
Abstract
Background
Hodgkin lymphoma (HL) is one of the most frequent lymphoma in the western world. Despite a good overall prognosis, some patients suffer relapsing tumors which are difficult to cure. Over a long period Vitamin D has been shown to be a potential treatment for cancer. Vitamin D acts via the vitamin D receptor, a nuclear receptor, acting as an inducible transcription factor. We aimed to investigate the expression of vitamin D receptor as a possible diagnostic marker and potential therapeutic target in HL as well as in B-cell derived non-Hodgkin lymphoma (B-NHL).
Methods
We used a panel of 193 formalin fixed tissues of lymphoma cases consisting of 55 cases of HL and 138 cases on several B-NHL entities.
Results
Vitamin D receptor is strongly expressed in tumor cells of HL, regardless of the sub entity with an overall positivity of 80% of all HL cases. In contrast, only about 17% of the analyzed origin-NHL showed positivity for vitamin D receptor. The detection of nuclear localization of vitamin D receptor in the tumor cells of HL suggests activated status of the vitamin D receptor.
Conclusions
Our study suggests VDR as a specific marker for tumor cells of HL, but not of B-NHL subtypes. Further, the observed nuclear localization suggests an activated receptor status in tumor cells of HL. Further investigations of mutational status and functional studies may shed some light in functional relevance of vitamin D receptor signaling in HL.
Similar content being viewed by others


Background
Hodgkin lymphoma (HL) is one of the most frequent lymphomas. Most of cases belong to the classical (c) subtype of HL (cHL), while only few percent of cases represent nodular lymphocyte predominant HL (NLPHL) [1]. The group of cHL is further subdivided into 4 distinct histological subgroups namely into nodular sclerosis (NS), mixed cellularity (MC), lymphocyte-depleted and lymphocyte-rich HL. In NLPHL, the tumor cells the so called lymphocyte predominant cells (LP), and also the tumor cells of cHL the Hodgkin and Reed-
Sternberg (HRS) cells account for a few percent of total tumor tissue the maiority of the cellular infiltrate comprises of T-lymphocytes, histiocytes, eosinophilic granulocytes and plasma cells [1]. All LP cells and mostly HRS cells are of B-cell origin [2–4]. Despite the HRS cells being mostly of B-cell origin they are lacking mostly of the B-cell expression program [5].
The genetic alterations involved in the pathogenesis of HL are still largely unknown. HRS cells in most if not all cases show numerical chromosome aberrations, amplifications and deletions of chromosomal subregions [6, 7]. In addition, genetic alterations that contribute to constitutive NFκB activation in HRS cells have been identified [8–11]. For a large fraction of cases, infection of HRS-cell precursors with the Epstein-Barr virus (EBV) and expression of the viral latent membrane proteins (LMPs) 1 and 2a is an important pathogenetic event [12]. In HRS cells, many different signaling molecules and pathways such as Notch1, several receptor tyrosine kinases, the PI3K and MEK/ERK pathways and the transcription factors NFκB, STAT and AP-1 are aberrantly activated [13]. These pathways are frequently driven by autocrine as well as paracrine activation loops [14].
Vitamin D3 is a lipophilic molecule that belongs to the family of steroid hormones. Its endocrine functions in regulation of the calcium and phosphate utilization in bone metabolism are known for a long time. Apart from this endocrine character, newer studies added further functions for vitamin D3 especially in the pathogenesis of cancer. It has been shown that vitamin D3 is capable to modulate proliferation, differentiation, metastasis, invasion, angiogenesis and apoptosis via autocrine and paracrine mechanisms [15–18].
Vitamin D3 effects are mainly mediated by its receptor, the vitamin D receptor (VDR) to whom it binds in a high-affinity. Ligand binding is associated with conformational changes of the protein, allowing the interaction with dimerization partners and subsequent nuclear import [19]. VDR is a member of the superfamily of nuclear receptors for steroid hormones and acts as a heterodimer by binding to vitamin D responsive elements as a ligand inducible transcription factor [20]. Nuclear localization saves the active receptor complex from proteasomic degradation [19]. VDR expression can be induced by vitamin D3[21–23].
