell growth, albeit to a lesser extent than higher genistein doses or E2 treatment [67]. Fourth, it is not clear to what extent the existing MCF-7 xenoplants in nude mice reflect tumors in breast cancer patients. These tumors are fully transformed and composed of cells that are extremely sensitive to the growth-stimulating effects of estrogen. Finally, other potentially relevant rodent models [76–78] have shown that isoflavones or isolated soy protein (ISP, by definition is >90% protein) suppress, rather than stimulate, the growth of tumors in mice implanted with MCF-7 cells and even enhance the efficacy of tamoxifen [79, 80].
Clinical studies
Breast tissue is highly regulated by sex hormones, particularly estrogens and progestogens, and breast epithelial proliferation is widely used as an indicator of hormonal exposure or effect. Epithelial cell proliferation also serves as an important prognostic marker in human breast cancer [81] and may help predict risk associated with different hormonal agents [82]. A common method for evaluating proliferation is the immunohistochemical marker Ki67 (also called MIB1), which is a nuclear protein expressed by cells in all active phases of the cycle but not in quiescent or resting cells [83]. Ki67 labeling correlates significantly with higher carcinoma grade, clinical response to endocrine therapy, higher risk of relapse, and worse survival in patients with early breast cancer [84–87].
Four trials, two involving breast cancer patients [88, 89], one in healthy subjects [61], and one in women undergoing breast biopsy or definitive surgery for breast cancer [90] were identified in which breast biopsies were taken before and after exposure to either isoflavone supplements or ISP (Table 1). In no case did the intervention lead to an increase in breast epithelial cell proliferation, which was used in these studies as a marker of potential tumor promotion. Daily isoflavone intake in these trials ranged from 36 [61, 91] to >100 mg [88, 89] and study duration from 2 weeks [90] to one year [89]. In comparison, postmenopausal ET results in modest variable increases in proliferation, while estrogen plus progestin therapy (EPT) results in more significant increases in breast cell proliferation [92, 93].
In one of the trials performed in healthy subjects, 28 postmenopausal women consumed 60 g textured vegetable (soy) protein containing 45 mg isoflavones for 2 weeks. No statistically significant effects on cell proliferation or several other estrogen-responsive markers were found, including progesterone receptor expression, Bcl-expression, and cells undergoing apoptosis and mitosis. However, levels of the estrogen-regulated protein pS2 significantly increased subsequent to soy consumption within breast nipple aspirate (NAF) [69]. The second trial was a 12-week Swedish study in which 51 healthy postmenopausal women took a daily placebo or a supplement that provided 36 mg/d isoflavones [61]. No statistically significant effects of isoflavone treatment were seen on cell proliferation or several other indicators of estrogenic effect (Table 1).
Two other pilot studies involving breast cancer patients also failed to find an effect of isoflavone supplements on breast cell proliferation. The intervention period averaged 23 days in one study [88] and a year in the other [89]. In both studies subjects were exposed to ≥ 100 mg isoflavones per day; however, the one-year study included only 9 women per group and is published only as an abstract. Interestingly, in this study, biopsies taken from the contralateral breast revealed an increase in breast cell proliferation at baseline, which supports the idea that the "healthy" contralateral breast of breast cancer patients may be at an increased risk of developing a tumor [94].
In addition to the lack of effect on cell proliferation, none of the five studies conducted (three in premenopausal [95–98], one in postmenopausal women [97] and one involving both pre- and postmenopausal women [99]) found that isoflavone exposure from soyfoods, ISP, or soybean- or red clover-derived supplements significantly affected breast tissue density (Table 1). Greater breast tissue density is associated with increased breast cancer risk and as was the case for cell proliferation, the lack of effects of isoflavones on breast tissue density generally contrasts with the effects of ET and EPT (see below) [100, 101].
Two additional clinical trials are worthy of comment (Table 1). In one, breast NAF was collected for a total of one year [102]. Samples were taken over three months prior to soy exposure, then for 6 months during which women consumed 37.5 g ISP that provided 75 mg isoflavones daily, and then for 3 months after discontinuation of soy intake [102]. Hyperplastic epithelial cells were noted in 7 of 24 (29.2%) women (4 premenopausal and 3 postmenopausal) while consuming soy whereas prior to soy consumption hyperplastic cells were noted in only 1 of 24 women (4.2%) [102]. The authors concluded that these findings suggest that soybean isoflavones exert an estrogenic stimulus on breast tissue. However, it is important to point out that this was a pilot study with several limitations including the lack of a control group, a high dropout rate (only 15 of 37 subjects finished the 12-month regimen), and the fact that hyperplastic epithelial cells in the NAF persisted far beyond cessation of soy protein intake. Furthermore, a more recent study involving 34 premenopausal women found that isoflavones had no impact on breast cell cytology after one month exposure to either ~24 or 42 mg/d isoflavones [103]. While the available trials examining breast proliferation and density have found no statistically significant effects of isoflavone-containing products it is important to recognize that many of these studies involved small sample sizes or were relative short in duration.
Finally, two epidemiologic studies were identified that examined the relationship between soy or isoflavone intake and breast cancer survival. The first found that soyfood intake was unrelated to survival over the 5.2 year follow-up period [104]. In this study, approximately 63% of the 1001 Chinese breast cancer cases (out of 1459 subjects in the total cohort) for whom data on receptor status was available were ER+. In the other study, when comparing the fifth versus the first intake quintiles, isoflavone intake was associated with a reduced risk of all-cause mortality over the approximate 5-year follow-up period [105]. Isoflavone intake was also associated with a marginal reduction in risk of breast cancer-specific mortality, although the effect was not statistically significant. Of note, the isoflavone intake cutoffs for the fifth quintile were only 7.48 and 0.60 mg/d for all-cause and breast cancer-specific mortality, respectively, and the percentage of ER+ patients among the 1210 subjects was not indicated.
