
Abstract
Background
Detecting intracellular bacterial symbionts can be challenging when they persist at very low densities. Wolbachia, a widespread bacterial endosymbiont of invertebrates, is particularly challenging. Although it persists at high titers in many species, in others its densities are far below the detection limit of classic end-point Polymerase Chain Reaction (PCR). These low-titer infections can be reliably detected by combining PCR with DNA hybridization, but less elaborate strategies based on end-point PCR alone have proven less sensitive or less general.
Results
We introduce a multicopy PCR target that allows fast and reliable detection of A-supergroup Wolbachia - even at low infection titers - with standard end-point PCR. The target is a multicopy motif (designated ARM: A-supergroup r epeat m otif) discovered in the genome of w Mel (the Wolbachia in Drosophila melanogaster). ARM is found in at least seven other Wolbachia A-supergroup strains infecting various Drosophila, the wasp Muscidifurax and the tsetse fly Glossina. We demonstrate that end-point PCR targeting ARM can reliably detect both high- and low-titer Wolbachia infections in Drosophila, Glossina and interspecific hybrids.
Conclusions
Simple end-point PCR of ARM facilitates detection of low-titer Wolbachia A-supergroup infections. Detecting these infections previously required more elaborate procedures. Our ARM target seems to be a general feature of Wolbachia A-supergroup genomes, unlike other multicopy markers such as insertion sequences (IS).
Similar content being viewed by others


Background
Detecting endosymbionts such as the widespread alphaproteobacterium Wolbachia in its host cell environment requires reliable and ideally simple but still sensitive molecular marker systems. When such bacteria are present at high titers, classic end-point PCR is sufficient to unambiguously determine infection status of an unknown specimen. Particularly for Wolbachia, a quite comprehensive set of diagnostic PCR markers has been developed and applied successfully. The most commonly used among these makers is the multi locus sequence typing (MLST) system [1–3] and the four hypervariable regions (HVRs) of the Wolbachia outer surface protein gene wsp[4, 5]. Both MLST, comprising a set of five singlecopy Wolbachia genes, and the wsp locus were demonstrated to be highly useful for Wolbachia infection determination and consequent diversity assessment. However, those marker systems are limited if the endosymbiont persists at very low titers within the host, either only during a certain ontogenetic stage [6] or throughout all life stages. In both cases proper detection of the endosymbiont is hindered and this points towards the need of an alternative strategy for efficient, robust and fast Wolbachia detection. One approach to address this issue is to use multicopy Wolbachia gene markers for PCR analyses. Particularly insertion sequences (IS; [7, 8]) represent a good strategy to increase the detection threshold [9, 10]. However, this approach relies on the conservation of such elements and their copy-numbers in diverse strains, which might not be the case over longer evolutionary distances due to the mobile nature of these elements. Another approach to cope with the detection problem introduced by low-titer infections is ‘nested PCR’. This method might help to increase the detection threshold but is also highly prone to contamination [6]. A third strategy combines standard PCR with consequent hybridization [6, 11, 12], which increases overall detection limit by four orders of magnitude [6]. On the other hand, this is an elaborate and time-consuming technique. Hence, we set out to find a more sensitive marker for detection of low-titer Wolbachia infections using standard PCR and identified ARM as such a simple but ‘ultra-sensitive’ marker for A-supergroup Wolbachia.
