
Abstract
Microbially induced calcite precipitation (MICP) technique utilizes ureolytic bacteria to decompose urea and generate carbonate ions for metal combination. MICP can remediate heavy metal (e.g., Cd) contaminated soils while maintaining or even improving soil functions, but its efficiency in agricultural soil practical application still needs to be enhanced. Here, we constructed a biochar-bacteria (2B) partnership in which biochar provides high nutrition and diverse sorption sites. Using the 2B system, Cd immobilization effectiveness and the underlying mechanism were examined along with the soil properties and soil functions. Results showed that compared to the single biochar and ureolytic bacteria systems, soil Cd mobility was reduced by 23.6% and 45.8% through co-precipitating with CaCO3 as otavite (CdCO3) in the 2B system, whereas soil fertility, bacterial diversity, and richness increased by 11.7–90.2%, 5.4–16.1%, and 6.8–54.7%, respectively. Moreover, the abundances of Proteobacteria and Firmicutes were enhanced in the 2B system. Notably, Sporosarcina and Bacillus (Firmicutes genus) that carry the ureC gene were boosted in the system, further implicating the microbiological mechanism in reducing Cd migration and its bioavailability in soil. Overall, the constructed 2B system was efficient in soil Cd immobilization by strengthening the ureolytic bacteria growth and their nutrient supply in the bacteria-rich soil ecosystem.
Highlights
-
A biochar-bacteria (2B) system was constructed based on MICP.
-
Cd was immobilized (23.6–45.8%) in the 2B system.
-
2B system facilitated the formation of insoluble otavite (CdCO3).
-
Bacteria carrying the ureC gene were boosted to lower the Cd mobility.

Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Agricultural soil contamination by cadmium (Cd) causes considerable health problems for humans via food chain exposure (Zhao et al. 2022a). Due to the no-biodegradability, bioaccumulation, and high toxicity, Cd can cause damage to the human brain, bones, and liver (Ali et al. 2020; Fang et al. 2021; Zhao et al. 2022a). High levels of Cd in soils mostly result from anthropogenic activities such as plating, refineries, and mine discharges (Li et al. 2012). To date, numerous physical and chemical methods have been used to remove Cd from the soil, including excavation and landfill, soil washing, soil flushing, chemical precipitation, and chemical oxidation (Jiang et al. 2019; Ali et al. 2020; Liu et al. 2021b). However, most technologies are disruptive to the surrounding environment and more expensive, making them impractical (Li et al. 2012; Guo et al. 2021). Because of their autotrophy for removing Cd from the soil, bio-immobilization of Cd by microbes is more cost-effective and environmentally benign than these methods (Govarthanan et al. 2019; Cuaxinque-Flores et al. 2020; Álvarez-Ayuso et al. 2021; Qiao et al. 2021; Usmani et al. 2021). Furthermore, it is viewed as a feasible approach for agricultural soil remediation because it does not necessitate the suspension of agricultural activities during the restoration process (Ma et al. 2020). As a result, there has recently been a lot of buzz about Cd immobilization in soils using microbes.
Microbially induced calcite precipitation technology (MICP) has been widely used in the bio-immobilization of heavy metals and is considered an efficient and cost-effective remediation approach (Achal et al. 2012a; Jiang et al. 2019; Naveed et al. 2020; de Oliveira et al. 2021; Graddy et al. 2021; Feng et al. 2022). In the process of MICP, ureolytic bacteria produce urease that hydrolyzes urea into ammonium and carbonate, leading to calcium carbonate (CaCO3) precipitation. Cd2+ which has an ion radius close to Ca2+ might be incorporated into the calcite crystal by replacement of Ca2+, thus making the immobilized Cd hard to release (Zhao et al. 2015) and slowing Cd transport into the environment (Achal et al. 2012a; Han et al. 2020b; Guo et al. 2021). In addition, the ureolytic bacteria hydrolyze urea into ammonium and carbonate associated with pH elevation, leading to stabilization of heavy metals (e.g., Cd) (Fang et al. 2021; Hu et al. 2021; Qiao et al. 2021). Co-precipitation of heavy metal with carbonate minerals in the MICP process is the dominant mechanism for heavy metals immobilization (Jiang et al. 2019), and the hydrolysis of the substrate via enzymes secreted by ureolytic bacteria is the core reaction of precipitation (Han et al. 2020b; Qiao et al. 2021). Ureolytic bacteria indigenously exist in the natural environment (Jiang et al. 2019; Chung et al. 2020; Han et al. 2020b), and the hydrolysis of urea by bacteria was 1014 times faster in the MICP process than the natural chemical decomposition of urea (de Oliveira et al. 2021). Besides, in the MICP process, ureolytic bacteria do not need external energy for self-growth (Jiang et al. 2019; de Oliveira et al. 2021; Guo et al. 2021). Thus, MICP has been proven to be a potential green, effective and environmental method for soil remediation with ecological benefits (Guo et al. 2021).
MICP process plays a central role in the N cycle because the reaction of MICP produces an amount of NH4+ and NH3 (Han et al. 2020b; Zhao et al. 2022b), which are beneficial for maintaining soil fertility. Therefore, MICP is a promising pathway for agricultural soil remediation (Qiao et al. 2021). Nowadays, MICP has been broadly employed in engineering, such as sand consolidation, soil stabilization, heavy metal passivation, crack healing in cement-based materials, CO2 sequestration, erosion mitigation, and dust suppression (Achal et al. 2012b; Phillips et al. 2016; Cuaxinque-Flores et al. 2020; Fang et al. 2021). However, the practical application of MICP for agricultural soil remediation is very limited because it may make soil coagulation, enhance mineral crystallization, increase soil strength, and reduce porosity and permeability (Jiang et al. 2019; Chung et al. 2020; Cuaxinque-Flores et al. 2020; Proudfoot et al. 2022), which may affect the soil properties, and tend to be disruptive to soil functions, thus not conducive to soil cultivation. Besides, contaminated soil usually suppresses microorganism growth because of the nutritional deficiency and high toxicity of pollutants (Ma et al. 2020). Thus, we need to develop a feasible strategy to overcome these defects so that the MICP can be highly efficient for heavy metal remediation for agricultural soils.
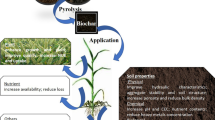
Biochar has high internal and external porosity which can conserve plentiful nutrients, thus making itself an efficient matrix for soil amelioration and bacteria thrive (Phillips et al. 2022; Ok et al. 2021; Woods et al. 2006; Ali et al. 2020). In addition, biochar can serve as a platform for bioremediation (Chen et al. 2020). Therefore, the combination of biochar and bacteria can be supportive for both plant growth and Cd immobilization (Li et al. 2022). Although Cd bio-immobilization using MICP has been studied extensively (Zhu et al. 2016; Do et al. 2020; Fang et al. 2021), no research has been conducted on the combined effect of biochar and MICP on soil fertility, soil functions, and Cd immobilization. Therefore, we combined bacteria and biochar as the prevalent soil ameliorant to develop a simple strategy to overcome the disadvantages of MICP, and modify the MICP-based Cd bio-immobilization pathway.