Estrogen and breast cancer risk
Since the estrogen-like effects of isoflavones are at the core of the soy-breast cancer controversy, understanding the relationship between estrogen and breast cancer provides a potentially useful perspective. There is a large amount of evidence that endogenous estrogens are involved in the etiology of certain types of breast cancer [106, 107]. Endogenous estrogens increase breast epithelial proliferation and may facilitate growth of estrogen-sensitive neoplastic or preneoplastic cells [108, 109]. Many of the major epidemiologic risk factors for breast cancer also relate to endogenous estrogen exposure. For example, greater lifelong exposure to ovarian estrogen – as occurs with early menarche and late menopause – is associated with increased breast cancer risk [110–112], whereas oophorectomy reduces risk in premenopausal women [113–115]. In postmenopausal women, higher endogenous circulating concentrations of estrogen [116, 117] are associated with increased risk, as are obesity and alcohol intake, both of which result in higher endogenous estrogen levels [112, 118]. Conversely, treatment with tamoxifen and raloxifene, which inhibits ER activity in the breast, and aromatase inhibitors, which reduce endogenous estrogen production, are effective for treating and preventing ER+ breast cancer [119, 120].
The risk of breast cancer associated with exogenous estrogen exposure is less clear, however, due in part to recent results of the Women's Health Initiative (WHI). This study consisted of two large parallel randomized, double-blind, placebo-controlled clinical trials of hormone therapy designed to evaluate effects of conjugated equine estrogens (CEE) alone (for women with prior hysterectomy) or in combination with the progestin medroxyprogesterone acetate (MPA). In the WHI Estrogen + Progestin Trial, use of CEE + MPA led to a 26% increase in breast cancer risk (38 vs 30 cases per 10,000 person-years) which was highly significant in the weighted analysis (P < 0.001) [121]. However, in the WHI Estrogen-Alone Trial, after an average of 7.1 years of follow-up, women assigned to CEE alone at 0.625 mg/d were 18% less likely to develop invasive breast cancer compared to women in the placebo group (26 vs 33 cases per 10,000 person-years; P = 0.09) [122]. When the latter analysis was restricted to adherent subjects, risk in the CEE group was reduced by one-third (P = 0.03), while the incidence of localized breast carcinoma and ductal carcinoma were lower by 31% and 29%, respectively [123].
The reason for the marginal reduction in breast cancer risk associated with estrogen-alone therapy in the WHI trial is currently unknown. Prior epidemiologic evidence regarding ET effects on breast cancer risk is mixed but generally indicates either no significant effect or a modest increase in risk with long-term exposure [124–128]. Variation within and across observational studies may relate to a variety of factors, including subject selection, screening frequency, duration of hormone use, hormone formulations and doses, and patient characteristics such as reproductive history, body mass index, and background endogenous estrogen context. Nevertheless, overall risks from observational studies are generally small for ET and notably lower than those reported for combined EPT, consistent with WHI results. Importantly, studies of ET use in breast cancer survivors (generally for periods < 5–10 years) also indicate minimal if any risk for recurrence or mortality [129–135].
Direct effects of ET (CEE in particular) on breast proliferation and density are generally modest and less than those seen with EPT. In one of the few clinical studies to assess breast proliferation following ET and EPT, postmenopausal women taking EPT but not ET had significantly greater breast epithelial Ki67 expression in terminal ductal lobular areas [82]. In this study, ET was associated with modestly higher percent breast epithelial area (~15%) compared to the control group (~7%; P = 0.01), while EPT resulted in greater density beyond that seen with ET (~24%; P = 0.02 compared to ET).
Consistent with these findings, the Postmenopausal Estrogen/Progestin Interventions (PEPI) randomized placebo-controlled clinical trial reported a non-significant change in mammographic density of +1.2% after 1 year of CEE treatment compared to significant increases of +3.1 to +4.8% for different EPT regimens [136]. In the WHI, absolute changes in mammographic density were not reported, although CEE resulted in a greater overall percentage of women with abnormal mammograms (36.2% for CEE compared to 28.1% for placebo) [123].
In conclusion, while there is general agreement that endogenous estrogen exposure has an important role in the etiology of breast cancer, the extent to which postmenopausal exogenous estrogen exposure affects risk is much less certain. Current evidence suggests that use of oral ET (particularly CEE) by relatively healthy postmenopausal women for periods < 10 years has very low if any risk for breast cancer and minimal to no effect on breast cancer recurrence or mortality in breast cancer survivors. This information provides a sensible context for considering the potential adverse effects on dietary soyfoods or isoflavones. Given the low overall risk associated with pharmacologic estrogen exposure, how reasonable is it to expect that any weak estrogen-like effects of soy-derived isoflavones (which have yet to be clearly demonstrated in the breast) may increase breast cancer risk or worsen the prognosis of breast cancer patients?
Summary and conclusion
Isoflavones are phytoestrogens which interact with ERs and generally function as weak estrogens in rodent and cell culture models. These estrogen-like effects have raised concern regarding soy/isoflavone consumption, particularly in the case of postmenopausal women at high risk for breast cancer. Currently there is little evidence to suggest that any potential weak estrogenic effects of dietary isoflavones have a clinically relevant impact on breast tissue in healthy women. Limited data suggest this is also the case for breast cancer survivors. This evidence includes multiple trials showing no effects on breast proliferation or mammographic density and considerable epidemiologic data showing either no effect or a modest protective role of soy/isoflavone intake on breast cancer risk. Tangential support for this idea is also provided by recent clinical trial findings regarding exogenous ET (in the form of CEE) showing a marginal decrease in risk of invasive breast cancer. Based on this evidence it seems unlikely that isoflavone consumption at dietary levels (i.e. <100 mg/day) elicits adverse breast cancer-promoting effects in healthy women or breast cancer survivors not undergoing active treatment. Findings from one rodent study showed that genistein may interfere with concurrent tamoxifen treatment, suggesting that breast cancer patients taking a SERM may need to limit soyfood intake and avoid isoflavone supplements. Currently there are no data to support the idea that soyfoods or isoflavone supplements improve the prognosis of breast cancer patients. Available data for ET effects on breast cancer recurrence and mortality provide some assurance for breast cancer patients that soyfoods/isoflavone supplements, when taken at dietary levels, do not contribute to recurrence rates although more data are clearly needed to better address this issue.
Abbreviations
- CEE:
-
conjugated equine estrogens
- E2:
-
17β-estradiol
- ER:
-
estrogen receptor
- ER+:
-
estrogen receptor positive
- ET:
-
estrogen therapy
- EPT:
-
estrogen plus progestin therapy
- ISP:
-
isolated soy protein
- MPA:
-
medroxyprogesterone acetate
- NAF:
-
nipple aspirate fluid
- NSE:
-
no statistically significant effect
- SERM:
-
selective estrogen receptor modulator
- WHI:
-
Women's Health Initiative.
References
Messina M, Barnes S: The role of soy products in reducing risk of cancer. J Natl Cancer Inst. 1991, 83: 541-546. 10.1093/jnci/83.8.541.
