
Abstract
Background
In many species males face a higher predation risk than females because males display elaborate traits that evolved under sexual selection, which may attract not only females but also predators. Females are, therefore, predicted to avoid such conspicuous males under predation risk. The present study was designed to investigate predator-induced changes of female mating preferences in Atlantic mollies (Poecilia mexicana). Males of this species show a pronounced polymorphism in body size and coloration, and females prefer large, colorful males in the absence of predators.
Results
In dichotomous choice tests predator-naïve (lab-reared) females altered their initial preference for larger males in the presence of the cichlid Cichlasoma salvini, a natural predator of P. mexicana, and preferred small males instead. This effect was considerably weaker when females were confronted visually with the non-piscivorous cichlid Vieja bifasciata or the introduced non-piscivorous Nile tilapia (Oreochromis niloticus). In contrast, predator experienced (wild-caught) females did not respond to the same extent to the presence of a predator, most likely due to a learned ability to evaluate their predators' motivation to prey.
Conclusions
Our study highlights that (a) predatory fish can have a profound influence on the expression of mating preferences of their prey (thus potentially affecting the strength of sexual selection), and females may alter their mate choice behavior strategically to reduce their own exposure to predators. (b) Prey species can evolve visual predator recognition mechanisms and alter their mate choice only when a natural predator is present. (c) Finally, experiential effects can play an important role, and prey species may learn to evaluate the motivational state of their predators.
Similar content being viewed by others

Background
Female mate choice has long been recognized as a major driver for character displacement [1–3], or the evolution of novel male traits [4, 5], but can also play a vital role during speciation processes by promoting reproductive isolation through assortative mating [6, 7]. Many mating preferences are innate [1]; still, various extrinsic factors (ecological constraints) may affect individual mating decisions [8–10], such as altered possibilities for mate quality assessment due to increased costs of mate searching [11, 12]. Additionally, the social environment of the choosing individual is known to affect the strength [13–17] or even the direction of mating preferences [18–20].
Another decisive factor acting upon the expression of (female) mating preferences in natural systems is predation risk [21–26]. Female sand gobies (Pomatoschistus minutus), for example, normally prefer larger and more colorful males, but were found to be less choosy when exposed to a predator [27]. Decreased choosiness due to the presence of predators was also reported for male mate choice in the sex-role reversed pipefish Syngnathus typhle [28]. Furthermore, studies on guppies (Poecilia reticulata) revealed that females when facing a predator switch towards associating with less colorful males [29, 30], and females of the green swordtail (Xiphophorus hellerii) that usually prefer males with long swords, switch their preference towards males with short swords when exposed to videos showing successful predator attacks [31].
Such behavioral alterations can be interpreted as a tactic employed by the choosing individuals to reduce their own exposure to predators, as brightly colored males attract predators to the area, and by having more predators in the area females' predation risk is increased [32, 33]. For example, brightly colored males in the Trinidadian guppy are more vulnerable to predation by the predatory cichlids Aequidens pulcher and Crenicichla alta than drabber ones [34–40] and females that preferentially associate with such brightly colored males will obviously face an equally high predation risk.
Our present study was designed to investigate predator-induced changes of female mating preferences in the Atlantic molly (Poecilia mexicana, Poeciliidae). Poecilia mexicana males show a pronounced polymorphism in body size and coloration [41–43], and females prefer larger, more colorful, dominant males as mating partners [42, 44]. At the same time, large molly males are more conspicuous to predators, as exemplified by studies of avian predation on the related sailfin molly, P. latipinna [45], or predation by giant water bugs on P. mexicana [46, 47]. Hence, female mollies might increase their own risk of being attacked by a predator when associating with larger males.
The present study involved piscine predators of P. mexicana, and used cichlid species that regularly co-occur with Atlantic mollies in the same habitats [48–50]. Even though prey choice experiments on the cichlid species considered here (especially 'Cichlasoma' salvini) still need to be conducted, it seems straightforward to assume a preference for larger prey, as another cichlid, the aforementioned pike cichlid (Crenicichla alta), also prefers larger guppies as prey [51, 52]. Adult C. salvini and female P. mexicana show roughly comparable relationships in body size as do C. alta and female P. reticulata (ratio SL P. reticulata, 15-28 mm [53] v. maximum SL C. alta, about 160 mm [53, 54]: 0.10-0.18; ratio SL P. mexicana, 35-40 mm [[55], this study] v. maximum SL C. salvini, about 220 mm [54]: 0.16-0.18), so it seems reasonable to argue that C. salvini would not be gape-limited when preying on average-sized P. mexicana females. Based on these considerations, we predicted that P. mexicana females should alter their mate choice behavior when facing predation risk by piscivorous cichlids; specifically, females should associate with small rather than large males when a piscine predator is around to minimize their own risk of being attacked.
Simultaneously, we asked whether females are able to recognize piscivorous predators and distinguish them from similar non-piscivorous species on the basis of visual cues. We hypothesized that only piscivorous predators would lead to a reversal of female preferences. To test our predictions, we conducted dichotomous female mate choice tests (association preference tests) and repeated the tests while either a natural molly predator ('Cichlasoma' salvini, Cichlidae) or a non-piscivorous fish (three types: two cichlids and another poeciliid, the green swordtail, Xiphophorus hellerii) were presented. This design allowed us to compare changes in the expression of female mating preferences from the 1st to the 2nd part of the tests among four different contexts (i.e. predator treatments). At least for the treatment involving green swordtails-a mainly detritivorous fish [56] that often occurs in the same microhabitats as P. mexicana in southern México and is of similar body size [48, 49]-we predicted that P. mexicana females should not alter their preferences. This treatment, therefore, served as a control to test whether females would be consistent in their mate choice behavior, or if any changes occurred over the course of the experiment that would not be attributable to the presence of a predator.
