
Summary
Background
α1-antitrypsin (AAT) is one of the major serine proteinase inhibitors controlling proteinases in many biological pathways. There is increasing evidence that AAT is able to exert other than antiproteolytic effects. To further examine this question we compared how various doses of the native (inhibitory) and the polymerised (non-inhibitory) molecular form of AAT affect pro-inflammatory responses in human monocytes, in vitro. Human monocytes isolated from different donors were exposed to the native or polymerised form of AAT at concentrations of 0.01, 0.02, 0.05, 0.1, 0.5 and 1 mg/ml for 18 h, and analysed to determine the release of cytokines and to detect the activity of NF-κB.
Results
We found that native and polymerised AAT at lower concentrations, such as 0.1 mg/ml, enhance expression of TNFα (10.9- and 4.8-fold, p < 0.001), IL-6 (22.8- and 23.4-fold, p < 0.001), IL-8 (2.4- and 5.5-fold, p < 0.001) and MCP-1 (8.3- and 7.7-fold, p < 0.001), respectively, compared to buffer exposed cells or cells treated with higher doses of AAT (0.5 and 1 mg/ml). In parallel to increased cytokine levels, low concentrations of either conformation of AAT (0.02–0.1 mg/ml) induced NF-κB p50 activation, while 1 mg/ml of either conformation of AAT suppressed the activity of NF-κB, compared to controls.
Conclusions
The observations reported here provide further support for a central role of AAT in inflammation, both as a regulator of proteinase activity, and as a signalling molecule for the expression of pro-inflammatory molecules. This latter role is dependent on the concentration of AAT, rather than on its proteinase inhibitory activity.
Similar content being viewed by others


Background
α1-antitrypsin (AAT) is one of the major serine proteinase inhibitors in human plasma, synthesised primarily in the liver, but also in extra-hepatic tissues and cells, including neutrophils, monocytes and alveolar macrophages [1–4]. Under normal conditions the daily production of AAT is 34 mg per kg body weight. The average concentration of AAT in plasma in healthy individuals is estimated to be 1.3 mg/ml, with a half-life of 3 to 5 days [5]. The concentration of AAT during acute phase processes rises by three- to fourfold above normal while, for example, local levels of AAT are shown to increase up to 11-fold [4]. The concentration of AAT in plasma also increases during oral contraceptive therapy and pregnancy [6]. A large number of studies has proven the importance of this rapid and high magnitude increase in AAT concentrations for the local regulation of serine proteinase activity and tissue protection against proteolytic destruction [7, 8]. AAT/proteinase complex formation and inhibitor inactivation presumably act as signals for inhibitor production and turnover in the acute phase state [9]. Human neutrophils, monocytes and alveolar macrophages can increase the expression of AAT in response to inflammatory mediators, such as IL-6 and endotoxins [1, 7].
The distribution of AAT in tissue is not uniform, for example, it is reduced to approximately 10% of the plasma levels in the fluid of the lower respiratory tract [10], which explains the extremely low levels of the protein in alveolar fluid of individuals with inherited AAT deficiency. AAT also diffuses through endothelial and epithelial cell walls and is shown to be present in the epithelial lining fluid at 10 to 15% of total serum AAT concentration [11]. Moreover, by using a monoclonal antibody that specifically recognizes AAT polymers, we demonstrated that vascular, endothelium-bound AAT is in a polymeric form [12].
A change of a single amino acid in certain domains of the AAT molecule can block changes in the structure necessary for the protein folding and normal inhibitory activity, and can lead to the polymerization of mutant serpin [13]. Severe AAT deficiency of the homozygous PiZ phenotype, which differs from the normal M variant in the substitution of Glu-342 by Lys, is recognized as a hereditary condition predisposing to disease on the basis of low plasma levels (10% of normal) of the protein arising not from the lack of AAT synthesis, but from a blockage of its secretion [14]. Polymerisation of AAT is known to be involved in AAT deficiency-related diseases, such as liver cirrhosis, neonatal hepatitis and hepatocellular carcinoma, which are characterised by the formation of intracellular inclusions of polymerised AAT [15–17]. Recent studies provide evidence that AAT polymers are also present in the lungs of emphysema patients with Z-AAT deficiency and in the circulation of PiZ AAT carriers [12, 18]. The normal M variant of AAT also forms polymers in vitro, and the process of polymer formation is shown to be dependent on temperature, pH and protein concentration [19].