Pisani P, Bray F, Parkin DM: Estimates of the world-wide prevalence of cancer for 25 sites in the adult population. Int J Cancer. 2002, 97: 72-81. 10.1002/ijc.1571.
Folman Y, Pope GS: The interaction in the immature mouse of potent oestrogens with coumestrol, genistein and other utero-vaginotrophic compounds of low potency. J Endocrinol. 1966, 34: 215-225.
Lee HP, Gourley L, Duffy SW, Esteve J, Lee J, Day NE: Dietary effects on breast-cancer risk in Singapore. Lancet. 1991, 337: 1197-2000. 10.1016/0140-6736(91)92867-2.
Barnes S, Grubbs C, Setchell KD, Carlson J: Soybeans inhibit mammary tumors in models of breast cancer. Prog Clin Biol Res. 1990, 347:: 239-253.
Messina MJ, Loprinzi CL: Soy for breast cancer survivors: a critical review of the literature. J Nutr. 2001, 131: 3095S-108S.
Muthyala RS, Ju YH, Sheng S, Williams LD, Doerge DR, Katzenellenbogen BS, Helferich WG, Katzenellenbogen JA: Equol, a natural estrogenic metabolite from soy isoflavones: convenient preparation and resolution of R- and S-equols and their differing binding and biological activity through estrogen receptors alpha and beta. Bioorg Med Chem. 2004, 12: 1559-1567. 10.1016/j.bmc.2003.11.035.
Jefferson WN, Newbold RR: Potential endocrine-modulating effects of various phytoestrogens in the diet. Nutrition. 2000, 16: 658-662. 10.1016/S0899-9007(00)00306-3.
An J, Tzagarakis-Foster C, Scharschmidt TC, Lomri N, Leitman DC: Estrogen Receptor beta -Selective Transcriptional Activity and Recruitment of Coregulators by Phytoestrogens. J Biol Chem. 2001, 276: 17808-17814. 10.1074/jbc.M100953200.
Fioravanti L, Cappelletti V, Miodini P, Ronchi E, Brivio M, Di Fronzo G: Genistein in the control of breast cancer cell growth: insights into the mechanism of action in vitro. Cancer Lett. 1998, 130: 143-152. 10.1016/S0304-3835(98)00130-X.
Miodini P, Fioravanti L, Di Fronzo G, Cappelletti V: The two phyto-oestrogens genistein and quercetin exert different effects on oestrogen receptor function. Br J Cancer. 1999, 80: 1150-1155. 10.1038/sj.bjc.6690479.
Le Bail JC, Champavier Y, Chulia AJ, Habrioux G: Effects of phytoestrogens on aromatase, 3beta and 17beta-hydroxysteroid dehydrogenase activities and human breast cancer cells. Life Sci. 2000, 66: 1281-1291. 10.1016/S0024-3205(00)00435-5.
Hsieh CY, Santell RC, Haslam SZ, Helferich WG: Estrogenic effects of genistein on the growth of estrogen receptor- positive human breast cancer (MCF-7) cells in vitro and in vivo. Cancer Res. 1998, 58: 3833-3838.
Zava DT, Duwe G: Estrogenic and antiproliferative properties of genistein and other flavonoids in human breast cancer cells in vitro. Nutr Cancer. 1997, 27: 31-40.
Santell RC, Chang YC, Nair MG, Helferich WG: Dietary genistein exerts estrogenic effects upon the uterus, mammary gland and the hypothalamic/pituitary axis in rats. J Nutr. 1997, 127: 263-269.
Wood CE, Barnes S, Cline JM: Phytoestrogen actions in the breast and uterus. Phytoestrogens and Health. Edited by: Gilani GS and Anderson JJB. 2002, Champaign, IL, AOCS Press, 440-469.
Wu AH, Yu MC, Tseng CC, Pike MC: Epidemiology of soy exposures and breast cancer risk. Br J Cancer. 2008, 98: 9-14. 10.1038/sj.bjc.6604145.
Meng L, Maskarinec G, Wilkens L: Ethnic differences and factors related to breast cancer survival in Hawaii. Int J Epidemiol. 1997, 26: 1151-1158. 10.1093/ije/26.6.1151.
Yonemoto RH: Breast cancer in Japan and United States: epidemiology, hormone receptors, pathology, and survival. Arch Surg. 1980, 115: 1056-1062.
Morrison AS, Lowe CR, MacMahon B, Ravnihar B, Yuasa S: Some international differences in treatment and survival in breast cancer. Int J Cancer. 1976, 18: 269-273. 10.1002/ijc.2910180302.
Ohsumi S, Sakamoto G, Takashima S, Koyama H, Shin E, Suemasu K, Nishi T, Nakamura S, Iino Y, Iwase T, Ikeda T, Teramoto S, Fukutomi T, Komaki K, Sano M, Sugiyama K, Miyoshi K, Kamio T, Ogita M: Long-term results of breast-conserving treatment for early-stage breast cancer in Japanese women from multicenter investigation. Jpn J Clin Oncol. 2003, 33: 61-67. 10.1093/jjco/hyg014.
Kanemori M, Prygrocki M: Results of breast conservation therapy from a single-institution community hospital in Hawaii with a predominantly Japanese population. Int J Radiat Oncol Biol Phys. 2005, 62: 193-197. 10.1016/j.ijrobp.2004.08.054.
Doyle C, Kushi LH, Byers T, Courneya KS, Demark-Wahnefried W, Grant B, McTiernan A, Rock CL, Thompson C, Gansler T, Andrews KS: Nutrition and physical activity during and after cancer treatment: an american cancer society guide for informed choices. CA Cancer J Clin. 2006, 56: 323-353.
Affenito SG, Kerstetter J: Position of the American Dietetic Association and Dietitians of Canada: women's health and nutrition. J Am Diet Assoc. 1999, 99: 738-751. 10.1016/S0002-8223(99)00178-9.
Murkies A, Dalais FS, Briganti EM, Burger HG, Healy DL, Wahlqvist ML, Davis SR: Phytoestrogens and breast cancer in postmenopausal women: a case control study. Menopause. 2000, 7: 289-296.
The role of isoflavones in menopausal health: consensus opinion of The North American Menopause Society. Menopause. 2000, 7: 215-229.
De Lemos M: Safety issues of soy phytoestrogens in breast cancer patients. J Clin Oncol. 2002, 20: 3040-1; discussion 3041-2.
