
Abstract
Background
The erythrocyte binding antigen-175 (EBA-175) on Plasmodium falciparum merozoites mediates sialic acid dependent binding to glycophorin A on host erythrocytes and, therefore, plays a crucial role in cell invasion. Dimorphic allele segments have been found in its encoding gene with a 342 bp segment present in FCR-3 strains (F-segment) and a 423 bp segment in CAMP strains (C-segment). Possible associations of the dimorphism with severe malaria have been analysed in a case-control study in northern Ghana.
Methods
Blood samples of 289 children with severe malaria and 289 matched parasitaemic but asymptomatic controls were screened for eba- 175 F- and C-segments by nested polymerase chain reaction.
Results
In children with severe malaria, prevalences of F-, C- and mixed F-/C-segments were 70%, 19%, and 11%, respectively. The C-segment was found more frequently in severe malaria cases whereas mixed infections were more common in controls. Infection with strains harbouring the C-segment significantly increased the risk of fatal outcome.
Conclusion
The results show that the C-segment is associated with fatal outcome in children with severe malaria in northern Ghana, suggesting that it may contribute to the virulence of the parasite.
Similar content being viewed by others


Introduction
Malaria parasites invade host erythrocytes via interaction between merozoite surface ligands and erythrocyte receptors [1, 2]. In Plasmodium falciparum, the erythrocyte binding antigen-175 (EBA-175) is one of the most important ligands mediating erythrocyte invasion by sialic acid dependent binding to glycophorin A [3, 4]. The 175 kDa protein is located in micronemes at the apical end of the merozoite [5]. Together with the Duffy-binding protein of Plasmodium vivax (PvDBP), EBA-175 belongs to a family of parasite adhesion molecules, the Duffy-binding-like erythrocyte-binding proteins (DBL-EBP) [6]. This protein family is characterized by a N-terminal signal peptide, cystein-rich Duffy-binding-like (DBL) domains, a carboxyl cysteine-rich region (c-cys), a transmembrane domain and a short cytoplasmatic tail. Encoding genes are members of the erythrocyte-binding-like (ebl) gene family. The eba-175 gene is located on chromosome 7 and consists of four exons [7, 8]. A large exon 1 is divided into regions I-VI and codes for a tandem array of two DBL domains in region II (Fl and F2) forming the putative ligand site as well as the c-cys domain, region VI. Both domains have numerous conserved cystein and hydrophobic amino acid residues supporting their functional relevance. They are separated by highly divergent dimorphic segments in region III [9]. This dimorphism is characterized by an insertion of either a 342 bp segment in FCR-3 strains (F-segment) or a 423 bp segment in CAMP strains (C-segment) [7, 10]. Both segments share little homology and are located at slightly different positions with the F-segment being 273 bp upstream of the C-segment. These two variants are conserved in all strains isolated so far and since Plasmodium merozoites are haploid either one or the other, but never both or neither segment is present in one individual parasite clone [10]. The functional relevance of the F-/C-segment dimorphism remains unknown. Human IgG antibodies recognize epitopes on the dimorphic F- and C-segments but are not associated with protection from clinical malaria in The Gambia [11]. Population genetic analysis showed a C-segment allele frequency of 73% in eba-175 of P. falciparum in a Sudanese population whereas more than 70% of isolates from The Gambia, Nigeria, Cameroon, Gabon, Tanzania and South Africa carried the F-segment [12, 13].
The two variants of the eba-175 dimorphism are evolutionarily conserved suggesting functional relevance. F- and C-segment frequencies vary in different ethnicities and geographic regions. It is, therefore, hypothesized that this dimorphism may influence clinical disease and outcome.