Individuals with homozygous PiZ AAT deficiency have not only low baseline serum levels of AAT, but also an attenuated acute-phase response [20]. They are particularly susceptible to lung damage during bacterial exacerbations when there is a significant inflammatory cell influx and a release of various proteases. Normally, all of these proteolytic enzymes are counteracted by anti-proteinases, such as AAT, in amounts sufficient to prevent lung injury. Therefore, it is widely accepted that inherited AAT deficiency results in proteinase-antiproteinase imbalance and that this is the proximal cause of the observed pathologies. On the other hand, while low levels of native AAT may result in uncontrolled proteolytic activity, dysfunctional forms of AAT, such as the polymerised, may have biological activities that differ from those of native AAT at physiological concentrations.
Various observations indicate that the native inhibitory form of AAT exerts its effects not only by inhibiting neutrophil elastase and other leukocyte derived proteinases. AAT was found to stimulate fibroblast proliferation and procollagen synthesis [21], up-regulate human B cell differentiation into IgE-and IgG4-secreting cells [22], interact with the proteolytic cascade of enzymes involved in apoptosis [23] and to express contrasting effects on the post-transcriptional regulation of iron between erytroid and monocytic cells [24]. In addition, AAT can inhibit neutrophil superoxide production [25], induce macrophage-derived interleukin-1 receptor antagonist release [26], and reduce TNFα-induced lethality [27].
Together these findings lead to an idea that the development of certain diseases might not only be ascribed to deficiency of AAT inhibitory activity, but also other properties of this protein, realised under inflammatory conditions. To address this possibility, we have tested whether native and polymerised forms of AAT differ in vitro in their effects on primary human monocytes, dependent on the protein concentration used. We find that both the native (inhibitory) and polymerised (non-inhibitory) form of AAT have similar effects as monocyte activators, with pro-inflammatory stimulation in low doses, and attenuation of pro-inflammatory activities at physiologically normal doses.
Results and discussion
AAT has been reported to influence cell function and behaviour via both direct and indirect mechanisms dependent on its concentration [28, 29]. For example, native AAT has been shown to stimulate and inhibit cell proliferation, depending on cell type and the AAT concentration examined [29]. Similarly, earlier studies by Aoshiba and colleagues have shown that AAT has stimulatory and inhibitory effects on blood polymorphonuclear cells migration depending on its concentration [30]. AAT impaired chemotactic responsiveness at 2 and 10 mg/ml, but induced chemokinesis and chemotaxis with maximum potency at 0.2 mg/ml [30]. In addition, conformationally modified molecular forms of AAT, such as proteolytically cleaved, complexed with elastase or polymerised, were shown to act as potent activators and chemoattractants for neutrophils [31–33] and were therefore suggested to play a role in sustaining inflammatory reactions at the site of tissue injury.
In order to further investigate if AAT can express pro-and/or anti-inflammatory effects in a dose-and conformation-dependent fashion we designed a model in which primary human monocytes were exposed to various concentrations of the native or the polymerised form of AAT for 18 h. Samples of native and polymerised AAT were subjected to native 7.5% PAGE analysis. Fig. 1, lane 1, shows a single molecular weight band corresponding to native AAT. Heating of native AAT for 3 h at 60°C resulted in the generation of different size polymers of AAT (Fig. 1, lane 2). As illustrated in Figures 2 and 3, monocytes exposed to native or polymerised preparation of the AAT at lower concentration (0.1 mg/ml) increase release of TNFα (10.9- and 4.8-fold, p < 0.001), IL-6 (22.8- and 23.4-fold, p < 0.001), IL-8 (2.4- and 5.5-fold, p < 0.001) and MCP-1 (8.3- and 7.7-fold, p < 0.001), respectively, compared to buffer exposed cells or cells treated with higher doses of either conformation of AAT (0.5 and 1 mg/ml). Thus, our data provide evidence that native and polymerised AAT at lower concentrations, i.e. 0.1 mg/ml and below, are sufficient to elicit an increase in pro-inflammatory cytokine (IL-6 and TNF-α) and chemokine (IL-8 and MCP-1) release from monocytes in vitro, whereas the cells treated with higher doses of either AAT manifest no activation at all. Although low-dose AAT induced cytokine and chemokine release is less pronounced relative to LPS (10 ng/ml) by approximately 10- to 100-fold (data not shown), it is still important to point out that the observed AAT effects are reproducible and statistically significant. In addition, statistical comparison between relative effects of native versus polymerised AAT (0.1 mg/ml) showed that native AAT is significantly more potent stimulator of TNFα and MCP-1 release, compared to polymerised AAT whereas polymerised AAT expresses stronger effects on IL-6 and IL-8 release (Table 1). These findings may have biological importance and need to be further investigated.