Duffy C, Perez K, Partridge A: Implications of phytoestrogen intake for breast cancer. CA Cancer J Clin. 2007, 57: 260-277.
Murphy PA, Song T, Buseman G, Barua K, Beecher GR, Trainer D, Holden J: Isoflavones in retail and institutional soy foods. J Agric Food Chem. 1999, 47: 2697-2704. 10.1021/jf981144o.
Messina M, Nagata C, Wu AH: Estimated Asian adult soy protein and isoflavone intakes. Nutr Cancer. 2006, 55: 1-12. 10.1207/s15327914nc5501_1.
van Erp-Baart MA, Brants HA, Kiely M, Mulligan A, Turrini A, Sermoneta C, Kilkkinen A, Valsta LM: Isoflavone intake in four different European countries: the VENUS approach. Br J Nutr. 2003, 89 Suppl 1: S25-30.
van der Schouw YT, Kreijkamp-Kaspers S, Peeters PH, Keinan-Boker L, Rimm EB, Grobbee DE: Prospective study on usual dietary phytoestrogen intake and cardiovascular disease risk in Western women. Circulation. 2005, 111: 465-471. 10.1161/01.CIR.0000153814.87631.B0.
Boker LK, Van der Schouw YT, De Kleijn MJ, Jacques PF, Grobbee DE, Peeters PH: Intake of dietary phytoestrogens by Dutch women. J Nutr. 2002, 132: 1319-1328.
Kuiper GG, Carlsson B, Grandien K, Enmark E, Haggblad J, Nilsson S, Gustafsson JA: Comparison of the ligand binding specificity and transcript tissue distribution of estrogen receptors alpha and beta. Endocrinology. 1997, 138: 863-870. 10.1210/en.138.3.863.
Kuiper GG, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT, van der Burg B, Gustafsson JA: Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology. 1998, 139: 4252-4263. 10.1210/en.139.10.4252.
Takashima N, Miyanaga N, Komiya K, More M, Akaza H: Blood isoflavone levels during intake of a controlled hospital diet. J Nutr Sci Vitaminol (Tokyo). 2004, 50: 246-252.
Mathey J, Lamothe V, Coxam V, Potier M, Sauvant P, Pelissero CB: Concentrations of isoflavones in plasma and urine of post-menopausal women chronically ingesting high quantities of soy isoflavones. J Pharm Biomed Anal. 2006, 41: 957-965. 10.1016/j.jpba.2006.01.051.
Pasqualini JR, Chetrite G, Blacker C, Feinstein MC, Delalonde L, Talbi M, Maloche C: Concentrations of estrone, estradiol, and estrone sulfate and evaluation of sulfatase and aromatase activities in pre- and postmenopausal breast cancer patients. J Clin Endocrinol Metab. 1996, 81: 1460-1464. 10.1210/jc.81.4.1460.
Margeat E, Bourdoncle A, Margueron R, Poujol N, Cavailles V, Royer C: Ligands Differentially Modulate the Protein Interactions of the Human Estrogen Receptors alpha and beta. J Mol Biol. 2003, 326: 77-92. 10.1016/S0022-2836(02)01355-4.
Kostelac D, Rechkemmer G, Briviba K: Phytoestrogens modulate binding response of estrogen receptors alpha and beta to the estrogen response element. J Agric Food Chem. 2003, 51: 7632-7635. 10.1021/jf034427b.
Pike AC, Brzozowski AM, Hubbard RE, Bonn T, Thorsell AG, Engstrom O, Ljunggren J, Gustafsson JA, Carlquist M: Structure of the ligand-binding domain of oestrogen receptor beta in the presence of a partial agonist and a full antagonist. Embo J. 1999, 18: 4608-4618. 10.1093/emboj/18.17.4608.
Brzezinski A, Adlercreutz H, Shaoul R, Rösler R, Shmueli A, Tanos V, Schenker JG: Short-term effect of phytoestrogen-rich diet on postmenopausal women. Menopause. 1997, 4: 89-94. 10.1097/00042192-199704020-00005.
Diel P, Geis RB, Caldarelli A, Schmidt S, Leschowsky UL, Voss A, Vollmer G: The differential ability of the phytoestrogen genistein and of estradiol to induce uterine weight and proliferation in the rat is associated with a substance specific modulation of uterine gene expression. Mol Cell Endocrinol. 2004, 221: 21-32. 10.1016/j.mce.2004.04.006.
Yildiz MF, Kumru S, Godekmerdan A, Kutlu S: Effects of raloxifene, hormone therapy, and soy isoflavone on serum high-sensitive C-reactive protein in postmenopausal women. Int J Gynaecol Obstet. 2005, 90: 128-133. 10.1016/j.ijgo.2005.05.005.
Ho JY, Chen MJ, Sheu WH, Yi YC, Tsai AC, Guu HF, Ho ES: Differential effects of oral conjugated equine estrogen and transdermal estrogen on atherosclerotic vascular disease risk markers and endothelial function in healthy postmenopausal women. Hum Reprod. 2006, 21: 2715-2720. 10.1093/humrep/del245.
Lakoski SG, Brosnihan B, Herrington DM: Hormone therapy, C-reactive protein, and progression of atherosclerosis: data from the Estrogen Replacement on Progression of Coronary Artery Atherosclerosis (ERA) trial. Am Heart J. 2005, 150: 907-911. 10.1016/j.ahj.2004.11.025.
Helgason S, Damber JE, Damber MG, von Schoultz B, Selstam G, Sodergard R: A comparative longitudinal study on sex hormone binding globulin capacity during estrogen replacement therapy. Acta Obstet Gynecol Scand. 1982, 61: 97-100.
Serin IS, Ozcelik B, Basbug M, Aygen E, Kula M, Erez R: Long-term effects of continuous oral and transdermal estrogen replacement therapy on sex hormone binding globulin and free testosterone levels. Eur J Obstet Gynecol Reprod Biol. 2001, 99: 222-225. 10.1016/S0301-2115(01)00398-0.
Reid IR, Eastell R, Fogelman I, Adachi JD, Rosen A, Netelenbos C, Watts NB, Seeman E, Ciaccia AV, Draper MW: A comparison of the effects of raloxifene and conjugated equine estrogen on bone and lipids in healthy postmenopausal women. Arch Intern Med. 2004, 164: 871-879. 10.1001/archinte.164.8.871.
