
Abstract
Background
The aim of this study was to investigate the role of TNF genetic variants and the combined effect between TNF gene and cigarette smoking in the development of gastric cancer in the Korean population.
Methods
We selected 84 incident gastric cancer cases and 336 matched controls nested within the Korean Multi-Center Cancer Cohort. Six SNPs on the TNF gene, TNF-α-238 G/A, -308 G/A, -857 C/T, -863 C/A, -1031 T/C, and TNF-β 252 A/G were genotyped. The ORs (95% CIs) were calculated using unconditional logistic regression model to detect each SNP and haplotype-pair effects for gastric cancer. The combined effects between the TNF gene and smoking on gastric cancer risk were also evaluated. Multi dimensionality reduction (MDR) analyses were performed to explore the potential TNF gene-gene interactions.
Results
TNF-α-857 C/T containing the T allele was significantly associated with an increased risk of gastric cancer and a linear trend effect was observed in the additive model (OR = 1.6, 95% CI 1.0–2.5 for CT genotype; OR = 2.6, 95% CI 1.0–6.4 for TT genotype). All haplotype-pairs that contained TCT or CCC of TNF-α-1031 T/C, TNF-α-863 C/A, and TNF-α-857 C/T were associated with a significantly higher risk for gastric cancer only among smokers. In the MDR analysis, regardless of smoking status, TNF-α-857 C/T was included in the first list of SNPs with a significant main effect.
Conclusion
TNF-α-857 C/T polymorphism may play an independent role in gastric carcinogenesis and the risk for gastric cancer by TNF genetic effect is pronounced by cigarette smoking.
Similar content being viewed by others


Background
Gastric cancer is a multi-factorial disease that requires the study of environmental, genetics, and host-related factors in order to understand its pathology. The strongest risk factor for gastric cancer is Helicobacter pylori (H. pylori) which is labeled a group I human gastric cancer carcinogen by the International Agency for Research on Cancer [1]. However, a high prevalence of H. pylori infection does not always result in a high incidence of gastric cancer. Only a small portion of H. pylori infected persons have gastric cancer, thereby suggesting that other susceptible factors such as genetic variants or environmental differences must additionally be involved in gastric carcinogenesis.
Genetic variants of inflammation-related cytokines are a potential risk factor because H. pylori infection induces chronic inflammation in gastric mucosa which is a critical step in gastric carcinogenesis. One of the major cytokines associated with H. pylori infection is the tumor necrosis factor (TNF) expressed by TNF-α and TNF-β genes [2–6]. Many studies have extensively investigated the association between TNF and gastric cancer [5–15]. Although studies have reported TNF can modify the risk of gastric cancer [5, 8–11], the exact role of TNF as a gastric carcinogen is still controversial.
In terms of environmental factors, cigarette smoking has been suggested to play a crucial role in increasing the risk of gastric cancer. Previous epidemiologic studies indicated that cigarette smoking was an independent risk factor for gastric cancer development [16–21]. Moreover, cigarette smoking may promote gastric cancer development by activating systemic inflammation [22]. Hamajima et al. proposed that cigarette smoking and cytokines such as TNF-α and Interleukin-1B may change normal mucosa to H. pylori infected mucosa that may be a first step towards gastric carcinogenesis [22].
Genetic variants of TNF cytokines and cigarette smoking may play a role in gastric carcinogenesis. They seem to play independent or synergetic roles in gastric cancer but the mechanism is still unclear. Thus, we hypothesized that genetic variants of TNF underlies the association with gastric cancer risk and/or their combined effect with cigarette smoking may modify the risk of gastric cancer. The aim of this study was to assess the role of TNF gene variants and the combined effect between TNF genes and cigarette smoking in the development of gastric cancer in the Korean population.
Methods
Study population and data collection
In this population-based nested case-control study, we selected subjects from the Korean Multi-Center Cancer Cohort (KMCC), a prospective cohort of participants (N = 19,688) recruited from four urban and rural areas in Korea (Haman, Chungju, Uljin, and Youngil) [23]. The participants signed a consent form and completed a detailed standardized interview-based questionnaire on general lifestyle, diet, medical history, and other environmental factors, and blood and spot urine samples were collected and stored at -70°C and -20°C, respectively. All information was from baseline survey questionnaires that were collected before gastric cancer diagnosis. Cancer ascertainment was identified by passive surveillance through record linkages to the national cancer registry, the national death certificate, and the health insurance medical records databases [23]. A total of 136 gastric cancer cases were ascertained in December 2002. Of the 136 cases, 36 cases diagnosed before recruitment and 16 cases that lacked DNA for genotyping were excluded. Eligible non-cancer controls were randomly selected from the KMCC population. Four controls were matched to each gastric cancer case by four variables that were age (± 5 years), sex, residential district, and enrollment year. Finally, 84 gastric cancer cases and 336 controls were included in this study.