Native and polymerised AAT (10 μg/ml) analysed by 7.5% native PAGE. Lane M, molecular size markers, lane 1, native AAT and lane 2, polymerised AAT (incubated at 60°C, 3 h).

Cytokines released from monocytes stimulated with various concentrations of native (A) or polymerized (B) form of AAT. Each point represents the mean ± SD of six separate experiments. Significantly high up-regulation of the pro-inflammatory cytokines (TNF-α and IL-6) was found when cells were treated with low doses (0.05 and 0.1 mg/ml) of native or polymerized AAT, respectively, compared to control treated with equivalent amounts of buffer alone. The buffer control levels are indicated on the graphs (TNFα in blue and IL-6 in red).

Chemokines released from monocytes stimulated with various concentrations of native (A) or polymerized (B) form of AAT. Each point represents the mean ± SD of six separate experiments. Significantly high up-regulation of the pro-inflammatory chemokines (IL-8 and MCP-1) was found when cells were treated with low doses (0.05 and 0.1 mg/ml) of native or polymerized AAT, respectively, compared to controls treated with equivalent amounts of buffer alone. The buffer control levels are indicated on the graphs (MCP-1 in blue and IL-8 in red).
It is known that nuclear transcription factor-κB (NF-κB) regulates host inflammatory and immune responses and cellular growth properties by increasing the expression of specific cellular genes [34]. Several studies have demonstrated that in monocytes, NF-κB activation results in the transcription of immediate early genes that encode IL-6, TNFα, MCP-1, as well as several other molecules [35]. Numerous stimuli are known to activate NF-κB, including the bacterial component LPS and other pro-inflammatory factors such as cytokines [36, 37].
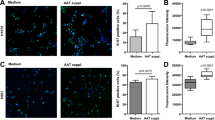
We have also studied the effects of native and polymerised AAT on the expression of transcription factor NF-κB. Monocytes exposed to native or polymerised form of AAT at lower concentrations, such as 0.05 and 0.1 mg/ml activate NF-κB expression, compared to control, buffer-stimulated cells (Figure 4), while the serum physiological level of either AAT (1 mg/ml) down-regulates NF-κB expression to almost undetectable level. These findings correlate with the stimulatory effects of low dose of AAT on pro-inflammatory cytokine and chemokine release in monocytes described above (Figures 2 and 3) confirming the interrelationship between transcription factor, NF-κB, activation and pro-inflammatory cytokine release in response to native and polymerised AAT.

A and B. Monitoring of NFκB p50 activation in monocytes after 18 h of exposure to different concentrations of native or polymerized AAT by ELISA-based TransAM kitAM. Cell nuclear extracts were prepared from monocytes alone or treated with different concentrations of AAT. NFκB activity assay was performed in 96-well plates with 10 μg of cell extract per well. Absorbance was measured by spectrophotometry at 450 nm. Specificity controls were performed by adding a molar excess (20 pmol/well) of mutant NFκB oligonucleotide (the positive signal remained unaffected) and wild-type NFκB oligonucleotide (a signal was abolished). A and B represent results from two independent experiments. Each point represents the mean of two experiments.
Surprisingly, the effects of both native and polymerised AAT on monocyte activity were parallel for all parameters measured and at all doses, although relative effects on specific cytokine or chemokine secretion were found to be different (Table 1). Equally important was the observation that the cellular responses showed a marked inverse dependence on dose, and were not dependent on the conformational form of AAT. Low AAT concentrations in vivo appear to be related to a state of general AAT deficiency, but may also reflect local depletion at different stages of inflammation. This latter may be the evolutionary basis for the inverse dependence on AAT concentration of the expression of pro-inflammatory molecules. The production of cytokines, chemokines, cell surface receptor/adhesion proteins, and other molecules by activated monocytes, macrophages and neutrophils can serve to orchestrate the immune response and is essential to host defence. The increased expression of a defined set of cytokines, such as IL-6 and TNFα, is believed to be one of the major pathological mediators of inflammatory diseases. Chemotactic cytokines (chemokines) are also generated locally, and this stimulates monocytes to migrate into the site of inflammation. In particular, IL-8 and MCP-1 have been implicated in diseases characterised by monocyte-rich infiltrates because of its ability to induce monocyte trafficking and activation [38]. IL-6, TNFα, IL-8 and MCP-1 are all hallmarks of inflammation and sustain the multiple inflammation-associated processes of lymphocyte attraction, differentiation, immunoglobulin secretion, primary monocyte recruitment, neutrophil attraction and up-regulation of extracellular matrix-degrading properties [39]. They are all most likely regulated by genes controlled from the NF-κB promoter, since their expression correlates strongly with NF-κB activation in our monocyte stimulation model. Based on our results, it is very likely that at low concentrations native as well as polymerised AAT may contribute to inflammatory events in vivo.