Methods
Study area and parasites
Samples were collected between August and November 2002 in Tamale and its vicinity, Northern Region, Ghana. Climate and vegetation are savannah-type. Malaria transmission is perennial but highly seasonal and infection rates are consistent with a hyperendemic malaria situtation (Mockenhaupt et al., unpublished data). Blood samples and clinical data were obtained from 290 children (aged 6 months to 9 years) with severe falciparum malaria (SM) according to the current WHO definition [14]. One sample was lost for genetic analysis. The clinical and parasitological characteristics of these children are described in detail elsewhere [15]. For each patient with SM, one age- and sex-matched asymptomatic but parasitaemic control was randomly selected from a survey representative for Tamale and the surrounding six districts. For that purpose, 30 communities and census units were recruited following a two-stage cluster sampling strategy with probability proportional to population size. Within each unit, ≥70 children aged 6 months to 9 years were randomly selected, clinical examinations were performed and venous blood samples were taken. Parasites were counted per ≥200 white blood cells on Giemsa-stained thick blood films and P. falciparum was substantiated by specific PCR assays [16].
DNA extraction and genotyping
Blood was stabilized (AS1, Qiagen, Hilden, Germany), stored at 4°C and DNA was extracted by commercial kits (QIAmp blood kit, Qiagen, Hilden, Germany). Genotyping of the eba dimorphism was performed by nested PCR using primers EBA1 5'-CAAGAAGCAGTTCCTGAGGAA-3' (forward) and EBA2 5'-TCTCAACATTCATATTAACAATTC-3' (reverse) for the first amplification and EBA3 5'-GAGGAAAACACTGAAATAGCACAC-3' (forward) and EBA4 5'-CAATTCCTCC-AGACTGTTGAACAT-3' (reverse) for the second amplification [17]. The following mixture was used for 20 μl reaction volume [18]: 4 μl (50–200 ng) of DNA-template for the first amplification and 2 μl PCR-product for the nested PCR; 1 μl of each primer (l00 μM), 1 U HotStarTaq Polymerase (Qiagen®, USA); 1x PCR-Buffer, 1,5 mM magnesium chloride; and 200 μM dNTP. PCR was performed on T3 Thermocylcer (Biometra®, Germany) using the following conditions: Initial 5-min denaturation at 94°C followed by 29 cycles with 1-min at 94°C, 1-min at 56°C and 2-min 72°C and a 3-min final extension at 72°C. Amplicons were separated on 1.5% ethidiumbromide stained agarose gels and visualized using UV-translumination. A single band of 795 or 714 bp identified infection with parasites containing the F- or C-segment, respectively. Samples showing both bands reflected mixed infection with a minimum of two different clones.
Statistical analysis
Geometric mean parasite density (GMPD) and 95% confidence intervals (95% CI) were calculated after log10 transformation. Genotype frequencies were compared between case and control groups using Pearson Chi-Square test and Fisher's exact test as applicable. Subgroup analysis of the risk of individual symptoms was performed by conditional logistic regression models. Associations between genotypes and fatal outcome in severe malaria cases were corrected for known predictors of mortality (impaired consciousness, circulatory collapse, hypoglycaemia and malnutrition [15]) by multiple logistic regression analysis. A P-value < .05 was considered as statistically significant.
Results
The median age of the 289 children with severe malaria (154 girls, 135 boys) was 24 months (range 6–102). Mean rectal temperature (±SD) was 38.6°C (±1.1). GMPD was 30,200/μL (95% CI, 22,474–40,581) and hyperparasitaemia (>250,000/μL) was present in 64 children (22%). The GMPD of parasitaemic controls was 1,738/μL (95% CI, 1,387–2,178). Cases exhibited the following symptoms defining severe malaria (WHO 2000): severe anemia (55%), prostration (34%), respiratory distress (23%), multiple convulsions (20%), impaired consciousness (19%), jaundice (12%), circulatory collapse (3%), and haemoglobinuria (3%). Case fatality rate was 11% (32/284). For five children, no data on outcome were available since they absconded during hospitalization.