Genotyping
TNF-α gene was genotyped for five single nucleotide polymorphisms (SNPs) that were -238 G/A(rs361525), -308 G/A(rs1800629), -857 C/T(rs1799724), -863 C/A(rs1800630), and -1031 T/C(rs1799964). These five polymorphisms were located in the promoter region of TNF-α gene. The TNF-β gene that is synonymous with Lymphotoxin-α (LTA) was genotyped for 252 A/G (rs909253) SNP. Genotyping was performed using SNaPshot method. Ten samples were randomly selected and genotyped for each SNP to check for reliability. All assays were 100% concordant. More than 95% of the total subjects were successfully genotyped for all SNPs.
H. pyloriserum assays
Serum was evaluated to identify IgG antibodies specific for H. pylori and seropositivity of CagA/VacA by Helico Blot 2.1™ (MP Biomedicals Asia Pacific, Singapore) according to the manufacturer's protocol. [24, 25]. The sensitivity and specificity for detecting seropositivity of H. pylori, CagA and VacA of Helico Blot 2.1™ were very high (for sensitivity, 99%, 99% and 93%, respectively; for specificity, 98%, 90% and 88%, respectively) [26].
Statistical analysis
Unconditional logistic regression model was used to calculate the odds ratios (ORs) and 95% confidence intervals (95% CIs) of selected characteristics such as follow time (years), H. pylori seropositivity, alcohol consumption, and cigarette smoking status for gastric cancer risk. To evaluate significant covariates, we reviewed the literature and conducted logistic regression analysis; smoking status, H. pylori infection, and CagA seropositivity were selected. Because of the significant association between age/sex and smoking status, we also included age and sex as major covariates. In the analysis, all five covariates were selected adjusted variables, and smoking status was also selected as a stratified variable.
The Hardy-Weinberg equilibrium assessed allele frequencies using the chi-square test. Both single SNPs and haplotype analysis were performed. To detect each SNP effect for gastric cancer, three genetic models, dominant, co-dominant and additive model, were used. Tests for significance were computed after adjusting for age, sex, smoking status, H. pylori infection, and CagA seropositivity. Also, in order to correct for multiple comparisons, false discovery rate (FDR) that controls the expected proportion of incorrectly rejected null hypotheses (type I errors) was computed [27].
To evaluate putative haplotype blocks, linkage disequilibrium (LD) between loci expressing genetic variation was analyzed using Haploview 4.0. Moreover, pairwise Lewontin's D' and r2 values were calculated with 95% CIs when the genotype frequencies were in Hardy-Weinberg equilibrium [28]. Three haplotype blocks were generated which included the following: 1) only three TNF-α SNPs -857, -863, and -1031; 2) three TNF-α SNPs in block 1 and TNF-β 252; and 3) all six SNPs on TNF-α and TNF-β were generated by the Haploview software using the method suggested by Gabriel et al [29] (Additional file 1).
Based on haplotype analysis, each individual haplotype-pair was estimated with haplotype frequencies greater than 5%. We stratified according to smoking status to assess the combined effect of the TNF gene and smoking for gastric cancer. To evaluate the association between risk for gastric cancer and TNF haplotype-pairs effects, unconditional logistic regression model was used to calculate the ORs and 95% CIs with adjustment for age, sex, H. pylori infection and CagA seropositivity.
Multi dimensionality reduction (MDR) analyses were performed to explore the potential TNF gene-gene interactions. MDR is a non-parametric data mining approach for detecting a potential gene-gene or gene-environment interaction and providing selective models of high-order combination of genes [30, 31]. MDR methodology is described in detail elsewhere [30–32]. TNF gene-gene interaction was evaluated using a naïve Bayes classifier in the context of a 10-fold cross-validation to estimate the test accuracy (TA) of the best 1-, 2-, 3-, and 4- factor models (Additional file 2). The best single interaction model was selected by maximized TA, lowest prediction error or higher cross-validation consistency (CVC). Significance was evaluated using a sign test and was determined at the 0.05 level. Finally, we conducted logistic regression to assess the association between genotype combinations suggested by MDR and gastric cancer with the MDR ORs and 95% CIs. According to stratification by smoking status, MDR ORs were also calculated after adjusting for age, sex, H. pylori infection and CagA seropositivity. All statistical analyses using logistic regression models were performed with SAS version 9.1 and MDR analyses were implemented using MDR software version 11.0 (available at http://www.epistasis.org).
