
Abstract
The epithelial to mesenchymal transition (EMT) generates tumor cells with stem cell characteristics and with phenotypes similar to those of cancer stem cells (CSCs). Evidence suggests CSCs are in an intermediate state of EMT expressing reduced levels of E-cadherin and with mesenchymal features including invasiveness associated with metastasis. These findings suggest mechanisms regulating EMT and stemness are closely integrated. Recent reports from several laboratories have identified novel mechanisms regulating EMT and stemness involving epigenetics, microenvironment, and dedifferentiation. Circulating tumor cells (CTCs) have also been shown to have features of EMT, but it is unclear which fraction has the properties of CSCs. EMT characteristics of both CSCs and CTCs are associated with resistance to current clinical treatments, indicating that therapy targeting CSCs in addition to more differentiated tumor cells are required for durable responses. Thus, EMT characteristics of CTCs may prove useful biomarkers for effective therapy for many cancers.
Similar content being viewed by others


Introduction
The epithelial to mesenchymal transition (EMT) is a reversible developmental program responsible for the formation of the tissues and organs of the body whereby nonmotile epithelial cells with strong cell-to-cell adhesion and apical–basal polarity become individual, motile mesenchymal cells with front-to-back polarity (Fig. 1) [1]. Molecular markers of EMT include loss of the epithelial marker E-cadherin and gain of EMT-inducing transcription factors and mesenchymal markers including N-cadherin, fibronectin, and vimentin [2]. EMT is associated with the reorganization of the actin cytoskeleton including loss of cortical actin and gain of filamentous actin. However, the most important marker of the gain of mesenchymal features is the acquisition of motility [3]. Examples of EMT in development include implantation, gastrulation, neural crest formation, and cardiac valve formation. EMT also occurs as a normal physiological response to tissue injury that can become pathological, as seen with tissue and organ fibrosis [4]. Importantly, the developmental program of EMT is reactivated in cancer, such that epithelial cells undergo an EMT, acquiring mesenchymal features that enable them to migrate and invade. This reactivation of the developmental program of EMT in cancer is believed to be responsible for metastasis of the primary tumor to distant sites, which is responsible for most cancer-related deaths. The process of EMT is reversible through a mesenchymal to epithelial transition (MET) in which the motile mesenchymal cells, once reaching their destination, revert to an epithelial phenotype. MET is essential in normal development.

Molecular characteristics of EMT and stemness in development versus cancer. The similar features of EMT in both development and cancer are illustrated. Differences between the maintenance and/or acquisition of stemness in development and cancer are also shown
The primary model explaining tumorigenesis and tumor heterogeneity has been the clonal evolution theory whereby cells acquire changes, genetic or epigenetic, that provide them with a growth advantage. These cells become the dominant clone in the tumor and are responsible for its tumorigenic capacity. More recently, a cancer stem cell (CSC) model has been proposed to explain tumor heterogeneity [5]. Unlike the clonal evolution model, the CSC hypothesis is based on hierarchical organization such that only a small subset of cells in a tumor is responsible for tumorigenesis. CSCs are cancer cells with the stem cell properties of self-renewal and differentiation. CSCs can self-renew to generate more CSCs and can differentiate to generate all the non-CSC progeny in the tumor. Importantly, CSCs do not necessarily originate from transformation of a stem cell. In addition, CSCs can arise from the transformation and dedifferentiation of non-CSCs via genetic or epigenetic changes to non-CSCs. The presence of CSCs has been demonstrated in hematopoietic, breast, pancreatic, and colon cancers. In addition, CSCs have been implicated in several other cancers, but are not likely to be responsible for all cancer types.
EMT Promotes Stem Cell Features and Induces CSCs
A critical topic in the field of EMT and CSCs is the intersection between EMT and stemness. Mani et al. [6, 7] demonstrated that induction of EMT in human mammary epithelial cells by overexpression of the transcription factors Snail or Twist generated stem cell-like cells (Fig. 1). These stem cell-like cells continued to have properties of EMT. Induction of EMT in mammary cells induced a stem cell phenotype similar to CSCs. They went on to show that EMT gives rise to metastatic CSCs. Post-therapy breast tissue was found to be enriched in tumor cells with CSC-like and mesenchymal features [8]. Recent studies by Rhim et al. [9∙] and Abell et al. [10] have shown that CSCs may be a population of cells in an intermediate state of EMT (Fig. 1). These cells express low to moderate levels of the epithelial marker E-cadherin and at the same time, have mesenchymal features. Abell et al. [10] showed that mutant stem cells in an intermediate stage of EMT and claudin-low breast cancer (CLBC) cells with mesenchymal features shared a gene expression signature. These cells were similar to cancer stem cells, maintaining stemness features and tumor-initiating properties, and were highly invasive [11]. Similarly, Rhim et al. [9∙] found that pancreatic ductal adenocarcinoma cells (PDACs) had features of an intermediate stage of EMT, coexpressing both epithelial and mesenchymal markers. They found that PDACs metastasized “early”, before histological evidence of malignancy. Further, circulating PDACs had pancreatic stem cell properties, being CD24/CD44 positive and being able to grow in pancreatosphere assays. Together, these studies suggest that cancer cells passing through EMT acquire self-renewing stem cell traits.
