
Abstract
Fueled by the promise of regenerative medicine, currently there is unprecedented interest in stem cells. Furthermore, there have been revolutionary, but somewhat controversial, advances in our understanding of stem cell biology. Stem cells likely play key roles in the repair of diverse lung injuries. However, due to very low rates of cellular proliferation in vivo in the normal steady state, cellular and architectural complexity of the respiratory tract, and the lack of an intensive research effort, lung stem cells remain poorly understood compared to those in other major organ systems. In the present review, we concisely explore the conceptual framework of stem cell biology and recent advances pertinent to the lungs. We illustrate lung diseases in which manipulation of stem cells may be physiologically significant and highlight the challenges facing stem cell-related therapy in the lung.
Similar content being viewed by others


Introduction
According to Greek mythology, the immortal Prometheus stole fire from the Gods as a gift for humankind. As punishment, he was shackled to a rock, whereupon each day for 30,000 years an eagle consumed as much of his liver as would regenerate. There is some debate whether the eagle ate his liver or heart, but what if the bird had a taste for lung? And what if Prometheus was a mere mortal?
Analogous to Prometheus and the eagle, the ambient air-exposed lung is subject to an array of potentially damaging agents, including chemical oxidants and proteolytic enzymes. Presumably, daily oxidant and protease wear and tear on structural components such as elastin and collagen contributes to inevitable age-related declines in pulmonary function in normal individuals [1, 2]. Acute and chronic lung disease, or its treatment with oxygen and positive pressure ventilation, may further damage lung tissue in excess of the capacity for orderly repair, resulting in characteristic pathologic changes including tissue destruction or fibrotic scarring [3–5]. But what determines the lungs' capacity for repair? Certainly, one factor must be the ability of stem cells to proliferate and differentiate to replace damaged cells and tissues. As discussed later in this review, the traditional view is that, during development, self-renewing tissues are imbued with resident, tissue-specific stem cells, so-called adult somatic stem cells. However, recent but highly controversial evidence suggests that stem cells from one type of tissue may generate cells typical of other organs. In this fashion, circulating cells derived from bone marrow may augment resident stem cells, and we comprehensively review such data from lung. Finally, there is great hope that embryonic stem cells, embryonic germ cells, or even adult somatic stem cells can be engineered as an unlimited source of cells to enhance organ-specific repair or replace lost tissues. Below, we concisely review stem cell biology, focusing on recent findings relevant to the lungs. Diseases in which alterations in stem cells contribute to lung dysfunction are discussed, as are the challenges facing the nascent field of pulmonary regenerative medicine.
Embryonic and adult (somatic) stem cells
For links to more in-depth information on general principles in stem cell biology, a comprehensive glossary, and the latest updates in this quick moving field, the reader is referred to the International Society for Stem Cell Biology http://www.isscr.org. During embryonic development, the inner cell mass of the blastocyst forms three primary germ layers, which generate all fetal tissue lineages (reviewed in [6], illustrated in Figure 1, path 1). Embryonic stem cells (derived from the blastocyst inner cell mass), or embryonic germ cells (derived from the gonadal ridge), when cultured on embryonic mouse fibroblast feeder cell layers in the presence of a differentiation-suppressing cytokine (leukemia inhibitory factor), proliferate indefinitely and remain pluripotent. Manipulation of culture conditions can coax the cells to undergo differentiation characteristic of many tissue types (Figure 1, paths 2 and 3). Theoretically, pluripotent embryonic cells can serve as an unlimited resource for therapeutic applications [7, 8].

Cell lineage determination during embryogenesis and generation of pluripotent embryonic cells. The three primary germ layers form during normal development (path 1). Embryonic stem cells from the inner cell mass (path 2) or embryonic germ cells from the gonadal ridge (path 3) can be cultured and manipulated to generate cells of all three lineages.
