
Abstract
Background
Many cloned animals have been created by transfer of differentiated cells at G0/G1 or M phase of the cell cycle into enucleated M II oocytes having high maturation/meiosis/mitosis-promoting factor activity. Because maturation/meiosis/mitosis-promoting factor activity during oocyte maturation is maximal at both M I and M II, M I oocytes may reprogram differentiated cell nuclei as well. The present study was conducted to examine the developmental ability in vitro of porcine embryos reconstructed by transferring somatic cells (ear fibroblasts) into enucleated M I or M II oocytes.
Results
Analysis of the cell cycle stages revealed that 91.2 ± 0.2% of confluent cells were at the G0/G1 phase and 54.1 ± 4.4% of nocodazole-treated cells were at the G2/M phase, respectively. At 6 h after activation, nuclear swelling was observed in 50.0-88.9% and 34.4-39.5% of embryos reconstituted with confluent cells and nocodazole-treated cells regardless of the recipient oocytes, respectively. The incidence of both a swollen nucleus and polar body was low (6.3-10.5%) for all nocodazole-treated donor cell regardless of the recipient oocyte. When embryos reconstituted with confluent cells and M I oocytes were cultured, 2 (1.5%) blastocysts were obtained and this was significantly (P < 0.05) lower than that (7.6%) of embryos produced by transferring confluent cells into M II oocytes. No reconstructed embryos developed to the blastocyst stage when nocodazole-treated cells were used as donors.
Conclusions
Porcine M I oocytes have a potential to develop into blastocysts after nuclear transfer of somatic cells.
Similar content being viewed by others
Background
Many cloned animals have been created using M II oocytes as the recipient cytoplasm (sheep [1,2], cattle [3,4,5], mouse [6,7], goat [8] and pig [9,10,11]). Maturation/meiosis/mitosis-promoting factor (MPF) activity of recipient M II oocytes appears to be important for the reprogramming of nuclei of reconstructed embryos. Two distinct and different protocols emerge for embryo reconstruction by nuclear transfer when using M II oocytes as recipients. The first is the transfer of nuclei in G1, S or G2 phase into the preactivated recipients after reducing MPF activity and the second is the transfer of nuclei in GO or G1 phase directly into M II oocytes having high MPF activity [12]. Recently, it has been shown that when mouse embryonic stem cells [13] and bovine somatic cells [14] in the M phase were transferred into non-treated recipients, the chromosome constitution of reconstructed embryos was normal in that the second polar body was excluded after parthenogenetic activation and the embryos developed to offspring. The first protocol is effective for production of blastomeres derived cloned embryos because cell cycle synchronization of embryonic nuclei is difficult and the majority of blastomeres are in the S phase at any given time [15,16,17]. In the preactivated oocyte, the nuclear membrane of the donor cells remains intact due to the low activity of MPF and DNA synthesis occurs according to the original cell cycle stage at the time of nuclear transfer [12] and nuclear reprogramming occurs during the expansion of the donor nucleus [18,19]. Additionally, nuclei of cells of established embryonic cell lines are reprogrammed in preactivated recipient oocytes, porcine embryos reconstituted with blastocyst-derived cells and preactivated recipients have the ability to develop to blastocysts [20]. However, there is only one report in which cloned animals have been produced from embryos reconstructed by transferring differentiated cells into preactivated recipient oocytes [8] and the ability of preactivated recipient oocytes to reprogram differentiated cells is now in debate. When bovine somatic cells were transferred into preactivated recipient oocytes, the resulting reconstructed embryo development was limited, since all embryos arrested at the 8-cell stage regardless of the cell cycle of the donor cells [14]. Because embryonic genome activation of bovine embryos occurs between the 8- to 16-cell stages [21], preactivated recipients may not reprogram somatic nuclei. In contrast, when nuclei in G0 or G1 phase are transferred into non-treated recipients, reconstructed embryos can develop to offspring in many species [1,2,3,4,5,6,8,9,10,11]. In the M II oocyte, the membrane of the donor nucleus is broken down and the chromosomes are prematurely condensed due to the high activity of MPF. After parthenogenetic activation, the membrane reforms and DNA synthesis begins [22]. MPF activity during oocyte maturation is maximal at both M I and M II [12]. Because many studies suggest that exposure to high activity of MPF is effective for reprogramming a donor nucleus from a differentiated cell [1,2,3,4,5,6,7,8,9,10,11,14], M I oocytes may reprogram somatic cell nuclei as well. In the amphibian, the greatest yield and most advanced tadpoles came from differentiated somatic cell nuclei injected into M I oocytes [23,24] compared to M II oocytes [23,25,26], although adults could not be generated from these reconstructed embryos. However, there are no reports that experimentally tested this hypothesis in mammals. In the present study, we examined the ability of porcine M I oocytes to reprogram somatic cell nuclei.