The inverse dependence on AAT concentration observed for the expression of pro-inflammatory reactance suggests a possible receptor-dependent feedback loop in which AAT is the key ligand. The cellular receptor for AAT has not been identified to date, although the cell surface receptors, termed serpin-enzyme complex (SEC) receptor [9], low density lipoprotein related (LRP) receptor [40], low density lipoprotein (LDL) receptor [41] and very low density lipoprotein (VLDL) receptor [42] which bind AAT- and other serpin-enzyme complexes and are involved in its internalisation and degradation, have been described. It is also known that AAT biosynthesis is responsive to levels of pro-inflammatory cytokines and to levels of inactivated forms of AAT through their uptake at the SEC receptor [7, 9, 43]. A parallel feedback loop may exist for inhibitory-active AAT, and as we show here for the polymerised form of AAT, in which rising levels of these molecular forms are signals for a plateau in inflammation. By saturating a receptor(s), these forms of AAT at increasing concentrations may signal down-regulation of the expression of pro-inflammatory reactants, i.e. a pro-inflammatory role of AAT at low concentrations can be linked to the phenomenon of partial agonism [44].
The observations reported here provide further support for a central role of AAT in inflammation, both as a regulator of protease activity and as a signalling molecule. The fact that polymerised AAT has parallel effects to native AAT suggests a putative common receptor acting as a sensor for rising levels of un-reacted, non-degraded molecular forms of AAT. This idea is partially supported by our earlier studies showing that low doses of oxidised (non-inhibitory) form of AAT induce pro-inflammatory monocyte activation in vitro [45].
Conclusions
Here we show that AAT may play multiple roles at sites of inflammation, which may be determined not only by the conformational modifications of AAT, but also directly by the reduction in its concentration. It is well established that native AAT is the most abundant circulating proteinase inhibitor, which is responsible for the regulation of proteolytic enzymes and the maintenance of homeostasis in humans. A substantial number of pathological problems are suggested to result from any mechanism causing the reduction in the levels of the inhibitory active form of AAT, including inactivation by non-target proteinases, oxidative inactivation, polymerisation, or genetic aberrations in structure [46–48]. Our data show that at low concentrations (at approximately 10% of normal) AAT exerts direct pro-inflammatory effects in human monocytes, leading to the hypothesis that inherited and/or acquired AAT deficiency in itself may be a consequence of the development of chronic inflammatory processes. It also explains the greater susceptibility of individuals with severe Z-AAT deficiency to develop chronic inflammatory processes. Therefore, a marked increase in AAT concentration (from 4 to 5-fold and locally up to 11-fold) during acute inflammatory reactions appears to be an anti-inflammatory condition, and the further investigation of it may provide a new target for the development of therapeutic agents to regulate inflammation.
Methods
Specific reagents
Human plasma AAT (purity > 95%, and inhibitory activity > 75%) was obtained from Calbiochem, USA and from Clinical chemistry laboratory, University Hospital Malmo, Sweden. Native AAT was diluted in PBS, pH 7.4, and, to ensure the complete removal of endotoxins, was decontaminated using endotoxin removing gel (Detoxi-Gel columns, Pierce, USA). AAT polymers were prepared as described previously [49] by incubating native AAT (final concentration 1 mg/ml) at 60°C for 3 h with gentle shaking. Polymer formation was confirmed by non-denaturing 7.5% PAGE and a concomitant loss of inhibitory activity against pancreatic elastase. The AAT-polymer mixture was reheated as necessary to ensure that the polymers were not contaminated with monomeric AAT. Protein preparations used in all assays were assayed for endotoxins using the LAL, Lumulus Amebocyte Lysate, Endochrome™ test (Sigma Chemical company, USA). In all samples of native and polymerized AAT used in the subsequent experiments endotoxin levels were less than 0.3 EU/mg of protein.