Based on the theory of MICP, we constructed a biochar-bacteria (2B) system, and verified its high efficiency in Cd immobilization in soils, together with the effects on soil properties and functions. In particular, Cd-contaminated paddy soil was used in this study. Ureolytic bacteria (i.e., Sporosarcina pasteurii), and 0 or 1% biochar (w/w) were applied to the soil, and the mixtures were incubated for 20 days. Finally, we also used the 16S rRNA method to evaluate the short-term effects on soil microbial community structure to make sure that there are no adverse effects on critical soil functions.
2 Materials and methods
2.1 Ureolytic bacteria cultivation
A ureolytic bacterium Sporosarcina pasteurii (S. pasteurii) was used for Cd bio-immobilization because it can produce a large amount of urease (de Oliveira et al. 2021; Fang et al. 2021). S. pasteurii was purchased from China General Microbiological Culture Collection Center. The ureolytic bacterium was cultivated in standard 250 mL Erlenmeyer culture flasks containing 125 mL of Tris-YE growth medium (20 g L−1 yeast extract, 10 g L−1 (NH4)2SO4, 15 g L−1 Tris, and 1 g 24 mL−1 urea) (Liu et al. 2021a). The bacterial cultures were aerobically incubated at 30 °C for 48 h with shaking (130 rpm). The optical density was then measured using UV–visible spectroscopy (V-730, Japan) to assess bacterial growth. The intensity of the bacterium was 1.52 × 108 cell mL−1 for further use.
2.2 Soil and biochar preparation and characterization
The Cd-contaminated soil used for the incubation experiment was collected from the top layer (0–20 cm) of a paddy field located in Mianzhu, China (31°25′ N, 104°15′ E). Soil samples were air-dried, milled, and sieved to < 2 mm for incubation and characterization.
Biochar was derived using rice straw by pyrolysis. The raw material was oven-dried at 60 °C until a constant weight was obtained, ground to pass through a 0.15-mm sieve, and then pyrolyzed at 300 °C in a muffle furnace for 2 h with limited air. To keep bacteria thriving, the temperature of pyrolysis was 300 °C when the polysized biochar has a large number of functional groups. The biochar was then cooled and ground to pass through a 100-mesh size before use. The properties of soil and biochar are shown in Table 1.
2.3 Experiment setup
To prevent MICP from negatively affecting the soil properties and disrupting the soil environment, Sporosarcina pasteurii (S. pasteurii) strain was used for the bioremediation of Cd in soil, and biochar was used to maintain soil fertility and function. The S. pasteurii and biochar system was referred to as the biochar-bacteria (2B) system. We directly applied extra strain and biochar to the polluted soil for creating a simple and workable pathway. The steps of the experiments are listed below.
The incubation experiment was conducted in beakers and each beaker contained 100 g of air-dried soil. After that, 0.694 mg urea and 1.25 mg CaCl2 were added to all soils to activate the MICP process (Li et al. 2014). One gram of biochar (1%, w/w) and 10 mL of bacterium solution were added to the soil and thoroughly mixed (BSW), but no bacteria were introduced to the controlled soil (BCK). Additionally, soil with 1% biochar (SW) and soil with no amendment (CK) were included. Each treatment was conducted in triplicate. The experiment was operated in an incubator under 25 °C with 12/12 h day/night light. During the incubation, soil samples were watered every day to keep a 60% water-holding capacity. After 20 days of incubation, soil samples were collected and then air-dried or kept at −4 °C or −80 °C before analyses. Air-dried soil was used for chemical analysis; fresh soils kept at −4 °C were used for the determination of ammonium (NH4+-N), nitrate (NO3−-N), and enzyme activities; and fresh soils kept at −80 °C were used for DNA extraction.
2.4 Soil property analyses
0.01 M CaCl2 was used to extract soil-soluble Cd (Lu et al. 2017). The speciation of the Cd fractions was determined using a modified Community Bureau of Reference (BCR) sequential extraction method, which defines Cd in four chemical forms, i.e., acid exchangeable fraction, reducible fraction, oxidizable fraction, and residual fraction (Rauret et al. 1999; Wan et al. 2017). HAC, NH2OH·HCl, H2O2, and HCl: HNO3 (V/V, 7.0:2.3) were used as chemical reagents for each batch of extractions. Cd concentration in the extracts of each step was determined with inductively coupled plasma mass spectrometry (ICP-MS, 7700, PerkinElmer, USA). To determine the crystalline phase of precipitated samples, fresh soil samples were freeze-dried, and then characterized by X-ray diffractometer (Bruker, Germany).
Soil pH was measured in a 1:25 soil: water (w/v) ratio using a pH meter. Organic matter (OM) was quantified by the K2Cr2O7 wet oxidation method (Walkley et al. 1934). Total nitrogen (TN) was measured using Kjeldahl method (Kammann et al. 2015). Total phosphorus (TP) was estimated by colorimetry procedure after H2SO4-HClO4 digestion (Murphy et al. 1962). Total potassium (TK) was measured using a Flame photometer (JK-FP6510, China) after H2SO4-HClO4 digestion. NH4+-N and NO3−- N concentrations were extracted using 2 M KCl at 1:25 soil: water (w/v) ratio by shaking at 150 r min−1 at 25 °C for 45 min, and then were measured through a continuous flow analyzer (SAN + + , Holland) (Bello et al. 2021). Available phosphorus (AP) was estimated by colorimetry procedure after 0.5 M NaHCO3 extraction (Murphy et al. 1962). Available potassium (AK) was extracted by 1 M NH4OAC (pH = 7.0) and then measured using a flame photometer (JK-FP6510, China).
The activities of soil enzymes (i.e., urease, dehydrogenase, and phosphatase) were evaluated to reflect soil qualities and functions. Urease activity was measured by indophenol blue method (Kandeler et al. 1988), and the activity of urease was expressed as mg g−1 24 h−1. Dehydrogenase activity was measured spectrophotometrically at 492 nm by triphenylformazan (TPF) method, and the result was expressed as μg TPF g−1 h−1 (Benefield et al. 1977). Phosphatase activity was measured spectrophotometrically at 400 nm (van Aarle et al. 2010), and the result was expressed as mg g−1 24 h−1.
2.5 16S rRNA
Soil total DNA was extracted from fresh soil using a DNA extraction Kit (E.Z.N.A.® soil DNA Kit, Omega Bio-Tek, Norcross, GA, U.S.). Soil DNA was then amplified. 16S rRNA gene was amplified with primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) by thermocycler PCR system (GeneAmp 9700, USA). Amplification was performed in triplicate using 20 μL mixtures, which contained 4 μL PCR buffer, 2 μL of 2.5 mM dNTPs, 0.8 μL of each primer (5 μM), 0.4 μL of FastPfu Polymerase, and 10 ng of template DNA. The PCR amplification conditions were: 3 min of denaturation at 95 °C, followed by 27 cycles of 30 s at 95 °C and 30 s for annealing at 55 °C, 45 s for elongation at 72 °C, and a final extension at 72 °C for 10 min. The PCR products were conditioned for sequencing through Illumina's high-throughput sequencing platform. Alpha diversity and richness indices were calculated to estimate the microbial diversity within an individual sample.
2.6 QA/QC and data analysis
For quality control, sample blanks and standard reference materials (GBW07428) were included when determining Cd concentration in the soil samples. The recovery rate from standard materials for Cd was 80–120%. Statistical analysis was performed using IBM SPSS Statistics 22.0 software. Data were analyzed by two-way analysis of variance ANOVA and the means were compared by Duncan method at the 0.05 level. Principal component analysis (PCA) and redundancy analysis (RDA) of the correlations between the environmental factors and the microbial groups were performed using Canoco 4.5 software.