IRB approval
The study protocols for the KMCC study and the present nested case-control study were approved by the institutional review boards of Seoul National University Hospital and the National Cancer Center of Korea (H-0110-084-002, and C-0603-161-170, respectively).
Results
Table 1 presents the odds ratios and 95% CIs of the selected characteristics for gastric cancer risk. There was no significant difference between cases and controls according to age, gender, median follow up period (FU years), education level, H. pylori infection, CagA/VacA seropositivity, and alcohol consumption. All smoking related factors were associated with an increased risk for gastric cancer. Current smokers had a significantly higher risk for gastric cancer compared to never smokers (OR = 1.8, 95% CI 1.0–3.2). Moreover, a dose-response relationship between smoking intensity and gastric cancer risk was found. Smoking more cigarettes per day in addition to longer smoking duration showed a significantly higher risk compared to never smokers (OR = 3.9, 95% CI 1.4–10.7 for more than 20 cigarettes per day; OR = 1.9, 95% CI 1.1–3.5 for more than 20 years). 41 or more pack-years of smoking was related to a statistically increased risk for gastric cancer (OR = 2.3, 95% CI 1.1–4.9).
Table 2 shows the distribution of TNF polymorphisms among cases and controls and the ORs (95% CIs) for gastric cancer risk in relation to TNF genetic polymorphisms. All genotype frequencies of each SNP did not deviate from the Hardy-Weinberg equilibrium (p > 0.1). In single SNPs analysis, TNF-α-857 C/T containing the T allele was significantly associated with an increased risk of gastric cancer. CT genotype of TNF-α-857 showed a 1.7 fold increased risk for gastric cancer in the co-dominant and dominant models that were statistically significant (95% CI 1.0–2.8, identically in both models). However the TT genotype of TNF-α-857 showed an insignificantly increased risk in the co-dominant model (OR = 1.7, 95% CI 0.3–9.2). In the additive model which showed a linear trend effect, the CT genotype of TNF-α-857 showed a 1.6 fold increased risk for gastric cancer (95% CI 1.0–2.5) and the TT genotype of TNF-α-857 had a 2.6 fold increased risk (95% CI 1.0–6.4). Both increase risks for the T allele in the additive model were statistically significant. Under multiple comparison tests by FDR, all p-values of TNF SNPs in each additive model were greater than 0.2.
Table 3 shows the association between haplotype-pairs of TNF gene and gastric cancer risk according to smoking status. In block 1 that is composed of TNF-α-1031 T/C, TNF-α-863 C/A and TNF-α-857 C/T (D' = 1.0 and r2 = 0.037), compared to TCC-TCC without a variant allele, TCC-TCT showed a significantly increased risk for gastric cancer among total subject and smokers (OR = 2.1, 95% CI 1.1–3.9; OR = 2.6, 95% CI 1.2–5.8; respectively) but was not associated with gastric cancer risk among non-smokers (OR = 1.5, 95% CI 0.5–4.0). TCC-CCC was associated with an increased risk only among smokers (OR = 2.9, 95% CI 1.0–8.9). Similar results were represented in block 2 that is composed of the three TNF-α SNPs in block 1 and TNF-β 252 A/G (D' = 1.0 and r2 = 0.184), and block 3 that is composed of all six TNF SNPs (D' = 1.0 and r2 = 0.068). Compared to the most frequent haplotype-pairs GTCC-GTCC in block 2, ATCT-XXXX or ACCC-XXXX that contained TCT or CCC of TNF-α-1031 T/C, TNF-α-863 C/A and TNF-α-857 C/T showed a significantly higher risk among smokers but not among non-smokers (OR = 2.5, 95% CI 1.0–6.8 for ATCT haplotype-pairs; OR = 4.1, 95% CI 1.2–14.2 for ACCC haplotype-pairs). Particularly, among non-smokers, other haplotype-pairs which did not contain ATCT or ACCC were associated with a significantly reduced risk for gastric cancer compared to the reference haplotype-pairs (OR = 0.2, 95% CI 0.1–0.7). Similarly for block 1 and 2, ATCTGG or ACCCXX haplotype-pairs that contained TCT or CCC of TNF-α-1031 T/C, TNF-α-863 C/A and TNF-α-857 C/T were also associated with a significantly higher risk for gastric cancer only among smokers (OR = 3.1, 95% CI 1.0–10.0 for ATCTGG; OR = 4.7, 95% CI 1.2–18.2 for ACCCXX).