Finally, we asked the interrelated question of whether visual predator recognition (and the correlated specific responses of females to piscivorous as opposed to non-piscivorous fishes) is innate, or whether also experiential effects/learned predator avoidance could play a role. Comparing the responses of lab-reared (predator-naïve) and wild-caught (predator-experienced) females allowed us to disentangle innate and learned components of predator recognition [57, 58]. We hypothesized that (a) if visual predator recognition mechanisms are entirely innate, then both lab-reared and wild-caught females should respond to the presence of a molly predator during their mate choice, while (b) if experiential effects are important, then a response might not be observable in one of the two different female groups (predator-naïve or wild caught).
In summary, our study aims to answer the following questions: (1) Do P. mexicana females change their mating preferences when exposed to a visually presented piscine predator? (2) Are molly females able to distinguish between predatory species and similar-shaped non-predatory ones on the basis of visual cues alone? (3) Does predator experience affect females' responses to a predator during mate choice?
Methods
Origin and maintenance of study animals
Predator-naïve (lab-reared) test fish were first generation descendents of fish collected from the Río Oxolotán near the town of Tapijulapa, Tabasco, México. They were reared in large (6,000 liters) fish culture tanks at the aquaculture facilities of the Academic Division for Biological Sciences at Universidad Juárez Autónoma de Tabasco (DACBIOL-UJAT) in Villahermosa. Prior to the mate choice experiments fish were kept separated by sex in well-aerated 70-liter tanks at a temperature of 27°C under a natural, approximately 12: 12 hours light: dark cycle.
Predator-experienced (wild-caught) fish were collected in the Río Ixtapangajoya near the city of Teapa, which just like the Río Oxolotán is a tributary of the Río Grijalva. Streams in the Río Grijalva system are widely interconnected in the lowlands at least during the wet season [59], and both collection sites at the Río Oxolotán and the Río Ixtapangajoya are within 20 km river distance. Furthermore, extensive haplotype sharing (based on mt-DNA markers) between mollies from both rivers was recently uncovered [60], so it was legitimate to treat fish from both sampling sites as representatives of the same population for the purpose of this study. Upon capture, fish were transferred into closed and aerated 38 L (43 × 31 × 32 cm) black Sterilite® containers and brought immediately to the laboratory at DACBIOL-UJAT, where they were kept separated by sex in aerated 70-liter tanks for 24 hours to allow acclimation to lab conditions. In the laboratory all fish were fed once a day ad libitum with commercially available flake food.
There is no direct way of easily assessing reproductive status in wild-caught females. However, due to their size (mean SL 34.5 ± 0.6 SEM), all females used in this study were most likely in their reproductive stage (see [55]). Certainly, they may have varied in their reproductive status, but judging from their abdominal distention most of them were likely pregnant. However, as poeciliid females are more receptive to male approaches for few days after giving birth [61, 62], we avoided using post partum females for our tests.
Mate choice tests
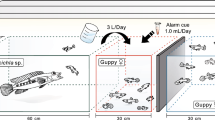
Mate choice experiments were conducted as part of the University course 'Tropical Ecology' of the University of Frankfurt between September 11th and October 17th 2010 at DACBIOL-UJAT, Villahermosa, Tabasco, México. Tests were conducted in three identical portable test tanks (42.6 × 30 × 16.5 cm) built from UV-transparent Plexiglas. Each tank was visually divided into three equally-sized zones by black marks on the outside. The central zone was designated the neutral zone, the two lateral zones as preference zones. Two stimulus males were presented in two smaller auxiliary tanks (19.5 × 30 × 14.5 cm) on either side of the test tank. Hence, the focal female could choose a mate on the basis of visual cues. To avoid disturbance from the outside, we set up all test tanks in large oval tubs that were filled with water to the level inside the test tanks (Figure 1). The entire set-up was placed on a shelf of about 1 m height, and the observer was standing approximately 1.5 to 2 m away from the test apparatus and observed the fish from diagonally above. Therefore, the test fish maximally saw the observer's head, which further helped minimize disturbance of the test fish.

Schematic view of the experimental set-up used to determine female preferences under predation risk. A. During the 1st part of the tests the focal female could choose to associate with either of two stimulus males (large and small). B. Tests were repeated while a predator (here: C. salvini) was presented in an adjacent tank (2nd part). Fishes are not drawn to scale.
Molly males constantly attempt to mate with females (e.g., even directly upon capture [63]; D.B., R.R. and M.P. personal observation). Hence, it is likely that time spent by a female in association with a given male (leading to physical proximity) facilitates male copulation attempts by that particular male. Furthermore, a recent study by Walling et al. [64] experimentally demonstrated that female association preferences actually do translate into male reproductive success in green swordtails, Xiphophorus hellerii (Poeciliidae). We are therefore confident that association preferences in fact translate into more copulations with the preferred male also in P. mexicana.