3 Results
3.1 Effect of different treatments on Cd mobility
Compared with single biochar and ureolytic bacteria systems, using the 2B system reduced CaCl2 extractable Cd in soil by 23.6% and 45.8% (Fig. 1A). The notable decrease in the reducible Cd (F = 19.393, p = 0.002) and increase in the residual Cd (F = 6.049, p = 0.039) were examined after adding bacteria and biochar (Fig. 1B), respectively. When compared to the single ureolytic bacteria system, the 2B system reduced reducible Cd by 14.9% while increasing residual Cd by 16.7% (p < 0.05).

CaCl2-Cd concentration (A) and the fraction of Cd (B) in soils between the treatments. CK: control; SW: soil treated with 1%-biochar; BCK: soil treated with S. pasteurii; BSW: soil treated with S. pasteurii and 1%-biochar. Error bars represent the standard deviation of the means (n = 3)
As illustrated in Fig. 2, the calcite peaks (31.1°) were observed in all soils. Other crystalline polymorphs of calcium carbonate such as vaterite and aragonite were also present in all soil samples. The peaks at 23.4°, 36.4°, 43.8°, 61.2° and 72.9° in samples confirmed that Cd existed in soil primarily as CdCO3. Calcite (22.3°) and CdCO3 (43.8° and 61.2°) were confirmed to be present in all soils, with more calcite and CdCO3 showing in BSW than in other treatments.

XRD patterns of the soil samples. CK: control; SW: soil treated with 1%-biochar; BCK: soil treated with S. pasteurii; BSW: soil treated with S. pasteurii and 1%-biochar
3.2 Effect of different treatments on soil properties and enzymatic activities
Bacteria addition increased soil pH (F = 8.849, p = 0.020). Both biochar and bacteria increased soil OM (Fbiochar = 8.344, p = 0.020; Fbacteria = 6.644, p = 0.022), TP (Fbiochar = 13.889, p = 0.010; Fbacteria = 5.763, p = 0.049), and NH4+-N (Fbiochar = 11.213, p = 0.010; Fbacteria = 874.687, p = 0.000). Compared with single biochar and ureolytic bacteria systems, the 2B system enhanced TK (F = 9.500, p = 0.015).
Notably, SW, BCK, and BSW treatments caused a dramatic increase of NH4+-N (82.5%, 98.0%, and 98.3%) as compared with CK. Biochar addition increased NO3−-N content (F = 33.942, p = 0.000), while bacteria addition significantly reduced NO3−-N (F = 56.914, p = 0.000). Biochar addition increased AK (F = 94.994, p = 0.000), while it did not affect AP. However, CEC and TN did not significantly differ between the treatments.
Bacteria addition reduced urease by 81.1% and 25.9% in non-biochar and biochar soils (F = 48.551, p = 0.000), respectively (Fig. 3). Biochar addition inhibited dehydrogenase activity (F = 13.728, p = 0.006), while bacteria addition increased dehydrogenase activity (F = 26.656, p = 0.001). Biochar significantly increased phosphatase activity (F = 11.210, p = 0.010). However, bacteria addition and the 2B system did not affect phosphatase activity.

Soil enzyme activities in soils under different treatments. CK: control; SW: soil treated with 1%-biochar; BCK: soil treated with S. pasteurii; BSW: soil treated with S. pasteurii and 1%-biochar. Error bars represent the standard deviation of the means (n = 3)
3.3 Effect of different treatments on soil bacterial diversity and community
The alpha diversity of bacteria is shown in Table 2. As compared with CK, BCK reduced Chao1, Faith_pd, Shannon, and Simpson by 48.7%, 38.2%, 23.5%, and 2.3% (p < 0.05), respectively. Both SW and BCK significantly decreased Shannon (15.7–23.5%) and Simpson (2.3–2.7%) as compared with CK (p < 0.05). As compared with single biochar and ureolytic bacteria systems, the 2B system had higher Chao1 (6.8–54.7%), Faith_pd (3.4–42.0%), and Shannon (5.4–16.1%).
The PCA plot showed that soil bacterial beta diversity was significantly influenced by biochar and bacteria addition (Fig. 4A). RDA analysis was performed for all soil factors and the top 14 phyla. Specifically, urease, TP, and NH4+-N were the main factors shifting bacteria community, explaining 47.5% (F = 8.0, p < 0.05), 42.4% (F = 6.6, p < 0.05) and 41.9% (F = 6.5, p < 0.05) of the total variation, respectively (Fig. 4B).

The PCA plots of the beta diversity of soil bacteria (A) and redundancy analysis (RDA) of the correlations between bacterial communities and environmental parameters (B). CK: control; SW: soil treated with 1%-biochar; BCK: soil treated with S. pasteurii; BSW: soil treated with S. pasteurii and 1%-biochar
3.4 Effect of different treatments on soil bacterial community
The dominant phyla in soil under different treatments were Proteobacteria (28.5–63.4%), Actinobacteria (11.3–34.1%), Firmicutes (3.0–19.3%), Chloroflexi (2.4–11.5%), and Gemmatimonadetes (1.9–7.4%), which accounted for more than 80% of the total (Fig. 5A). The abundances of Proteobacteria and Firmicutes significantly increased by 27.0–93.0% (F = 50.244, p = 0.000) and 10.9–45.0% (F = 10.267, p = 0.013) in biochar treated soil as compared with biochar-free soil. The relative abundance of Firmicutes in bacteria-treated soils was 3.4–4.5 times higher than that in biochar-free soils (F = 149.922, p = 0.000). Actinobacteria (F = 31.873, p = 0.000) and Chloroflexi (F = 23.244, p = 0.001) were significantly reduced after adding bacteria. Besides, Firmicutes (31.0–77.8%), and Nitrospirae (25.7–68.5%) showed significant increases in the 2B system as compared with single biochar and ureolytic bacteria systems.

Relative abundance of bacterial community composition at the phylum (A) and genus (B) levels. CK: control; SW: soil treated with 1%-biochar; BCK: soil treated with S. pasteurii; BSW: soil treated with S. pasteurii and 1%-biochar. Error bars represent the standard deviation of the means (n = 3)
These genera dominated the community with Bacillus composing 2.0–7.5% of the total community, followed by Sporosarcina at 0.2–6.4%, Lysobacter at 2.0–5.9%, Chelativorans at 0–10.7%, and Arthrobacter at 0.3–6.2%, respectively. The relative abundances of Bacillus (37.7–62.6%), Sporosarcina (22.6–97.8%), Arthrobacter (23.0–89.5%), and Nitrospira (24.4–64.5%) were much more in 2B system than in the single biochar and ureolytic bacteria systems.
4 Discussion
MICP is known as an effective remediation strategy for remediating heavy metal contaminated soil (Zhu et al. 2016; Han et al. 2020; Zhao et al. 2022b). However, the practical application of MICP for agricultural soil remediation is very limited because it reduces soil ecosystem function. To address the issue that the practical application of MICP may affect the soil properties and tend to be disruptive to the soil environment, we constructed a biochar-bacteria (2B) system based on the theory of MICP and verified the efficiency of the 2B system in Cd immobilization in soils, together with the effects on soil properties and functions.