Shulman LP: Effects of progestins in different hormone replacement therapy formulations on estrogen-induced lipid changes in postmenopausal women. Am J Cardiol. 2002, 89: 47E-54E; discussion 54E-55E. 10.1016/S0002-9149(02)02413-X.
Marqusee E, Braverman LE, Lawrence JE, Carroll JS, Seely EW: The effect of droloxifene and estrogen on thyroid function in postmenopausal women. J Clin Endocrinol Metab. 2000, 85: 4407-4410. 10.1210/jc.85.11.4407.
Abech DD, Moratelli HB, Leite SC, Oliveira MC: Effects of estrogen replacement therapy on pituitary size, prolactin and thyroid-stimulating hormone concentrations in menopausal women. Gynecol Endocrinol. 2005, 21: 223-226. 10.1080/09513590500279717.
Davies GC, Huster WJ, Shen W, Mitlak B, Plouffe L, Shah A, Cohen FJ: Endometrial response to raloxifene compared with placebo, cyclical hormone replacement therapy, and unopposed estrogen in postmenopausal women. Menopause. 1999, 6: 188-195.
Meuwissen JH, van Langen H: Monitoring endometrial thickness during estrogen replacement therapy with vaginosonography. Radiology. 1992, 183: 284-
Kaari C, Haidar MA, Junior JM, Nunes MG, Quadros LG, Kemp C, Stavale JN, Baracat EC: Randomized clinical trial comparing conjugated equine estrogens and isoflavones in postmenopausal women: a pilot study. Maturitas. 2006, 53: 49-58. 10.1016/j.maturitas.2005.02.009.
D'Anna R, Baviera G, Corrado F, Cancellieri F, Crisafulli A, Squadrito F: The effect of the phytoestrogen genistein and hormone replacement therapy on homocysteine and C-reactive protein level in postmenopausal women. Acta Obstet Gynecol Scand. 2005, 84: 474-477. 10.1111/j.0001-6349.2005.00661.x.
Garrido A, De la Maza MP, Hirsch S, Valladares L: Soy isoflavones affect platelet thromboxane A2 receptor density but not plasma lipids in menopausal women. Maturitas. 2006, 54: 270-276. 10.1016/j.maturitas.2005.12.002.
Khaodhiar L, Ricciotti HA, Li L, Pan W, Schickel M, Zhou J, Blackburn GL: Daidzein-rich isoflavone aglycones are potentially effective in reducing hot flashes in menopausal women. Menopause. 2008, 15: 125-32.
Hall WL, Vafeiadou K, Hallund J, Bugel S, Reimann M, Koebnick C, Zunft HJ, Ferrari M, Branca F, Dadd T, Talbot D, Powell J, Minihane AM, Cassidy A, Nilsson M, Dahlman-Wright K, Gustafsson JA, Williams CM: Soy-isoflavone-enriched foods and markers of lipid and glucose metabolism in postmenopausal women: interactions with genotype and equol production. Am J Clin Nutr. 2006, 83: 592-600.
Katz DL, Evans MA, Njike VY, Hoxley ML, Nawaz H, Comerford BP, Sarrel PM: Raloxifene, soy phytoestrogens and endothelial function in postmenopausal women. Climacteric. 2007, 10: 500-507. 10.1080/13697130701750123.
Cheng G, Wilczek B, Warner M, Gustafsson JA, Landgren BM: Isoflavone treatment for acute menopausal symptoms. Menopause. 2007, 14: 468-473. 10.1097/GME.0b013e31802cc7d0.
Bruce B, Messina M, Spiller GA: Isoflavone supplements do not affect thyroid function in iodine-replete postmenopausal women. J Med Food. 2003, 6: 309-316. 10.1089/109662003772519859.
Marini H, Minutoli L, Polito F, Bitto A, Altavilla D, Atteritano M, Gaudio A, Mazzaferro S, Frisina A, Frisina N, Lubrano C, Bonaiuto M, D'Anna R, Cannata ML, Corrado F, Adamo EB, Wilson S, Squadrito F: Effects of the phytoestrogen genistein on bone metabolism in osteopenic postmenopausal women: a randomized trial. Ann Intern Med. 2007, 146: 839-847.
Sammartino A, Di Carlo C, Mandato VD, Bifulco G, Di Stefano M, Nappi C: Effects of genistein on the endometrium: ultrasonographic evaluation. Gynecol Endocrinol. 2003, 17: 45-49. 10.1080/713603185.
Ju YH, Allred CD, Allred KF, Karko KL, Doerge DR, Helferich WG: Physiological Concentrations of Dietary Genistein Dose-Dependently Stimulate Growth of Estrogen-Dependent Human Breast Cancer (MCF-7) Tumors Implanted in Athymic Nude Mice. J Nutr. 2001, 131: 2957-2962.
Allred CD, Ju YH, Allred KF, Chang J, Helferich WG: Dietary genistin stimulates growth of estrogen-dependent breast cancer tumors similar to that observed with genistein. Carcinogenesis. 2001, 22: 1667-1673. 10.1093/carcin/22.10.1667.
Allred CD, Allred KF, Ju YH, Virant SM, Helferich WG: Soy diets containing varying amounts of genistein stimulate growth of estrogen-dependent (MCF-7) tumors in a dose-dependent manner. Cancer Res. 2001, 61: 5045-5050.
Allred CD, Allred KF, Ju YH, Goeppinger TS, Doerge DR, Helferich WG: Soy processing influences growth of estrogen-dependent breast cancer tumors. Carcinogenesis. 2004, 25: 1649-1657. 10.1093/carcin/bgh178.
Allred CD, Allred KF, Ju YH, Clausen LM, Doerge DR, Schantz SL, Korol DL, Wallig MA, Helferich WG: Dietary genistein results in larger MNU-induced, estrogen-dependent mammary tumors following ovariectomy of Sprague-Dawley rats. Carcinogenesis. 2004, 25: 211-218. 10.1093/carcin/bgg198.
Selvaraj V, Zakroczymski MA, Naaz A, Mukai M, Ju YH, Doerge DR, Katzenellenbogen JA, Helferich WG, Cooke PS: Estrogenicity of the isoflavone metabolite equol on reproductive and non-reproductive organs in mice. Biol Reprod. 2004, 71: 966-972. 10.1095/biolreprod.104.029512.
Cline JM, Franke AA, Register TC, Golden DL, Adams MR: Effects of dietary isoflavone aglycones on the reproductive tract of male and female mice. Toxicol Pathol. 2004, 32: 91-99. 10.1080/01926230490265902.