General principles of tissue renewal by adult stem cells have been reviewed recently [9] and can be summarized as follows. The traditional view of cell lineages is that adult somatic stem cells maintain cell populations in adult tissues. The adult lung falls into the category in which cell proliferation is very low in the normal steady-state but can be induced dramatically by injury (see [10, 11] for recent reviews of lung stem cells). The conditional nature of lung cell proliferation complicates the search for lung stem cells. Cell lineages are much better understood in continuously proliferating tissues such as the gut, skin and hematopoietic system (reviewed in [12–14], respectively). The long-standing view, developed from these other organs, is that stem cells reside in well-protected, innervated, and vascularized niches that provide cues regulating cell fate decisions such as proliferation, migration, and differentiation [15]. Adult stem cells are capable of abundant self-renewal and can also generate the specific cell lineages within the tissue compartment (Figure 2). Proportional to tissue needs, stem cells may undergo asymmetric cell division, in which they generate one stem cell and a committed progenitor. The capacity for self-renewal decreases progressively as committed progenitors differentiate. The wisdom of the body is to conserve stem cells. They cycle infrequently and the majority of cell replacement is accomplished by committed progenitors within the so-called transiently amplifying compartment. Eventually, individual cells become incapable of further cell division. In tissues, there are specific temporal and spatial hierarchic relationships between stem cells in their niches and their differentiated progeny. Within this axis, cell proliferation, migration, differentiation, function, death, and removal are tightly regulated to maintain tissue homeostasis.

Traditional view of cell lineage in adult renewing tissues. Organ-specific (somatic) stem cells generate characteristic cell types through a linear set of commitment and differentiation steps. Arrow thickness represents self-renewal potential.
Cell compartments in the lung and functional integration
In the architecturally complex lung, cells of multiple germinal lineages interact both during morphogenesis and to maintain adult lung structure. Even within derivatives of a single germ layer, cells become subdivided into separate cell lineage "zones". For example, the endoderm generates least four distinct epithelial regions, each with a different cellular composition (Figure 3). Additional cell types, including airway smooth muscle, fibroblasts, and the vasculature, are derived from mesoderm. Airway and alveolar architecture, and in turn, function, result from interaction among epithelium, smooth muscle, fibroblasts, and vascular cells, all within an elaborate structural matrix of connective tissue. The complexity of even this oversimplified view, which omits pulmonary neuroepithelial cells and bodies, innervation, and classical hematopoietically-derived cells such as dendritic cells, mast cells, and macrophages, has hindered identification of lung stem cells and patterns of cell migration during tissue renewal. Nevertheless, the prevailing view is that airway basal and Clara cells and alveolar type II cells serve as epithelial progenitors [11, 16–19]. Cell lineages in the mesodermal compartments remain less well understood.

Stem cell compartments in the lungs. The endoderm-derived epithelium can be subdivided into at least 4 types whereas smooth muscle, fibroblasts, and vascular cells are derived from mesoderm. The coordinated interaction of multiple cell types, including alveolar epithelium, interstitial fibroblasts, myofibroblasts and pulmonary endothelium, is necessary to form alveolar septa.
Stem cell plasticity and the lung
Recent studies challenge the view that tissues are maintained solely by organ-specific stem cells. There is evidence that adult stem cells from a variety of sources can generate not only their own lineages, but those of other tissues, sometimes crossing barriers of embryonic derivation previously thought impenetrable [20, 21, 8]. There are a few controversial reports that adult stem cells from outside the bone marrow may reconstitute the hematopoietic system, but most of the evidence flows in the other direction- namely, that cells from the bone marrow can generate diverse non-hematopoietic cell types. Both experimental studies in animals and human clinical studies, summarized in Table 1, provide evidence for, and against, circulatory delivery of lung progenitor cells. While bone marrow-derived cells, such as alveolar macrophages, dendritic cells, mast cells, and lymphocytes, normally migrate to the lung, the surprise in the recent literature is that under certain circumstances circulating cells can apparently generate lung resident cells, including epithelial, endothelial, and myofibroblast cells. The technical approach towards identification of these cells is often technically challenging and involves co-localization of a donor cell marker, for example, the Y chromosome, in sex-mismatched transplantation, or a genetically engineered marker in mouse experiments, and proteins characteristic of the differentiated cell type in the lung, for example, keratin in epithelial cells or collagen in fibroblasts. As discussed below, the results are highly variable and often contradictory, depending on factors including the starting cell population, the methods for marker detection, and the amount of injury to the lung.