Table 4 presents the summary of MDR analysis and association between high risk genotype combinations suggested by MDR analysis and gastric cancer risk. We selected separate best models according to smoking status. The best MDR model for TNF SNPs among total subjects included TNF-α-1031 T/C, TNF-α-863 C/A, TNF-α-857 C/T and TNF-α-308. This model had maximum TA of 0.602 and maximum CVC of 9/10 (p = 0.0015). The global OR adjusted for the age, sex, smoking status, H. pylori infection, and CagA seropositivity was 2.2 (95% CI 1.3–3.6). Regardless of smoking status, TNF-α-857 C/T was included in the first list of SNPs with a significant main effect. TNF-α-857 model among total subjects had TA of 0.554 and CVC of 9/10 but was not significant at the level of 0.0799. Similar results were shown among non-smokers and smokers. Among non-smokers, the model included only TNF-α-857 and had a maximum CVC of 9/10 and a higher prediction for gastric cancer risk of (OR = 1.7, 95% CI 1.0–2.7) though showing minimum TA of 0.603 (p = 0.0636). In contrast, although the model included TNF-β 252 A/G, TNF-α-1031 T/C, TNF-α-857 C/T, and TNF-α-308 G/A had a higher CVC of 8/10 and maximum TA of 0.697 (p = 0.0009), it had a lower predictability for gastric cancer risk since the adjusted global OR by this model was not significant (OR = 1.2, 95% CI 0.7–2.0). The best model of smokers was selected with TNF-α-1031 T/C, TNF-α-863 C/A and TNF-α-857 C/T. This model had a maximum CVC of 10/10, TA of 0.615 (p = 0.0023) and adjusted global OR of 2.0 (95% CI 1.2–3.3). Although the model included TNF-α-1031 T/C, TNF-α-863 C/A, TNF-α-857 C/T, and TNF-α-308 G/A had maximum TA with the lowest p value, its CVC was relatively low (7/10).
Discussion
In our study, TNF-α-857 C/T genetic variant containing the T allele was associated with a significantly increased risk for gastric cancer. SNP-SNP interaction in the TNF gene including TNF-α-857 C/T or TNF-α-1031 T/C genetic variant was associated with a risk for gastric cancer among smokers but not among non-smokers. Moreover this effect of the TNF-α-857 C/T genetic variant was ascertained in both haplotype and MDR analysis.
Previous studies reported that TNF gene polymorphisms modified the risk of gastric cancer [5, 8–11], while others did not find a significant association [6, 7, 12–15]. This inconsistency may be due to the small number of cases, differences in ethnic populations and SNP selection. Two recent meta-analyses assessed the association between TNF-α polymorphisms and risk of gastric cancer [33, 34]. According to these papers, TNF-α-308 polymorphism had a significantly increased risk for gastric cancer but TNF-α-857 polymorphism only showed a marginally significant risk due to the small number of studies. In contrast to the meta-analysis, our results indicated that TNF-α-857 polymorphism was associated with a significantly increased risk for gastric cancer but other SNPs including TNF-α-308 did not show a significant association. Our findings about the single SNP effect of TNF-α-857 polymorphism can help clarify results of the previous meta-analyses. Inconsistency in TNF-α-308 polymorphism may be due to an ethnic difference since TNF-α-308 AA polymorphism is rare in the East Asian population. Also, we were not able to clearly evaluate TNF-α-308 AA polymorphism and gastric cancer because none of our gastric cancer cases had the TNF-α-308 AA polymorphism.
In haplotype-pairs and MDR analysis, we observed a genetic combined effect among TNF SNPs. Individual haplotype-pairs including TNF-α-1031, -863 and -857 were consistently associated with a significantly increased risk of gastric cancer. Compared to the single SNP effect of the TNF-α-857 polymorphism, a greater odds ratio for haplotype-pairs including TNF-α-857 indicated that a synergistic interaction among TNF SNPs was more strongly associated with gastric cancer development. These results were nearly replicated in the MDR analysis. Our consistent findings from the different statistical methodologies are quite meaningful.
TNF cytokine may interact with cigarette smoking to promote gastric cancer development. Biologically, cigarette smoking can activate systemic inflammations [35] and augment the level of TNF-α through changes in inflammatory markers or cytokine level in animal models [36–38]. Although these findings have not been fully reproduced in humans, several papers suggest an indirect connection between cigarette smoking and TNF gene in the etiology of gastric cancer [39–42]. Some studies reported that circulating concentrations of TNF-α were increased and higher TNF-α level was associated with such diseases in smokers compared to non-smokers [39–42]. In one of these studies, higher TNF-α levels were presented among smokers, especially subjects with the 857 T allele and rare haplotype of the TNF-α promoter [39]. Similar to our findings, the study concluded that TNF-α-857 polymorphism was especially susceptible to the hazards of smoking and the TNF cytokine was strongly affected by cigarette smoking.