4.1 The efficiency of the 2B system in Cd immobilization
When compared with single biochar and ureolytic bacteria systems, the 2B system significantly reduced CaCl2 extractable Cd in soil by 23.6% and 45.8%, implying that the 2B system reduced soluble Cd. Moreover, using the 2B system can reduce reducible Cd while increasing residual Cd, thus converting soil Cd from reducible to residual fraction (Ma et al. 2020). The data indicated that Cd was reduced to a state with lower mobility, and stable mineralization occurs with soluble Cd becoming inherently stable phases, which can be explained by the biochar promoting MICP process and leading to Cd mineralization. The 2B system provided more nutrients for carbonate precipitation, leading to higher efficiency for Cd immobilization (Li et al. 2022). Besides, Cd can inhibit bacteria growth even at low concentrations (Chen et al. 2020), while biochar addition protected bacteria against Cd (de Oliveira et al. 2021), thus promoting MICP. Our study was consistent with the findings of Li et al. (2022), who reported that biochar addition effectively immobilized Cd in soil when using strain TZ5 as immobilizing bacteria. Therefore, the 2B system was able to immobilize a larger amount of soluble Cd into insoluble Cd through carbonate precipitation.
Detailed XRD patterns of the soil samples were sufficient enough to confirm the carbonate precipitation and mineral formation (Fig. 2). The calcite peaks (31.1°) were observed in all soils, indicating that MICP occurred. The peaks at 23.4°, 36.4°, 43.8°, 61.2° and 72.9° in samples suggested that the MICP process immobilized Cd mainly in the form of CdCO3 and S. pasteurii was able to sequester Cd along with calcium carbonate biomineralization (Zhao et al. 2022b). The formation of CdCO3 was caused by the isomorphic substitution of Ca by Cd (Zhu et al. 2016; Chen et al. 2019). Cd incorporation into biologically synthesized calcite was also confirmed in other studies (Zeng et al. 2021; Lyu et al. 2022). Calcite (22.3°) and CdCO3 (43.8° and 61.2°) were presented in all soils, with more calcite and CdCO3 showing in BSW than in other treatments. This suggested that the 2B system was successful in promoting bioremediation, accelerating Cd precipitation, and lowering soluble Cd in soil (Peng et al. 2020).
4.2 Performance of the 2B system in regulating soil properties
Soil properties are imperative for evaluating agricultural soil remediation performance. Meanwhile, the bioremediation process is strongly affected by soil properties (Usmani et al. 2021). Thus, soil properties after bioremediation should be understood (Table 3). Soil OM (11.7–12.9%), TP (13.7–15.2%), TK (30.1–67.7%), and NH4+-N (18.2–90.2%) were significantly improved in the 2B system, as compared with single biochar and ureolytic bacteria systems, which confirmed that the 2B system significantly improved the soil fertility of Cd-contaminated soil. Soil properties such as OM, TP, and TK can be enhanced by biochar addition, which has been extensively studied in recent years (Agegnehu et al. 2016b; Yan et al. 2022; Bello et al. 2021). Because biochar is a carbon-rich material, it is not surprising thats the 2B system had higher OM, which was consistent with previous reports (Agegnehu et al. 2016a; Plaza et al. 2016). 2B system also significantly increased TP but did not affect AP, which may account for the lower availability of P within biochar. Previous studies have demonstrated that after pyrolysis at > 200℃, soluble P in biochar was reduced due to the formation of Ca-Mg-P crystallization (Cao et al. 2010; Bekiaris et al. 2016). Unlike P, the availability of K was not decreased after pyrolysis as K is the most mobile element, therefore, the 2B system increased TK.
Further, bacteria addition increased soil pH and NH4+-N, while significantly reducing NO3−-N, which supported the assumption that the MICP process was enhanced by bacteria addition. Ureolytic bacteria hydrolyze urea into ammonium associated with pH elevation, which has been previously described (Fang et al. 2021; Hu et al. 2021; Qiao et al. 2021). Similarly, NH4+-N increased in soils with biochar addition as compared with biochar-free soil, indicating that biochar also promoted the MICP process. Biochar addition increased NO3−-N probably due to its high absorption capacity. Notably, SW, BCK, and BSW treatments caused a dramatic increase of NH4+-N as compared with CK, indicating that all treated soils can stimulate the MICP process and produce NH4+-N, with the 2B system being more successful at doing so. Besides, the higher levels of NO3−-N (60.7%) in the BSW treatment compared to the BCK treatment suggested that the 2B system enhanced NH4+-N content and generated a significant amount of NO3−-N, which encouraged nitrification. Overall, the 2B system was shown to be more suited than single biochar and ureolytic bacteria systems for bioremediation.
4.3 Performance of the 2B system in maintaining soil functions
Urease, dehydrogenase, and phosphatase are extracellular enzymes and are involved in soil biochemical reactions and nutrient cycles such as N, C, and P cycles, which reflect soil functions (Ma et al. 2020; Álvarez-Ayuso et al. 2021). Therefore, the activities of urease, dehydrogenase, and phosphatase after bioremediation were analyzed.
Urease, a key enzyme in MICP, involves the urea hydrolysis to produce ammonia that leads to Cd immobilization (Li et al. 2014; Álvarez-Ayuso et al. 2021), which is dominantly secreted by ureolytic bacteria (Li et al. 2022). In our study, bacteria addition significantly reduced urease (Fig. 3), which was consistent with previous (Ma et al. 2020; Tu et al. 2020). The decrease in urease was probably due to the consumption of urease after bioremediation. After the addition of biochar, the reduced percentage of urease gradually decreased, suggesting the adverse effects can be mitigated by adding biochar. The data demonstrated that the MICP process was improved by adding biochar to the soil, which was consistent with previous findings of Li et al. (2022), who used corncob as an additive and found that urease activity was improved by corncob addition.
A strong negative relationship between urease and CaCl2-Cd (r = −0.512, p < 0.05) implied that urease was involved in the MICP process and Cd immobilization. As mentioned above, the 2B system led to a lower reducible Cd, but a higher residual Cd content. The data also supported the assumption that biochar promoted the passivation of Cd by stimulating ureolytic microbes to create urease and CO32−, which then converted soluble Cd to stable Cd (Zhu et al. 2016; Peng et al. 2020). Using the 2B system, the concentration of urease in the soil released from bacteria was significantly increased, increasing carbonate content and converting solution soluble Cd to state Cd (Fig. 1). Besides, urease is not only related to Cd immobilization but also affects the N cycle (Peng et al. 2020). The increase of NH4+-N in soil could support this point.
Dehydrogenase is an important indicator for assessing soil microbial activation (Peng et al. 2020). Bacteria addition increased dehydrogenase, which was mainly due to a higher number of ureolytic bacteria, and then promoted the MICP process. MICP process led to the release of OH− and then increased pH, increasing dehydrogenase activity (Zhu et al. 2016; Peng et al. 2020). However, biochar addition inhibited the bacteria from secreting dehydrogenase, which corresponded to the previous study (Peng et al. 2020). The decrease in dehydrogenase was likely caused by biochar absorption, which decreased the amount of substrate that was available for bacteria to release dehydrogenase (Li et al. 2022).