Results and discussion
Identification of a multicopy marker associated with tandem repeats in A-supergroup Wolbachia genomes (ARM)
To find a marker that serves a highly sensitive detection method of low-titer Wolbachia strains we identified multicopy regions in the A-supergroup w Mel genome (Wolbachi a of Drosophila melanogaster; GenBank NC_002978). An intergenic region of 440 bp associated with the recently described hypervariable tandem repeat region (Figure 1; [13]) was the most promising candidate, hereafter called ARM (A-supergroup r epeat m otif) as it was found in 24 almost identical copies dispersed throughout the w Mel genome (Additional file 1). However, for a marker to be useful as a general tool it also needs to be conserved and present in multiple copies in other strains and we therefore used the w Mel repeat sequence to search an additional 13 draft and complete Wolbachia genomes from four different Wolbachia supergroups for the same sequence. We were able to identify the presence of the repeat in seven A-supergroup Wolbachia genomes (w Ha, w Ri, w Wil, w Ana, w Uni, w Suzi and w Gmm; see Table 1), albeit in variable copy numbers. In the Drosophila associated Wolbachia strains, the copy numbers were around 20 per genome (Table 1), whereas the other two A-supergroup genomes (w Uni and w Gmm) contained about half the amount of copies. Low number of hits in w Uni is most likely explained by the incomplete status of the genome resulting in an underestimation of the actual copy number. In the B- (w No, w VitB, w Pip), C- (w Oo, w Ov), and D-supergroup (w Bm) genomes, ARM was not found. Even though some of the genomes in supergroups B, C, and D are incomplete, the total absence of the repeat in all genomes from these supergroups suggests that this motif might be Wolbachia A-supergroup specific. Additionally, VNTR-tandem repeats associated with ARM in A-supergroup infections are also absent from genomes of B- to D-supergroups, further indicating that this feature might indeed be A-supergroup specific.

Schematic presentation of ARM. (A) Position of ARM in association with VNTR-105 locus plus flanking regions in the w Mel genome (GenBank NC_002978). Scheme for VNTR-105 repeat region was adapted from [13] (see this publication for detailed description of VNTR-105 structural features). Black arrows indicate the full 105 bp core repeat segment. Dashed box represents a disrupted segment. ARM (highlighted in yellow) is located within the intergenic region containing the VNTR-105 repeat region. ARM plus repeat region are flanked by WD_1129 (red; NADH-ubiquinone oxidoreductase, putative) on the 5’-prime end and WD_1131 (green; conserved hypothetical protein, degenerate) on the 3’-prime end. (B) Detailed scheme of ARM. The 315 bp PCR amplicon is generated by primer ARM-F (21-mer) and ARM-R (18-mer). Both primers are displayed above and below the PCR amplicon (indicated in yellow).
ARM facilitates detection of low-titer Wolbachia from A-supergoup
ARM-targeting primer were tested via end-point PCR screen on DNA from high- and low-titer Wolbachia infections in Drosophila and Glossina (tsetse fly) species (Additional file 2). As shown in Figure 2, the classic Wolbachia singlecopy gene marker wsp (Wolbachia outer surface protein gene) is only applicable for samples with high-titer infections, since Wolbachia was only detected in high-titer D. paulistorum Orinocan semispecies (OR, Figure 2A) as well as in D. willistoni (Dw +, Figure 2B), D. melanogaster (Dm +, Figure 2B), D. simulans (Ds +, Figure 2B) and Glossina morsitans morsitans (Gmm, Figure 2B). The wsp primer failed to detect Wolbachia in low-titer strains like D. paulistorum Amazonian (AM) and Centroamerican (CA) semispecies plus Glossina swynnertoni (Figure 2A,B), indicating that a singlecopy gene like wsp is not suited for tracking low-titer infections. As multicopy gene markers like insertion sequences (IS) can be used to increase the detection limit, we ran PCR using primer for Insertion Sequence 5 (IS5; [8–10] on the same sample set. We observed increased sensitivity compared to wsp-PCR since Wolbachia was detected in low-titer CA2 (Figure 2A) and in the A/O hybrid samples. However, IS5 primer failed at amplifying the target sequence in all three Glossina samples (Gmm, Gsw and Gs/Gm hybrid; Figure 2B) despite the overall high Wolbachia titer in Gmm[12].