The combined effect between smoking and TNF gene was definitely expressed in our haplotype-pairs and MDR analysis. Haplotype-pairs formed with TNF-α-1031, -863 and -857 were associated with a significantly greater increased risk for gastric cancer only among smokers but not among non-smokers. Both single SNP and haplotype-pair effect of TNF-α-857 T allele were related with an increased risk among smokers. Haplotype-pairs including TNF-α-1031 and -863 genetic variation that did not show significance for single SNP effect were also associated with a significantly increased risk of gastric cancer among smokers only. In contrast to TNF-α-857, TNF-α-1031 and -863 may interact mutually based on haplotype specificity and not allele specificity, and this effect may be strongly affected by smoking. If only allele specificity by high-producing or mutant alleles plays a role as risk factors for gastric cancer, haplotype-pairs composed of a greater number of high-producing or mutant alleles may have an even greater risk. Based on this hypothesis, CAT haplotype should be the most powerful risk factor for gastric cancer development and CAC haplotype composed of two high-producing alleles should show a greater risk than CCC or TCT haplotype that includes only one high-producing allele. However, we were not able to fully test this hypothesis because only four haplotypes, TCC, TCT, CCC and CAC, were observed in our population. Considering our results, independent allele specificity of TNF-α-857 T and mutual haplotype specificity of TNF-α-1031 and -863 may be more important risk factors rather than the total number of high-producing alleles, especially for smokers. Moreover, these results were reproduced in all haploblocks regardless of different SNP combinations and were nearly replicated in the MDR analysis. This supports our conclusion that the interaction between the TNF gene and smoking may play a crucial role in the etiology of gastric cancer.
Although, to our knowledge, this is the first study to report an interaction between the TNF gene and cigarette smoking on gastric cancer development, our study had several limitations. First, because of a small number of gastric cancer cases and small sample size, we did not have sufficient statistical power and were not able to stratify on different factors. Second, although we had information on cardiac or non-cardiac cancer of gastric cancer cases, because of our small number of cases, we were not able to observe a difference for gastric cancer risk according to cancer type. Moreover, we did not collect information on cancer histology. Finally, a part of SNPs which is related to signal pathway (NF-κB) from H. pylori to cytokine gene expression were genotyped. Other cytokines, such as interleukins and GM-CSF, which are involved in NF-κB pathway, were not considered for analysis so we examined only a small portion of signal pathway for gastric carcinogenesis.
In spite of these limitations, our study had several strengths. This is a population-based, nested case-control study that is free of biases that are common in retrospective studies. Additionally, we matched cases and controls according to basic confounders, such as age and sex, and significant confounding factors selected in the full model. Furthermore, we used various approaches to detect the potential association between genetic and environmental factors on gastric cancer, and derived consistent results through different approaches. Finally, the minor allele frequencies (MAFs) of all cytokine genes analyzed in our study showed very similar results in the Korean, Chinese, and Japanese Hapmap projects [43, 44] and thus, our results are applicable to most East-Asian populations. On the basis of this study results, we will be able to make more conclusive evidence in future studies.
Conclusion
This study demonstrates that TNF-α-857 C/T polymorphism may play an independent role in gastric carcinogenesis and the gene-gene interaction of TNF also affects gastric cancer development. The combined effect between TNF gene and cigarette smoking can be a major risk factor for gastric cancer. Tailored smoking cessation programs should be targeted for smokers with TNF genetic variants. Additional studies with a greater number of cases and information about gastric cancer type and various genes will allow us to conduct stratified analysis to obtain more detailed results that will further clarify the role of genetic and environment factors on gastric cancer.
References
IARC: IARC Monographs on the evaluation of carcinogenic risks to human: Schistosomes, liver flukes and Helicobacter pylori. 1994, Intl Agency for Research on Cancer. Lyon, 61: 177-240.
Zambon CF, Basso D, Navaglia F, Belluco C, Falda A, Fogar P, Greco E, Gallo N, Rugge M, Di Mario F, Plebani M: Pro- and anti-inflammatory cytokines gene polymorphisms and Helicobacter pylori infection: interactions influence outcome. Cytokine. 2005, 29: 141-152. 10.1016/j.cyto.2004.10.013.
Kunstmann E, Epplen C, Elitok E, Harder M, Suerbaum S, Peitz U, Schmiegel W, Epplen JT: Helicobacter pylori infection and polymorphisms in the tumor necrosis factor region. Electrophoresis. 1999, 20: 1756-1761. 10.1002/(SICI)1522-2683(19990101)20:8<1756::AID-ELPS1756>3.0.CO;2-B.
Yea SS, Yang YI, Jang WH, Lee YJ, Bae HS, Paik KH: Association between TNF -alpha promoter polymorphism and Helicobacter pylori cagA subtype infection. J Clin Pathol. 2001, 54: 703-706.