VDR is detectable by immunohistochemistry in dendritic cells [24]. In proliferating T-cells, VDR expression was shown to be high and correlating with the proliferative status, whereas VDR expression in B-cells is low in general [25, 26]. Dendritic cells with tolerogenic properties can be induced by VDR [27, 28].
Immunohistochemical studies revealed that VDR expression was detectable in most analyzed B-cell Non Hodgkin Lymphoma (B-NHL) cases; however expression was at very low levels compared with normal breast tissue as well as with breast carcinoma in which VDR has been shown to be of importance [29, 30]. In diffuse large B-Cell lymphoma derived cell lines, treatment with vitamin D3 and a selective VDR-agonist reduced cell growth whereas again observed VDR expression was low [29].
In the present study, we investigated the expression of vitamin D receptor in a panel of HL and B-NHL in an immunohistochemical approach in order to identify a potential diagnostic marker and possible new therapeutical target for B-cell derived lymphomas.
Methods
Tissue samples
Tissue samples were retrieved from the files of the Department of Pathology of the University of Frankfurt and were originally submitted for diagnostic purposes. Cases were analyzed after approval was obtained from the University of Frankfurt School of Medicine Institutional Review Board for these studies. The study was performed according to the updated Helsinki declaration and informed consent was obtained from all patients.
Antibodies
Primary antibodies were obtained from Santa Cruz (VDR, #sc13133, Santa Cruz, Heidelberg, Germany) and from Sigma (ß-Actin, Sigma, Deisenhofen, Germany). Secondary antibodies were purchased from Dako (Hamburg, Germany).
Western blot analysis
Tissue lysates were prepared as described [31]. Proteins were separated by electrophoresis on 12,5% polyacrylamide gels containing 0,1% sodium dodecyl sulfate and were transferred to polyvinylidene difluoride membranes (Bio Rad, Munich, Germany) using tank blotting technique. Membranes were blocked with phosphate-buffered saline (PBS) containing 5% dry milk powder and 0,05% Tween20. Primary antibodies were used at dilutions of 1/5000 (for VDR) and 1:10000 (for ß-Actin) in PBS with 5% milk powder, and bound antibody was detected with the appropriate secondary HRP-coupled antibodies (Dako) in 5% milk supplemented with 0,05% Tween20 for 60 min. Signals were detected by using the ECL plus system (GE Healthcare, Munich, Germany), High Performance ECL chemiluminescence films (GE Healthcare) and Readymatic developer and fixer (Carestream Health Inc., NY, USA).
Immunohistochemistry
Immunohistochemistry was performed with 5 μm sections of tissue of complete lymph node biopsies fixed in 5% buffered formalin embedded in paraffin. After dewaxing with xylene, antigen was retrieved by boiling the sections for 10 minutes in a microwave oven in 10 mM Na citrate at pH 6.0. Subsequently sections were blocked with BSA (5% in Tris buffered saline (TBS, Sigma). Sections were incubated for 12 hours with the primary antibody in at 4°C in TBS. Secondary, enzyme coupled mouse Ig-specific antibody (Dako) was incubated for 1 hour at room temperature. Staining was visualized with a catalyzed signal amplification Kit (K1500; Dako). For picture acquisition, an Olympus BX-51 microscope (Olympus, Hamburg, Germany) equipped with UPlanFL and PlanApo objective lenses (10x/0,3, 20x/0,5, and 40x/0,65) was used. Fotographs were processed with Adobe Photoshop 7.0 (Adobe, San Jose, CA).
Reactions lacking primary antibody did not show any noteworthy non specific background.
Statistical analysis
Data analysis was performed using the GraphPad Prism 3.0 software (GraphPad, San Diego, CA). P-value in group comparison was determined using the two-sided Fisher's exact test. The criterion of significance was p < 0.05.
Results and discussion
VDR can be detected in formalin fixed tissue by immunohistochemistry
Like most cell types which are responsive to vitamin D3, also T-lymphocytes express constitutively VDR [32]. B-cells normally lack VDR, but receptor expression can be up regulated by specific stimuli [33–35]. Using RT-PCR, VDR expression could be demonstrated in naïve, germinal center and memory B-cells at low levels [36]. VDR expression in non activated B-cells in tonsils could only be detected after induction by vitamin D3 and interleukin 4 [25, 36, 37].