Before each trial, a large and a small stimulus male was placed into either auxiliary tank [mean (± SEM) SL; large: 45.2 ± 0.5 mm; small: 32.8 ± 0.5 mm]. Then, a female was introduced into the test tank (SL 34.5 ± 0.6 mm). The choice tanks used in this study were relatively small, so focal females were able to see both stimuli at all times. Test fish would typically freeze on the bottom of the test tank for few seconds (to some minutes) after they were introduced, so a trial began only after the focal female had started to swim freely in the water column. We measured the time the female spent in each preference zone during a 5-min observation period. To detect side biases, the stimuli were switched between sides immediately after the first 5-min observation period and measurement was repeated. This episode is henceforth called the 1st part of the tests.
During the 2nd part of our choice tests, we presented a heterospecific audience in a transparent box (a plastic mouse cage measuring 23 × 15 × 16.5 cm) next to the neutral zone outside of the main test tank (Figure 1). The audience was one of four different fish species (treatments 1-4), three of which are found in sympatry in natural P. mexicana habitats in the Río Grijalva drainage [48–50]. The four species differed in nutritional ecology (piscivorous v. non-piscivorous) as well as body shape and coloration (see inserted drawings in Figures 2 and 3). Females were assigned randomly to each of the four treatments.

Time females spent in association with the large and the small stimulus males during the 1st part of the choice tests ( left ) and during the 2nd part of all four predator treatments [treatments (1)-(4), from left to right ]. Depicted are association times (± SEM). Test results are from paired t-tests.

Changes in Atlantic molly females' individual mating preferences when confronted with a piscine predator. Depicted are preference scores (see main text), whereby negative values indicate that female preferences for the initially preferred male decreased in strength.
In treatment (1), females were confronted with a green swordtail (X. hellerii) female (SL: 38.9 ± 1.4 mm) which served as a control for consistency of female preferences, since X. hellerii is a related species with a similar size, appearance, and ecology. In treatment (2) we presented focal females with a Nile tilapia (Oreochromis niloticus; 71.0 ± 1.0 mm). This species was introduced to Mexican freshwaters during the second half of the 20th century [65, 66], and the related species O. aureus is found sympatric with P. mexicana [48]. Nile tilapia is described as a phytoplanktivorous filter feeder [67] and also ingests various plants, detritus, and insects like chironomid larvae [68]. In treatment (3), we confronted females with the algi- and detrivorous cichlid Vieja bifasciata (91.9 ± 2.8 mm; [69, 70]), which is also common in the natural habitats of P. mexicana [48]. Finally, in treatment (4) we used 'Cichlasoma' salvini (98.2 ± 4.0 mm), which is a native omnivorous cichlid in southern México [70, 54] and also includes mollies in its diet (M.P. personal observation). We chose this cichlid species because it is common at both sampling sites ([48]; authors, personal observation) while other, more specialized piscivorous predators (such as Centropomus undecimalis [48]) are comparatively rare and, due to their large body size, less suited for laboratory experiments. We repeated measurements of female association times (including switching of side-assignments of the two stimulus males) as described for the 1st part of the experiments.
Statistical analysis
All data were tested for normality using Kolmogorov-Smirnov-tests. Data are generally given as mean ± standard error (SEM). To evaluate female preferences for large male body size we compared the amount of time focal females spent near the large and small stimulus males during the initial preference test (1st part) as well as during the 2nd part of all four treatments using paired t-tests.
Our central question was whether females would alter their individual choice decisions under the influence of a heterospecific audience. We, therefore, calculated a score expressing the change of female mating decisions [13] as the difference between individual females' relative association times near the initially preferred male during the 2nd part (with predator present) and relative association times near the same male during the 1st part (without predator), such that no change in female preferences would lead to a score of zero, negative values would indicate that the focal females spent less time near the initially preferred male in the 2nd part of a trial, and positive values would indicate that females spent relatively more time near the initially preferred male. Scores were compared among treatments using a fully factorial univariate General Linear Model (GLM) with 'treatment' and 'predator experience' as independent variables. We included 'focal female body size', 'audience body size' as well as 'stimulus male body size difference' (SL large stimulus male-SL small stimulus male) as covariates in our first analysis, but removed 'stimulus male body size difference' (F1,144 = 0.16, P = 0.69) and 'audience body size' (F1,135 = 1.74, P = 0.19) from our final model as they had no statistically significant effects. Where covariates were significant in our final model, standardized residuals from GLM were used to calculate post hoc Pearson correlations.
Results
Female preference for large male body size
Focal females, both from the lab and from the wild, spent significantly more time in association with larger males during the 1st part of the preference tests (Figure 2). Wild-caught females retained a significant preference for the larger of the two stimulus males during the 2nd part in treatment (1) (X. hellerii), treatment (2) (O. niloticus), and treatment (3) (V. bifasciata). Females also spent more time with the larger male in treatment (4) (C. salvini), but this effect was not statistically significant (P < 0.1; Figure 2B).
During the 2nd part of the tests lab-reared (i.e. predator-naïve) females tended to spend more time in association with the larger male in treatment (1), while females tended to spend more time with the smaller male in treatments (2) and (3), but in neither case was the difference statistically significant (Figure 2A). Notably, females spent significantly more time near the smaller male in treatment (4) (with C. salvini).