Transplantation studies in mice can be performed using whole donor bone marrow, the fraction that adheres in culture, termed marrow stromal cells (MSC), or preparations enriched for hematopoietic stem cells (HSC). Whole body irradiation, which may injure lung tissue, is typically used to deplete the host bone marrow. Importantly, lung injury apparently enhances engraftment into lung [22–29]. Whole bone marrow, MSC, or HSC have all been reported to reconstitute lung parenchymal cells. MSC transplantation resulted in collagen I expressing donor cells in the lung [30], and in the presence of bleomycin injury, MSC reportedly generated type I [23] or type II pneumocytes [22]. Transplantation with HSCs yielded up to 20% donor-derived pneumocytes and 4% bronchial epithelial cells [31]. However, other investigators have identified only hematopoeitic chimerism by HSCs [32]. Whole bone marrow infusion generated type II pneumocytes [33], or fibroblasts and type I pneumocytes [34]. Radiation pneumonitis augmented whole bone marrow generation of type II pneumocytes and bronchial epithelial cells [25] or fibroblasts [26]. Bleomycin lung injury enhanced formation of type I collagen-producing cells [24] from whole bone marrow, whereas elastase-induced emphysema stimulated formation of alveolar epithelium and endothelium [28]. Lung injury alone, without bone marrow transplantation, may promote stem cell migration. For example, in the ovalbumin model of asthma, circulating fibrocytes were recruited into bronchial tissue [29], and in a bovine model of hypoxic pulmonary hypertension, cells capable of generating endothelial and smooth muscle cells in vitro were found in the circulation [27].
Sex-mismatched lung and bone marrow transplantation in humans provides a natural model for analysis of donor and recipient cell behavior. Bronchial epithelial and gland cells and type II pneumocytes of host origin were reported in one study of lung allografts [35], but not another [36]. After bone marrow transplantation, epithelial cells of donor origin were not detected in the nasal passages [37]. Similar to lung allografts, following bone marrow transplantation, epithelium and endothelium of donor origin were found in one study [38], but not another [35].
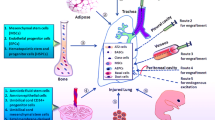
Many questions remain unanswered. The mechanism whereby cells assume lung cell phenotypes remains uncertain. Several studies have demonstrated that cell fusion occurs both in vitro and in vivo, which likely explains why some of the cells contain both donor and lung cell markers [see [39] for a study of fusion of MSCs and lung epithelium and [40, 41] for recent reviews]. Alternatively, cells may reprogram in the lung environment- a concept termed "transdifferentiation", which is defined as the ability of a particular cell from one tissue type to differentiate into a cell type characteristic of another tissue. It has been suggested that many of the events previously attributed to transdifferentiation may actually represent cell fusions, particularly due to the influx of fusion-prone myeloid cells into damaged tissues from the repopulated bone marrow [40]. New, more stringent, criteria have been put forth for demonstration of transdifferentiation [41]. Bone marrow harbors a generalized pluripotent stem cell [42] and the bone marrow cell responsible for lung engraftment has not been identified with certainty. It is possible that rare transdifferentiation events represent migration of a pluripotent bone marrow cell type resembling an embryonic stem or embryonic germ cell still harbored in the adult bone marrow. It remains unknown whether bone marrow cells must transit through an intermediate compartment prior to lung colonization (Figure 4) or whether circulating stem cells can be mobilized from sources other than bone marrow. It is important to note that bone marrow derived cells of typical hematopoietic lineage, chimeric cells created by fusion, or lung cells generated by transdifferentiation may all play a role in lung repair by promoting the local production of stem cells or reparative function of lung-specific cell types. A compelling study suggests that mesenchymal stem cells from bleomycin-resistant mice can mitigate the pro-fibrotic effects of bleomycin in sensitive mice [22], while another study suggests that bone marrow cells actively contribute to the formation of fibrotic tissue [24]. Mitigating or exacerbating roles for bone marrow derived cells in lung repair or fibrosis are not mutually exclusive. The important concepts of whether the lungs' capacity for repair is dependent on circulating cells, and whether exogenously delivered cells can enhance resistance to injury or promote healing, remain unanswered and controversial.

Evolving view of cell lineages in the lungs. The functional significance of circulating cells towards lung cell maintenance or tissue repair remains unknown, as does the precise mechanism whereby circulating cells generate lung cell types.