Results
Cell Cycle Analysis of Donor Cells
The cell cycle stages of ear fibroblasts after reaching confluence or treatment with nocodazole were analyzed using flow cytometry. Analysis of the cell cycle stages in confluent cells revealed that 91.2 ± 0.2% of the cells were at the G0/G1 phase, 2.5 ± 0.1% were at the S phase and 6.4 ± 0.1% were at the G2/M phase. In the population of cells collected after nocodazole treatment, 31.0 ± 3.6% were at the G0/G1 phase, 14.8 ± 3.1% were at the S phase and 54.1 ± 4.4% were at the G2/M phase. Experiments were repeated 3 times and the data are expressed as mean ± SEM.
Behaviour before Activation of Donor Cell Nuclei
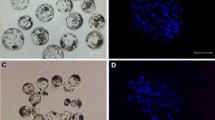
When confluent cells were transferred into M I oocytes, the fusion rate (60.2%) was significantly (P < 0.05) higher than that (25.8%) of embryos reconstituted with nocodazole-treated cells and M I oocytes (Table 1). At 15 h after fusion, spots of condensed chromatin (Figure 1A and 1B) were observed in the majority (85.0%) of embryos reconstructed by transferring confluent cells and this percentage was significantly (P < 0.05) higher than that (63.3%) of embryos reconstituted with nocodazole-treated cells. When nocodazole-treated cells were transferred into M I oocytes, one spot of condensed chromatin and one polar body (Figure 1C) was observed in 13.3% of reconstructed embryos. Regardless of the cell cycle of the donor cells, the percentage (3.3-5.0%) of embryos with swollen nuclei (Figure 1D and 1E) was low.

Nuclear swelling and polar body formation in porcine reconstructed embryos. Embryos containing one (A) or two (B) spots of condensed chromatin, one spot of condensed chromatin and one polar body (C), one (D) or two (E) swollen nuclei or one swollen nucleus and one polar body (F). Embryos were reconstructed by transferring confluent (A and D) or nocodazole-treated (B, C, E and F) cells into enucleated M I oocytes. Bar 100 μm.
Behaviour after Activation of Donor Cell Nuclei
As shown in Table 2, when confluent cells were used as donors, the fusion rate (60.2-62.7%) was significantly (P < 0.05) higher than that (29.0-33.6%) of nocodazole-treated cells regardless of the maturation stage of the recipient oocytes. At 6 h after activation, swollen nuclei were observed in 50.0 and 88.9% of embryos produced by transferring confluent cells into M I or M II oocytes with no significant differences. Although 34.4-39.5% of embryos reconstituted with nocodazole-treated cells had swollen nuclei, the percentage of embryos with both the swollen nucleus and polar body (Figure 1F) was only 6.3-10.5%.
Development of Reconstructed Embryos
The cleavage rate (38.2%) of embryos reconstituted with confluent cells and M II oocytes was significantly (P < 0.05) higher than that (5.3-9.7%) of embryos produced by transferring confluent or nocodazole-treated cells into M I oocytes (Table 3). However, this percentage (38.2%) did not significantly differ from that of embryos reconstituted with nocodazole-treated cells and M II oocytes (15.4%). When embryos reconstituted with confluent cells and M I oocytes were cultured, 2 (1.5%) blastocysts (Figure 2) were obtained and this percentage was significantly (P < 0.05) lower than that of embryos produced by transferring confluent cells into M II oocytes (7.6%). There were no significant differences in mean numbers of cells in the blastocysts between different maturation stages of the recipient oocytes. No embryos developed to the blastocyst stage when nocodazole-treated cells were used as donors regardless of the maturation stage of the recipient oocytes.

A porcine blastocyst developed from embryos reconstituted with confluent adult somatic cells and enucleated M I oocytes matured in a protein-free medium. Bar 100 μm.
Discussion
The results of the present study demonstrated the in vitro developmental ability to blastocysts of porcine embryos reconstituted with somatic cells and enucleated M I oocytes. To our knowledge, this is the first report that describes blastocyst formation of embryos reconstituted with M I oocytes in mammals.
When donor nuclei are transferred into non-treated M II recipient oocytes that have high MPF activity, various changes referred to as nuclear remodeling occur in the introduced nuclei. This phenomenon is considered to involve nuclear structural changes, such as nuclear envelope breakdown, premature chromosome condensation and pronuclear development and swelling [27,28,29]. Nuclear remodeling is thought to be a result of the exchange of proteins between the nuclear and cytoplasmic compartments [30] and is pre-requisite to or is part of the reprogramming of gene expression [31,32,33]. Therefore, it is generally considered that the phenomenon of nuclear swelling is one essential component of the more complex process of genetic reprogramming [34]. In the present study, when confluent or nocodazole-treated cells were transferred into enucleated M I oocytes, nuclear swelling was observed in 34.4-50.0% of reconstructed embryos after activation. This result suggests that porcine M I oocytes can initiate reprogramming of both confluent and nocodazole-treated cells. Moreover, some reconstructed embryos produced by transferring confluent cells developed to the blastocyst stage. Because activation of the embryonic genome occurs by the 4-cell stage in the pig [21], it is evident that donor nuclei were at least partially reprogrammed in these blastocysts. Therefore, our results clearly indicate that porcine M I oocytes like M II oocytes have the ability to reprogram confluent somatic cell nuclei as determined by nuclear remodeling and in vitro development.