Changes in individual female preferences
The degree to which individual female preferences changed from the 1st to 2nd part of the tests (preference score; Figure 3) differed significantly between the two types of females (i.e. lab-reared and wild-caught fish; Table 1) as well as among audience treatments (see effect of the factor 'treatment' in Table 1). However, as indicated by a significant interaction term of 'treatment by predator experience' (Table 1), females of the two groups responded differently to the four different types of audience. The pattern is revealed in Figure 3: predator-naïve, lab-reared females altered their preferences when C. salvini was presented, while wild-caught females did not show such responses.
In the GLM 'focal female body size', too, had a significant effect (Table 1), and a post hoc Pearson correlation using standardized residuals revealed a significant negative correlation (rP = -0.18, P = 0.031, n = 145; Figure 4); in other words: larger females were generally less consistent in their mate choice.

The correlation between the strength of change of female preference (preference score) and female body size (SL). Standardized residuals were obtained from a General Linear Model (see Table 1). Test results are from Pearson correlation. Note that larger females were more likely to change their preferences over the course of the experiment.
Discussion
Predation plays a central role as a natural selection factor [71] and affects various aspects of prey species' ecology and behavior. In many cases the male sex is more susceptible to predation [21, 72–76]. For instance, several studies on Trinidadian guppies (Poecilia reticulata) suggest that predation risk has a profound impact on the kind of mating tactics employed by males as well as the evolution of color patterns [62, 77, 78]. Males from 'high-predation' streams, which are characterized by the presence of large cichlid predators, show less courtship behavior and more sneaky copulations, while evolving less conspicuous coloration [62, 79–81].
In the present study we asked (a) if female Atlantic mollies (Poecilia mexicana) would adjust their mating preferences strategically to the presence of a predator, (b) if females would distinguish between piscivorous and similar-shaped non-piscivorous fishes on the basis of visual cues alone by responding alternatively, and (c) if prior experience with predators affects females' responses. We found predator naïve (lab-reared) females to spend more time near the initially non-preferred (smaller) male when confronted with a cichlid during the 2nd part of the tests, but the strongest response-with females exhibiting a significant preference for smaller stimulus males-was found in treatment (4), which involved C. salvini, an omnivorous species that is known to also prey on mollies. A similar effect was reported for green swordtail (Xiphophorus hellerii) females [31]. Females of that species showed a predilection for males with an elongated caudal fin (a sword-like structure) when no predator was around, but preferred males sporting short swords when a predatory cichlid (Petenia splendida) was presented. Since another visual, diurnal swordtail predator, the characid Astyanax mexicanus, has been shown to prefer males with long swords [33], females are thought to reduce their own predation risk by associating with small-sworded males under predation threat.
Females in our study did not change their preferences in treatment (1), which involved a swordtail female as audience. This result is congruent with a previous study reporting on highly consistent mating preferences of P. mexicana females, irrespective of whether choice tests were conducted in front of a con- or heterospecific poeciliid female, or without an audience [82]. By contrast, male poeciliids strongly respond to the presentation of a conspecific audience male by reducing their sexual activity and preference expression [83, 84]-a response that has been interpreted as a tactic employed by males to prevent rivals from copying their mate choice [83, 85, 86]. Our treatment (1), therefore, served as a baseline to make sure that focal females in our present study would indeed be consistent in mate choice over the course of the experiment when no predator was present.
Larger females were generally less consistent in their mate choice than smaller ones (see Figure 4). We do not have a compelling explanation for this surprising finding at hand. However, we tentatively argue that as large P. mexicana females are preferred by males [13, 14, 87] and thus, will attract more males in natural populations (Bierbach et al., unpublished data), larger females could simply be less inclined to associate with a high quality mate for an extended period of time since they are more likely to be approached by yet another (large) male in the near future.
Innate and experiential effects
Our finding that at least lab-reared (predator-naïve) females responded differentially to the four types of audience points towards an innate component of visual predator recognition [88]. Surprisingly, wild-caught (predator experienced) females in our study did not respond to the same extent to a predator as did naïve females. Indeed, wild-caught females still tended to express a preference for large male body size during the 2nd part of the tests even when the predatory C. salvini was presented. At first sight these results seem to contradict other studies on guppies reporting on stronger anti-predator responses in predator experienced (wild-caught) fish compared to lab-reared fish from the same population [89, 90].
So, why did field-collected females (i.e. females that had experienced predators in their natural habitat) not respond to the visual presence of C. salvini, while lab-reared females did so? First, for our experiments we had to use subadult predator specimens that were certainly smaller than the size classes typically preying on mollies in natural habitats. Secondly, even though C. salvini is common in natural molly habitats in southern México [48, 70], sympatric P. mexicana may not always be under immanent predation threat [91]: For example, predators will not represent any risk for some time after a successful catch. Accordingly, we regularly observe groups of P. mexicana (and other poeciliids) in close proximity to groups of C. salvini in their natural habitat, where mollies typically would not show any obvious fright responses even towards large, adult predators (Figure 5). Similar observations were made in the predator-prey interaction between pike cichlids (Crenicichla spp.) and Trinidadian guppies (P. reticulata) [92]. We, therefore, propose that females in nature learn to evaluate the actual level of threat posed by surrounding piscine predators. Experiential effects on anti-predator behaviors are well known from fishes [58, 88], and context-dependent anti-predator responses may be adaptive as they allow saving energy and time that would be wasted when prey species respond indiscriminately [62]. Guppies, for example, display stronger anti-predator responses toward a hungry predator, and predator-experienced fish from a high-predation locality respond more strongly to a hungry predator than those from a low predation locality [91]. We suggest that wild-caught P. mexicana females in our present study have learned to evaluate the predators' motivational state. Predators in our experiment were certainly somewhat stressed due to handling and the relatively small dimensions of their experimental compartment and, therefore, could have presented focal females with visual cues regarding their (lack of) motivation for imminent predation. Future studies will have to answer the question of how exactly mollies determine predators' motivation, but changes in color and movement patterns are the most likely candidates [88].