In children with severe malaria, the F- and C-segment were present in 70% (201/289) and 19% (55/289), respectively (see Table 1). Both segments were found in 11% (33/289) representing mixed infections. In the parasitaemic controls, the prevalences for the F- and C-segment were 61% (176/289) and 17% (49/289), respectively. Mixed infections were significantly less frequent in severe cases than in parasitaemic controls (11% (33/289) versus 22% (64/289); OR .5, [95% CI .3–.7], P = .003; F-segment used as reference). Matched pair analysis revealed no significant associations of the C- or F-segment with individual symptoms and conditions of SM (Table 1). However, for all symptoms and conditions of SM except for haemoglobinuria, the C-segment was more common in severe cases while mixed infections occured more often in respective parasitaemic controls (not significant).
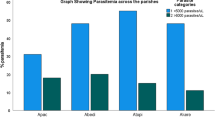
A trend towards a higher risk of dying of severe malaria was found in children with the C-segment in matched pair analysis (OR 4.0, [95% CI .9–19.4], P = .1; Table 1). Associations of the eba-175 dimorphism with symptoms, conditions, and outcome within the group of severe malaria cases were re-tested (Figure 1). The C-segment was significantly associated with fatal outcome (case fatality rates: F-segment: 8%, F-/C-segment: 13%, C-segment: 24%; _2trend = 11.3, P = .0008). Logistic regression analysis adjusting for known predictors of death in these children [15] confirmed a six-fold increased risk of fatal outcome in patients infected with C-segment parasites as compared to F-segment parasites (OR 5.6, [95% CI 1.8–17.1], P < .003; other predictors were: impaired consciousness, OR, 9.3; 95% CI, 3.4–25.9; P < .0001; circulatory collapse, OR, 35.9; 95% CI, 4.5–290; P = .0008; hypoglycemia, OR, 3.7; 95% CI, 1.3–10.3; P = .01; malnutrition, OR, 95% CI, 2.5; 1.0–6.3; P = .06).

eba-175 dimorphism (F-segment, F-/C-mixed infection, C-segment): Percentage (absolute numbers) of severe malaria cases with respective symptoms, conditions, and outcome.
Discussion
This study aimed at evaluating the frequencies of a dimorphic allele segment in the gene encoding the erythrocyte binding antigen-175 on P. falciparum merozoites in children with severe malaria in northern Ghana. Second, associations of the eba-175 dimorphism with symptoms, conditions, and outcome of severe disease were assessed in a case-control analysis.
In 289 Ghanaian children with severe malaria, the F-segment predominated (70%). C-segment and mixed infections were present in 19% and 11%, respectively. These findings are consistent with recently published data from West Africa [13].
In the case-control analysis, mixed infections were significantly more common in controls than in cases. This is in accordance with findings from other areas with high transmission [19, 20] and is considered to indicate acquired immunity [21]. However, the presence of either the F- or the C-segment alone does not exclude the possibility of infection with different strains and, therefore, limits validity of this finding. The overall distribution of the F- and C-alleles did not differ significantly between groups arguing against a role for the eba dimorphism in mediating severe malaria per se. This is consistent with the finding in the Gambia that antibodies against the dimorphism do not protect against clinical malaria [11]. Yet, within the group of children with severe malaria, fatal outcome was significantly associated with the C-segment (Figure 1). These findings suggest that the C-segment of the eba-175 dimorphism on P. falciparum merozoites is associated with fatality in severe malaria rather than promoting disease progression from asymptomatic parasitaemia towards severe disease. A longitudinal study is needed to confirm this.
Unlike in West Africa, in a Sudanese population the C-segment was most frequent [13]. This part of Africa represents an ethnically distinct region with lower malaria endemicity. In a study conducted in Laos, the dimorphism was unequally distributed in distinct ethnic minorities [18]. Random shifts in parasite allele frequencies in different geographic regions can serve as an explanation. Also, susceptibility to infection and disease severity vary in different regions as does genetic variability in host populations. On the other hand, not only genetic differences in the host but also in the parasite may influence disease severity and outcome. Although the functional mechanism of the eba-175 dimorphism regarding disease severity remains unclear one might postulate that this dimorphism offers alternative ways of erythrocyte invasion to the parasite and therefore allows more rapid replication. Since the eba-175 dimorphism appears not to be involved in the binding to neuraminidase- or trypsin-sensitive erythrocyte receptors [22] it may recognize other, as yet unknown, receptors. This explanation is supported by the finding that the C-segment was associated at borderline statistical significance with hyperparasitaemia (OR 3.7 [1.1–13.1] P = .06. see Table 1).