The blastocyst formation rate of reconstructed embryos produced by transferring confluent cells into M I oocytes was significantly lower than that of embryos reconstituted with confluent cells and M II oocytes although there were no significant differences in percentages of embryos with swollen nuclei between different maturation stages of recipient oocytes. These results suggest that the reprogramming of donor nuclei may be started but is not completed in some embryos reconstituted with M I oocytes. It has been suggested that nuclear swelling may not necessarily be a sufficient indicator of developmental potential in a reconstructed embryo [28]. In addition to MPF, mitogen-activated protein (MAP) kinase is also known as an important protein kinase that regulates the meiotic cell cycle [35]. In mouse oocytes, MAP kinase plays a more important role in controlling chromatin and microtubule behavior than MPF [36]. However, MAP kinase and MPF activities do not vary between M I and M II of porcine oocytes because phosphorylations of MAP kinase and histone H1 kinase which corresponds to MPF activity reach the peak level at M I and remain high beyond M II [37,38]. Potentially, an unknown factor(s) is required for reprogramming of nuclei, which is present in M II oocytes but may not be present or active in M I oocytes. It is known that pronuclear exchange between zygotes does not prevent development of the reconstructed embryos [39] but transfer of donor nuclei from later developmental stages into enucleated zygotes brings about restricted development [40,41]. One explanation of this phenomenon is that a factor(s) required for reprogramming of donor nuclei is removed with the pronuclei from recipient zygotes. Similarly, the factor(s) would be present near the chromosomes at M I and removed with the M I plate during enucleation in many oocytes. In contrast, the factor(s) is believed to be in cytoplasts derived from M II oocytes because embryos reconstituted with donor nuclei from later developmental stages and enucleated M II oocytes can develop to adults [42]. An alternative cause is the effect of cytochalasin B, a microfilament polymerization inhibitor. In the present study, reconstructed embryos produced by transferring confluent cells into M I oocytes were cultured in the maturation medium supplemented with cytochalasin B to prevent loss of chromosomes by polar body formation. However, it has been reported recently that morphologically normal M I spindles are formed but further meiotic progression does not occur in porcine oocytes matured in a cytochalasin B-containing medium [38]. Therefore, it is possible that reprogramming of nuclei transferred into M I oocytes is inhibited by cytochalasin B. Thirdly, M I and M II oocytes may differ in cytoplasmic competence. When porcine M I oocytes are inseminated with spermatozoa in vitro, the formation rate of male and female pronuclei is significantly lower than that in M II oocytes [43]. Finally, optimal conditions for activation may vary between M I and M II oocytes. To our knowledge, there are no reports that describe activation of MI oocytes in mammals. Parameters of the activation pulse used in the present study were also optimized in a preliminary experiment using M II oocytes. It is possible that optimization of methods for activation improves the development of embryos reconstituted with M I oocytes.
In contrast, no reconstructed embryos developed to the blastocyst stage regardless of the maturation stage of the recipient oocytes when nocodazole-treated cells were transferred, although nuclear swelling was observed in 34.4-39.5% of the embryos. When cells in M phase are used as donors, reconstructed embryos with a polar body are diploid but embryos without a polar body are tetraploid [14]. In the present study, the percentage of embryos with both a swollen nucleus and a polar body was low after transfer of nocodazole-treated cells. In the bovine, the potential of reconstructed embryos with or without the polar body to develop to the blastocyst stage is not different [14]. However, this study suggests that porcine tetraploid embryos may not develop to blastocysts.
In the present study, there was a significant difference in fusion rates between the cell cycles of donor cells. The plasma membrane of nocodazole-treated cells was weak compared to that of confluent cells and many cells were broken before and after applying a fusion pulse. Parameters of the fusion pulse were optimized in a preliminary experiment using confluent cells. Therefore, it is expected that optimal conditions for fusion vary among cell cycles of donor cells in the pig.
In general, in vitro maturation (IVM) media used to prepare recipient oocytes for production of cloned embryos are usually supplemented with fetal calf serum (FCS) [3,4,5,14] or porcine follicular fluid (pFF) [11,20,44]. It is a well known fact that growth factors and hormones, in addition to many other factors, are abundant in serum [45,46] and follicular fluid [45,47,48]. Therefore, supplementation of IVM media with FCS or pFF presents the potential problems in proper quality control and repeatability among laboratories or replicates. Recently, a defined IVM system for porcine oocytes was developed in order to eliminate such variability [49,50]. In the present study, 7.6% of reconstructed embryos produced by transferring confluent adult somatic cells into enucleated M II oocytes matured in the protein-free medium developed to the blastocyst stage. The blastocyst formation rate in our study is similar to results from the recent reports in which porcine adult somatic (cumulus) cells (5-10%) [29,51] or fetal fibroblasts (1-16%) [11,29,51,52,53,54,55,56,57] were transferred into enucleated M II oocytes matured in media supplemented with pFF. In addition, quality of the blastocysts based on the mean number of cells (45.1 cells) equals if not exceeds results from previous studies (cumulus cells: 28.9 cells [51]; fetal fibroblasts: 12.5-66.0 cells [11,51,52,53,54,55,56]). Although this study did not compare defined IVM medium with complex medium the results indicate that protein-free IVM medium is suitable for maturing recipient oocytes for the production of cloned embryos. The defined conditions will allow us to examine factors affecting nuclear reprogramming of reconstructed embryos more precisely and better understand the basic molecular mechanisms of the process.