Lung "stem cell" diseases
Major lung diseases likely involving stem cells and the cellular targets for stem cell therapy are summarized in Table 2. These may be broadly categorized whether they involve stem cell deficiency, hyper-proliferation or possibly, a combination of both. For example, impaired pulmonary endothelial and/or epithelial barrier function may contribute to the pathophysiology of adult respiratory distress syndrome. Mobilization of endogenous endothelial or epithelial stem/progenitor cells or delivery of adult somatic stem cells, embryonic stem cells, or embryonic germ cells may theoretically improve barrier function, supporting the notion of treating a "stem cell deficiency". Similarly, toxic, viral or alloimmune destruction of the bronchiolar epithelium suggests stem cell deficiency in bronchiolitis obliterans. However, fibrotic reactions and scarring in response to epithelial injury can be viewed as fibroblast "stem cell hyper-proliferation". The general concept is that augmentation of stem cells may minimize lung injury, augment repair, or possibly regenerate lost tissue. However, one must also consider that inhibiting excessive growth of stem cells may be a valid therapeutic goal when hyper-proliferation contributes to disease pathophysiology, as in fibrosis, smooth muscle hyperplasia or lung cancer.
Challenges for lung regenerative medicine
What are the realistic prospects for beneficial stem cell therapy of the lung? First, we must conclusively identify lung diseases/cases/timing in which cell and tissue damage occurs in excess of the capacity for timely endogenous repair. Second, we must establish standardized sources of relevant stem/progenitor cells and methods for their delivery to the appropriate lung sub-compartment. Once delivered, therapeutic cells must home to microscopic sites of need and integrate to serve a beneficial function. There is clearly potential for adverse effects, as exemplified by the propensity of embryonic stem cells to form teratomas when implanted in vivo [43]. Major lung diseases potentially addressable by stem cell therapy may pose unique challenges. Reversal of lung developmental anomalies resulting in hypoplasia, or repair of chronic lung disease of prematurity and advanced pulmonary emphysema in adults, will require neogenesis of alveolar septa in which the endogenous "tissue blueprint" never developed, or was completely destroyed. Until we gain a much better understanding of lung tissue morphogenesis, we must rely on stem cells intrinsically "knowing" where to go and "how" to recreate alveolar septal architecture to ultimately restore higher order complex three dimensional relationships amongst alveoli, airways, and vessels. Stem cell therapy to cure cystic fibrosis will require heterologous, or gene corrected autologous, stem cells to colonize the airway, proliferate, and differentiate into columnar cells covering a significant portion of the airway lumen. However, most evidence thus far suggests that cells from the circulation may generate isolated, single airway basal cells. Stem cell therapy to mitigate respiratory distress syndrome (RDS) will require cells capable of restoring alveolar endothelial and epithelial function in the face of evolving injury. Whereas injury is thought to promote stem cell recruitment, the relevant question is whether it can occur quickly enough to meaningfully reverse acute, widespread cellular dysfunction typical of RDS.
Conclusion
Provocative, but controversial, recent evidence suggests that circulating stem cells may home to the lung. There is great excitement and hope that exogenous and/or mobilized endogenous stem cells may be harnessed to prevent or treat acute and chronic lung diseases and even regenerate abnormally developed or lost tissue. Our understanding of lung stem cells and the regulation of lung morphogenesis is still rudimentary, and the complex, integrated function of multiple cell types underlying normal lung structure and function poses unique challenges. Thus, the therapeutic prospects for stem cell therapy in lungs appear more distant than in some other organs. This realization should stimulate meaningful new studies from the lung research community. Unlike the mythical hero Prometheus, patients with lung disease cannot wait 30,000 years!
References
Mahler DA, Rosiello RA, Loke J: The aging lung. Clin Geriatr Med 1986, 2:215–225.
Mahler DA, Fierro-Carrion G, Baird JC: Evaluation of dyspnea in the elderly. Clin Geriatr Med 2003, 19:19–33.
Jeffery PK: Remodeling in asthma and chronic obstructive lung disease. Am J Respir Crit Care Med 2001, 164:S28-S38.
Ozkan M, Dweik RA, Ahmad M: Drug-induced lung disease. Cleve Clin J Med 2001, 68:782–795.
Chow CW, Herrera Abreu MT, Suzuki T, Downey GP: Oxidative stress and acute lung injury. Am J Respir Cell Mol Biol 2003, 29:427–431.