Phosphatase catalyzes organic phosphomonoesters' hydrolysis to form soluble P (Li et al. 2022). Biochar addition significantly increased phosphatase, suggesting that biochar addition was beneficial to the P cycle. Nonetheless, we did not observe an increase in AP in biochar-treated soil (Table 1), which might be attributed to Cd chelation. A previous study reported that soil soluble P in the soil can chelate heavy metals to immobilize form (Li et al. 2022), thus depletion of soil AP seems to be the primary reason for the limited increase of AP. The increase of residual Cd in biochar-treated soil (Fig. 1B) can support the point. In addition, as mentioned above, soluble P in biochar was limited, leading to a negligible change in AP.
4.4 Diversity of bacteria to soil factors
The alpha analysis reflects the richness and diversity of bacterial communities (Chen et al. 2019). Shannon and Simpson were significantly lower in SW and BCK treatments compared to CK, suggesting that bacterial richness and diversity were decreased after the MICP process even in biochar and bacteria-treated soils, which was consistent with previous findings (Peng et al. 2020; Lyu et al. 2022). The addition of biochar and bacteria triggered the MICP process, promoted the growth of specific taxonomic groups of bacteria such as ureolytic bacteria, and thus decreased the diversity and richness of bacteria in the soil. Further, as compared with single biochar and ureolytic bacteria systems, the 2B system had higher Chao1, Faith_pd, and Shannon, indicating higher bacterial diversity and richness. It also showed that, compared with CK, α-diversity indices (i.e., Chao1, Faith_pd, and Shannon) were reduced in BCK treatment, while they were increased in BSW treatment as compared with BCK (p < 0.05). The data indicated that bacterial diversity and richness were decreased after the MICP process (Peng et al. 2020; Lyu et al. 2022), while biochar addition could alleviate this negative effect, and then increased microbial community diversity and richness, which was consistent with previous results (Mierzwa-Hersztek et al. 2016; Chen et al. 2019; Jiang et al. 2019; Álvarez-Ayuso et al. 2021; Li et al. 2022). The increases in microbial diversity and richness in the 2B system might be explained in terms of the following aspects: (1) habitat effect and (2) nutrient supply (Li et al. 2022). Biochar had a large specific surface area, which was conducive to the attachment of microorganisms (Li et al. 2022). The high number of microbes secreted more urease to promote the MICP process, which meant biochar induced the increase of bacterial diversity and was beneficial for Cd immobilization (Fig. 1) (Chen et al. 2019). In addition, biochar contains sufficient nutrient elements, which might be involved in the microbially induced carbonate precipitation (Li et al. 2022), thus promoting Cd immobilization. Furthermore, soil microbial biomass directly affects soil enzymes, thus the enhancement of richness and diversity in the 2B system resulted in the improvement of activities of soil enzymes, and therefore there were no adverse effects on soil functions. Together, the 2B system improved bacterial variety and richness, enhancing soil functions and preserving soil production. Additionally, this stimulated the MICP process and immobilized Cd.
The PCA plot showed that soil bacterial beta diversity was significantly influenced by biochar and bacteria addition (Fig. 4A), which was consistent with the changing trends in the composition and alpha diversity of soil bacterial communities. Soil physicochemical variables have been previously described to be the major factors for soil microbial dynamics and structure (Bello et al. 2021; Hu et al. 2021). The bacterial variations in soil samples were caused by the nutrients cascade and competition as well as habitat supply (Ma et al. 2020; Bello et al. 2021). RDA analysis showed that urease, TP, and NH4+-N were the main drivers shaping and controlling bacteria species. This result supported the findings of Hu et al. (2021), who reported that available N and urease showed a significant correlation with bacterial composition and diversity in soils.
4.5 Community structure of bacteria
The bacterial community had significant changes (Fig. 5A), indicating its sensitivity to environmental conditions. The relative abundance percentage analysis of microorganisms showed that Proteobacteria, Actinobacteria, and Firmicutes were the most dominant phyla in all samples, which was consistent with previous reports (Hu et al. 2021; Koner et al. 2022).
Proteobacteria can survive in harsh conditions and become dominant in heavy metal contaminated agricultural soil since they can tolerate higher heavy metal concentrations and utilize multiple C and N sources in a complex survival mode, which has been reported previously (Hu et al. 2021). As such, it was not surprising that Proteobacteria had the highest abundance in our study. The abundance of Proteobacteria significantly increased in biochar-treated soil as compared with biochar-free soil, which may be related to its habit. Proteobacteria prefer a eutrophic environment (Hu et al. 2021). Biochar contains plentiful nutrients (C, N, P, and K), which exerted beneficial effects on Proteobacteria growth.
Firmicutes can adapt to harsh conditions, and most ureolytic microorganisms belonged to Firmicutes (Yin et al. 2021; Koner et al. 2022). In our study, the relative abundance of Firmicutes in all soils was 3.2–16.5%, indicating that ureolytic microorganisms could survive in an environment with Cd contamination, which had the potential for the use of Cd biomineralization (Hu et al. 2021). Meanwhile, the relative abundance of Firmicutes in bacteria soils was 3.4–4.5 times higher than that in the soils without bacteria, indicating that ureolytic microorganisms were inoculated successfully. Additionally, biochar addition significantly increased Firmicutes abundance by 10.9–45.0%, which implied that biochar addition was an effective way to promote Firmicutes growth, thus promoting heavy metal biomineralization.
Actinobacteria and Chloroflexi were significantly reduced in bacteria-treated soil, indicating that the MICP process had adverse effects on soil microorganisms, which has been reported (Lyu et al. 2022). Besides, Nitrospirae showed significant increases in the 2B system as compared with single biochar and ureolytic bacteria systems, suggesting that the 2B system played important role in the N cycle.
4.6 Construction of the biochar-bacteria (2B) system and the immobilization mechanism
To further reveal the mechanism of Cd immobilization by MICP, the genera ranked in the top 19 in terms of abundance were analyzed (Fig. 5B). The result showed that Bacillus was the dominant genus, followed by Sporosarcina, Lysobacter, Chelativorans, and Arthrobacter. Species identified as significantly positively correlated with NH4+-N (r = 0.953, p < 0.01) and negatively correlated with NO3−-N (r = − 0.509, p < 0.05) belonged to phylum Firmicutes. Most of the ureolytic genera that can produce urease belong to this phylum in soil (Hu et al. 2021). Therefore, the ureolytic genera in Firmicutes such as Bacillus and Sporosarcina can carry the ureC gene and have a high urease-producing ability (Graddy et al. 2021; Yin et al. 2021), which can immobilize heavy metals by secreting urease that decomposes urea and produce CO32− that binds with Cd to form precipitates, resulting in the Cd immobilization in soil (Han et al. 2020a; Hu et al. 2021). In the 2B system, the abundances of Firmicutes were enhanced, which again suggested that the 2B system stimulated ureolytic genera growth and accelerated the MICP process. It is worth noting that species Bacillus and Sporosarcina had significant positive correlations with NH4+-N with the correlation coefficients being 0.871 and 0.986 (p < 0.01), implying that these genera were important functional genera that could be involved in the MICP process. Bacillus and Sporosarcina were the dominant genera in all soils, indicating that ureolytic genera were inoculated successfully.