Conclusions
We have shown that porcine M I oocytes have a potential to develop into blastocysts after nuclear transfer of somatic cells. However, the process of nuclear reprogramming may be different between M I and M II oocytes because developmental abilities of reconstructed embryos vary between the maturation stages of recipient oocytes. Comparison of reprogramming events between M I and M II oocytes would bring about important information to understand the mechanisms of nuclear reprogramming in reconstructed embryos. The use of defined IVM medium will be useful for these studies. Taken together, the use of a defined medium and optimal reprogramming conditions such as the type of donor nuclei and recipient oocyte will lead to improvements in porcine cloning outcomes.
Materials and Methods
Donor Cells
Porcine fibroblasts were harvested from an ear skin biopsy obtained from an adult pig. The tissue was finely minced and digested in 0.125% (w/v) trypsin/0.02% (w/v) EDTA solution (Sigma Chemical Co., St. Louis, MO) containing 1 mg/ml collagenase (Sigma) and 0.3 mg/ml hyaluronidase (Sigma) for 2 h at 37°C. After digestion, the mixture was filtered through four layers of sterile gauze and cells were pelleted by centrifugation. Cells were cultured in DMEM/F-12 medium (Sigma) supplemented with 20% (v/v) FCS (Biowhittaker Inc., Walkersville, MD) under 5% CO2 in air at 37°C. After reaching confluence, cells were passaged. Passage 2 fibroblasts were trypsinized, suspended in the culture medium with 10% (v/v) dimethyl sulfoxide (Sigma) and stored as frozen aliquots. Donor cells were used for nuclear transfer between passages 3 and 9 of culture. For preparation of donors in G0/G1 phase, the cells were allowed to grow to confluency by culturing for 6 days and a single cell suspension was prepared by standard trypsinization. The cells were used for nuclear transfer within 5 days after reaching confluence. After 2-4 days of subculturing, the cells (approximately 50% of confluency) were cultured in the medium supplemented with 0.3 μg/ml nocodazole (Sigma), a microtubule polymerization inhibitor, for 10 h to induce G2/M phase arrest. After gentle pipetting, cells floating in the medium were collected. Both donor cells were prepared immediately prior to nuclear transfer.
Recipient Oocytes
Ovaries were collected from a local slaughterhouse and transported in 0.9% saline at approximately 32°C. Follicles greater than 3 mm in diameter were aspirated with an 18-gauge needle using vacuum suction (100 mmHg; 28 ml/min). Aspirated oocytes that had an evenly granulated cytoplasm and were surrounded by at least two uniform layers of compact cumulus cells were selected and washed three times in Hepes-buffered synthetic oviductal fluid medium [58]. Oocytes were transferred into tubes containing Hepes-buffered TCM-199 (Gibco BRL, Grand Island, NY) supplemented with 0.57 mM cysteine, 10 ng/ml epidermal growth factor (Sigma), 100 IU/ml penicillin, 100 μg/ml streptomycin, 0.25 ng/ml amphotericin, 0.01 IU/ml porcine FSH (Sioux Biochemicals, Sioux Center, IA), 0.01 IU/ml porcine LH (Sioux Biochemicals) and 0.1% (w:v) polyvinyl alcohol (maturation medium) [50], and shipped to the laboratory overnight (20-24 h) at 38.5°C. Oocytes continued to be cultured in the tubes under 38.5°C until enucleation.
Nuclear Transfer
Reconstructed embryos were produced using a modification of the method described by Miyoshi et al. [44]. Cumulus cells were removed by vortexing with 0.1% (w:v) hyaluronidase and denuded oocytes were transferred into 100 μl of Hepes-buffered TCM-199 supplemented with 7.5 μg/ml cytochalasin B (Sigma) and 10% FCS with the osmolarity adjusted to 300 mOsm by adding sorbitol (manipulation medium) at 29 or 42 h after IVM. The M I plate or the first polar body and M II plate were removed by aspiration with a 15-μm inner diameter glass pipette. The oocytes had been previously stained in the manipulation medium supplemented with 5 μg/ml Hoechst 33342 (Sigma) for 20 min and confirmation of successful enucleation was achieved by visualizing the cytoplast and removed cytoplasm under ultraviolet light. After enucleation, a donor cell was inserted into the perivitelline space of each enucleated oocyte using the same glass pipette. When nocodazole-treated cells were used as donors, large cells (cell diameter was >15 μm) were selected. In our preliminary experiment, such a relatively large cytoplasmic volume was typical of cells at the G2/M phase as described by Wakayama et al. [13]. Cell-oocyte complexes were washed in TCM-199 supplemented with 10% FCS, transferred to the same medium and kept under 5% CO2 in air at 38.5°C until fusion.