Loebel DAF, Watson CM, De Young RA, Tam PPL: Lineage choice and differentiation in mouse embryos and embryonic stem cells. Dev Biol 2003, 264:1–14.
West JA, Daley GQ: Human embryonic stem cells. Bone Marrow Transplant 2004, 33:135.
Daley GQ, Goodell MA, Snyder EY: Realistic prospects for stem cell therapeutics. Hematology (Am Soc Hematol Educ Program) 2003, 398–418.
Stem Cells Handbook(Edited by: Sell S). Totowa, Humana Press 2003.
Otto WR: Lung epithelial stem cells. J Pathol 2002, 197:527–535.
Ten Have-Opbroek AAW, Randell SH, Stripp BR: Stem cells in lung morphogenesis, regeneration, and carcinogenesis. Stem Cells Handbook (Edited by: SellS). Totowa, Humana Press 2003, 455–472.
Marshman E, Booth C, Potten CS: The intestinal epithelial stem cell. Bioessays 2002, 24:91–98.
Alonso L, Fuchs E: Stem cells of the skin epithelium. Proc Natl Acad Sci U S A 2003, 100:11830–11835.
Kondo M, Wagers AJ, Manz MG, Prohaska SS, Scherer DC, Beilhack GF, Shizuru JA, Weissman IL: Biology of hematopoietic stem cells and progenitors: implications for clinical application. Annu Rev Immunol 2003, 21:759–806.
Watt FM, Hogan BL: Out of Eden: stem cells and their niches. Science 2000, 287:1427–1430.
Schoch KG, Lori A, Burns KA, Eldred T, Olsen JC, Randell SH: A subset of mouse tracheal epithelial basal cells generates large colonies in vitro. Am J Physiol Lung Cell Mol Physiol 2004, 286:631–642.
Hong KU, Reynolds SD, Watkins S, Fuchs E, Stripp BR: Basal cells are a multipotent progenitor capable of renewing the bronchial epithelium. Am J Pathol 2004, 164:577–588.
Hong KU, Reynolds SD, Watkins S, Fuchs E, Stripp BR: In vivo differentiation potential of tracheal basal cells: evidence for multipotent and unipotent subpopulations. Am J Physiol Lung Cell Mol Physiol 2004, 286:L643-L649.
Hong KU, Reynolds SD, Giangreco A, Hurley CM, Stripp BR: Clara cell secretory protein-expressing cells of the airway neuroepithelial body microenvironment include a label-retaining subset and are critical for epithelial renewal after progenitor cell depletion. Am J Respir Cell Mol Biol 2001, 24:671–681.
Koerbling M, Estrov Z: Adult stem cells for tissue repair - A new therapeutic concept? N Engl J Med 2003, 349:570–582.
Herzog EL, Chai L, Krause DS: Plasticity of marrow-derived stem cells. Blood 2003, 102:3483–3493.
Ortiz LA, Gambelli F, McBride C, Gaupp D, Baddoo M, Kaminski N, Phinney DG: Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc Natl Acad Sci U S A 2003, 100:8407–8411.
Kotton DN, Ma BY, Cardoso WV, Sanderson EA, Summer RS, Williams MC, Fine A: Bone marrow-derived cells as progenitors of lung alveolar epithelium. Development 2001, 128:5181–5188.
Hashimoto N, Jin H, Liu T, Chensue SW, Phan SH: Bone marrow-derived progenitor cells in pulmonary fibrosis. J Clin Invest 2004, 113:243–252.
Theise ND, Henegariu O, Grove J, Jagirdar J, Kao PN, Crawford JM, Badve S, Saxena R, Krause DS: Radiation pneumonitis in mice: a severe injury model for pneumocyte engraftment from bone marrow. Exp Hematol 2002, 30:1333–1338.
Epperly MW, Guo H, Gretton JE, Greenberger JS: Bone marrow origin of myofibroblasts in irradiation pulmonary fibrosis. Am J Respir Cell Mol Biol 2003, 29:213–224.
Davie NJ, Crossno JT, Frid MG, Hofmeister SE, Reeves JT, Hyde DM, Carpenter TC, Brunetti JA, McNiece IK, Stenmark KR: Hypoxia-induced pulmonary artery adventitial remodeling and neovascularization: potential contribution of progenitor cells. Am J Physiol Lung Cell Mol Physiol 2003, 286:L668-L678.