A shoal of adult Atlantic mollies, P. mexicana ( left arrow, 1 ), as well as another poeciliid ( Heterandria bimaculata; middle, 2 ) in close proximity to a group of C. salvini ( right, 3 ). Note that the mollies showed no obvious fright response. The photo was taken at a site in a clear-water affluent to the "El Azufre", a sulfidic stream in the Cueva del Azufre system [48]. Traces of calcium sulphate give the water a milky appearance.
Conclusion
Overall, our present study highlights that (a) predatory fish can influence the expression of mating preferences of their prey, and females may alter their mate choice behavior strategically to reduce their own exposure to predators. (b) Prey species can evolve visual predator recognition mechanisms and alter their mate choice according to the identity of the audience. (c) Finally, experiential effects also play a role, and prey species may learn to evaluate the motivational state of their predators. Altogether then, our present study underscores the important role played by environmental factors on the expression of mating preferences and adds to our understanding of the multiple pathways by which predation affects prey populations.
References
Andersson M: Sexual selection. 1994, Princeton, Princeton University Press
Gibson RM, Langen TA: How do animals choose their mates?. Trends Ecol Evol. 1996, 11: 468-470. 10.1016/0169-5347(96)10050-1.
Clutton-Brock TH: Sexual Selection in males and females. Science. 2007, 318: 1882-1885. 10.1126/science.1133311.
Basolo AL: Female preference predates the evolution of the sword in swordtail fish. Science. 1990, 250: 808-810. 10.1126/science.250.4982.808.
Schlupp I, Riesch R, Tobler M, Plath M, Parzefall J, Schartl M: A novel sexually selected trait in poeciliid fishes: Female preference for mustache-like, rostral filaments in male Poecilia sphenops. Behav Ecol Sociobiol. 2010, 64: 1849-1855. 10.1007/s00265-010-0996-y.
Tobler M, Riesch R, Tobler CM, Schulz-Mirbach T, Plath M: Natural and sexual selection against immigrants maintains differentiation among micro-allopatric populations. J Evol Biol. 2009, 22: 2298-2304. 10.1111/j.1420-9101.2009.01844.x.
Plath M, Riesch R, Oranth A, Dzienko J, Karau N, Schießl A, Stadler S, Wigh A, Zimmer C, Arias-Rodriguez L, Schlupp I, Tobler M: Complementary effect of natural and sexual selection against immigrants maintains differentiation between locally adapted fish. Naturwissenschaften. 2010, 97: 769-774. 10.1007/s00114-010-0691-x.
Jennions MD, Petrie M: Variation in mate choice and mating preferences: a review of causes and consequences. Biol Rev Camb Phil Soc. 1997, 72: 283-327. 10.1017/S0006323196005014.
Widemo F, Sæther SA: Beauty is in the eye of the beholder: causes and consequences of variation in mating preferences. Trends Ecol Evol. 1999, 14: 26-31. 10.1016/S0169-5347(98)01531-6.
Cotton S, Small J, Pomiankowski A: Sexual selection and condition-dependent mate preferences. Curr Biol. 2006, 16: R755-R765. 10.1016/j.cub.2006.08.022.
Milinski M, Bakker TCM: Costs influence sequential mate choice in stickleback, Gasterosteus aculeatus. Proc R Soc Lond Series B. 1992, 250: 229-233. 10.1098/rspb.1992.0153.
Wong BBM, Jennions MD: Costs influence male mate choice in a freshwater fish. Proc R Soc Lond Biol Sci. 2003, 270: 36-38. 10.1098/rsbl.2003.0003.
Plath M, Blum D, Schlupp I, Tiedemann R: Audience effect alters mating preferences in Atlantic molly (Poecilia mexicana) males. Anim Behav. 2008, 75: 21-29. 10.1016/j.anbehav.2007.05.013.
Plath M, Richter S, Tiedemann R, Schlupp I: Male fish deceive competitors about mating preferences. Curr Biol. 2008, 18: 1138-1141. 10.1016/j.cub.2008.06.067.
Ziege M, Mahlow K, Hennige-Schulze C, Kronmarck C, Tiedemann R, Streit B, Plath M: Audience effects in the Atlantic molly (Poecilia mexicana)-prudent male mate choice in response to perceived sperm competition risk?. Front Zool. 2009, 6: 17-10.1186/1742-9994-6-17.
Bierbach D, Girndt A, Hamfler S, Klein M, Müksch F, Penshorn M, Schwinn M, Zimmer C, Schlupp I, Streit B, Plath M: Male fish use prior knowledge about rivals to adjust their mate choice. Biology Letters. 2011, online first
Bierbach D, Kronmarck C, Hennige-Schulz C, Stadler S, Plath M: Sperm competition risk affects male mate choice copying. Behav Ecol Sociobiol. 2011, online first
Dugatkin LA: Sexual selection and imitation: females copy the mate choice of others. Am Nat. 1992, 139: 1384-1389. 10.1086/285392.