Ju YH, Fultz J, Allred KF, Doerge DR, Helferich WG: Effects of dietary daidzein and its metabolite, equol, at physiological concentrations on the growth of estrogen-dependent human breast cancer (MCF-7) tumors implanted in ovariectomized athymic mice. Carcinogenesis. 2006, 27: 856-863. 10.1093/carcin/bgi320.
Ju YH, Doerge DR, Allred KF, Allred CD, Helferich WG: Dietary Genistein Negates the Inhibitory Effect of Tamoxifen on Growth of Estrogen-dependent Human Breast Cancer (MCF-7) Cells Implanted in Athymic Mice. Cancer Res. 2002, 62: 2474-2477.
Guo TL, Chi RP, Hernandez DM, Auttachoat W, Zheng JF: Decreased 7,12-dimethylbenz[a]anthracene-induced carcinogenesis coincides with the induction of antitumor immunities in adult female B6C3F1 mice pretreated with genistein. Carcinogenesis. 2007, 28: 2560-2566. 10.1093/carcin/bgm223.
Ju YH, Allred KF, Allred CD, Helferich WG: Genistein stimulates growth of human breast cancer cells in a novel, postmenopausal animal model, with low plasma estradiol concentrations. Carcinogenesis. 2006, 27: 1292-1299. 10.1093/carcin/bgi370.
Zhou JR, Yu L, Zhong Y, Nassr RL, Franke AA, Gaston SM, Blackburn GL: Inhibition of orthotopic growth and metastasis of androgen-sensitive human prostate tumors in mice by bioactive soybean components. Prostate. 2002, 53: 143-153. 10.1002/pros.10141.
Zhou JR, Yu L, Mai Z, Blackburn GL: Combined inhibition of estrogen-dependent human breast carcinoma by soy and tea bioactive components in mice. Int J Cancer. 2004, 108: 8-14. 10.1002/ijc.11549.
Hawrylewicz EJ, Zapata JJ, Blair WH: Soy and experimental cancer: animal studies. J Nutr. 1995, 125: 698S-708S.
Mai Z, Blackburn GL, Zhou JR: Soy Phytochemicals Synergistically Enhance the Preventive Effect of Tamoxifen on the Growth of Estrogen-Dependent Human Breast Carcinoma in Mice. Carcinogenesis. 2007, 28: 1217-1223. 10.1093/carcin/bgm004.
Gotoh T, Yamada K, Ito A, Yin H, Kataoka T, Dohi K: Chemoprevention of N-nitroso-N-methylurea-induced rat mammary cancer by miso and tamoxifen, alone and in combination. Jpn J Cancer Res. 1998, 89: 487-495.
Urruticoechea A, Smith IE, Dowsett M: Proliferation marker Ki-67 in early breast cancer. J Clin Oncol. 2005, 23: 7212-7220. 10.1200/JCO.2005.07.501.
Hofseth LJ, Raafat AM, Osuch JR, Pathak DR, Slomski CA, Haslam SZ: Hormone replacement therapy with estrogen or estrogen plus medroxyprogesterone acetate is associated with increased epithelial proliferation in the normal postmenopausal breast. J Clin Endocrinol Metab. 1999, 84: 4559-4565. 10.1210/jc.84.12.4559.
Scholzen T, Gerdes J: The Ki-67 protein: from the known and the unknown. J Cell Physiol. 2000, 182: 311-322. 10.1002/(SICI)1097-4652(200003)182:3<311::AID-JCP1>3.0.CO;2-9.
de Azambuja E, Cardoso F, de Castro G, Colozza M, Mano MS, Durbecq V, Sotiriou C, Larsimont D, Piccart-Gebhart MJ, Paesmans M: Ki-67 as prognostic marker in early breast cancer: a meta-analysis of published studies involving 12 155 patients. Br J Cancer. 2007, 96: 1504-1513. 10.1038/sj.bjc.6603756.
Weidner N, Moore DH, Vartanian R: Correlation of Ki-67 antigen expression with mitotic figure index and tumor grade in breast carcinomas using the novel "paraffin"-reactive MIB1 antibody. Hum Pathol. 1994, 25: 337-342. 10.1016/0046-8177(94)90140-6.
Chang J, Powles TJ, Allred DC, Ashley SE, Makris A, Gregory RK, Osborne CK, Dowsett M: Prediction of clinical outcome from primary tamoxifen by expression of biologic markers in breast cancer patients. Clin Cancer Res. 2000, 6: 616-621.
Veronese SM, Gambacorta M: Detection of Ki-67 proliferation rate in breast cancer. Correlation with clinical and pathologic features. Am J Clin Pathol. 1991, 95: 30-34.
Sartippour MR, Rao JY, Apple S, Wu D, Henning S, Wang H, Elashoff R, Rubio R, Heber D, Brooks MN: A pilot clinical study of short-term isoflavone supplements in breast cancer patients. Nutr Cancer. 2004, 49: 59-65. 10.1207/s15327914nc4901_8.
Palomares MR, Hopper L, Goldstein L, Lehman CD, Storer BE, Gralow JR: Effect of soy isoflavones on breast proliferation in postmenopausal breast cancer survivors. Breast Cancer Res Treatment. 2004, 88 : 4002-
Hargreaves DF, Potten CS, Harding C, Shaw LE, Morton MS, Roberts SA, Howell A, Bundred NJ: Two-week dietary soy supplementation has an estrogenic effect on normal premenopausal breast. J Clin Endocrinol Metab. 1999, 84: 4017-4024. 10.1210/jc.84.11.4017.
Messina M: The safety and benefits of soybean isoflavones. A natural alternative to conventional hormone therapy?. Menopause. 2007, 14: 958; author reply 958-9. 10.1097/gme.0b013e31812e5258.
Conner P, Skoog L, Soderqvist G: Breast epithelial proliferation in postmenopausal women evaluated through fine-needle-aspiration cytology. Climacteric. 2001, 4: 7-12. 10.1080/713605030.
Conner P, Soderqvist G, Skoog L, Graser T, Walter F, Tani E, Carlstrom K, von Schoultz B: Breast cell proliferation in postmenopausal women during HRT evaluated through fine needle aspiration cytology. Breast Cancer Res Treat. 2003, 78: 159-165. 10.1023/A:1022987618445.