Isolation and culture of monocytes
Human monocytes were isolated from buffy coats from different donors by the Ficoll-Hypaque procedure as previously detailed [50]. Cell purity was > 97% as determined on an AC900EO AutoCounter (Swelab Instruments, AB); cell viability was analysed by staining with 0.4 % trypan blue. Monocytes were plated at a density of 4 × 106 cells/ml into plastic dishes. After removal of non-adhering cells, the remaining adherent monocytes were cultured in RPMI 1640 (Gibco, Life Technologies, Paisley, Scotland) supplemented with 2 mM N-acetyl-L-alanyl-L-glutamine, 100 U/ml penicillin, 100 μg/ml streptomycin, 1% non-essential amino acid, 2% sodium pyruvate and 20 mM Hepes (Fluka, Chemie AG) without serum at 37°C in an atmosphere of 5% CO2. Monocytes were treated with native or polymerised AAT at concentrations of 0.01, 0.02, 0.05, 0.1, 0.5 and 1 mg/ml for 18 h. Control wells contained the same volumes of PBS or were incubated in medium alone. At the end of the incubations cell-free supernatants were collected and stored at -20°C until analysis. It must be pointed out that no difference was observed between control cells alone and treated with PBS therefore all presented data are calculated from the buffer controls. Each experiment was done at a minimum in triplicate for each condition.
Cytokine and chemokine asssys
Cell culture supernatants from monocytes treated with various concentrations of native and polymerised AAT were analysed to determine TNF-α, IL-6, IL-8 and MCP-1 by using sandwich DuoSet ELISA kits according to the manufacturer's instructions (R&D Systems, England). The detection limits for the assays were as follows: IL-8, 15.6 pg/ml, MCP-1, 7.8 pg/ml, TNF-α, 7.8 pg/ml, IL-6, 2.3 pg/ml.
Nuclear extract preparation and NFκB activation monitoring assay
Nuclear extracts from monocytes were prepared exclusively as described previously [51]. Protein concentrations were determined spectrophotometrically at 260 nm and using the Comassie Plus protein assay reagent according to instructions from the manufacturer (Pierce, IL, USA).
NF-κB in nuclear extracts were analysed using TransAm kits (Activemotif, CA, USA) specific for p50 subunit according to manufactures instructions. Detection limit <0.5 μg cell extract per well.
Statistical analysis
The differences in the means in experimental results were analysed for their statistical significance by one-way analysis of variance (ANOVA) combined with a multiple comparisons procedure (Scheffé multiple range test) with the overall significance level of α = 0.05. The Statistical Package for Social Sciences (SPSS for Windows, release 11.0) was used for the calculations.
Abbreviations
- AAT:
-
alpha1-antitrypsin, PiZZ, homozygous type Z deficiency variant
- PiMM:
-
normal AAT gene variant
- MCP-1:
-
monocyte chemoattractant protein-1
- TNFα:
-
tumour necrosis factor-α
- IL:
-
interleukin, PBS, phosphate buffered saline
- NF-κB:
-
nuclear factor-κB, Ig, immunoglobulin
- PAGE:
-
polyacrylamide gel electrophoresis
- LPS:
-
lipopolysacharide
- SEC:
-
serpin-enzyme complex
- LRP:
-
low density lipoprotein related protein
- LDL:
-
low density lipoprotein
- VLDL:
-
very low density lipoprotein.
References
Perlmutter DH, Cole FS, Kilbridge TH, Rossing TH, Colten HR: The cellular defect in alpha 1-proteinase inhibitor (alpha 1-PI) deficiency is expressed in human monocytes and in Xenopus oocytes injected with human liver mRNA. Proc Natl Acad Sci USA. 1985, 82: 795-799.
Ray MB, Desmet VJ, Gepts W: alpha-1-Antitrypsin immunoreactivity in islet cells of adult human pancreas. Cell Tissue Res. 1977, 185: 63-68.
Geboes K, Ray MB, Rutgeerts P, Callea F, Desmet VJ, Vantrappen G: Morphological identification of alpha-1-antitrypsin in the human small intestine. Histopathology. 1982, 6: 55-60.