Earley RL, Dugatkin LA: Fighting, mating and networking: pillars of poeciliid sociality. Animal communication networks. Edited by: McGregor PK. 2005, Cambridge: Cambridge University Press, 84-113.
Witte K: Learning and mate choice. Fish cognition and behavior. Edited by: Brown C, Laland KN, Krause J. 2006, Oxford: Blackwell Publishing, 70-95.
Magnhagen C: Predation risk as a cost of reproduction. Trends Ecol Evol. 1991, 6: 183-186. 10.1016/0169-5347(91)90210-O.
Sih A: Predation risk and the evolutionary ecology of reproductive behavior. J Fish Biol. 1994, 45 (Suppl A): 111-130.
Candolin U: Predation risk affects courtship and attractiveness of competing threespine stickleback males. Behav Ecol Sociobiol. 1997, 41: 81-87. 10.1007/s002650050367.
Ellis JMS: Anti-predator signals as advertisements: evidence in white-throated magpie-jays. Ethology. 2009, 115: 522-532. 10.1111/j.1439-0310.2009.01631.x.
Kim TW, Christy JH, Dennenmoser S, Choe JC: The strength of a female mate preference increases with predation risk. Proc R Soc Lond Biol Sci. 2009, 276: 775-780. 10.1098/rspb.2008.1070.
Greig EI, Pruett-Jones S: Danger may enhance communication: predator calls alert females to male displays. Behav Ecol. 2010, 21: 1360-1366. 10.1093/beheco/arq155.
Forsgren E: Predation risk affects mate choice in a gobiid fish. Am Nat. 1992, 140: 1041-1049. 10.1086/285455.
Berglund A: Risky sex: male pipefishes mate at random in presence of a predator. Anim Behav. 1993, 46: 169-175. 10.1006/anbe.1993.1172.
Godin J-GJ, Briggs SE: Female mate choice under predation risk in the guppy. Anim Behav. 1996, 51: 117-130. 10.1006/anbe.1996.0010.
Gong A, Gibson RM: Reversal of a female preference after visual exposure to a predator in the guppy, Poecilia reticulata. Anim Behav. 1996, 52: 1007-1015. 10.1006/anbe.1996.0248.
Johnson JB, Basolo AL: Predator exposure alters female mate choice in the green swordtail. Behav Ecol. 2003, 14: 619-625. 10.1093/beheco/arg046.
Zuk M, Kolluru GR: Exploitation of sexual signals by predators and parasitoids. Quart Rev Biol. 1998, 73: 415-438. 10.1086/420412.
Rosenthal GG, Martinez TYF, de Leon FJG, Ryan MJ: Shared preferences by predators and females for male ornaments in swordtails. Am Nat. 2001, 158: 146-154. 10.1086/321309.
Haskins CP, Haskins EF, McLaughlin JJA, Hewitt RE: Polymorphism and population structure in Lebistes reticulatus, an ecological study. Vertebrate Speciation. Edited by: Blair WF. 1961, Austin: University of Texas Press, 320-395.
Seghers BH: An analysis of geographic variation in the antipredator adaptions of the guppy Poecilia reticulata. 1973, University of British Columbia, PhD Thesis
Endler JA: Natural selection on color patterns in Poecilia reticulata. Evolution. 1980, 34: 76-91. 10.2307/2408316.
Endler JA: Natural and sexual selection on color patterns in poeciliid fishes. Environ Biol Fish. 1983, 9: 173-190. 10.1007/BF00690861.
Gong A: The effects of predator exposure on the female choice of guppies (Poecilia reticulata) from a high-predation population. Behaviour. 1995, 134: 373-389.
Reznick DN, Butler MJ, Rodd FH, Ross P: Life-history evolution in guppies (Poecilia reticulata). Differential mortality as a mechanism for natural selection. Evolution. 1996, 50: 1651-1660. 10.2307/2410901.
Godin JGJ, McDonough HE: Predator preference for brightly colored males in the guppy: a viability cost for a sexually selected trait. Behav Ecol. 2003, 14: 194-200. 10.1093/beheco/14.2.194.
Parzefall J: Zur vergleichenden Ethologie verschiedener Mollienesia-Arten einschließlich einer Höhlenform von Mollienesia sphenops. Behaviour. 1969, 33: 1-37. 10.1163/156853969X00297.
Tobler M, Schlupp I, Plath M: Does divergence in female mate choice affect male size distributions in two cave fish populations?. Biol Lett. 2008, 4: 452-454. 10.1098/rsbl.2008.0259.
Riesch R, Plath M, Schlupp I: Toxic hydrogen sulphide and dark caves: pronounced male life-history divergence among locally adapted Poecilia mexicana (Poeciliidae). J Evol Biol. 2011, 24: 596-606. 10.1111/j.1420-9101.2010.02194.x.
Plath M, Parzefall J, Körner KE, Schlupp I: Sexual selection in darkness? Female mating preferences in surface- and cave-dwelling Atlantic mollies, Poecilia mexicana (Poeciliidae, Teleostei). Behav Ecol Sociobiol. 2004, 55: 596-601. 10.1007/s00265-003-0750-9.
Trexler J, Tempe R, Travis J: Size-selective predation of Sailfin mollies by two species of heron. Oikos. 1994, 69: 250-259. 10.2307/3546145.
Plath M, Parzefall J, Schlupp I: The role of sexual harassment in cave- and surface-dwelling populations of the Atlantic molly, Poecilia mexicana (Poeciliidae, Teleostei). Behav Ecol Sociobiol. 2003, 54: 303-309. 10.1007/s00265-003-0625-0.