Hemminki K, Ji J, Forsti A: Risks for familial and contralateral breast cancer interact multiplicatively and cause a high risk. Cancer Res. 2007, 67: 868-870. 10.1158/0008-5472.CAN-06-3854.
Maskarinec G, Williams AE, Carlin L: Mammographic densities in a one-year isoflavone intervention. Eur J Cancer Prev. 2003, 12: 165-169. 10.1097/00008469-200304000-00011.
Maskarinec G, Franke AA, Williams AE, Hebshi S, Oshiro C, Murphy S, Stanczyk FZ: Effects of a 2-year randomized soy intervention on sex hormone levels in premenopausal women. Cancer Epidemiol Biomarkers Prev. 2004, 13: 1736-1744.
Atkinson C, Warren RM, Sala E, Dowsett M, Dunning AM, Healey CS, Runswick S, Day NE, Bingham SA: Red-clover-derived isoflavones and mammographic breast density: a double-blind, randomized, placebo-controlled trial. Breast Cancer Res. 2004, 6: R170-9. 10.1186/bcr773.
Messina M, McCaskill-Stevens W, Lampe JW: Addressing the soy and breast cancer relationship: review, commentary, and workshop proceedings. J Natl Cancer Inst. 2006, 98: 1275-1284.
Powles TJ, Howell A, Evans DG, McCloskey EV, Ashley S, Greenhalgh R: Red clover isoflavones are safe and well tolerated in women with a family history of breast cancer. Menopause Int. 2008, 14: 6-12. 10.1258/mi.2007.007033.
Boyd NF, Lockwood GA, Martin LJ, Byng JW, Yaffe MJ, Tritchler DL: Mammographic density as a marker of susceptibility to breast cancer: a hypothesis. IARC Sci Publ. 2001, 154: 163-169.
Boyd NF, Martin LJ, Li Q, Sun L, Chiarelli AM, Hislop G, Yaffe MJ, Minkin S: Mammographic density as a surrogate marker for the effects of hormone therapy on risk of breast cancer. Cancer Epidemiol Biomarkers Prev. 2006, 15: 961-966. 10.1158/1055-9965.EPI-05-0762.
Petrakis NL, Barnes S, King EB, Lowenstein J, Wiencke J, Lee MM, Miike R, Kirk M, Coward L: Stimulatory influence of soy protein isolate on breast secretion in pre- and postmenopausal women. Cancer Epidemiol Biomarkers Prev. 1996, 5: 785-794.
Qin W, Zhu W, Shi H, Hewett JE, Ruhlen RL, MacDonald RS, Rottinghaus GE, Chien YC, Sauter ER: Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Am Inst Cancer Res November 1/2. 2007
Boyapati SM, Shu XO, Ruan ZX, Dai Q, Cai Q, Gao YT, Zheng W: Soyfood intake and breast cancer survival: a followup of the Shanghai Breast Cancer Study. Breast Cancer Res Treat. 2005, 92: 11-17. 10.1007/s10549-004-6019-9.
Fink BN, Steck SE, Wolff MS, Britton JA, Kabat GC, Gaudet MM, Abrahamson PE, Bell P, Schroeder JC, Teitelbaum SL, Neugut AI, Gammon MD: Dietary Flavonoid Intake and Breast Cancer Survival among Women on Long Island. Cancer Epidemiol Biomarkers Prev. 2007, 16: 2285-2292. 10.1158/1055-9965.EPI-07-0245.
Yager JD, Davidson NE: Estrogen carcinogenesis in breast cancer. N Engl J Med. 2006, 354: 270-282. 10.1056/NEJMra050776.
Clemons M, Goss P: Estrogen and the risk of breast cancer. N Engl J Med. 2001, 344: 276-285. 10.1056/NEJM200101253440407.
Yue W, Wang JP, Li Y, Bocchinfuso WP, Korach KS, Devanesan PD, Rogan E, Cavalieri E, Santen RJ: Tamoxifen versus aromatase inhibitors for breast cancer prevention. Clin Cancer Res. 2005, 11: 925s-30s.
Preston-Martin S, Pike MC, Ross RK, Jones PA, Henderson BE: Increased cell division as a cause of human cancer. Cancer Res. 1990, 50: 7415-7421.
Key TJ, Verkasalo PK, Banks E: Epidemiology of breast cancer. Lancet Oncol. 2001, 2: 133-140. 10.1016/S1470-2045(00)00254-0.
Russo J, Russo IH: The role of estrogen in the initiation of breast cancer. J Steroid Biochem Mol Biol. 2006, 102: 89-96. 10.1016/j.jsbmb.2006.09.004.
Chlebowski RT, Anderson GL, Lane DS, Aragaki AK, Rohan T, Yasmeen S, Sarto G, Rosenberg CA, Hubbell FA: Predicting risk of breast cancer in postmenopausal women by hormone receptor status. J Natl Cancer Inst. 2007, 99: 1695-1705. 10.1093/jnci/djm224.
Kauff ND, Barakat RR: Risk-reducing salpingo-oophorectomy in patients with germline mutations in BRCA1 or BRCA2. J Clin Oncol. 2007, 25: 2921-2927. 10.1200/JCO.2007.11.3449.
Kramer JL, Velazquez IA, Chen BE, Rosenberg PS, Struewing JP, Greene MH: Prophylactic oophorectomy reduces breast cancer penetrance during prospective, long-term follow-up of BRCA1 mutation carriers. J Clin Oncol. 2005, 23: 8629-8635. 10.1200/JCO.2005.02.9199.
Hulka BS: Epidemiologic analysis of breast and gynecologic cancers. Prog Clin Biol Res. 1997, 396: 17-29.
Hankinson SE, Eliassen AH: Endogenous estrogen, testosterone and progesterone levels in relation to breast cancer risk. J Steroid Biochem Mol Biol. 2007, 106: 24-30. 10.1016/j.jsbmb.2007.05.012.
Key T, Appleby P, Barnes I, Reeves G: Endogenous sex hormones and breast cancer in postmenopausal women: reanalysis of nine prospective studies. J Natl Cancer Inst. 2002, 94: 606-616.
Kendall A, Folkerd EJ, Dowsett M: Influences on circulating oestrogens in postmenopausal women: relationship with breast cancer. J Steroid Biochem Mol Biol. 2007, 103: 99-109. 10.1016/j.jsbmb.2006.07.011.