Using immunohistochemistry (IHC), VDR levels in B-cells in reactive lymph nodes or in hyperplastic tonsils were below detection limits (data not shown), while dendritic cells (DC), in line with previous data [24], stained positive as serving as an internal positive control (data not shown). High expression of VDR in DC served as a further control for the applied IHC method. To validate the IHC staining of VDR, Western blot analyses of B-NHL cases with high and low DC-content as obtained by IHC were performed using the corresponding frozen tissues. Signals for VDR were obtained at the expected molecular weight at around 48 kDa which appeared stronger in a DC-rich case of follicular lymphoma (FL). compared to a weaker signal in a FL-case with a low DC-count. (Figure 1)

Controls for the immunohistochemical staining of VDR. (a) Immunohistochemical staining of VDR of two cases of B-NHL, both follicular lymphoma subtype: one case appearing rich in DC displays strong VDR positivity (case a) compared to another showing only few VDR positive DC (case b). (b) Frozen material of these cases was lysed and analyzed for VDR content by western blotting. In line with the immunohistochemical staining results, high VDR amount was detected in the DC rich case (case a), while a faint band was detected in the DC poor case (case b).
VDR is highly expressed in most HL cases but only in few B-NHL cases
Effects of sunlight, especially ultraviolet radiation regarding the risk of development of lymphoma, are discussed contradictorily [38–41]. Expression of VDR in B-cell lymphoma derived cell lines has been shown immunohistochemically long ago. However, expression levels of VDR in HL are still unknown [29].
We used immunhistochemistry to analyze a panel of B-cell lymphomas being composed of a total of 193 cases, namely 55 cases of HL (26 NSHL, 20 MCHL, 9 NLPHL), 138 cases of B-NHL (38 follicular lymphomas, 59 diffuse large B cell lymphomas, 10 mantle cell lymphomas, 5 marginal zone lymphomas, 6 Burkitt lymphomas, 10 primary mediastinal B cell lymphomas, 10 B cell chronic lymphocytic leukemias). A case was rated positive when more than 50% of the tumor cells displayed VDR-immunoreaction. This threshold was set in order to avoid underestimation of VDR expression in lymphoma cases with inhomogeneous staining due to possible fixation artifacts of the tissue. As seen before, DC also showed positivity in lymphoma cases and therefore served as an internal positive control.
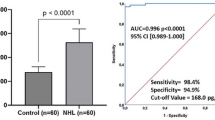
VDR-positivity of tumor cells was observed in 80% (44/55) of HL cases but only in 17.4% (24/138) of total B-NHL entities tested (Figures 2 and 3). Using Fisher’s exact test on contingency tables, statistical analysis of VDR-positive cases revealed statistical significance between the group of HL and B-NHL (p < 0.0001).

Immunohistochemical detection of VDR in HL. Exemplary cases of HL and B-NHL with expression of VDR are shown at low (10x) and high (40x, insert) magnification. The HL entities nodular sclerosing subtype (a), mixed cellularity subtype (b) and nodular lymphocyte predominant Hodgkin Lymphoma (c) are shown. Of B-NHL, samples of diffuse large B-cell lymphoma (d), follicular lymphoma (e), B-cell chronic lymphatic leukemia (f), marginal zone lymphoma (g) and Burkitt´s lymphoma (h) cases with staining reactions scored as positive are shown. In nodular sclerosing subtype of HL (A), also staining of activated T-cells and macrophages can be seen in the non neoplastic cellular background. Also in follicular lymphoma (E) macrophages staining strongly positive can be seen besides the tumor cells. B-NHL showed mostly cytoplasmatic positivity. Nuclear staining was found only in few cells of rare cases.

Immunohistochemical detection of VDR in B-cell lymphoma. 80% of HL showed strong nuclear positivity for VDR while positivity of B-NHL was only seen in 17.4% of cases (a). Among HL subtypes, VDR shows strongest positivity in classical subtypes NSHL (88.5%), followed by MCHL (75%). DLBCL with 22% and FL with 15.8% of positive cases are B-NHL subtypes with most prominent VDR reactivity. 33.3% of BL showed positivity for VDR, however only 6 cases have been analyzed (b).