Sporosarcina pasteurii (S. pasteurii) belongs to the Sporosarcina genus. Sporosarcina dominated the community in soils treated with bacteria, making up 4.1–6.4% of the total community, which was 22.7–46.7 times more than it was in soils not treated with bacteria, demonstrating the efficacy of the inoculation of the bacteria. Cd mineralization by S. pasteurii caused steady mineralization, which lowered the bioavailability of Cd in agricultural soil and reduced Cd toxicity. It was also obvious from Fig. 5B that Sporosarcina and Bacillus had high abundances in all soils, and they were significantly increased in the 2B system when compared with single biochar and ureolytic bacteria systems. Thus, it can be concluded that the 2B system not only enhanced Sporosarcina but also favored other ureolytic microorganism growth, such as Bacillus, which was consistent with previous studies (Chen et al. 2019; Li et al. 2022). In addition, the Bacillus genus is regarded as metal resistant bacteria (Ma et al. 2020), which can promote P solubilization (Pattnaik et al. 2020). Soil soluble P can chelate Cd to immobilize Cd (Ma et al. 2020), thus reducing Cd availability. Thus, the increase of Bacillus was also favorable for Cd immobilization.
Besides, Arthrobacter is also recognized as a Cd-resistant bacterium (Ma et al. 2020), which was also increased in the 2B system when compared with single biochar and ureolytic bacteria systems. Further, Nitrospira belongs to the Nitrospirae genera. Both Nitrospira and Nitrospirae are involved in nitrification because they are correlated positively with NO3−-N with values of correlation coefficient of 0.526 (p < 0.01) for Nitrospirae and 0.517 (p < 0.01) for Nitrospira, respectively. Increases in the abundance of Nitrospira and Nitrospirae in the 2B system suggested that biochar promoted nitrification.
Overall, the 2B system increased bacterial richness and diversity and increased Sporosarcina and Bacillus abundance, thus promoting the MICP process. Thus, the constructed 2B system is an easy and feasible pathway to remediate the Cd-contaminated soil. Besides, the 2B system also mediated the N cycle by altering Nitrospira growth. The overall reactions involved in biomineralization in the 2B system mainly included the following processes: (1) urea hydrolysis produced NH4+-N and CO32−; (2) Cd coprecipitated with CO32− by replacing Ca; and (3) NO3− was produced. Using the proposed system, Cd can be transformed into carbonate-bound and this process was enhanced (Ma et al. 2020). In the process, a large amount of NH4+-N led to NO3−-N production and promoted nitrification.
5 Conclusion
Based on the theory of MICP, we constructed a biochar-bacteria (2B) system, and verified its high efficiency in Cd immobilization in soils, together with the effects on soil properties and functions. Soil Cd mobility was reduced through co-precipitating with CaCO3 as otavite (CdCO3) in the 2B system. Using the 2B system, soil fertility, urease, bacterial diversity, and richness were significantly improved. Urease, TP, and NH4+-N were the main drivers shaping and controlling bacteria species. Moreover, in the 2B system, the abundances of Proteobacteria and Firmicutes were enhanced. Sporosarcina and Bacillus belonging to the Firmicutes genera and carrying the ureC gene were further boosted, explaining the microbiological mechanism in reducing soil Cd bioavailability. Thus, the 2B system promoted the passivation of Cd by stimulating ureolytic microbes to create urease and CO32−, which then converted soluble Cd to stable Cd. Besides, the 2B system enhanced NO3−-N content by increasing Nitrospira growth. The overall reactions involved in biomineralization in the 2B system mainly included the following processes: (1) urea hydrolysis produced NH4+-N and CO32−; (2) Cd coprecipitated with CO32− by replacing Ca; and (3) NO3− was produced. Taken together, the constructed 2B system is efficient in Cd immobilization by strengthening the ureolytic bacteria growth and nutrient supply. However, more research is required to enhance the 2B system's performance in remediating Cd-contaminated agricultural soil as well as its practical use.
Availability of data and materials
The datasets analyzed in the current study are available from the corresponding author upon reasonable request.
References
Achal V, Pan X, Fu Q, Zhang D (2012a) Biomineralization based remediation of As(III) contaminated soil by Sporosarcina ginsengisoli. J Hazard Mater 201–202:178–184. https://doi.org/10.1016/j.jhazmat.2011.11.067
Achal V, Pan X, Zhang D (2012b) Bioremediation of strontium (Sr) contaminated aquifer quartz sand based on carbonate precipitation induced by Sr resistant Halomonas sp. Chemosphere 89:764–768. https://doi.org/10.1016/j.chemosphere.2012.06.064
Agegnehu G, Bass AM, Nelson PN, Bird MI (2016a) Benefits of biochar, compost and biochar-compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci Total Environ 543:295–306. https://doi.org/10.1016/j.scitotenv.2015.11.054
Agegnehu G, Bass AM, Nelson PN, Bird MI (2016b) Benefits of biochar, compost and biochar–compost for soil quality, maize yield and greenhouse gas emissions in a tropical agricultural soil. Sci Total Environ 543:295–306. https://doi.org/10.1016/j.scitotenv.2015.11.054
Ali U, Shaaban M, Bashir S, Gao R, Fu Q, Zhu J, Hu H (2020) Rice straw, biochar and calcite incorporation enhance nickel (Ni) immobilization in contaminated soil and Ni removal capacity. Chemosphere 244:125418. https://doi.org/10.1016/j.chemosphere.2019.125418
Álvarez-Ayuso E, Abad-Valle P (2021) Application of different alkaline materials as polluted soil amendments: A comparative assessment of their impact on trace element mobility and microbial functions. Ecotoxicol Environ Saf 227: 112927. https://doi.org/10.1016/j.ecoenv.2021.112927
Bekiaris G, Peltre C, Jensen LS, Bruun S (2016) Using FTIR-photoacoustic spectroscopy for phosphorus speciation analysis of biochars. Spectrochim Acta A Mol Biomol 168:29–36. https://doi.org/10.1016/j.saa.2016.05.049
Bello A, Wang B, Zhao Y, Yang W, Ogundeji A, Deng L, Egbeagu UU, Yu S, Zhao L, Li D, Xu X (2021) Composted biochar affects structural dynamics, function and co-occurrence network patterns of fungi community. Sci Total Environ 775:145672. https://doi.org/10.1016/j.scitotenv.2021.145672
Benefield CB, Howard PJA, Howard DM (1977) The estimation of dehydrogenase activity in soil. Soil Biol Biochem 9:67–70. https://doi.org/10.1016/0038-0717(77)90063-3
Cao X, Harris W (2010) Properties of dairy-manure-derived biochar pertinent to its potential use in remediation. Bioresour Technol 101:5222–5228. https://doi.org/10.1016/j.biortech.2010.02.052