Vogel VG, Costantino JP, Wickerham DL, Cronin WM, Cecchini RS, Atkins JN, Bevers TB, Fehrenbacher L, Pajon ER, Wade JL, Robidoux A, Margolese RG, James J, Lippman SM, Runowicz CD, Ganz PA, Reis SE, McCaskill-Stevens W, Ford LG, Jordan VC, Wolmark N: Effects of tamoxifen vs raloxifene on the risk of developing invasive breast cancer and other disease outcomes: the NSABP Study of Tamoxifen and Raloxifene (STAR) P-2 trial. Jama. 2006, 295: 2727-2741. 10.1001/jama.295.23.joc60074.
Thurlimann B, Keshaviah A, Coates AS, Mouridsen H, Mauriac L, Forbes JF, Paridaens R, Castiglione-Gertsch M, Gelber RD, Rabaglio M, Smith I, Wardley A, Price KN, Goldhirsch A: A comparison of letrozole and tamoxifen in postmenopausal women with early breast cancer. N Engl J Med. 2005, 353: 2747-2757. 10.1056/NEJMoa052258.
Writing Group for the Women's Health Initiative Investigators: Risks and benefits of estrogen plus progestin in healthy postmenopausal women: principal results From the Women's Health Initiative randomized controlled trial. JAMA. 2002, 288: 321-333. 10.1001/jama.288.3.321.
Anderson GL, Limacher M, Assaf AR, Bassford T, Beresford SA, Black H, Bonds D, Brunner R, Brzyski R, Caan B, Chlebowski R, Curb D, Gass M, Hays J, Heiss G, Hendrix S, Howard BV, Hsia J, Hubbell A, Jackson R, Johnson KC, Judd H, Kotchen JM, Kuller L, LaCroix AZ, Lane D, Langer RD, Lasser N, Lewis CE, Manson J, Margolis K, Ockene J, O'Sullivan MJ, Phillips L, Prentice RL, Ritenbaugh C, Robbins J, Rossouw JE, Sarto G, Stefanick ML, Van Horn L, Wactawski-Wende J, Wallace R, Wassertheil-Smoller S: Effects of conjugated equine estrogen in postmenopausal women with hysterectomy: the Women's Health Initiative randomized controlled trial. Jama. 2004, 291: 1701-1712. 10.1001/jama.291.14.1701.
Stefanick ML, Anderson GL, Margolis KL, Hendrix SL, Rodabough RJ, Paskett ED, Lane DS, Hubbell FA, Assaf AR, Sarto GE, Schenken RS, Yasmeen S, Lessin L, Chlebowski RT: Effects of conjugated equine estrogens on breast cancer and mammography screening in postmenopausal women with hysterectomy. Jama. 2006, 295: 1647-1657. 10.1001/jama.295.14.1647.
Warren MP: A comparative review of the risks and benefits of hormone replacement therapy regimens. Am J Obstet Gynecol. 2004, 190: 1141-1167. 10.1016/j.ajog.2003.09.033.
Ross RK, Paganini-Hill A, Wan PC, Pike MC: Effect of hormone replacement therapy on breast cancer risk: estrogen versus estrogen plus progestin. J Natl Cancer Inst. 2000, 92: 328-332. 10.1093/jnci/92.4.328.
Schairer C, Lubin J, Troisi R, Sturgeon S, Brinton L, Hoover R: Estrogen-progestin replacement and risk of breast cancer. Jama. 2000, 284: 691-694. 10.1001/jama.284.6.691.
Li CI, Malone KE, Porter PL, Weiss NS, Tang MT, Cushing-Haugen KL, Daling JR: Relationship between long durations and different regimens of hormone therapy and risk of breast cancer. Jama. 2003, 289: 3254-3263. 10.1001/jama.289.24.3254.
Chen WY, Manson JE, Hankinson SE, Rosner B, Holmes MD, Willett WC, Colditz GA: Unopposed estrogen therapy and the risk of invasive breast cancer. Arch Intern Med. 2006, 166: 1027-1032. 10.1001/archinte.166.9.1027.
Vassilopoulou-Sellin R, Asmar L, Hortobagyi GN, Klein MJ, McNeese M, Singletary SE, Theriault RL: Estrogen replacement therapy after localized breast cancer: clinical outcome of 319 women followed prospectively. J Clin Oncol. 1999, 17: 1482-1487.
Vassilopoulou-Sellin R, Cohen DS, Hortobagyi GN, Klein MJ, McNeese M, Singletary SE, Smith TL, Theriault RL: Estrogen replacement therapy for menopausal women with a history of breast carcinoma: results of a 5-year, prospective study. Cancer. 2002, 95: 1817-1826. 10.1002/cncr.10913.
O'Meara ES, Rossing MA, Daling JR, Elmore JG, Barlow WE, Weiss NS: Hormone replacement therapy after a diagnosis of breast cancer in relation to recurrence and mortality. J Natl Cancer Inst. 2001, 93: 754-761. 10.1093/jnci/93.10.754.
Peters GN, Fodera T, Sabol J, Jones S, Euhus D: Estrogen replacement therapy after breast cancer: a 12-year follow-up. Ann Surg Oncol. 2001, 8: 828-832. 10.1007/s10434-001-0828-4.
Col NF, Hirota LK, Orr RK, Erban JK, Wong JB, Lau J: Hormone replacement therapy after breast cancer: a systematic review and quantitative assessment of risk. J Clin Oncol. 2001, 19: 2357-2363.
Creasman WT: Hormone replacement therapy after cancers. Curr Opin Oncol. 2005, 17: 493-499.
DiSaia PJ, Brewster WR, Ziogas A, Anton-Culver H: Breast cancer survival and hormone replacement therapy: a cohort analysis. Am J Clin Oncol. 2000, 23: 541-545. 10.1097/00000421-200012000-00001.
Greendale GA, Reboussin BA, Slone S, Wasilauskas C, Pike MC, Ursin G: Postmenopausal hormone therapy and change in mammographic density. J Natl Cancer Inst. 2003, 95: 30-37.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
M.M. is president of Nutrition Matters, Inc., a nutrition consulting company with clients involved in the manufacture and/or sale of soyfoods and isoflavone supplements.
Authors' contributions
MM and CEW were equally involved in the writing of this manuscript.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Messina, M.J., Wood, C.E. Soy isoflavones, estrogen therapy, and breast cancer risk: analysis and commentary. Nutr J 7, 17 (2008). https://doi.org/10.1186/1475-2891-7-17
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2891-7-17