Comparison of Wolbachia marker sensitivity by PCR. (A) The three Wolbachia markers wsp, IS5 and ARM were tested on the following specimens: New world Drosophila species from the Drosophila willistoni group including D. paulistorum Amazonian (AM1, AM2), and Centroamerican (CA1, CA2) semispecies. Orinocan semispecies (OR) served as Wolbachia positive control; Ds - as Wolbachia negative control. B = blank. Quality of DNA was assessed with universal primer set 12SCFR, 12SCRR targeting the mitochondrial 12S rRNA gene [20, 21]. Expected amplicon sizes for Wolbachia positive control (OR) are 631 bp (wsp), 752 bp (IS5), 315 bp (ARM) and 399 bp (12S rRNA). (B) Same markers as above were tested on additional samples including hybrids: A/O hybrid plus parents AM and OR; Glossina Gs/Gm hybrid plus parental strains Gsw and Gmm (Additional file 2). Drosophila New world members include D. willistoni Dw + and Dw -. Old world species are D. melanogaster Dm +; D. simulans Ds + and Ds -. B = blank. Note: IS5 primer set does not produce amplicons in all three Glossina samples due to complete absence of this IS element in symbionts of tsetse flies (see discussion).
We have recently shown that Wolbachia titers increase in D. paulistorum[11] and Glossina[12] hybrid backgrounds, which should significantly facilitate detection and strain characterization. Such titer increase was sufficient to detect Wolbachia with the IS5 primer set in A/O hybrids, but the low-titer Wolbachia infection in the AM mother still remained undetected (Figure 2B). Failure of IS5-amplification in the Gs/Gm hybrid plus parents is explained by lacking homology between primer sequences and target, as no matches with the IS5 primer sequence were found in the w Gmm genome [14]. This finding implies that IS5 is not suitable as a general Wolbachia A-supergroup marker.
Figure 2A and B show that the ARM-marker system can be applied to address aforementioned problems arising with wsp and IS5 primer: sensitivity during PCR is increased significantly and all tested A-supergroup infections are unambiguously detected. Wolbachia was traced in all low-titer New world Drosophila species (AM1, AM2; CA1, CA2) plus the A/O hybrid. In contrast to IS5, the ARM primer set amplified Wolbachia from all three Glossina samples (Gmm, Gsw and Gs/Gm hybrid). As anticipated, all samples from high-titer Wolbachia infections (OR, Dw +, Dm +, Ds +) showed bright bands with ARM, whereas Wolbachia-uninfected specimens (Dw -, Ds -) did not (Figure 2A,B). This argues for a high specificity of the ARM primer and against mis-amplification of a random host target rather than the specific symbiont target site.
Conclusions
We suggest that the new multicopy Wolbachia A-supergroup marker can be used as an ‘ultra-sensitive’ tool to trace low-titer infections by means of classic end-point PCR. First, ARM has the advantage of higher sensitivity compared to classic singlecopy Wolbachia markers like wsp and thus improves detection limit significantly. Particularly, ARM-PCR can be easily applied to screen larger numbers of untyped DNA specimens, even of low quality arising from long-term storage and/or storage in inappropriate media, from laboratory stocks or samples directly from nature. This is of pivotal interest since classical detection tools might yield false negatives when examining species harboring Wolbachia at very low densities, and thereby lead to underestimating natural prevalence of A-supergroup infections. Given that 80% of the Dipteran infections are supergroup A [15], our new method will significantly facilitate and improve the sensitivity of such surveys. In addition our approach is an advantage over the classic IS5-marker, which fails in Wolbachia from the tsetse fly Glossina. Taken together, we show that a Wolbachia sequence motif found in multiple copies associated with the VNTR loci facilitates reliable Wolbachia screening of samples from low-titer infections and might thus serve as a great tool for the Wolbachia research community. Furthermore a similar approach might be applied to detect other symbionts such as Sodalis glossinidius (secondary symbiont of Glossina) and the primary symbiont Candidatus Sodalis pierantonius str. SOPE of the weevil Sitophilus orizae. Both symbiont genomes exhibit more than 20% of repetitive DNA rendering them appropriate candidates for repeat-based PCR analysis [16, 17]. However, we anticipate that such a method reaches its limit when dealing with symbiont genomes, which have become highly streamlined in the course of tight host-symbiont coevolution.