Boskovic G, Twining SS: Local control of alpha1-proteinase inhibitor levels: regulation of alpha1-proteinase inhibitor in the human cornea by growth factors and cytokines. Biochim Biophys Acta. 1998, 1403: 37-46. 10.1016/S0167-4889(98)00018-4.
Laurell CB, Nosslin B, Jeppsson JO: Catabolic rate of alpha1-antitrypsin of Pi type M and Z in man. Clin Sci Mol Med. 1977, 52: 457-461.
Lisowska-Myjak B, Pachecka J: Antigenic and functional levels of alpha-1-antitrypsin in serum during normal and diabetic pregnancy. Eur J Obstet Gynecol Reprod Biol. 2003, 106: 31-35. 10.1016/S0301-2115(02)00201-4.
Perlmutter DH, Punsal PI: Distinct and additive effects of elastase and endotoxin on expression of alpha 1 proteinase inhibitor in mononuclear phagocytes. J Biol Chem. 1988, 263: 16499-16503.
Knoell DL, Ralston D, Coulter KR, Wewers MD: Alpha 1-antitrypsin and protease complexation is induced by lipopolysaccharide, interleukin-1beta, and tumor necrosis factor-alpha in monocytes. Am J Respir Crit Care Med. 1998, 157: 246-255.
Perlmutter DH, Glover GI, Rivetna M, Schasteen CS, Fallon RJ: Identification of a serpin-enzyme complex receptor on human hepatoma cells and human monocytes. Proc Natl Acad Sci USA. 1990, 87: 3753-3757.
Boudier C, Pelletier A, Gast A, Tournier JM, Pauli G, Bieth JG: The elastase inhibitory capacity and the alpha 1-proteinase inhibitor and bronchial inhibitor content of bronchoalveolar lavage fluids from healthy subjects. Biol Chem Hoppe Seyler. 1987, 368: 981-990.
Hubbard RC, Crystal RG: Strategies for aerosol therapy of alpha 1-antitrypsin deficiency by the aerosol route. Lung. 1990, 168: 565-578.
Janciauskiene S, Dominaitiene R, Sternby NH, Piitulainen E, Eriksson S: Detection of circulating and endothelial cell polymers of Z and wild type alpha 1-antitrypsin by a monoclonal antibody. J Biol Chem. 2002, 277: 26540-26546. 10.1074/jbc.M203832200.
Elliott PR, Pei XY, Dafforn TR, Lomas DA: Topography of a 2.0 A structure of alpha1-antitrypsin reveals targets for rational drug design to prevent conformational disease. Protein Sci. 2000, 9: 1274-1281.
Foreman RC: Alpha 1-antitrypsin deficiency – a defect in secretion. Biosci Rep. 1987, 7: 307-311.
Sveger T: Liver disease in alpha1-antitrypsin deficiency detected by screening of 200,000 infants. New Engl J Med. 1976, 294: 1316-1321.
Durr R, Caselmann WH: Carcinogenesis of primary liver malignancies. Langenbeck's Arch Surg. 2000, 385: 154-161. 10.1007/s004230050259.
Lomas DA, Evans DL, Finch JT, Carrell RW: The mechanism of Z alpha 1-antitrypsin accumulation in the liver. Nature. 1992, 357: 605-607. 10.1038/357605a0.
Elliott PR, Bilton D, Lomas DA: Lung polymers in Z alpha1-antitrypsin deficiency-related emphysema. Am J Respir Cell Mol Biol. 1998, 18: 670-674.
Lomas DA, Evans DL, Stone SR, Chang WS, Carrell RW: Effect of the Z mutation on the physical and inhibitory properties of alpha 1-antitrypsin. Biochemistry. 1993, 32: 500-508.
Hill AT, Campbell EJ, Bayley DL, Hill SL, Stockley RA: Evidence for excessive bronchial inflammation during an acute exacerbation of chronic obstructive pulmonary disease in patients with alpha(1)-antitrypsin deficiency (PiZ). Am J Respir Crit Care Med. 1999, 160: 1968-1975.
Dabbagh K, Laurent GJ, Shock A, Leoni P, Papakrivopoulou J, Chambers RC: Alpha-1-antitrypsin stimulates fibroblast proliferation and procollagen production and activates classical MAP kinase signalling pathways. J Cell Physiol. 2001, 186: 73-81. 10.1002/1097-4652(200101)186:1<73::AID-JCP1002>3.0.CO;2-Q.