Hamajima N, Shibata A, Katsuda N, Matsuo K, Ito H, Saito T, Tajima K, Tominaga S: Subjects with TNF -A-857TT and -1031TT genotypes showed the highest Helicobacter pylori seropositive rate compared with those with other genotypes. Gastric Cancer. 2003, 6: 230-236. 10.1007/s10120-003-0258-z.
Machado JC, Figueiredo C, Canedo P, Pharoah P, Carvalho R, Nabais S, Castro Alves C, Campos ML, Van Doorn LJ, Caldas C, Seruca R, Carneiro F, Sobrinho-Simões M: A proinflammatory genetic profile increases the risk for chronic atrophic gastritis and gastric carcinoma. Gastroenterology. 2003, 125: 364-371. 10.1016/S0016-5085(03)00899-0.
Qin Z, Krüger-Krasagakes S, Kunzendorf U, Hock H, Diamantstein T, Blankenstein T: Expression of tumor necrosis factor by different tumor cell lines results either in tumor suppression or augmented metastasis. J Exp Med. 1993, 178: 355-360. 10.1084/jem.178.1.355.
Jang WH, Yang YI, Yea SS, Lee YJ, Chun JH, Kim HI, Kim MS, Paik KH: The -238 tumor necrosis factor-alpha promoter polymorphism is associated with decreased susceptibility to cancers. Cancer Lett. 2001, 166: 41-46. 10.1016/S0304-3835(01)00438-4.
Lu W, Pan K, Zhang L, Lin D, Miao X, You W: Genetic polymorphisms of interleukin (IL)-1B, IL-1RN, IL-8, IL-10 and tumor necrosis factor {alpha} and risk of gastric cancer in a Chinese population. Carcinogenesis. 2005, 26: 631-636. 10.1093/carcin/bgh349.
Hou L, El-Omar EM, Chen J, Grillo P, Rabkin CS, Baccarelli A, Yeager M, Chanock SJ, Zatonski W, Sobin LH, Lissowska J, Fraumeni JF, Chow WH: Polymorphisms in Th1-type cell-mediated response genes and risk of gastric cancer. Carcinogenesis. 2007, 28: 118-123. 10.1093/carcin/bgl130.
El-Omar EM, Rabkin CS, Gammon MD, Vaughan TL, Risch HA, Schoenberg JB, Stanford JL, Mayne ST, Goedert J, Blot WJ, FraumeniJF Jr, Chow WH: Increased risk of noncardia gastric cancer associated with proinflammatory cytokine gene polymorphisms. Gastroenterology. 2003, 124: 1193-1201. 10.1016/S0016-5085(03)00157-4.
Sugimoto M, Furuta T, Shirai N, Nakamura A, Xiao F, Kajimura M, Sugimura H, Hishida A: Different effects of polymorphisms of tumor necrosis factor-alpha and interleukin-1 beta on development of peptic ulcer and gastric cancer. J Gastroenterol Hepatol. 2007, 22: 51-59. 10.1111/j.1440-1746.2006.04442.x.
García-González MA, Lanas A, Quintero E, Nicolás D, Parra-Blanco A, Strunk M, Benito R, Angel Simón M, Santolaria S, Sopeña F, Piazuelo E, Jiménez P, Pascual C, Mas E, Irún P, Espinel J, Campo R, Manzano M, Geijo F, Pellisé M, González-Huix F, Nieto M, Espinós J, Titó L, Bujanda L, Zaballa M, Spanish Gastroenterological Association AEG: Gastric cancer susceptibility is not linked to pro-and anti-inflammatory cytokine gene polymorphisms in whites: a Nationwide Multicenter Study in Spain. Am J Gastroenterol. 2007, 102: 1878-1892. 10.1111/j.1572-0241.2007.01423.x.
Kamangar F, Abnet CC, Hutchinson AA, Newschaffer CJ, Helzlsouer K, Shugart YY, Pietinen P, Dawsey SM, Albanes D, Virtamo J, Taylor PR: Polymorphisms in inflammation-related genes and risk of gastric cancer (Finland). Cancer Causes Control. 2006, 17: 117-125. 10.1007/s10552-005-0439-7.
Lee SG, Kim B, Yook JH, Oh ST, Lee I, Song K: TNF /LTA polymorphisms and risk for gastric cancer/duodenal ulcer in the Korean population. Cytokine. 2004, 28: 75-82. 10.1016/j.cyto.2004.06.009.
Ladeiras-Lopes R, Pereira AK, Nogueira A, Pinheiro-Torres T, Pinto I, Santos-Pereira R, Lunet N: Smoking and gastric cancer: systematic review and meta-analysis of cohort studies. Cancer Causes Control. 2008, 19: 689-701. 10.1007/s10552-008-9132-y.