Tobler M, Schlupp I, Plath M: Predation of a cave fish (Poecilia mexicana, Poeciliidae) by a giant water-bug (Belostoma, Belostomatidae) in a Mexican sulfur cave. Ecol Entomol. 2007, 32: 492-495. 10.1111/j.1365-2311.2007.00892.x.
Tobler M, Schlupp I, Heubel KU, Riesch R, García de León FJ, Giere O, Plath M: Life on the edge: Hydrogen sulfide and the fish communities of a Mexican cave and surrounding waters. Extremophiles. 2006, 10: 577-585. 10.1007/s00792-006-0531-2.
Riesch R, Schlupp I, Tobler M, Plath M: Reduction of the association preference for conspecifics in surface- and cave-dwelling Atlantic mollies, Poecilia mexicana. Behav Ecol Sociobiol. 2006, 60: 794-802. 10.1007/s00265-006-0223-z.
Riesch R, Duwe V, Herrmann N, Padur L, Ramm A, Scharnweber K, Schulte M, Schulz-Mirbach T, Ziege M, Plath M: Variation along the shy-bold continuum in extremophile fishes (Poecilia mexicana, P. sulphuraria). Behav Ecol Sociobiol. 2009, 63: 1515-1526. 10.1007/s00265-009-0780-z.
Pocklington R, Dill L: Predation on females or males: who pays for bright male traits. Anim Behav. 1995, 49: 1122-1124. 10.1006/anbe.1995.0141.
Johansson J, Turesson H, Persson A: Active selection for large guppies, Poecilia reticulata, by the pike cichlid, Crenicichla saxatilis. Oikos. 2004, 105: 595-605. 10.1111/j.0030-1299.2004.12938.x.
Reznick DN, Butler MJ, Rodd FH, Ross P: Life-history evolution in guppies (Poecilia reticulata) 6. Differential mortality as a mechanism for natural selection. Evolution. 1996, 50: 1651-1660. 10.2307/2410901.
Kullander SO: Cichlidae (Cichlids). Checklist of the Freshwater Fishes of South and Central America. Edited by: Reis RE, Kullander SO, Ferraris CJ Jr. 2003, BPorto Alegre, EDIPUCRS, 3605-3654.
Riesch R, Plath M, Schlupp I: Toxic hydrogen sulfide and dark caves: life history adaptations in a livebearing fish (Poecilia mexicana, Poeciliidae). Ecology. 2010, 95: 1494-1505.
Arthington AH: Diet of Gambusia affinis holbrooki, Xiphophorus helleri, X. maculatus and Poecilia reticulata (Pisces: Poeciliidae) in streams of southeastern Queensland, Australia. Asian Fish Sci. 1989, 2: 193-212.
Cox JG, Lima SL: Naiveté and an aquatic-terrestrial dichotomy in the effects of introduced predators. Trends Ecol Evol. 2006, 21: 674-680. 10.1016/j.tree.2006.07.011.
Brown GE, Chivers DP: Learning as an adaptive response to predation. Ecology of predator-prey interactions. Edited by: Barbosa P, Castellanos I. 2005, Oxford: Oxford University Press, 34-54.
Miller RR: Geographical distribution of Central American freshwater fishes. Copeia. 1966, 773-802.
Tobler M, Palacios M, Chapman L, Mitrofanov I, Bierbach D, Plath M, Arias-Rodriguez L, García de León FJ, Mateos M: Evolution in extreme environments: replicated phenotypic differentiation in livebearing fish inhabiting sulfidic springs. Evolution. 2011,
Plath M, Körner KE, Schlupp I, Parzefall J: Sex recognition and female preferences of cave mollies Poecilia mexicana (Poeciliidae, Teleostei) in light and darkness. Mém Biospéol. 2001, 28: 163-167.
Magurran AE: Evolutionary ecology: the Trinidadian guppy. 2005, Oxford, Oxford University Press
Riesch R, Schlupp I, Plath M: Female sperm-limitation in natural populations of a sexual/asexual mating-complex (Poecilia latipinna, P. formosa). Biol Lett. 2008, 4: 266-269. 10.1098/rsbl.2008.0019.
Walling CA, Royle NJ, Lindström J, Metcalfe NB: Do female association preferences predict the likelihood of reproduction?. Behav Ecol Sociobiol. 2010, 64: 541-548. 10.1007/s00265-009-0869-4.
Arredondo FJL: Especies animales acuáticas de importancia nutricional introducidas en México. Biotica. 1983, 8: 175-199.
Arredondo FJL, Lozano GS: El cultivo de la tilapia en México. Primer curso internacional de producción de tilapia. Edited by: Escamilla MM, Rana AP. 1996, Mexico, Facultad de Medicina Veterinaria y Zootecnia, UNAM, SEMARNAP, 7-18.
Robinson RL, Turner GF, Grimm AS, Pitcher TJ: A comparison of the ingestion rates of three tilapia species fed on a small planctonic alga. J Fish Biol. 1990, 36: 269-270. 10.1111/j.1095-8649.1990.tb05603.x.
Asano Y, Hayashizaki K, Eda H, Khonglaliang T, Kurokura H: Natural foods utilized by Nile tilapia, Oreochromis niloticus, in fertilizer-based fish ponds in Lao PDR identified through stable isotope analysis. Fish Sci. 2010, 76: 811-817. 10.1007/s12562-010-0271-1.