Methods
Drosophila and Glossina strains plus hybrid samples
Drosophila specimens included members of New world and Old world clades (Additional file 2). Representatives of the new world clade were Drosophila paulistorum semispecies AM, CA and OR, together with Wolbachia- infected (Dw +) and -uninfected (Dw -) D. willistoni (see Additional file 2 for details). The Old world clade was represented by Wolbachia- infected D. melanogaster (Dm +) and Wolbachia- infected (Ds +) and uninfected (Ds -) D. simulans (Additional file 2). Additionally, the tsetse fly species Glossina swynnertoni and G. morsitans morsitans (genus Glossina, superfamily Hippoboscoidea) and hybrids from D. paulistorum (A/O) and Glossina (Gs/Gm) were included (Additional file 2). Detailed descriptions of establishing hybrid samples can be found in [11, 12]. Drosophila strains are permanently maintained in the Laboratory of Genome Dynamics in Vienna, Glossina colonies are kept at the Insect Pest Control Laboratory, Joint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, Vienna, Austria.
Analysis of complete and draft Wolbachia genomes for candidate marker loci and primer design
Candidate multicopy marker regions were identified by running nucmer and repeat-match from the MUMmer 3 package [18] on the w Mel genome (Wolbachia, endosymbiont of Drosophila melanogaster; GenBank reference NC_002978). Searches were performed with the megablast algorithm using default settings against 14 Wolbachia genomes present in GenBank (see Table 1; http://www.ncbi.nlm.nih.gov) and other analyses were performed using Geneious 5.6.6 software (Biomatters, New Zealand).
Diagnostic wsp-, IS5-, ARM- and 12S rRNA-PCR
Primer pairs for diagnostic wsp-PCR were taken from [19] and the corresponding PCR set-up is described in [11]. Primers and PCR profile for IS5 can be found in [9]. We designed the following primer set targeting ARM: ARM-F 5’-TTCGCCAATCTGCAGATTAAA-3’ and ARM-R 5’-GTTTTAAACGCTTGACAA-3’. Both primers are positioned in the flanking regions of the VNTR-105 locus in w Mel [9, 13], and produce an amplicon of 315 bp constant size. Composition of the locus is shown in Figure 1. Diagnostic ARM-PCR was performed in 20 μl reactions containing 1x reaction buffer, 3.0 mM MgCl2, 0.4 μM of forward and reverse primer, 35 μM dNTPs, 0.4 U of Taq Polymerase (Promega) and 2 μl of DNA template. PCR was performed using a profile of 2 min initial denaturation at 94°C followed by 30 cycles consisting of 45 sec denaturation at 94°C, 45 sec annealing at 55°C, and 1 min extension at 72°C. Final extension was performed for 10 min at 72°C. In order to assess DNA quality, we amplified part of the mitochondrial 12S rRNA gene with primer set 12SCFR 5′-GAGAGTGACGGGCGATATGT-3′ and 12SCRR 5′-AAACCAGGATTAGATACCCTATTAT-3′ [20]. PCR conditions are outlined in [21]. PCR amplicons were examined using gel-electrophoresis on a 1% agarose gel pre-stained with 0.05 mg ethidium bromide.
Ethics statement
This study did not involve any subjects and materials that require approval by an ethics committee (human, vertebrate, regulated invertebrates). No genetically modified organisms were part of this study.
Abbreviations
- VNTR:
-
Variable number of tandem repeats
- wsp:
-
Wolbachia outer surface protein gene
- IS5:
-
Insertion sequence element 5
- ARM:
-
A-supergroup repeat motif.
References
Maiden MC, Bygraves JA, Feil E, Morelli G, Russell JE, Urwin R, Zhang Q, Zhou J, Zurth K, Caugant DA, Feavers IM, Achtman M, Spratt BG: Multilocus sequence typing: a portable approach to the identification of clones within populations of pathogenic microorganisms. Proc Natl Acad Sci U S A. 1998, 95: 3140-3145. 10.1073/pnas.95.6.3140.