Among the group of HL, the highest number of positive cases was found in NSHL with 88.5% (23/26) of cases, followed by MCHL with 75% (15/20) and NLPHL in 67.7% (6/9) of positive cases. The number of positive cases in B-NHL was clearly lower at 17.4% of cases (24/138). Of these, most positive cases were among DLBCL at 22% (13/59), FL at 15.8% (6/38) and BL 33.3% (2/6). (Figure 3b) However, only 6 BL could be analyzed so the number of positive cases might be different in a larger cohort. In the vast majority of HL B-NHL cases, HRS cells of HL and LP cells of NLPHL showed a cytoplasmic and also nuclear immunoreaction, whereas VDR was mainly detectable in the cytoplasm of the tumor cells B-NHL cases. Further, VDR staining intensity in tumor cells of HL was much stronger in general when compared to positive B-NHL cases. (Figure 2)
The strong VDR expression and nuclear localization in cHL as well as in NLPHL seems somehow unexpected as these lymphomas have different cellular origins and different marker profiles have been proven [1]. In a global gene expression analysis comparing NLPHL with other lymphoma types it was shown that NLPHL is despite the known differences very closely related to cHL. As cHL NLPHL is also characterized by a loss of marker molecules of B-cells. Both lymphoma entities show a strong constitutive NF-kB activity. Taken together it was shown that their global gene expression profiles differ only in few genes [42]. To go back to histological findings cHL and NLPHL both show a similar mixed cellular background of non neoplastic bystander cells which most likely provide the tumor cells of both entities with an analog pattern of paracrine stimuli.
Upon interaction with vitamin D3, retinoid-X-receptors are recruited to VDR, forming a receptor complex as heterodimerization partners. This complex migrates into the nucleus, binds to vitamin D responsive elements and activates transcription of dependent genes. Only few cell types have been described to show nuclear VDR without previous ligand induced activation [19, 43]. Staining reactions for VDR in HL show mainly nuclear signals. Therefore it seems most likely that also in HRS cells VDR is ligand bound and activated, and thus detectable in the nucleus. (Figure 2a-c) Vitamin D3 activation by sunlight could not be taken as a definite ligand source, since influence of ultraviolet radiation has not been proven to have an influence on lymphoma risk [44]. Thus, activation of VDR raises the question of a possible endogenous source of ligand in tumor tissue. Given the typically low, subnanomolar plasma concentration of vitamin D3, availability of hormone in a suitable, effective concentration requires the presence of an exogenous cell source. Macrophages and monocytes on the one hand have been shown to express vitamin D3 and also been identified to express 1α-hydroxylase, the enzyme that is responsible activating vitamin D3 into its metabolic active form [45, 46]. As these cell types are part the non neoplastic cellular infiltrate of HL, they might be a possible source of ligands acting in a paracrine manner.
About 40% of HL cases, mainly of MC subtype are infected with Epstein-Barr Virus (EBV) [1]. A recent study has shown that the EBV encoded protein EBNA3 is a possible binding partner of VDR, and that EBNA3 - VDR interaction blocks activation of VDR dependent gene expression, which protects EBV immortalized lymphoblast cell lines from vitamin D3 induced growth arrest and apoptosis [47]. As EBNA3 has not been detected in HRS cells, a relevant interaction of VDR with EBNA3 seems unlikely at least in HRS cells [12]. However, the interaction of EBNA3 with VDR in tumor cell progenitors of HL might be advantageous in earlier lymphoma development.
The VDR gene is located on chromosome 12q12-q14. Several single nucleotide polymorphisms of VDR, having possible influences on cancer pathogenesis, have been detected. Exemplarily, polymorphism rs2228570 results in a VDR variant which is three amino acids longer than its wild type and shows a lowered activity compared to the wild type [48]. Polymorphisms rs1544410 and rs731236 do not change VDR protein, but result in enhanced stability of VDR messenger RNA [49, 50]. Association to DLBCL, and increased risk for FL upon exposure to ultraviolet radiation was reported for rs731236 [51, 52]. As tumor cells in HL are very rare, micro dissection experiments would raise a possibility to elucidate genomic vdr mutation status in HL.
Conclusions
In summary, this is the first study to report the strong expression of VDR in the vast majority of HL cases, while only low or none VDR expression was detected in normal, non neoplastic B-cells and all analyzed B-NHL. Nuclear localization of VDR suggests the active form of detected VDR in HL. As VDR activity seems to be of importance in B-NHL, VDR function in HL remains to be elucidated.