Fusion was performed in a 100 mm dish filled with 15 ml of Zimmermann fusion medium [59]. Two stainless steel wires (100 μm diameter) attached to micromanipulators were used as electrodes. The single cell-oocyte complex was sandwiched between the electrodes and oriented with the contact surface between the cytoplast and the donor cell perpendicular to the electrodes. The distance between the electrodes was about 100 μm. Membrane fusion was induced by applying a single direct current pulse of 250 V/mm for a duration of 20 μsec with a prepulse of alternating current field of 5 V, 1 MHz for 2 sec using an LF 101 Fusion Machine (TR Tech Co., Tokyo, Japan). Following fusion, the complexes were washed in G 1.2 medium [60] and cultured for a period of 1 h in 100 μl of the same medium. Fusion rate was then determined by microscopic examination.
Activation of Reconstructed Embryos
Fused embryos were washed once in Zimmermann fusion medium and then placed between 2 wire electrodes (1 mm apart) of the fusion chamber slide with 15 ml of the fusion medium. Direct current pulses of 75 V/mm were applied twice to the embryos for a duration of 60 μsec at intervals of 5 sec.
Examination of Reconstructed Embryos
Before and after activation, fused embryos were stained with 5 μg/ml Hoechst 33342 for 20 min and then examined for nuclear swelling and polar body formation under ultraviolet light. The nucleus with a diameter that was at least twice bigger than the original size of donor cell nuclei was considered to be a swollen nucleus.
Experimental Studies
Cell Cycle Analysis of Donor Cells
In Experiment 1, the cell cycle stages of confluent and nocodazole-treated donor cells were analyzed using flow cytometry. Collected cells (approx. 2 × 106 cells) were centrifuged and the pellet was re-suspended in 1 ml of cold PBS. Then, cells were fixed by adding 4 ml of -20°C absolute ethanol and stored in -20°C. Before analysis, cells were centrifuged and the pellet was re-suspended in 1 ml of PBS. Then, 100 μl of DNase-free RNase A (200 μg/ml; Sigma) was added and samples were incubated for 30 min at 37°C. After incubation, 100 μl of propidium iodide (1 mg/ml; Sigma) was added into each sample and cells were analyzed using FACStartPLUS analyzer (Becton Dickinson, San Jose, CA).
Behavior before Activation of Donor Cell Nuclei
In Experiment 2, we examined the behavior before activation of donor cell nuclei transferred into enucleated M I oocytes. M I oocytes were enucleated at 29 h after IVM and confluent or nocodazole-treated cells were transferred. Fused embryos were cultured in the maturation medium for 15 h and then examined for nuclear swelling and polar body formation. When confluent cells were used as donors, fused embryos were cultured in the medium supplemented with 7.5 μg/ml cytochalasin B to prevent extrusion of polar bodies.
Behavior after Activation of Donor Cell Nuclei
In Experiment 3, the behavior of nuclei of confluent or nocodazole-treated cells transferred into enucleated M I or M II oocytes was examined after activation. Embryos reconstituted with M I oocytes were cultured in the maturation medium with or without cytochalasin B for 15 h and then activated. M II oocytes were enucleated at 42 h after IVM and embryos reconstituted with them were activated at 3 h after fusion. Activated embryos were cultured in 100 μl of G 1.2 medium for 6 h and then examined for nuclear swelling and polar body formation. When confluent cells were used as donors, embryos were cultured in G1.2 medium supplemented with 7.5 μg/ml cytochalasin B for 2 h after activation to prevent extrusion of polar bodies [44].
Development of Reconstructed Embryos
Experiment 4 was undertaken to compare in vitro developmental abilities of reconstructed embryos produced by transferring confluent or nocodazole-treated cells into enucleated M I or M II oocytes as described above. After culture in G1.2 medium for 3 days, the embryos were transferred into 100 μl of G2.2 medium [60] and continued to be cultured. The embryos were examined for cleavage and blastocyst formation at 2 and 7 days after culture, respectively. At the end of the culture period, the number of cells in blastocysts was examined. The blastocysts were placed on slides with a drop of mounting medium consisting of glycerol and PBS (9:1) containing 0.1 mg/ml Hoechst 33342. A cover slip was placed on the top of the blastocysts and the edge was sealed with fingernail polish. The numbers of nuclei were counted under ultraviolet light.
Statistical Analysis
All percentage data were subjected to an arcsin square root transformation in each replicate and the transformed values were analyzed using one-way (Experiment 2) or two-way (Experiments 3 and 4) ANOVA followed by Student-Newman-Keuls test. The numbers of cells in blastocysts were analyzed using Student t-test. A probability of P < 0.05 was considered statistically significant.