In conclusion, the C-segment of the eba-175 allelic dimorphism on P. falciparum merozites is not associated with severe malaria per se but confers a higher risk of fatal disease in northern Ghana. These findings may contribute to the development of a diagnostic tool predicting disease outcome in children. Further studies are necessary to reveal its functional relevance.
References
Bannister LH, Dluzewski AR: The ultrastructure of red cell invasion in malaria infections: a review. Blood Cells. 1990, 16: 257-292.
Chitnis CE: Molecular insights into receptors used by malaria parasites for erythrocyte invasion. Curr Opin Hematol. 2001, 8: 85-91. 10.1097/00062752-200103000-00005.
Camus D, Hadley TJ: A Plasmodium falciparum antigen that binds to host erythrocytes and merozoites. Science. 1985, 230: 553-556.
Orlandi PA, Klotz FW, Haynes JD: A malaria invasion receptor, the 175-kilodalton erythrocyte binding antigen of Plasmodium falciparum recognizes the terminal Neu5Ac(alpha 2–3)Gal-sequences of glycophorin A. J Cell Biol. 1992, 116: 901-909. 10.1083/jcb.116.4.901.
Sim BK, Chitnis CE, Wasniowska K, Hadley TJ, Miller LH: Receptor and ligand domains for invasion of erythrocytes by Plasmodium falciparum. Science. 1994, 264: 1941-1944.
Peterson DS, Miller LH, Wellems TE: Isolation of multiple sequences from the Plasmodium falciparum genome that encode conserved domains homologous to those in erythrocyte-binding proteins. Proc Natl Acad Sci U S A. 1995, 92: 7100-7104.
Sim BK, Orlandi PA, Haynes JD, Klotz FW, Carter JM, Camus D, Zegans ME, Chulay JD: Primary structure of the 175 K Plasmodium falciparum erythrocyte binding antigen and identification of a peptide which elicits antibodies that inhibit malaria merozoite invasion. J Cell Biol. 1990, 111: 1877-1884. 10.1083/jcb.111.5.1877.
Adams JH, Sim BK, Dolan SA, Fang X, Kaslow DC, Miller LH: A family of erythrocyte binding proteins of malaria parasites. Proc Natl Acad Sci U S A. 1992, 89: 7085-7089.
Orlandi PA, Sim BK, Chulay JD, Haynes JD: Characterization of the 175-kilodalton erythrocyte binding antigen of Plasmodium falciparum. Mol Biochem Parasitol. 1990, 40: 285-294. 10.1016/0166-6851(90)90050-V.
Ware LA, Kain KC, Lee Sim BK, Haynes JD, Baird JK, Lanar DE: Two alleles of the 175-kilodalton Plasmodium falciparum erythrocyte binding antigen. Mol Biochem Parasitol. 1993, 60: 105-109. 10.1016/0166-6851(93)90033-T.
Okenu DM, Riley EM, Bickle QD, Agomo PU, Barbosa A, Daugherty JR, Lanar DE, Conway DJ: Analysis of human antibodies to erythrocyte binding antigen 175 of Plasmodium falciparum. Infect Immun. 2000, 68: 5559-5566. 10.1128/IAI.68.10.5559-5566.2000.
Conway DJ, Cavanagh DR, Tanabe K, Roper C, Mikes ZS, Sakihama N, Bojang KA, Oduola AM, Kremsner PG, Arnot DE, Greenwood BM, McBride JS: A principal target of human immunity to malaria identified by molecular population genetic and immunological analyses. Nat Med. 2000, 6: 689-962. 10.1038/76272.