That CSCs may exist in an intermediate stage of EMT suggests mechanisms that regulate EMT and stemness are closely integrated. Recent papers from several laboratories have identified novel mechanisms regulating EMT and stemness that include epigenetic modifications, changes in the microenvironment, and dedifferentiation of non-CSCs to CSCs. Each of these potential mechanisms is discussed below.
Promotion of EMT and Stemness by Epigenetic Changes
An important focus of research into EMT and stem cell features of cancer cells relates to the mechanisms that regulate EMT to promote stemness in CSCs. Several recent publications have demonstrated the power of epigenetic changes to promote EMT and induce stemness in CSCs. Abell et al. [10] identified an epigenetic mechanism controlling EMT in stem cells and CLBC cells with stem cell features. MAP3K4, a MAP3K kinase regulating the JNK pathway, activates the acetyl transferase CBP, promoting acetylation of histone H2BK5 to maintain the epithelial phenotype. Loss of H2BK5 acetylation on a specific set of genes in both stem cells and CLBC resulted in an intermediate state of EMT. One of the genes regulated by H2BK5 acetylation is SMARCD3, a SWI/SNF chromatin-remodeling factor [12]. Knockdown of SMARCD3 in mesenchymal-like basal breast cancer cells partially restored epithelial features of these cells. Further, overexpression of SMARCD3 was sufficient to induce EMT in mammary epithelial cells and to induce a gene-expression signature similar to that of mesenchymal CLBC cells with stemness characteristics, being CD44 and ALDH1-positive and CD24-negative.
In addition to identification of acetyl transferases and chromatin modifiers that regulate the induction of EMT and stemness, recent studies by Song et al. [13] and Taube et al. [14] have demonstrated the role of specific microRNAs (miRs) in the regulation of stemness and EMT in breast cancer. Song et al. [13] showed the ability of miR-22 to promote EMT and stemness. miR-22 silenced the miR-200 family, which controls epithelial maintenance, by targeting the expression of the ten eleven translocation (TET) family of methylcytosine dioxygenases, inhibiting demethylation of the mir-200 promoter. miR-22 expression was sufficient to induce expansion of the mammary stem cell compartment, EMT features, and invasiveness, suggesting it is an initiator of tumor initiation. Overexpression of miR-22 correlated with poor patient outcomes in breast cancer, suggesting that miR-22 may be a viable therapeutic target for breast cancers with EMT and stemness features.
In contrast with EMT-inducing properties of miR-22, Taube et al. [14] has found that miR-203 represses EMT and stemness. Expression of miR-203 in human mammary epithelial cells was repressed by induction of EMT by TGFβ, or by overexpression of EMT-inducing transcription factors Twist or Snail. It was also repressed in CLBC cells and in CD44hi/CD24low stem cell-enriched fractions. Importantly, restoration of miR-203 expression inhibited stemness and EMT properties of CLBC cells, inhibiting invasiveness, mammosphere formation, tumorigenesis, and metastasis, suggesting the potential of miR-203 to suppress transformation and breast cancer.
Promotion of EMT and Stem Cell-Like Features by Signals From the Microenvironment
Recent work has identified factors produced and released by the tumor microenvironment that promote EMT and stemness. In colon cancer, hepatocyte growth factor (HGF) secreted by stromal myofibroblasts promoted EMT and stemness [15]. These colon CSCs expressed high levels of Wnt induced by factors secreted by stromal myofibroblasts. Colon CSC self-renewal capacity was apparent from tumor initiation capacity, spheroid culture, and stem cell markers. Significantly, these factors secreted by the microenvironment were sufficient to promote CSC formation from more differentiated non-CSC tumor cells, showing the ability of the microenvironment to control EMT and stemness in colon cancer. Scheel et al. [16] have identified signals that induce and/or maintain mesenchymal features and stem cell fate in the breast. In this study, they used previously identified spontaneously arising mesenchymal subpopulations of cells (MSPs) isolated from immortalized human mammary epithelial cells. They found that the mesenchymal and stem cell state of MSPs was maintained by increased TGFβ1, BMP antagonists and Wnt signaling, and by reduced BMPs and Wnt antagonists. Importantly, the self-renewal and motility properties of the MSPs could be reversed by inhibiting TGFβ or by adding back BMP4 or Wnt antagonists. These findings emphasize the importance of autocrine signaling to maintaining the stem cell fate of these MSPs. They also asked whether this autocrine loop applies to cancer stem cell tumorigenicity and metastasis. For example, do Ras-transformed MSPs behave like CSCs? They found that Ras-transformed MSPs have more tumor initiating potential and are more metastatic than Ras-transformed mammary epithelial cells. Importantly, blocking the autocrine loop created by TGFβ and Wnt that supports MSPs also blocked tumor initiation and metastasis in the Ras-transformed MSPs.