Paraskevopoulos C, Bordenstein SR, Wernegreen JJ, Werren JH, Bourtzis K: Toward a Wolbachia multilocus sequence typing system: discrimination of Wolbachia strains present in Drosophila species. Curr Microbiol. 2006, 53: 388-395. 10.1007/s00284-006-0054-1.
Baldo L, Dunning Hotopp JC, Jolley KA, Bordenstein SR, Biber SA Choudhury RR, Hayashi C, Maiden MC, Tettelin H, Werren JH: Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl Environ Microbiol. 2008, 72: 7098-7110.
Zhou W, Rousset F, O'Neil S: Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc Biol Sci. 1998, 265: 509-515. 10.1098/rspb.1998.0324.
Braig HR, Zhou W, Dobson SL, O'Neill SL: Cloning and characterization of a gene encoding the major surface protein of the bacterial endosymbiont Wolbachia pipientis. J Bacteriol. 1998, 180: 2373-2378.
Arthofer W, Riegler M, Schneider D, Krammer M, Miller WJ, Stauffer C: Hidden Wolbachia diversity in field populations of the European cherry fruit fly, Rhagoletis cerasi (Diptera, Tephritidae). Mol Ecol. 2009, 18: 3816-3830. 10.1111/j.1365-294X.2009.04321.x.
Masui S, Kamoda S, Sasaki T, Ishikawa H: The first detection of the insertion sequence ISW1 in the intracellular reproductive parasite Wolbachia. Plasmid. 1999, 42: 13-19. 10.1006/plas.1999.1407.
Wu M, Sun LV, Vamathevan J, Riegler M, Deboy R, Brownlie JC, McGraw EA, Martin W, Esser C, Ahmadinejad N, Wiegand C, Madupu R, Beanan MJ, Brinkac LM, Daugherty SC, Durkin AS, Kolonay JF, Nelson WC, Mohamoud Y, Lee P, Berry K, Young MB, Utterback T, Weidman J, Nierman WC, Paulsen IT, Nelson KE, Tettelin H, O'Neill SL, Eisen JA: Phylogenomics of the reproductive parasite Wolbachia pipientis w Mel: a streamlined genome overrun by mobile genetic elements. PLoS Biol. 2004, 2: E69-10.1371/journal.pbio.0020069.
Riegler M, Sidhu M, Miller WJ, O'Neill SL: Evidence for a global Wolbachia replacement in Drosophila melanogaster. Curr Biol. 2005, 15: 1428-1433. 10.1016/j.cub.2005.06.069.
Cordaux R: ISWpi1 from Wolbachia pipientis defines a novel group of insertion sequences within the IS5 family. Gene. 2008, 409: 20-27. 10.1016/j.gene.2007.10.035.
Miller WJ, Ehrman L, Schneider D: Infectious speciation revisited: impact of symbiont-depletion on female fitness and mating behavior of Drosophila paulistorum. PLoS Pathog. 2010, 6: e1001214-10.1371/journal.ppat.1001214.
Schneider DI, Garschall KI, Parker AG, Abd-Alla AM, Miller WJ: Global Wolbachia prevalence, titer fluctuations and their potential of causing cytoplasmic incompatibilities in tsetse flies and hybrids of Glossina morsitans subgroup species. J Invertebr Pathol. 2013, 112 (Suppl): S104-S115.
Riegler M, Iturbe-Ormaetxe I, Woolfit M, Miller WJ, O'Neill SL: Tandem repeat markers as novel diagnostic tools for high resolution fingerprinting of Wolbachia. BMC Microbiol. 2012, 12 (Suppl 1): S12-10.1186/1471-2180-12-S1-S12.