Binks RH, Baum J, Oduola AM, Arnot DE, Babiker HA, Kremsner PG, Roper C, Greenwood BM, Conway DJ: Population genetic analysis of the Plasmodium falciparum erythrocyte binding antigen-175 (eba-175) gene. Mol Biochem Parasitol. 2001, 114: 63-70. 10.1016/S0166-6851(01)00240-7.
World Health Organization (WHO): Severe falciparum malaria. Trans R Soc Trop Med Hyg. 2000, 94 (suppl 1):
Mockenhaupt FP, Ehrhardt S, Burkhardt J, Bosomtwe SY, Laryea S, Anemana SD, Otchwemah RN, Cramer J, Dietz E, Gellert S, Bienzle U: Manifestation and outcome of severe malaria in children in northern Ghana. Am J Trop Med Hyg.
Djimde A, Doumbo OK, Cortese JF, Kayentao K, Doumbo S, Diourte Y, Dicko A, Su XZ, Nomura T, Fidock DA, Wellems TE, Plowe CV, Coulibaly D: A molecular marker for chloroquine-resistant falciparum malaria. N Engl J Med. 2001, 344: 257-263. 10.1056/NEJM200101253440403.
Touré FS, Mavoungou E, Ndong JM, Tshipamba P, Deloron P: Erythrocyte binding antigen (EBA-175) of Plasmodium falciparum: improved genotype determination by nested polymerase chain reaction. Trop Med Int Health. 2001, 6: 767-769. 10.1046/j.1365-3156.2001.00789.x.
Dittrich S, Schwobel B, Jordan S, Vanisaveth V, Rattanaxay P, Christophel EM, Phompida S, Jelinek T: Distribution of the two forms of Plasmodium falciparum erythrocyte binding antigen-175 (eba-175) gene in Lao PDR. Malar J. 2003, 2: 23-10.1186/1475-2875-2-23.
Färnert A, Rooth I, Svensson A, Snounou G, Björkman A: Complexity of Plasmodium falciparum infections is consistent over time abd protects against clinical disease in Tanzanian children. J Infect Dis. 1999, 179: 989-995. 10.1086/314652.
Owusu-Agyei S, Smith T, Beck H-P, Amenga-Etego L, Felger I: Molecular epidemioloy of Plasmodium falciparum infections among asymptomatic inhabitants of a holoendemic malarious area in northern Ghana. Trop Med Int Health. 2002, 7: 421-8. 10.1046/j.1365-3156.2002.00881.x.
Smith T, Felger I, Tanner M, Beck HP: Premunition in Plasmodium falciparum infection: insights from the epidemiology of multiple infections. Trans R Soc Trop Med Hyg. 1999, 93 (Suppl 1): 59-64.
Binks RH, Conway DJ: The major allelic dimorphisms in four Plasmodium falciparum merozoite proteins are not associated with alternative pathways of erythrocyte invasion. Mol Biochem Parasitol. 1999, 103: 123-127. 10.1016/S0166-6851(99)00115-2.
Acknowledgements
We thank all participating children and their parents. We also thank all members of the Northern Region Malaria Projects (NORMAP) and the Medical Faculty Charité, Humboldt University Berlin, for financial support.
Financial support: Charité grant 2003-676
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
TJand FP Mockenhaupt had the original idea for the study. IM and SD did the polymorphism analysis. FPM, SE, UB and RNO were involved in the recruitment of cases and controls. ED and JPC did statistical analysis. JPC, FPM and TJ wrote the report with major contributions of the other authors.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Cramer, J.P., Mockenhaupt, F.P., Möhl, I. et al. Allelic dimorphism of the erythocyte binding antigen-175 (eba-175) gene of Plasmodium falciparum and severe malaria: Significant association of the C-segment with fatal outcome in Ghanaian children. Malar J 3, 11 (2004). https://doi.org/10.1186/1475-2875-3-11
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-3-11