References
WHO Classifination of Tumors of the Haematopoetic and Lymphoid Tissues. Edited by: Swerdlow SH, Campo E, Harris NL, Jaffe ES, Pileri SA, Stein H, Thiele J, Vardiman JW. 2008, Lyon, France: International Agency for Research on Cancer (IARC)
Braeuninger A, Kuppers R, Strickler JG, Wacker HH, Rajewsky K, Hansmann ML: Hodgkin and Reed-Sternberg cells in lymphocyte predominant Hodgkin disease represent clonal populations of germinal center-derived tumor B cells. Proc Natl Acad Sci U S A. 1997, 94 (17): 9337-9342. 10.1073/pnas.94.17.9337.
Kuppers R, Rajewsky K, Zhao M, Simons G, Laumann R, Fischer R, Hansmann ML: Hodgkin disease: Hodgkin and Reed-Sternberg cells picked from histological sections show clonal immunoglobulin gene rearrangements and appear to be derived from B cells at various stages of development. Proc Natl Acad Sci U S A. 1994, 91 (23): 10962-10966. 10.1073/pnas.91.23.10962.
Willenbrock K, Ichinohasama R, Kadin ME, Miura I, Terui T, Meguro K, Fukuhara O, DeCoteau JF, Hansmann ML: T-cell variant of classical Hodgkin's lymphoma with nodal and cutaneous manifestations demonstrated by single-cell polymerase chain reaction. Laboratory investigation; a journal of technical methods and pathology. 2002, 82 (9)): 1103-1109.
Schwering I, Brauninger A, Klein U, Jungnickel B, Tinguely M, Diehl V, Hansmann ML, Dalla-Favera R, Rajewsky K, Kuppers R: Loss of the B-lineage-specific gene expression program in Hodgkin and Reed-Sternberg cells of Hodgkin lymphoma. Blood. 2003, 101 (4): 1505-1512. 10.1182/blood-2002-03-0839.
Joos S, Menz CK, Wrobel G, Siebert R, Gesk S, Ohl S, Mechtersheimer G, Trumper L, Moller P, Lichter P, et al: Classical Hodgkin lymphoma is characterized by recurrent copy number gains of the short arm of chromosome 2. Blood. 2002, 99 (4): 1381-1387. 10.1182/blood.V99.4.1381.
Weber-Matthiesen K, Deerberg J, Poetsch M, Grote W, Schlegelberger B: Numerical chromosome aberrations are present within the CD30+ Hodgkin and Reed-Sternberg cells in 100% of analyzed cases of Hodgkin's disease. Blood. 1995, 86 (4): 1464-1468.
Cabannes E, Khan G, Aillet F, Jarrett RF, Hay RT: Mutations in the IkBa gene in Hodgkin's disease suggest a tumour suppressor role for IkappaBalpha. Oncogene. 1999, 18 (20): 3063-3070. 10.1038/sj.onc.1202893.
Emmerich F, Meiser M, Hummel M, Demel G, Foss HD, Jundt F, Mathas S, Krappmann D, Scheidereit C, Stein H, et al: Overexpression of I kappa B alpha without inhibition of NF-kappaB activity and mutations in the I kappa B alpha gene in Reed-Sternberg cells. Blood. 1999, 94 (9): 3129-3134.
Emmerich F, Theurich S, Hummel M, Haeffker A, Vry MS, Dohner K, Bommert K, Stein H, Dorken B: Inactivating I kappa B epsilon mutations in Hodgkin/Reed-Sternberg cells. J Pathol. 2003, 201 (3): 413-420. 10.1002/path.1454.
Jungnickel B, Staratschek-Jox A, Brauninger A, Spieker T, Wolf J, Diehl V, Hansmann ML, Rajewsky K, Kuppers R: Clonal deleterious mutations in the IkappaBalpha gene in the malignant cells in Hodgkin's lymphoma. J Exp Med. 2000, 191 (2): 395-402. 10.1084/jem.191.2.395.
Kuppers R: B cells under influence: transformation of B cells by Epstein-Barr virus. Nature reviews. 2003, 3 (10): 801-812. 10.1038/nri1201.