Brelsfoard C, Tsiamis G, Falchetto M, Gomulski L, Telleria E, Alam U, Ntountoumis E, Swain M, Malacrida A, Bourtzis K, Aksoy S: Wolbachia symbiont genome sequence and extensive chromosomal insertions described from the tsetse fly Glossina morsitans. 2014, in press: doi:10.1371/journal.pntd.0002728
Stahlhut JK, Desjardins CA, Clark ME, Baldo L, Russell JA, Werren JH, Jaenike J: The mushroom habitat as an ecological arena for global exchange of Wolbachia. Mol Ecol. 2010, 19: 1940-1952. 10.1111/j.1365-294X.2010.04572.x.
Belda E, Moya A, Bentley S, Silva FJ: Mobile genetic element proliferation and gene inactivation impact over the genome structure and metabolic capabilities of Sodalis glossinidius, the secondary endosymbiont of tsetse flies. BMC Genom. 2010, 11: 449-10.1186/1471-2164-11-449.
Oakeson KF, Gil R, Clayton AL, Dunn DM, von Niederhausern AC, Hamil C, Aoyagi A, Duval B, Baca A, Silva FJ, Vallier A, Jackson DG, Latorre A, Weiss RB, Heddi A, Moya A, Dale C: Genome degeneration and adaptation in a nascent stage of symbiosis. Genome Biol Evol. 2014, 6: 76-93. 10.1093/gbe/evt210.
Kurtz S, Phillippy A, Delcher AL, Smoot M, Shumway M, Antonescu C, Salzberg SL: Versatile and open software for comparing large genomes. Genome Biol. 2004, 5: R12-10.1186/gb-2004-5-2-r12.
Jeyaprakash A, Hoy MA: Long PCR improves Wolbachia DNA amplification: wsp sequences found in 76% of sixty-three arthropod species. Insect Mol Biol. 2000, 9: 393-405. 10.1046/j.1365-2583.2000.00203.x.
Hanner R, Fugate M: Branchiopod phylogenetic reconstruction from 12S rDNA sequence data. J Crustacean Biol. 1997, 17: 74-183.
Augustinos AA, Santos-Garcia D, Dionyssopoulou E, Moreira M, Papapanagiotou A, Scarvelakis M, Doudoumis V, Ramos S, Aguiar AF, Borges PA, Khadem M, Latorre A, Tsiamis G, Bourtzis K: Detection and characterization of Wolbachia infections in natural populations of aphids: is the hidden diversity fully unraveled?. PLoS One. 2011, 6: e28695-10.1371/journal.pone.0028695.
Klasson L, Westberg J, Sapountzis P, Näslund K, Lutnaes Y, Darby AC, Veneti Z, Chen L, Braig HR, Garrett R, Bourtzis K, Andersson SG: The mosaic genome structure of the Wolbachia w Ri strain infecting Drosophila simulans. Proc Natl Acad Sci U S A. 2009, 106: 5725-5730. 10.1073/pnas.0810753106.
Elegaard KM, Klasson L, Näslund K, Bourtzis K, Andersson SG: Comparative genomics of Wolbachia and the bacterial species concept. PLoS Genet. 2013, 9: e1003381-10.1371/journal.pgen.1003381.
Salzberg SL, Dunning Hotopp JC, Delcher AL, Pop M, Smith DR, Eisen MB, Nelson WC: Serendipitous discovery of Wolbachia genomes in multiple Drosophila species. Genome Biol 2005, 6:R23. Erratum in. Genome Biol. 2005, 6: 402-10.1186/gb-2005-6-7-402.
Siozios S, Cestaro A, Kaur R, Pertot I, Rota-Stabelli O, Anfora G: Draft Genome Sequence of the Wolbachia Endosymbiont of Drosophila suzukii. Genome Announc. 2013, 1: e00032-13-doi:10.1128/genomeA.00032-13
Kent BN, Salichos L, Gibbons JG, Rokas A, Newton IL, Clark ME, Bordenstein SR: Complete bacteriophage transfer in a bacterial endosymbiont (Wolbachia) determined by targeted genome capture. Genome Biol Evol. 2011, 3: 209-218. 10.1093/gbe/evr007.