Chen H, Tang L, Wang Z, Su M, Tian D, Zhang L, Li Z (2020) Evaluating the protection of bacteria from extreme Cd (II) stress by P-enriched biochar. Environ Pollut 263:114483. https://doi.org/10.1016/j.envpol.2020.114483
Chen X, Achal V (2019) Biostimulation of carbonate precipitation process in soil for copper immobilization. J Hazard Mater 368:705–713. https://doi.org/10.1016/j.jhazmat.2019.01.108
Chung H, Kim SH, Nam K (2020) Inhibition of urea hydrolysis by free Cu concentration of soil solution in microbially induced calcium carbonate precipitation. Sci Total Environ 740:140194. https://doi.org/10.1016/j.scitotenv.2020.140194
Cuaxinque-Flores G, Aguirre-Noyola JL, Hernández-Flores G, Martínez-Romero E, Romero-Ramírez Y, Talavera-Mendoza O (2020) Bioimmobilization of toxic metals by precipitation of carbonates using Sporosarcina luteola: An in vitro study and application to sulfide-bearing tailings. Sci Total Environ 724:138124. https://doi.org/10.1016/j.scitotenv.2020.138124
de Oliveira D, Horn EJ, Randall DG (2021) Copper mine tailings valorization using microbial induced calcium carbonate precipitation. J Environ Manage 298:113440. https://doi.org/10.1016/j.jenvman.2021.113440
Do H, Wang Y, Long Z, Ketehouli T, Li X, Zhao Z, Li M (2020) A psychrotolerant Ni-resistant Bacillus cereus D2 induces carbonate precipitation of nickel at low temperature. Ecotox Environ Safe 198:110672. https://doi.org/10.1016/j.ecoenv.2020.110672
Fang L, Niu Q, Cheng L, Jiang J, Yu YY, Chu J, Achal V, You T (2021) Ca-mediated alleviation of Cd2+ induced toxicity and improved Cd2+ biomineralization by Sporosarcina pasteurii. Sci Total Environ 787:147627. https://doi.org/10.1016/j.scitotenv.2021.147627
Feng C, Zhao S, Zong Y, He Q, Winarto W, Zhang W, Utada AS, Zhao K (2022) Microdroplet-based in situ characterization of the dynamic evolution of amorphous calcium carbonate during microbially induced calcium carbonate precipitation. Environ Sci Technol 56:11017–11026. https://doi.org/10.1021/acs.est.1c08858
Govarthanan M, Mythili R, Kamala-Kannan S, Selvankumar T, Srinivasan P, Kim H (2019) In-vitro bio-mineralization of arsenic and lead from aqueous solution and soil by wood rot fungus, Trichoderma sp. Ecotox Environ Safe 174:699–705. https://doi.org/10.1016/j.ecoenv.2019.03.034
Graddy CMR, Gomez MG, DeJong JT, Nelson DC (2021) Native bacterial community convergence in augmented and stimulated ureolytic MICP biocementation. Environ Sci Technol 55:10784–10793. https://doi.org/10.1021/acs.est.1c01520
Guo S, Zhang J, Li M, Zhou N, Song W, Wang Z, Qi S (2021) A preliminary study of solid-waste coal gangue based biomineralization as eco-friendly underground backfill material: Material preparation and macro-micro analyses. Sci Total Environ 770:145241. https://doi.org/10.1016/j.scitotenv.2021.145241
Han H, Cai H, Wang X, Hu X, Chen Z, Yao L (2020a) Heavy metal-immobilizing bacteria increase the biomass and reduce the Cd and Pb uptake by pakchoi (Brassica chinensis L.) in heavy metal-contaminated soil. Ecotoxicol Environ Safe 195:110375. https://doi.org/10.1016/j.ecoenv.2020.110375
Han L, Li J, Xue Q, Chen Z, Zhou Y, Poon CS (2020b) Bacterial-induced mineralization (BIM) for soil solidification and heavy metal stabilization: A critical review. Sci Total Environ 746:140967. https://doi.org/10.1016/j.scitotenv.2020.140967
Hu X, Liu X, Qiao L, Zhang S, Su K, Qiu Z, Li X, Zhao Q, Yu C (2021) Study on the spatial distribution of ureolytic microorganisms in farmland soil around tailings with different heavy metal pollution. Sci Total Environ 775:144946. https://doi.org/10.1016/j.scitotenv.2021.144946
Jiang NJ, Liu R, Du YJ, Bi YZ (2019) Microbial induced carbonate precipitation for immobilizing Pb contaminants: Toxic effects on bacterial activity and immobilization efficiency. Sci Total Environ 672:722–731. https://doi.org/10.1016/j.scitotenv.2019.03.294
Kammann CI, Schmidt HP, Messerschmidt N, Linsel S, Steffens D, Müller C, Koyro HW, Conte P, Joseph S (2015) Plant growth improvement mediated by nitrate capture in co-composted biochar. Sci Rep 5:11080. https://doi.org/10.1038/srep11080
Kandeler E, Gerber H (1988) Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol Fert Soils 6:68–72. https://doi.org/10.1007/BF00257924
Koner S, Chen JS, Hsu BM, Rathod J, Huang SW, Chien HY, Hussain B, Chan MWY (2022) Depth-resolved microbial diversity and functional profiles of trichloroethylene-contaminated soils for Biolog EcoPlate-based biostimulation strategy. J Hazard Mater 424:127266. https://doi.org/10.1016/j.jhazmat.2021.127266
Li H, Song Y, Li Q, He J, Song Y (2014) Effective microbial calcite precipitation by a new mutant and precipitating regulation of extracellular urease. Bioresour Technol 167:269–275. https://doi.org/10.1016/j.biortech.2014.06.011
Li JT, Baker AJ, Ye ZH, Wang HB, Shu WS (2012) Phytoextraction of Cd-contaminated soils: Current status and future challenges. Crit Rev Environ Sci Technol 42:2113–2152. https://doi.org/10.1080/10643389.2011.574105
Li W, Yang Y, Achal V (2022) Biochemical composite material using corncob powder as a carrier material for ureolytic bacteria in soil cadmium immobilization. Sci Total Environ 802:149802. https://doi.org/10.1016/j.scitotenv.2021.149802
Liu P, Zhang Y, Tang Q, Shi S (2021a) Bioremediation of metal-contaminated soils by microbially-induced carbonate precipitation and its effects on ecotoxicity and long-term stability. Biochem Eng J 166:107856. https://doi.org/10.1016/j.bej.2020.107856
Liu Y, Hu C, Li B, Ding D, Zhao Z, Fan T, Li Z (2021b) Subsurface drip irrigation reduces cadmium accumulation of pepper (Capsicum annuum L.) plants in upland soil. Sci Total Environ 755:142650. https://doi.org/10.1016/j.scitotenv.2020.142650
Lu K, Yang X, Gielen G, Bolan N, Ok YS, Niazi NK, Xu S, Yuan G, Chen X, Zhang X, Liu D, Song Z, Liu X, Wang H (2017) Effect of bamboo and rice straw biochars on the mobility and redistribution of heavy metals (Cd, Cu, Pb and Zn) in contaminated soil. J Environ Manage 186:285–292. https://doi.org/10.1016/j.jenvman.2016.05.068
Lyu C, Qin Y, Chen T, Zhao Z, Liu X (2022) Microbial induced carbonate precipitation contributes to the fates of Cd and Se in Cd-contaminated seleniferous soils. J Hazard Mater 423:126977. https://doi.org/10.1016/j.jhazmat.2021.126977
Ma H, Wei M, Wang Z, Hou S, Li X, Xu H (2020) Bioremediation of cadmium polluted soil using a novel cadmium immobilizing plant growth promotion strain Bacillus sp TZ5 loaded on biochar. J Hazard Mater 388:122065. https://doi.org/10.1016/j.jhazmat.2020.122065
Mierzwa-Hersztek M, Gondek K, Baran A (2016) Effect of poultry litter biochar on soil enzymatic activity, ecotoxicity and plant growth. Appl Soil Ecol 105:144–150. https://doi.org/10.1016/j.apsoil.2016.04.006