References
Campbell KHS, McWhir J, Ritchie WA, Wilmut I: Sheep cloned by nuclear transfer from a cultured cell line. Nature. 1996, 380: 64-66. 10.1038/380064a0.
Wilmut I, Schnieke AE, McWhir J, Kind AJ, Campbell KHS: Viable offspring derived from fetal and adult mammalian cells. Nature. 1997, 385: 810-813. 10.1038/385810a0.
Cibelli JB, Stice SL, Golueke PJ, Kane JJ, Jerry J, Blackwell C, Abel Ponce de Leon, Robl JM: Cloned transgenic calves produced from nonquiescent fetal fibroblasts. Science. 1998, 280: 1256-1258. 10.1126/science.280.5367.1256.
Kato Y, Tani T, Sotomaru Y, Kurokawa K, Kato J, Doguchi H, Yasue H, Tsunoda Y: Eight calves cloned from somatic cells of a single adult. Science. 1998, 282: 2095-2098. 10.1126/science.282.5396.2095.
Wells DN, Misica PM, Tervit HR: Production of cloned calves following nuclear transfer with cultured adult mural granulosa cells. Biol Reprod. 1999, 60: 996-1005.
Wakayama T, Perry ACF, Zuccotti M, Johnson KR, Yanagimachi R: Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature. 1998, 394: 369-374. 10.1038/28615.
Wakayama T, Yanagimachi R: Cloning of male mice from adult tail-tip cells. Nature genet. 1999, 22: 127-128. 10.1038/9632.
Baguisi A, Behboodi E, Melican DT, Pollock JS, Destrempes MM, Cammuso C, Williams JL, Nims SD, Porter CA, Midura P, Palacios MJ, Ayres SL, Denniston RS, Hayes ML, Ziomek CA, Meade HM, Godke RA, Gavin WG, Overstrom EW, Echelard Y: Production of goats by somatic cell nuclear transfer. Nature Biotech. 1999, 17: 456-461. 10.1038/8632.
Onishi A, Iwamoto M, Akita T, Mikawa S, Takeda K, Awata T, Hanada H, Perry ACF: Pig cloning by microinjection of fetal fibroblast nuclei. Science. 2000, 289: 1188-1190. 10.1126/science.289.5482.1188.
Polejaeva IA, Chen SH, Vaught TD, Page RL, Mullins J, Ball S, Dai Y, Boone J, Walker S, Ayares DL, Colman A, Campbell KHS: Cloned pigs produced by nuclear transfer from adult somatic cells. Nature. 2000, 407: 86-90. 10.1038/35024082.
Betthauser J, Forsberg E, Augenstein M, Childs L, Eilertsen K, Enos J, Forsythe T, Golueke P, Jurgella G, Koppang R, Lesmeister T, Mallon K, Mell G, Misica P, Pace M, Pfister-Genskow M, Strelchenko N, Voelker G, Watt S, Thompson S, Bishop M: Production of cloned pigs from in vitro systems. Nature Biotech. 2000, 18: 1055-1059. 10.1038/80242.
Campbell KHS, Loi P, Otaegui P, Wilmut I: Cell cycle co-ordination in embryo cloning by nuclear transfer. Rev Reprod. 1996, 1: 40-45. 10.1530/ror.0.0010040.
Wakayama T, Rodriguez I, Perry ACF, Yanagimachi R, Mombaerts P: Mice cloned from embryonic stem cells. Proc Natl Acad Sci USA. 1999, 96: 14984-14989. 10.1073/pnas.96.26.14984.
Tani T, Kato Y, Tsunoda Y: Direct exposure of chromosomes to nonactivated ovum cytoplasm is effective for bovine somatic cell nucleus reprogramming. Biol Reprod. 2001, 64: 324-330.
Campbell KHS, Loi P, Cappai P, Wilmut I: Improved development to blastocyst of ovine nuclear transfer embryos reconstructed during the presumptive S-phase of enucleated activated oocytes. Biol Reprod. 1994, 50: 1358-1359.
Kono T, Sotomaru Y, Aono F, Takahashi T, Ogiwara I, Sekizawa F, Arai T, Nakahara T: Effect of ooplast activation on the development of oocytes following nucleus transfer in cattle. Theriogenohgy. 1994, 41: 1463-1471. 10.1016/0093-691X(94)90197-Q.
Nagashima H, Ashman RJ, Nottle MB: Nuclear transfer of porcine embryos using cryopreserved delipated blastomeres as donor nuclei. Mol Reprod Dev. 1997, 48: 339-343. 10.1002/(SICI)1098-2795(199711)48:3<339::AID-MRD6>3.0.CO;2-S.
Stice SL, Robl JM: Nuclear reprogramming in nuclear transplant rabbit embryos. Biol Reprod. 1988, 39: 657-664.
Stice SL, Keefer CL, Matthews L: Bovine nuclear transfer embryos: oocyte activation prior to blastomere fusion. Mol Reprod Dev. 1994, 38: 61-68.
Miyoshi K, Taguchi Y, Sendai Y, Hoshi H, Sato E: Establishment of a porcine cell line from in vitro-produced blastocysts and transfer of the cells into enucleated oocytes. Biol Reprod. 2000, 62: 1640-1646.