Batty GD, Kivimaki M, Gray L, Smith GD, Marmot MG, Shipley MJ: Cigarette smoking and site-specific cancer mortality: testing uncertain associations using extended follow-up of the original Whitehall study. Ann Oncol. 2008, 19: 996-1002. 10.1093/annonc/mdm578.
Kim Y, Shin A, Gwack J, Jun JK, Park SK, Kang D, Shin HR, Chang SH, Yoo KY: Cigarette smoking and gastric cancer risk in a community-based cohort study in Korea. J Prev Med Public Health. 2007, 40: 467-474. 10.3961/jpmph.2007.40.6.467.
Ozasa K, Japan Collaborative Cohort Study for Evaluation of Cancer: Smoking and mortality in the Japan Collaborative Cohort Study for Evaluation of Cancer (JACC). Asian Pac J Cancer Prev. 2007, 8: 89-96.
Nishino Y, Inoue M, Tsuji I, Wakai K, Nagata C, Mizoue T, Tanaka K, Tsugane S, Research Group for the Development and Evaluation of Cancer Prevention Strategies in Japan: Tobacco smoking and gastric cancer risk: an evaluation based on a systematic review of epidemiologic evidence among the Japanese population. Jpn J Clin Oncol. 2006, 36: 800-807. 10.1093/jjco/hyl112.
Sjödahl K, Lu Y, Nilsen TI, Ye W, Hveem K, Vatten L, Lagergren J: Smoking and alcohol drinking in relation to risk of gastric cancer: a population-based, prospective cohort study. Int J Cancer. 2007, 120: 128-132. 10.1002/ijc.22157.
Hamajima N, Naito M, Kondo T, Goto Y: Genetic factors involved in the development of Helicobacter pylori-related gastric cancer. Cancer Sci. 2006, 97: 1129-1138. 10.1111/j.1349-7006.2006.00290.x.
Yoo KY, Shin HR, Chang SH, Lee KS, Park SK, Kang D, Lee DH: Korean Multi-center Cancer Cohort Study including a Biological Materials Bank (KMCC-I). Asian Pac J Cancer Prev. 2002, 3: 85-92.
Gwack J, Shin A, Kim CS, Ko KP, Kim Y, Jun JK, Bae J, Park SK, Hong YC, Kang D, Chang SH, Shin HR, Yoo KY: CagA-producing Helicobacter pylori and increased risk of gastric cancer: a nested case-control study in Korea. Br J Cancer. 2006, 95: 639-641. 10.1038/sj.bjc.6603309.
Shin A, Shin HR, Kang D, Park SK, Kim CS, Yoo KY: A nested case-control study of the association of Helicobacter pylori infection with gastric adenocarcinoma in Korea. Br J Cancer. 2005, 92: 1273-1275. 10.1038/sj.bjc.6602467.
Park CY, Cho YK, Kodama T, El-Zimaity HM, Osato MS, Graham DY, Yamaoka Y: New serological assay for detection of putative Helicobacter pylori virulence factors. J Clin Microbiol. 2002, 40: 4753-4756. 10.1128/JCM.40.12.4753-4756.2002.
Benjamini Y, Hochberg Y: Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc B. 1995, 57: 289-300.
Barrett JC, Fry B, Maller J, Daly MJ: Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005, 21: 263-265. 10.1093/bioinformatics/bth457.
Gabriel SB, Schaffner SF, Nguyen H, Moore JM, Roy J, Blumenstiel B, Higgins J, DeFelice M, Lochner A, Faggart M, Liu-Cordero SN, Rotimi C, Adeyemo A, Cooper R, Ward R, Lander ES, Daly MJ, Altshuler D: The structure of haplotype blocks in the human genome. Science. 2002, 296: 2225-2229. 10.1126/science.1069424.
Hahn LW, Ritchie MD, Moore JH: Multifactor dimensionality reduction software for detecting gene-gene and gene-environment interactions. Bioinformatics. 2003, 19: 376-382. 10.1093/bioinformatics/btf869.
Moore JH, Gilbert JC, Tsai CT, Chiang FT, Holden T, Barney N, White BC: A flexible computational framework for detecting, characterizing, and interpreting statistical patterns of epistasis in genetic studies of human disease susceptibility. J Theor Biol. 2006, 241: 252-261. 10.1016/j.jtbi.2005.11.036.
Moore JH: Computational analysis of gene-gene interactions using multifactor dimensionality reduction. Expert Rev Mol Diagn. 2004, 4: 795-803. 10.1586/14737159.4.6.795.