Klasson L, Walker T, Sebaihia M, Sanders MJ, Quail MA, Lord A, Sanders S, Earl J, O'Neill SL, Thomson N, Sinkins SP, Parkhill J: Genome evolution of Wolbachia strain w Pip from the Culex pipiens group. Mol Biol Evol. 2008, 25: 1877-1887. 10.1093/molbev/msn133.
Darby AC, Armstrong SD, Bah GS, Kaur G, Hughes MA, Kay SM, Koldkjær P, Rainbow L, Radford AD, Blaxter ML, Tanya VN, Trees AJ, Cordaux R, Wastling JM, Makepeace BL: Analysis of gene expression from the Wolbachia genome of a filarial nematode supports both metabolic and defensive roles within the symbiosis. Genome Res. 2012, 22: 2467-2477. 10.1101/gr.138420.112.
Desjardins CA, Cerqueira GC, Goldberg JM, Chandler M, Mahillon J: Insertion sequences revisited. Edited by Craig NL, Craigie R, Gellert M, Lambowitz AM. Edited by: Mobile DNAII. 2002, Washington, DC: American Society of Microbiology, 305-366.
Foster J, Ganatra M, Kamal I, Ware J, Makarova K, Ivanova N, Bhattacharyya A, Kapatral V, Kumar S, Posfai J, Vincze T, Ingram J, Moran L, Lapidus A, Omelchenko M, Kyrpides N, Ghedin E, Wang S, Goltsman E, Joukov V, Ostrovskaya O, Tsukerman K, Mazur M, Comb D, Koonin E, Slatko B: The Wolbachia genome of Brugia malayi: endosymbiont evolution within a human pathogenic nematode. PLoS Biol. 2005, 3: E121-10.1371/journal.pbio.0030121.
Ehrman L, Powell JR: The Drosophila willistoni species group. Ashburner, Carson, Thompson. 1981-1986, 193-225.
Miller WJ, Riegler M: Evolutionary dynamics of w Au-like Wolbachia variants in Neotropical Drosophila species. Appl Environ Microbiol. 2006, 72: 826-835. 10.1128/AEM.72.1.826-835.2006.
Kidwell MG, Novy JB: Hybrid dysgenesis in Drosophila melanogaster: sterility resulting from gonadal dysgenesis in the P-M system. Genetics. 1979, 92: 1127-1140.
Poinsot D, Montchamp-Moreau C, Merçot H: Wolbachia segregation rate in Drosophila simulans naturally bi-infected cytoplasmic lineages. Heredity (Edinb). 2000, 85 (Pt 2): 191-198.
Acknowledgements
We thank E. Kehrer and M. Leitner for careful maintenance of fly strains in the lab, A. G. Parker and A. M. M. Abd-Alla for providing Glossina material and S. Aksoy from Yale School of Public Health for sharing wGmm genome data. DIS and WJM were partly funded by research grant FWF P22634-B17 from the Austrian Science Fund (FWF).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
DIS and WJM conceived the study. DIS, LK, AEL and WJM designed and performed the experiments. WJM provided material. DIS, LK, AEL and WJM analyzed the data. DIS, LK and WJM wrote the manuscript. All authors read and approved the final version of the manuscript.
Electronic supplementary material
12866_2014_2269_MOESM1_ESM.pptx
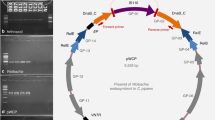
Additional file 1:Positions of ARM in the w Mel and w Ri genomes. Circular schemes of the w Ri (Wolbachia symbiont of Drosophila simulans; NC_012416; [22]) and w Mel genomes (Wolbachia, endosymbiont of D. melanogaster; NC_002978; [8]), showing that ARM (indicated by black bars) is equally dispersed throughout the genomes. (PPTX 171 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Schneider, D.I., Klasson, L., Lind, A.E. et al. More than fishing in the dark: PCR of a dispersed sequence produces simple but ultrasensitive Wolbachia detection. BMC Microbiol 14, 121 (2014). https://doi.org/10.1186/1471-2180-14-121
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2180-14-121