Telford NA, Watson AJ, Schultz GA: Transition from maternal to embryonic control in early mammalian development: a comparison of several species. Mol Reprod Dev. 1990, 26: 90-100.
Campbell KHS, Ritchie WA, Wilmut I: Nuclear-cytoplasmic interactions during the first cell cycle of nuclear transfer reconstructed bovine embryos: implications for deoxyribonucleic acid replication and development. Biol Reprod. 1993, 49: 933-942.
DiBerardino MA, Hoffner NJ: Gene reactivation in erythrocytes. Nuclear transplantation in oocytes and eggs of Rana. Science. 1983, 219: 862-864.
DiBerardino MA, Orr Hoffner N, McKinnell RG: Feeding tadpoles cloned from Rana erythrocyte nuclei. Proc Natl Acad Sci USA. 1986, 83: 8231-8234.
Gurdon JB, Laskey RA, Reeves OR: The developmental capacity of nuclei transplanted from keratinized skin cells of adult frogs. J Embryol Exp Morphol. 1975, 34: 93-112.
Brun RB: Developmental capacities of Xenopus eggs, provided with erythrocyte or erythroblast nuclei from adults. Dev Biol. 1978, 65: 271-284.
Tsunoda Y, Tokunaga T, Uchida T: Nuclear transplantation of male germ cells in the mouse. Development. 1989, 107: 407-411.
Prather RS, Sims MM, First NL: Nuclear transplantation in the pig embryo: nuclear swelling. J Exp Zool. 1990, 255: 355-358.
Uhm SJ, Chung HM, Kirn C, Shim H, Kirn NH, Lee HT, Chung KS: In vitro development of porcine enucleated oocytes reconstructed by the transfer of porcine fetal fibroblasts and cumulus cells. Theriogenology. 2000, 54: 559-570. 10.1016/S0093-691X(00)00371-X.
Prather RS, First NL: Cloning embryos by nuclear transfer. J Reprod Fertil Suppl. 1990, 41: 125-134.
Collas P, Robl JM: Relationship between nuclear remodeling and development in nuclear transplant rabbit embryos. Biol Reprod. 1991, 45: 455-465.
Hyttel P, Prochazka R, Smith S, Kanka J, Greve T: RNA synthesis in porcine blastomere nuclei introduced into in vitro matured ooplasm. Acta Vet Scand. 1993, 34: 159-167.
Niemann H, Reichelt B: Manipulating early pig embryos. J Reprod Fertil Suppl. 1993, 48: 75-94.
Liu L, Moor RM, Laurie S, Notarianni E: Nuclear remodelling and early development in cryopreserved, porcine primordial germ cells following nuclear transfer into in vitro-matured oocytes. Int J Dev Biol. 1995, 39: 639-644.
Sun QY, Breitbart H, Schatten H: Role of the MAPK cascade in mammalian germ cells. Reprod Fertil Dev. 1999, 11: 443-450. 10.1071/RD00014.
Verlhac MH, Kubiak JZ, Clarke HJ, Maro BH: Microtubule and chromatin behavior follow MAP kinase activity but not MPF activity during meiosis in mouse oocyte. Development. 1994, 120: 1017-1025.
Inoue M, Naito K, Aoki F, Toyoda Y, Sato E: Activation of mitogen-activated protein kinase during meiotic maturation in porcine oocytes. Zygote. 1995, 3: 265-271.
Sun QY, Lai L, Park KW, Kuhholzer B, Prather RS, Schatten H: Dynamic events are differently mediated by microfilaments, microtubules, and mitogen-activated protein kinase during porcine oocyte maturation and fertilization in vitro. Biol Reprod. 2001, 64: 879-889.
McGrath J, Solter D: Nuclear transplantation in the mouse embryo by microsurgery and cell fusion. Science. 1998, 220: 1300-1302.
McGrath J, Solter D: Nuclear transplantation in mouse embryos. J Exp Zool. 1993, 228: 355-362.
Wakayama T, Tateno H, Mombaerts P, Yanagimachi R: Nuclear transfer into mouse zygotes. Nature Genet. 2000, 24: 108-109. 10.1038/72749.
Willadsen SM: Nuclear transplantation in sheep embryos. Nature. 1986, 320: 63-65.
Wang WH, Abeydeera LR, Okuda K, Niwa K: Penetration of porcine oocytes during maturation in vitro by cryopreserved, ejaculated spermatozoa. Biol Reprod. 1994, 50: 510-515.
Miyoshi K, Saeki K, Sato E: Improvement in development of porcine embryos reconstituted with cells from blastocyst-derived cell lines and enucleated oocytes by optimization of reconstruction methods. Cloning. 2000, 2: 175-184. 10.1089/15204550045473510.1089/152045500454735.
Hsu CJ, Holmes SD, Hammond JM: Ovarian epidermal growth factor-like activity. Concentrations in porcine follicular fluid during follicular enlargement. Biochem Biophys Res Commun. 1987, 147: 242-247.