Brauninger A, Schmitz R, Bechtel D, Renne C, Hansmann ML, Kuppers R: Molecular biology of Hodgkin's and Reed/Sternberg cells in Hodgkin's lymphoma. Int J Cancer. 2006, 118 (8): 1853-1861. 10.1002/ijc.21716.
Khan G: Epstein-Barr virus, cytokines, and inflammation: a cocktail for the pathogenesis of Hodgkin's lymphoma?. Exp Hematol. 2006, 34 (4): 399-406. 10.1016/j.exphem.2005.11.008.
Deeb KK, Trump DL, Johnson CS: Vitamin D signalling pathways in cancer: potential for anticancer therapeutics. Nature reviews. 2007, 7 (9): 684-700. 10.1038/nrc2196.
Dusso AS, Brown AJ, Slatopolsky E: Vitamin D. Am J Physiol Renal Physiol. 2005, 289 (1): F8-F28. 10.1152/ajprenal.00336.2004.
Hansen CM, Frandsen TL, Brunner N, Binderup L: 1 alpha,25-Dihydroxyvitamin D3 inhibits the invasive potential of human breast cancer cells in vitro. Clin Exp Metastasis. 1994, 12 (3): 195-202. 10.1007/BF01753887.
Mantell DJ, Owens PE, Bundred NJ, Mawer EB, Canfield AE: 1 alpha,25-dihydroxyvitamin D(3) inhibits angiogenesis in vitro and in vivo. Circ Res. 2000, 87 (3): 214-220. 10.1161/01.RES.87.3.214.
Peleg S, Nguyen CV: The importance of nuclear import in protection of the vitamin D receptor from polyubiquitination and proteasome-mediated degradation. J Cell Biochem. 2010, 110 (4): 926-934. 10.1002/jcb.22606.
Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, Blumberg B, Kastner P, Mark M, Chambon P, et al: The nuclear receptor superfamily: the second decade. Cell. 1995, 83 (6): 835-839. 10.1016/0092-8674(95)90199-X.
Yu XP, Hustmyer FG, Garvey WT, Manolagas SC: Demonstration of a 1,25-dihydroxyvitamin D3-responsive protein in human lymphocytes: immunologic crossreactivity and inverse regulation with the vitamin D receptor. Proc Natl Acad Sci U S A. 1991, 88 (19): 8347-8351. 10.1073/pnas.88.19.8347.
Strom M, Sandgren ME, Brown TA, DeLuca HF: 1,25-Dihydroxyvitamin D3 up-regulates the 1,25-dihydroxyvitamin D3 receptor in vivo. Proc Natl Acad Sci U S A. 1989, 86 (24): 9770-9773. 10.1073/pnas.86.24.9770.
Lee Y, Inaba M, DeLuca HF, Mellon WS: Immunological identification of 1,25-dihydroxyvitamin D3 receptors in human promyelocytic leukemic cells (HL-60) during homologous regulation. J Biol Chem. 1989, 264 (23): 13701-13705.
Tetlow LC, Smith SJ, Mawer EB, Woolley DE: Vitamin D receptors in the rheumatoid lesion: expression by chondrocytes, macrophages, and synoviocytes. Ann Rheum Dis. 1999, 58 (2): 118-121. 10.1136/ard.58.2.118.
Veldman CM, Cantorna MT, DeLuca HF: Expression of 1,25-dihydroxyvitamin D(3) receptor in the immune system. Arch Biochem Biophys. 2000, 374 (2): 334-338. 10.1006/abbi.1999.1605.
Kizaki M, Norman AW, Bishop JE, Lin CW, Karmakar A, Koeffler HP: 1,25-Dihydroxyvitamin D3 receptor RNA: expression in hematopoietic cells. Blood. 1991, 77 (6): 1238-1247.
Mathieu C, Adorini L: The coming of age of 1,25-dihydroxyvitamin D(3) analogs as immunomodulatory agents. Trends Mol Med. 2002, 8 (4): 174-179. 10.1016/S1471-4914(02)02294-3.
Penna G, Adorini L: 1 Alpha,25-dihydroxyvitamin D3 inhibits differentiation, maturation, activation, and survival of dendritic cells leading to impaired alloreactive T cell activation. J Immunol. 2000, 164 (5): 2405-2411.