Ishizawa K, Kubo H, Yamada M, Kobayashi S, Numasaki M, Ueda S, Suzuki T, Sasaki H: Bone marrow-derived cells contribute to lung regeneration after elastase-induced pulmonary emphysema. FEBS Lett 2004, 556:249–252.
Schmidt M, Sun G, Stacey MA, Mori L, Mattoli S: Identification of circulating fibrocytes as precursors of bronchial myofibroblasts in asthma. J Immunol 2003, 171:380–389.
Pereira RF, Halford KW, O'Hara MD, Leeper DB, Sokolov BP, Pollard MD, Bagasra O, Prockop DJ: Cultured adherent cells from marrow can serve as long-lasting precursor cells for bone, cartilage, and lung in irradiated mice. Proc Natl Acad Sci U S A 1995, 92:4857–4861.
Krause DS, Theise ND, Collector MI, Henegariu O, Hwang S, Gardner R, Neutzel S, Sharkis SJ: Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell 2001, 105:369–377.
Wagers AJ, Sherwood RI, Christensen JL, Weissman IL: Little evidence for developmental plasticity of adult hematopoietic stem cells. Science 2002, 297:2256–2259.
Grove JE, Lutzko C, Priller J, Henegariu O, Theise ND, Kohn DB, Krause DS: Marrow-derived cells as vehicles for delivery of gene therapy to pulmonary epithelium. Am J Respir Cell Mol Biol 2002, 27:645–651.
Abe S, Lauby G, Boyer C, Rennard S, Sharp J: Transplanted BM and BM side population cells contribute progeny to the lung and liver in irradiated mice. Cytotherapy 2003, 5:523–533.
Kleeberger W, Versmold A, Rothamel T, Glockner S, Bredt M, Haverich A, Lehmann U, Kreipe H: Increased chimerism of bronchial and alveolar epithelium in human lung allografts undergoing chronic injury. Am J Pathol 2003, 162:1487–1494.
Bittmann I, Dose T, Baretton GB, Muller C, Schwaiblmair M, Kur F, Lohrs U: Cellular chimerism of the lung after transplantation. An interphase cytogenetic study. Am J Clin Pathol 2001, 115:525–533.
Davies JC, Potter M, Bush A, Rosenthal M, Geddes DM, Alton EWFW: Bone marrow stem cells do not repopulate the healthy upper respiratory tract. Pediatr Pulmonol 2002, 34:251–256.
Suratt BT, Cool CD, Seris AE, Chen L, Varella-Garcia M, Shpall EJ, Brown KK, Worthen GS: Human pulmonary chimerism after hematopoietic stem cell transplantation. Am J Respir Crit Care Med 2003, 168:318–322.
Spees JL, Olson SD, Ylostalo J, Lynch PJ, Smith J, Perry A, Peister A, Wang MY, Prockop DJ: Differentiation, cell fusion, and nuclear fusion during ex vivo repair of epithelium by human adult stem cells from bone marrow stroma. Proc Natl Acad Sci U S A 2003, 100:2397–2402.
Camargo FD, Chambers SM, Goodell MA: Stem cell plasticity: from transdifferentiation to macrophage fusion. Cell Prolif 2004, 37:55–65.
Wagers AJ, Weissman IL: Plasticity of adult stem cells. Cell 2004, 116:639–648.
Jiang Y, Jahagirdar BN, Reinhardt RL, Schwartz RE, Keene CD, Ortiz-Gonzalez XR, Reyes M, Lenvik T, Lund T, Blackstad M, Du J, Aldrich S, Lisberg A, Low WC, Largaespada DA, Verfaillie CM: Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418:41–49.
Takahashi K, Mitsui K, Yamanaka S: Role of ERas in promoting tumour-like properties in mouse embryonic stem cells. Nature 2003, 423:541–545.
Acknowledgements
The authors thank Lisa Brown for outstanding assistance with graphics, editing, and manuscript production.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
None declared.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Neuringer, I.P., Randell, S.H. Stem cells and repair of lung injuries. Respir Res 5, 6 (2004). https://doi.org/10.1186/1465-9921-5-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1465-9921-5-6