Younis AI, Brackett BG, Fayrer-Hosken RA: Influence of serum and hormones on bovine oocyte maturation and fertilization in vitro. Gamete Res. 1989, 23: 189-201.
Ainsworth L, Tsang BK, Downey BR, Marcus GJ, Armstrong DT: Interrelationships between follicular fluid steroid levels, gonadotropic stimuli, and oocyte maturation during preovulatory development of porcine follicles. Biol Reprod. 1980, 23: 621-627.
Ding J, Foxcroft GR: Follicular heterogeneity and oocyte maturation in vitro in pigs. Biol Reprod. 1992, 47: 648-655.
Abeydeera LR, Wang WH, Prather RS, Day BN: Maturation in vitro of pig oocytes in protein-free culture media: fertilization and subsequent embryo development in vitro. Biol Reprod. 1998, 58: 1316-1320.
Abeydeera LR, Wang WH, Cantley TC, Rieke A, Murphy CN, Prather RS, Day BN: Development and viability of pig oocytes matured in a protein-free medium containing epidermal growth factor. Theriogenology. 2000, 54: 787-797. 10.1016/S0093-691X(00)00390-3.
Koo DB, Kang YK, Choi YH, Park JS, Kirn HN, Kim T, Lee KK, Han YM: Developmental potential and transgene expression of porcine nuclear transfer embryos using somatic cells. Mol Reprod Dev. 2001, 58: 15-21. 10.1002/1098-2795(200101)58:1<15::AID-MRD3>3.0.CO;2-Y.
Tao T, Boquest AC, Machaty Z, Petersen AL, Day BN, Prather RS: Development of pig embryos by nuclear transfer of cultured fibroblast cells. Cloning. 1999, 1: 55-62. 10.1089/1520455995002009410.1089/15204559950020094.
Tao T, Machaty Z, Boquest AC, Day BN, Prather RS: Development of pig embryos reconstructed by microinjection of cultured fetal fibroblast cells into in vitro matured oocytes. Anim Reprod Sci. 1999, 56: 133-141. 10.1016/S0378-4320(99)00037-8.
Koo DB, Kang YK, Choi YH, Park JS, Han SK, Park IY, Kim SU, Lee KK, Son DS, Chang WK, Han YM: In vitro development of reconstructed porcine oocytes after somatic cell nuclear transfer. Biol Reprod. 2000, 63: 986-992.
Kuhholzer B, Tao T, Machaty Z, Hawley RJ, Greenstein JL, Day BN, Prather RS: Production of transgenic porcine blastocysts by nuclear transfer. Mol Reprod Dev. 2000, 56: 145-148. 10.1002/(SICI)1098-2795(200006)56:2<145::AID-MRD4>3.0.CO;2-8.
Verma PJ, Du ZT, Crocker L, Faast R, Grupen CG, McIlfatrick SM, Ashman RJ, Lyons IG, Nottle MB: In vitro development of porcine nuclear transfer embryos constructed using fetal fibroblasts. Mol Reprod Dev. 2000, 57: 262-269. 10.1002/1098-2795(200011)57:3<262::AID-MRD8>3.0.CO;2-X.
Uhm SJ, Kim NH, Kim T, Chung HM, Chung KH, Lee HT, Chung KS: Expression of enhanced green fluorescent protein (EGFP) and neomycin resistant (Neo(R)) genes in porcine embryos following nuclear transfer with porcine fetal fibroblasts transfected by retrovirus vector. Mol Reprod Dev. 2000, 57: 331-337. 10.1002/1098-2795(200012)57:4<331::AID-MRD4>3.0.CO;2-7.
Tervit HR, Whittingham DG, Rowson LEA: Successful culture in vitro of sheep and cattle ova. J Reprod Ferttil. 1972, 30: 493-497.
Zimmemiann U, Vienken J: Electric field-induced cell-to-cell fusion. J Membr Biol. 1982, 67: 165-182.
Gandhi AP, Lane M, Gardner DK, Krisher RL: Substrate utilization in porcine embryos cultured in NCSU23 and G1.2/G2.2 sequential culture media. Mol Reprod Dev. 2001, 58: 269-275. 10.1002/1098-2795(200103)58:3<269::AID-MRD4>3.0.CO;2-L.
Acknowledgments
The authors thank Dr. DK Gardner, Colorado Center for Reproductive Medicine for providing G1.2 and G2.2 media. We also thank RA Waltenburg, MC Tumlin and DS Respess for assisting with nuclear transfer and SK Bridges for transporting oocytes. Supported by NIH SBIR grant (No. R43HL65806-01). KM is a recipient of the Japan Society for the Promotion of Science Fellowship (No. 9801517).
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Miyoshi, K., Rzucidlo, S.J., Gibbons, J.R. et al. Development of porcine embryos reconstituted with somatic cells and enucleated metaphase I and II oocytes matured in a protein-free medium. BMC Dev Biol 1, 12 (2001). https://doi.org/10.1186/1471-213X-1-12
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-213X-1-12