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36. https://doi.org/10.1016/S0003-2670(00)88444-5
Naveed M, Duan J, Uddin S, Suleman M, Hui Y, Li H (2020) Application of microbially induced calcium carbonate precipitation with urea hydrolysis to improve the mechanical properties of soil. Ecol Eng 153:105885. https://doi.org/10.1016/j.ecoleng.2020.105885
Ok YS, Palansooriya KN, Yuan X, Rinklebe J (2021) Special issue on biochar technologies, production, and environmental applications. Crit Rev Environ Sci Technol 52(19):3375–3383. https://doi.org/10.1080/10643389.2021.1990446
Pattnaik S, Dash D, Mohapatra S, Pattnaik M, Marandi AK, Das S, Samantaray DP (2020) Improvement of rice plant productivity by native Cr(VI) reducing and plant growth promoting soil bacteria Enterobacter cloacae. Chemosphere 240:124895. https://doi.org/10.1016/j.chemosphere.2019.124895
Peng D, Qiao S, Luo Y, Ma H, Zhang L, Hou S, Wu B, Xu H (2020) Performance of microbial induced carbonate precipitation for immobilizing Cd in water and soil. J Hazard Mater 400:123116. https://doi.org/10.1016/j.jhazmat.2020.123116
Phillips AJ, Cunningham AB, Gerlach R, Hiebert R, Hwang C, Lomans BP, Westrich J, Mantilla C, Kirksey J, Esposito R (2016) Fracture sealing with microbially-induced calcium carbonate precipitation: A field study. Environ Sci Technol 50:4111–4117. https://doi.org/10.1021/acs.est.5b05559
Phillips CL, Meyer KM, Garcia-Jaramillo M, Weidman CS, Stewart CE, Wanzek T, Grusak MA, Watts DW, Novak J, Trippe KM (2022) Towards predicting biochar impacts on plant-available soil nitrogen content. Biochar 8:2417. https://doi.org/10.1007/s42773-022-00137-2
Plaza C, Giannetta B, Fernández JM, López-de-Sá EG, Polo A, Gascó G, Méndez A, Zaccone C (2016) Response of different soil organic matter pools to biochar and organic fertilizers. Agr Ecosyst Environ 225:150–159. https://doi.org/10.1016/j.agee.2016.04.014
Proudfoot D, Brooks L, Gammons CH, Barth E, Bless D, Nagisetty RM, Lauchnor EG (2022) Investigating the potential for microbially induced carbonate precipitation to treat mine waste. J Hazard Mater 424:127490. https://doi.org/10.1016/j.jhazmat.2021.127490
Qiao S, Zeng G, Wang X, Dai C, Sheng M, Chen Q, Xu F, Xu H (2021) Multiple heavy metals immobilization based on microbially induced carbonate precipitation by ureolytic bacteria and the precipitation patterns exploration. Chemosphere 274:129661. https://doi.org/10.1016/j.chemosphere.2021.129661
Rauret G, López-Sánchez JF, Sahuquillo A, Rubio R, Davidson C, Ure A, Quevauviller P (1999) Improvement of the BCR three step sequential extraction procedure prior to the certification of new sediment and soil reference materials. J Environ Monit 1:57–61. https://doi.org/10.1039/a807854h
Tu C, Wei J, Guan F, Liu Y, Sun Y, Luo Y (2020) Biochar and bacteria inoculated biochar enhanced Cd and Cu immobilization and enzymatic activity in a polluted soil. Environ Int 137:105576. https://doi.org/10.1016/j.envint.2020.105576
Usmani Z, Kulp M, Lukk T (2021) Bioremediation of lindane contaminated soil: Exploring the potential of actinobacterial strains. Chemosphere 278:130468. https://doi.org/10.1016/j.chemosphere.2021.130468
van Aarle IM, Plassard C (2010) Spatial distribution of phosphatase activity associated with ectomycorrhizal plants is related to soil type. Soil Biol Biochem 42:324–330. https://doi.org/10.1016/j.soilbio.2009.11.011
Walkley A, Black IA (1934) An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38. https://doi.org/10.1097/00010694-193401000-00003
Wan X, Dong H, Feng L, Lin Z, Luo Q (2017) Comparison of three sequential extraction procedures for arsenic fractionation in highly polluted sites. Chemosphere 178:402–410. https://doi.org/10.1016/j.chemosphere.2017.03.078
Woods WI, Falcão NPS, Teixeira WG (2006) Biochar trials aim to enrich soil for smallholders. Nature 443:144. https://doi.org/10.1038/443144b
Yan SH, Zhang SL, Yan PK, Aurangzeib M (2022) Effect of biochar application method and amount on the soil quality and maize yield in Mollisols of Northeast China. Biochar 56:1552. https://doi.org/10.1007/s42773-022-00180-z
Yin T, Lin H, Dong Y, Li B, He Y, Liu C, Chen X (2021) A novel constructed carbonate-mineralized functional bacterial consortium for high-efficiency cadmium biomineralization. J Hazard Mater 401:123269. https://doi.org/10.1016/j.jhazmat.2020.123269
Zeng G, Qiao S, Wang X, Sheng M, Wei M, Chen Q, Xu H, Xu F (2021) Immobilization of cadmium by Burkholderia sp QY14 through modified microbially induced phosphate precipitation. J Hazard Mater 412:125156. https://doi.org/10.1016/j.jhazmat.2021.125156
Zhao C, Fu Q, Song W, Zhang D, Ahati J, Pan X, Al-Misned FA, Mortuza MG (2015) Calcifying cyanobacterium (Nostoc calcicola) reactor as a promising way to remove cadmium from water. Ecol Eng 81:107–114. https://doi.org/10.1016/j.ecoleng.2015.04.012
Zhao D, Wang P, Zhao FJ (2022a) Dietary cadmium exposure, risks to human health and mitigation strategies. Crit Rev Environ Sci Technol. https://doi.org/10.1080/10643389.2022.2099192
Zhao J, Csetenyi L, Gadd GM (2022b) Fungal-induced CaCO3 and SrCO3 precipitation: a potential strategy for bioprotection of concrete. Sci Total Environ 816:151501. https://doi.org/10.1016/j.scitotenv.2021.151501
Zhu X, Li W, Zhan L, Huang M, Zhang Q, Achal V (2016) The large-scale process of microbial carbonate precipitation for nickel remediation from an industrial soil. Environ Pollut 219:149–155. https://doi.org/10.1016/j.envpol.2016.10.047
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 4210070784), and the Key Program of Chengdu Science and Technology Bureau (Grant Nos. 2020-YF09-00023-SN and 2020-YF09-00012-SN).
Author information
Authors and Affiliations
Contributions
MX: Conceptualization, formal analysis, methodology, experimental operation, visualization, writing—original draft. JM: Investigation. XZ: Writing—review and editing. GY: Data curation, writing—review and editing. LL: Writing—review and editing. CC: Analysis, writing—review and editing. CS: Formal analysis, writing—review and editing. JW: Resources, supervision. PG: Writing—review and editing. DG: Writing—review an editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Handling editor: Lukas Trakal.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Xu, M., Ma, J., Zhang, XH. et al. Biochar-bacteria partnership based on microbially induced calcite precipitation improves Cd immobilization and soil function. Biochar 5, 20 (2023). https://doi.org/10.1007/s42773-023-00222-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s42773-023-00222-0