Zhang J, Dou C, Song Y, Ji C, Gu S, Xie Y, Mao Y: Polymorphisms of tumor necrosis factor-alpha are associated with increased susceptibility to gastric cancer: a meta-analysis. J Hum Genet. 2008, 53: 479-489. 10.1007/s10038-008-0273-3.
Gorouhi F, Islami F, Bahrami H, Kamangar F: Tumour-necrosis factor-A polymorphisms and gastric cancer risk: a meta-analysis. Br J Cancer. 2008, 98: 1443-1451. 10.1038/sj.bjc.6604277.
Levitzky YS, Guo CY, Rong J, Larson MG, Walter RE, Keaney JF, Sutherland PA, Vasan A, Lipinska I, Evans JC, Benjamin EJ: Relation of smoking status to a panel of inflammatory markers: The Framingham offspring. Atherosclerosis. 2008, 201: 217-224. 10.1016/j.atherosclerosis.2007.12.058.
Zhang C, Cai S, Chen P, Chen JB, Wu J, Wu SJ, Zhou R: Inhibition of tumor necrosis factor-alpha reduces alveolar septal cell apoptosis in passive smoking rats. Chin Med J (Engl). 2008, 121: 597-601.
Brandsma CA, Hylkema MN, Luinge MA, Geerlings M, Klok PA, Cassee FR, Timens W, Postma DS, Kerstjens HA: Nitrogen dioxide exposure attenuates cigarette smoke-induced cytokine production in mice. Inhal Toxicol. 2008, 20: 183-189. 10.1080/08958370701821342.
Churg A, Dai J, Tai H, Xie C, Wright JL: Tumor necrosis factor-alpha is central to acute cigarette smoke-induced inflammation and connective tissue breakdown. Am J Respir Crit Care Med. 2002, 166: 849-854. 10.1164/rccm.200202-097OC.
Bennet AM, van Maarle MC, Hallqvist J, Morgenstern R, Frostegård J, Wiman B, Prince JA, de Faire U: Association of TNF -alpha serum levels and TNF A promoter polymorphisms with risk of myocardial infarction. Atherosclerosis. 2006, 187: 408-414. 10.1016/j.atherosclerosis.2005.09.022.
Gingo MR, Silveira LJ, Miller YE, Friedlander AL, Cosgrove GP, Chan ED, Maier LA, Bowler RP: Tumour necrosis factor gene polymorphisms are associated with COPD. Eur Respir J. 2008, 31: 1005-1012. 10.1183/09031936.00100307.
Jang Y, Koh SJ, Kim OY, Kim BK, Choi D, Hyun YJ, Kim HJ, Chae JS, Lee JH: Effect of the 252A>G polymorphism of the lymphotoxin-alpha gene on inflammatory markers of response to cigarette smoking in Korean healthy men. Clin Chim Acta. 2007, 377: 221-227. 10.1016/j.cca.2006.10.002.
Tappia PS, Troughton KL, Langley-Evans SC, Grimble RF: Cigarette smoking influences cytokine production and antioxidant defences. Clin Sci (Lond). 1995, 88: 485-489.
The International HapMap Project Website. [http://www.hapmap.org/]
The Korean HapMap Project Website. [http://www.khapmap.org/]
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2407/9/238/prepub
Acknowledgements
This study was supported by a grant from the National R&D Program for Cancer Control, Ministry of Health & Welfare, Republic of Korea (0520140), Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2007-313-E00175), and the Seoul National University Hospital (SNUH) research fund (04-2007-030).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
All the authors participated in the design, conduct, and analysis of the study, and approved the final version of this paper.
Electronic supplementary material
12885_2008_1572_MOESM1_ESM.jpeg
{kind=link}
Additional file 1: LD blocks of TNFgene: A) only three TNF-α SNPs -857, -863, and -1031; B) three TNF-α SNPs in block 1 and TNF-β 252; and C) all six SNPs on TNF-α and TNF-β. These LD blocks are generated by the Haploview software using the method suggested by Gabriel et al. (JPEG 67 KB)
12885_2008_1572_MOESM2_ESM.png
{kind=link}
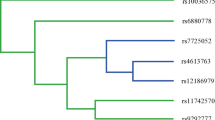
Additional file 2: Dendrograms for TNF genetic variants among A) total subjects, B) smokers, and C) non-smokers modeled by MDR analysis. These dendrograms show high-order combination of TNF genes and indicate TNF gene-gene interaction. (PNG 170 KB)
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Yang, J.J., Ko, KP., Cho, L.Y. et al. The role of TNFgenetic variants and the interaction with cigarette smoking for gastric cancer risk: a nested case-control study. BMC Cancer 9, 238 (2009). https://doi.org/10.1186/1471-2407-9-238
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2407-9-238