Conkel D: Cichlids of North and Central America. 1993, USA, T.F.H. Publications
Miller RR, Minckley WL, Norris SM: Freshwater Fishes of México. 2005, Chicago, Chicago University Press
Endler JA: Natural Selection in the Wild. Monographs in Population Biology. 1986, Princeton, Princeton University Press, 21:
Burk T: Evolutionary significance of predation on sexually signalling males. Florida Entomol. 1982, 65: 90-104. 10.2307/3494148.
Macías Garcia C, Saborío E, Berea C: Does male biased predation lead to male scarcity in viviparous fish?. J Fish Biol. 1998, 53: 104-117. 10.1111/j.1095-8649.1998.tb01021.x.
Lode T, Holveck M, Lesbarreres D, Pagano A: Sex-biased predation by polecats influences the mating system of frogs. Proc R Soc Lond Series B. 2004, 271: S399-S401. 10.1098/rsbl.2004.0195.
Costantini D, Bruner E, Fanfani A, Dell'Omo G: Male-biased predation of western lizards by Eurasian kestrels. Naturwissenschaften. 2007, 94: 1015-1020. 10.1007/s00114-007-0284-5.
Tobler M, Franssen C, Plath M: Male-biased predation of a cave fish by a giant water bug. Naturwissenschaften. 2008, 95: 775-779. 10.1007/s00114-008-0382-z.
Endler JA: A predator's view of animal color patterns. Evol Biol. 1978, 11: 319-364.
Endler JA, Houde AE: Geographic variation in female preferences for male traits in Poecilia reticulata. Evolution. 1995, 49: 456-468. 10.2307/2410270.
Endler JA: Predation, light intensity and courtship behaviour in Poecilia reticulata (Pisces: Poeciliidae). Anim Behav. 1987, 35: 1376-1385. 10.1016/S0003-3472(87)80010-6.
Houde AE: Sex, color, and mate choice in guppies. 1997, Princeton, Princeton University Press
Evans JP, Kelly JL, Ramnarine IW, Pilastro A: Female behavior mediates male courtship under predation risk in the guppy (Poecilia reticulata). Behav Ecol Sociobiol. 2002, 52: 496-502. 10.1007/s00265-002-0535-6.
Plath M, Kromuszczynski K, Tiedemann R: Audience effect alters male but not female mating preferences. Behav Ecol Sociobiol. 2009, 63: 381-390. 10.1007/s00265-008-0672-7.
Plath M, Richter S, Tiedemann R, Schlupp I: Male fish deceive competitors about mating preferences. Curr Biol. 2008, 18: 1138-1141. 10.1016/j.cub.2008.06.067.
Plath M, Blum D, Schlupp I, Tiedemann R: Audience effect alters mating preferences in Atlantic molly (Poecilia mexicana) males. Anim Behav. 2008, 75: 21-29. 10.1016/j.anbehav.2007.05.013.
Plath M, Schlupp I: Misleading mollies-the effect of an audience on the expression of mating preferences. Comm Integr Biol. 2008, 1: 199-203. 10.4161/cib.1.2.7199.
Plath M, Bierbach D: Sex and the public-social eavesdropping, sperm competition risk, and male mate choice. Comm Integr Biol. 2011, 4: 1-5.
Plath M, Seggel U, Burmeister H, Heubel KU, Schlupp I: Choosy males from the underground: male mate choice in surface- and cave dwelling Atlantic mollies, Poecilia mexicana (Poeciliidae, Teleostei). Naturwissenschaften. 2006, 93: 103-109. 10.1007/s00114-005-0072-z.
Kelley JL, Magurran AE: Learned predator recognition and antipredator responses in fish. Fish Fisheries. 2003, 4: 216-226. 10.1046/j.1467-2979.2003.00126.x.
Magurran AE, Seghers BH: Population differences in predator recognition and attack cone avoidance in the guppy Poecilia reticulata. Anim Behav. 1990, 40: 443-452. 10.1016/S0003-3472(05)80524-X.
Kelley JL, Magurran AE: Effects of relaxed predation pressures on visual predator recognition in the guppy. Behav Ecol Sociobiol. 2003, 54: 225-232. 10.1007/s00265-003-0621-4.
Licht T: Discriminating between hungry and satiated predators: The response of guppies (Poecilia reticulata) from high and low predation sites. Ethology. 1989, 82: 238-243.
Seghers BH: Schooling behavior in the guppy (Poecilia reticulata)-evolutionary response to predation. Evolution. 1974, 28: 486-489. 10.2307/2407174.
Acknowledgements
Financial support came from the Deutsche Forschungsgemeinschaft (DFG; PL 470/3-1), the program Nachwuchswissenschaftler im Fokus of the University of Frankfurt (to MP), and from the Herrmann-Willkomm-Foundation (to SS and MP). The experiments reported here comply with the current laws of the Estados Unidos de México.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
All authors collected fish and jointly performed the experiments at DACBIOL-UJAT. DB, RR, MT, NH, MS and MP wrote the first draft of the manuscript; drawings are by MZ. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Bierbach, D., Schulte, M., Herrmann, N. et al. Predator-induced changes of female mating preferences: innate and experiential effects. BMC Evol Biol 11, 190 (2011). https://doi.org/10.1186/1471-2148-11-190
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2148-11-190