Jeannin P, Lecoanet-Henchoz S, Delneste Y, Gauchat JF, Bonnefoy JY: Alpha-1 antitrypsin up-regulates human B cell differentiation selectively into IgE- and IgG4-secreting cells. Eur J Immunol. 1998, 28: 1815-1822. 10.1002/(SICI)1521-4141(199806)28:06<1815::AID-IMMU1815>3.0.CO;2-5.
Ikari Y, Mulvihill E, Schwartz SM: alpha 1-Proteinase inhibitor, alpha 1-antichymotrypsin, and alpha 2-macroglobulin are the antiapoptotic factors of vascular smooth muscle cells. J Biol Chem. 2001, 276: 11798-11803. 10.1074/jbc.M008503200.
Weiss G, Graziadel I, Urbanek M, Grunewald K, Vogel W: Divergent effects of alpha 1-antitrypsin on the regulation of iron metabolism in human erythroleukaemic (K562) and myelomonocytic (THP-1) cells. Biochem J. 1996, 319: 897-902.
Bucurenci N, Blake DR, Chidwick K, Winyard PG: Inhibition of neutrophil superoxide production by human plasma alpha 1-antitrypsin. FEBS Lett. 1992, 300: 21-24. 10.1016/0014-5793(92)80156-B.
Tilg H, Vannier E, Vachino G, Dinarello CA, Mier JW: Antiinflammatory properties of hepatic acute phase proteins: preferential induction of interleukin 1 (IL-1) receptor antagonist over IL-1 beta synthesis by human peripheral blood mononuclear cells. J Exp Med. 1993, 178: 1629-1636.
Jie Z, Cai Y, Yang W, Jin M, Zhu W, Zhu C: Protective effects of alpha(1)-antitrypsin on acute lung injury in rabbits induced by endotoxin. Chin Med J (Engl). 2003, 116: 1678-1682.
Qing-Bai S, Mukherjee JJ, Crilly KS, Kiss Z: alpha(1)-antitrypsin can increase insulin-induced mitogenesis in various fibroblast and epithelial cell lines. FEBS Lett. 2000, 473: 33-36. 10.1016/S0014-5793(00)01492-7.
Yao J, Baecher-Allan CM, Sharon J: Serpins identified as cell growth inhibitors in human plasma. Mol Cell Biol Res Commun. 2000, 3: 76-81. 10.1006/mcbr.2000.0192.
Aoshiba K, Nagai A, Ishihara Y, Kagawa J, Takizawa T: Effects of alpha 1-proteinase inhibitor on chemotaxis and chemokinesis of polymorphonuclear leukocytes: its possible role in regulating polymorphonuclear leukocyte recruitment in human subjects. J Lab Clin Med. 1993, 122: 333-340.
Parmar JS, Mahadeva R, Reed BJ, Farahi N, Cadwallader KA, Keogan MT, Bilton D, Chilvers ER, Lomas DA: Polymers of alpha(1)-antitrypsin are chemotactic for human neutrophils: a new paradigm for the pathogenesis of emphysema. Am J Respir Cell Mol Biol. 2002, 26: 723-730.
Bitterman PB, Saltzman LE, Adelberg S, Ferrans VJ, Crystal RG: Alveolar macrophage replication. One mechanism for the expansion of the mononuclear phagocyte population in the chronically inflamed lung. J Clin Invest. 1984, 74: 460-469.
Banda MJ, Rice AG, Griffin GL, Senior RM: Alpha 1-proteinase inhibitor is a neutrophil chemoattractant after proteolytic inactivation by macrophage elastase. J Biol Chem. 1988, 263: 4481-4484.
Yamamoto Y, Gaynor RB: Therapeutic potential of inhibition of the NF-kappaB pathway in the treatment of inflammation and cancer. J Clin Invest. 2001, 107: 135-142.
Pahl HL: Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999, 18: 6853-6866. 10.1038/sj.onc.1203239.
Dinarello CA: Interleukin-18. Methods. 1999, 19: 121-132. 10.1006/meth.1999.0837.
Hall AJ, Vos HL, Bertina RM: Lipopolysaccharide induction of tissue factor in THP-1 cells involves Jun protein phosphorylation and nuclear factor kappaB nuclear translocation. J Biol Chem. 1999, 274: 376-383. 10.1074/jbc.274.1.376.