Hickish T, Cunningham D, Colston K, Millar BC, Sandle J, Mackay AG, Soukop M, Sloane J: The effect of 1,25-dihydroxyvitamin D3 on lymphoma cell lines and expression of vitamin D receptor in lymphoma. Br J Cancer. 1993, 68 (4): 668-672. 10.1038/bjc.1993.406.
Lopes N, Sousa B, Martins D, Gomes M, Vieira D, Veronese LA, Milanezi F, Paredes J, Costa JL, Schmitt F: Alterations in Vitamin D signalling and metabolic pathways in breast cancer progression: a study of VDR, CYP27B1 and CYP24A1 expression in benign and malignant breast lesions Vitamin D pathways unbalanced in breast lesions. BMC Cancer. 2010, 10: 483-10.1186/1471-2407-10-483.
Al-Shami A, Gilbert C, Barabe F, Gaudry M, Naccache PH: Preservation of the pattern of tyrosine phosphorylation in human neutrophil lysates. J Immunol Methods. 1997, 202 (2)): 183-191.
Jeffery LE, Burke F, Mura M, Zheng Y, Qureshi OS, Hewison M, Walker LS, Lammas DA, Raza K, Sansom DM: 1,25-Dihydroxyvitamin D3 and IL-2 combine to inhibit T cell production of inflammatory cytokines and promote development of regulatory T cells expressing CTLA-4 and FoxP3. J Immunol. 2009, 183 (9): 5458-5467. 10.4049/jimmunol.0803217.
Bhalla AK, Amento EP, Clemens TL, Holick MF, Krane SM: Specific high-affinity receptors for 1,25-dihydroxyvitamin D3 in human peripheral blood mononuclear cells: presence in monocytes and induction in T lymphocytes following activation. J Clin Endocrinol Metab. 1983, 57 (6): 1308-1310. 10.1210/jcem-57-6-1308.
Morgan JW, Morgan DM, Lasky SR, Ford D, Kouttab N, Maizel AL: Requirements for induction of vitamin D-mediated gene regulation in normal human B lymphocytes. J Immunol. 1996, 157 (7): 2900-2908.
Provvedini DM, Tsoukas CD, Deftos LJ, Manolagas SC: 1,25-dihydroxyvitamin D3 receptors in human leukocytes. Science (New York, NY. 1983, 221 (4616)): 1181-1183.
Morgan JW, Kouttab N, Ford D, Maizel AL: Vitamin D-mediated gene regulation in phenotypically defined human B cell subpopulations. Endocrinology. 2000, 141 (9): 3225-3234. 10.1210/en.141.9.3225.
Morgan JW, Sliney DJ, Morgan DM, Maizel AL: Differential regulation of gene transcription in subpopulations of human B lymphocytes by vitamin D3. Endocrinology. 1999, 140 (1): 381-391. 10.1210/en.140.1.381.
Nordstrom M, Hardell L, Magnusson A, Hagberg H, Rask-Andersen A: Occupation and occupational exposure to UV light as risk factors for hairy cell leukaemia evaluated in a case-control study. Eur J Cancer Prev. 1997, 6 (5): 467-472. 10.1097/00008469-199710000-00006.
Bentham G: Association between incidence of non-Hodgkin's lymphoma and solar ultraviolet radiation in England and Wales. BMJ. 1996, 312 (7039): 1128-1131. 10.1136/bmj.312.7039.1128.
Weihkopf T, Becker N, Nieters A, Mester B, Deeg E, Elsner G, Blettner M, Seidler A: Sun exposure and malignant lymphoma: a population-based case-control study in Germany. Int J Cancer. 2007, 120 (11): 2445-2451. 10.1002/ijc.22492.
Grandin L, Orsi L, Troussard X, Monnereau A, Berthou C, Fenaux P, Marit G, Soubeyran P, Huguet F, Milpied N, et al: UV radiation exposure, skin type and lymphoid malignancies: results of a French case-control study. Cancer Causes Control. 2008, 19 (3): 305-315. 10.1007/s10552-007-9093-6.