Rollins BJ: Monocyte chemoattractant protein 1: a potential regulator of monocyte recruitment in inflammatory disease. Mol Med Today. 1996, 2: 198-204. 10.1016/1357-4310(96)88772-7.
Burger D, Dayer JM: Cytokines, acute-phase proteins, and hormones: IL-1 and TNF-alpha production in contact-mediated activation of monocytes by T lymphocytes. Ann N Y Acad Sci. 2002, 966: 464-473.
Poller W, Willnow TE, Hilpert J, Herz J: Differential recognition of alpha 1-antitrypsin-elastase and alpha 1-antichymotrypsin-cathepsin G complexes by the low density lipoprotein receptor-related protein. J Biol Chem. 1995, 270: 2841-2845. 10.1074/jbc.270.6.2841.
Kounnas MZ, Church FC, Argraves WS, Strickland DK: Cellular internalization and degradation of antithrombin III-thrombin, heparin cofactor II-thrombin, and alpha 1-antitrypsin-trypsin complexes is mediated by the low density lipoprotein receptor-related protein. J Biol Chem. 1996, 271: 6523-6529. 10.1074/jbc.271.11.6523.
Rodenburg KW, Kjoller L, Petersen HH, Andreasen PA: Binding of urokinase-type plasminogen activator-plasminogen activator inhibitor-1 complex to the endocytosis receptors alpha2-macroglobulin receptor/low-density lipoprotein receptor-related protein and very-low-density lipoprotein receptor involves basic residues in the inhibitor. Biochem J. 1998, 329: 55-63.
Morgan K, Kalsheker NA: Regulation of the serine proteinase inhibitor (SERPIN) gene alpha 1-antitrypsin: a paradigm for other SERPINs. Int J Biochem Cell Biol. 1997, 29: 1501-1511. 10.1016/S1357-2725(97)00118-0.
Pliska V: Partial agonism: mechanisms based on ligand-receptor interactions and on stimulus-response coupling. J Recept Signal Transduct Res. 1999, 19: 597-629.
Moraga F, Janciauskiene S: Activation of primary human monocytes by the oxidized form of alpha1-antitrypsin. J Biol Chem. 2000, 275: 7693-7700. 10.1074/jbc.275.11.7693.
Enghild JJ, Valnickova Z, Thogersen IB, Pizzo SV: Complexes between serpins and inactive proteinases are not thermodynamically stable but are recognized by serpin receptors. J Biol Chem. 1994, 269: 20159-20166.
Crowther DC, Evans DL, Carrell RW: Serpins: implications of a mobile reactive centre. Curr Opin Biotechnol. 1992, 3: 399-407.
Stein PE, Carrell RW: What do dysfunctional serpins tell us about molecular mobility and disease?. Nat Struct Biol. 1995, 2: 96-113.
Dafforn TR, Mahadeva R, Elliott PR, Sivasothy P, Lomas DA: A kinetic mechanism for the polymerization of alpha1-antitrypsin. J Biol Chem. 1999, 274: 9548-9555. 10.1074/jbc.274.14.9548.
Janciauskiene S, Moraga F, Lindgren S: C-terminal fragment of alpha1-antitrypsin activates human monocytes to a pro-inflammatory state through interactions with the CD36 scavenger receptor and LDL receptor. Atherosclerosis. 2001, 158: 41-51. 10.1016/S0021-9150(00)00767-X.
Dichtl W, Moraga F, Ares MP, Crisby M, Nilsson J, Lindgren S, Janciauskiene S: The carboxyl-terminal fragment of alpha1-antitrypsin is present in atherosclerotic plaques and regulates inflammatory transcription factors in primary human monocytes. Mol Cell Biol Res Commun. 2000, 4: 50-61. 10.1006/mcbr.2000.0256.
Abdelmonem A Afifi, Stanley P Azen: Statistical analysis: A Computer oriented approach. Academic press. 1997
Acknowledgments
This work was supported by grants from the Department of Medicine, Lund University and AstraZeneca R&D, Lund, Sweden.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
RA carried out the experimental work and wrote the preliminary draft of the manuscript. LJ participated in the design of the study. SJ designed and coordinated the study, performed statistical analysis of the results, made major revision of the manuscript.
All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Aldonyte, R., Jansson, L. & Janciauskiene, S. Concentration-dependent effects of native and polymerised α1-antitrypsin on primary human monocytes, in vitro. BMC Cell Biol 5, 11 (2004). https://doi.org/10.1186/1471-2121-5-11
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2121-5-11