Brune V, Tiacci E, Pfeil I, Döring C, Eckerle S, van Noesel CJM, Klapper W, Falini B, Von Heyderbeck AMetzler D, et al: Origin and pathogenesis of nodular lymphocyte-predominant Hodgkin lymphoma as revealed by global gene expression analys. J Exp Med. 2008, 205 (10)): 2251-2268.
Ismail A, Nguyen CV, Ahene A, Fleet JC, Uskokovic MR, Peleg S: Effect of cellular environment on the selective activation of the vitamin D receptor by 1alpha,25-dihydroxyvitamin D3 and its analog 1alpha-fluoro-16-ene-20-epi-23-ene-26,27-bishomo-25-hydroxyvitamin D3 (Ro-26-9228). Mol Endocrinol. 2004, 18 (4): 874-887. 10.1210/me.2003-0310.
Smedby KE, Eloranta S, Duvefelt K, Melbye M, Humphreys K, Hjalgrim H, Chang ET: Vitamin D Receptor Genotypes, Ultraviolet Radiation Exposure, and Risk of Non-Hodgkin Lymphoma. Am J Epidemiol. 2011, 173 (1): 48-54. 10.1093/aje/kwq340.
Pryke AM, Duggan C, White CP, Posen S, Mason RS: Tumor necrosis factor-alpha induces vitamin D-1-hydroxylase activity in normal human alveolar macrophages. J Cell Physiol. 1990, 142 (3): 652-656. 10.1002/jcp.1041420327.
Cohen MS, Gray TK: Phagocytic cells metabolize 25-hydroxyvitamin D3 in vitro. Proc Natl Acad Sci U S A. 1984, 81 (3): 931-934. 10.1073/pnas.81.3.931.
Yenamandra SP, Hellman U, Kempkes B, Darekar SD, Petermann S, Sculley T, Klein G, Kashuba E: Epstein-Barr virus encoded EBNA-3 binds to vitamin D receptor and blocks activation of its target genes. Cell Mol Life Sci. 2010, 67 (24)): 4249-4256.
Whitfield GK, Remus LS, Jurutka PW, Zitzer H, Oza AK, Dang HT, Haussler CA, Galligan MA, Thatcher ML: Encinas Dominguez C et al: Functionally relevant polymorphisms in the human nuclear vitamin D receptor gene. Mol Cell Endocrinol. 2001, 177 (1-2): 145-159. 10.1016/S0303-7207(01)00406-3.
Fang Y, Van Meurs JB, D'Alesio A, Jhamai M, Zhao H, Rivadeneira F, Hofman A, van Leeuwen JP, Jehan F, Pols HA, et al: Promoter and 3'-untranslated-region haplotypes in the vitamin d receptor gene predispose to osteoporotic fracture: the rotterdam study. Am J Hum Genet. 2005, 77 (5)): 807-823.
Uitterlinden AG, Fang Y, Van Meurs JB, Pols HA, Van Leeuwen JP: Genetics and biology of vitamin D receptor polymorphisms. Gene. 2004, 338 (2): 143-156. 10.1016/j.gene.2004.05.014.
Purdue MP, Lan Q, Kricker A, Vajdic CM, Rothman N, Armstrong BK: Vitamin D receptor gene polymorphisms and risk of non-Hodgkin's lymphoma. Haematologica. 2007, 92 (8): 1145-1146. 10.3324/haematol.11053.
Purdue MP, Hartge P, Davis S, Cerhan JR, Colt JS, Cozen W, Severson RK, Li Y, Chanock SJ, Rothman N, et al: Sun exposure, vitamin D receptor gene polymorphisms and risk of non-Hodgkin lymphoma. Cancer Causes Control. 2007, 18 (9): 989-999. 10.1007/s10552-007-9039-z.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2407/12/215/prepub
Acknowledgements
We thank Sabine Albrecht for excellent technical assistance and Udo Rüb for help with statistical analyses.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
All authors declare that they have no competing interests.
Authors’ contributions
CR and AHB performed the experimental work. CR and MLH analyzed the data and drafted the manuscript. CR and MLH designed the study. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Renné, C., Benz, A.H. & Hansmann, M.L. Vitamin D3receptor is highly expressed in Hodgkin’s lymphoma. BMC Cancer 12, 215 (2012). https://doi.org/10.1186/1471-2407-12-215
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2407-12-215