EMT Promotes Stem Cell Like Features in Cancer Cells by Dedifferentiation
Until recently, CSCs were believed to exist at the top of a cellular hierarchy giving rise both to new CSCs with high tumor-initiating properties and to more differentiated non-CSC tumor cells. More recently, several studies have suggested that non-CSCs can acquire CSC features. Initial studies by Scheel et al. [16] and Chaffer et al. [17∙] described the ability of normal and non-CSC cells to convert to stem-like cells that were dependent on signals from the microenvironment. They predicted that this conversion and/or dedifferentiation may be promoted by signals that drive EMT. Similarly, as described above, studies of colon CSCs showed the ability of HGF to reprogram non-CSCs to CSCs [15]. This conversion or dedifferentiation was dependent on the factors secreted by stromal cells like HGF and on Wnt signals. These studies were extended to basal-like breast cancers, for which it was shown that conversion of non-CSCs to CSCs was frequent in specific subtypes of breast cancer, for example basal-like breast cancers. These conversions were dependent on EMT-inducing factors TGFβ and ZEB1, which promoted high levels of expression of the stem cell marker CD44. The ability of specific cancer cell types to interconvert between non-CSC and CSC states, thus controlling the tumorigenic and invasive potential of the cancer cells, is an important mechanism that may be targeted therapeutically to inhibit both tumorigenesis and metastasis.
Circulating Tumor Cells (CTCs), CSCs, and EMT: Mesenchymal Features of CTCs and How CTCs Relate to CSCs
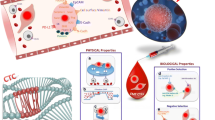
Recent studies of multiple cancer types have demonstrated EMT and stem cell properties of CTCs (Fig. 2). In pancreatic cancer, most pancreatic CTCs were found to exclusively express mesenchymal markers and to have stemness properties, including being CD24+/CD44+ and forming pancreatospheres [9∙]. In prostate cancer, CTCs were identified with stemness markers [18]. Different levels of epithelial and mesenchymal marker coexpression were observed for these CTCs, suggesting the prostate cancer CTCs were in different transitional states. In metastatic breast cancer patients, a major proportion of CTCs were found to have EMT and tumor stem cell properties [19]. This study was further extended to CTCs from breast cancer patients without diagnosis of metastasis, suggesting that acquisition of EMT and stemness can occur early in the dissemination process, similarly to pancreatic cancer [9∙, 20, 21]. These studies have been significantly extended by a recent study by Yu et al. [22∙], who showed that CTCs from metastatic breast cancer have features of EMT (Fig. 2). Importantly, they found that EMT in CTCs correlated with breast cancer treatment responses and patient relapse. They suggested the importance of EMT as a potential biomarker of therapeutic resistance and as a drug target in breast cancer. They also found that CTCs with mesenchymal markers often circulated as clusters instead of individual cells (Fig. 2). It remains unclear what proportion of CTCs are CSCs and what are the consequences to patient treatment, response, and survival. A major hurdle in the use of CTCs to monitor patient response to treatment and/or relapse is the presence of subpopulations of CTCs that are dedifferentiated CSCs with mesenchymal features. These cells, which have undergone EMT, are not likely to be detected by current CTC isolation approaches that rely heavily on such epithelial markers as EpCAM and cytokeratins, which may be lost in aggressive CTC and/or CSC phenotypes that have lost their epithelial features and gained mesenchymal characteristics. This is very relevant to patient response to current therapy, because drug-resistant CTCs clearly have mesenchymal features associated with EMT. Productive response to therapeutic intervention correlated with reduced mesenchymal CTCs, whereas therapeutic resistance correlated with increased mesenchymal CTCs, indicating the importance of being able to monitor, study, and treat these subpopulations in patients [22∙].

EMT, CSCs, and CTCs. Mesenchymal CTCs are likely to represent CTCs and CSCs with tumor-initiating potential. Epithelial tumor cells lacking mesenchymal features are shown in orange. Tumor cells in an intermediate state of EMT with both epithelial and mesenchymal features are shown in green. Individual or sheets of tumor cells primarily with mesenchymal features are shown in blue. It is postulated that it is the sheets of mesenchymal tumor cells [1], and not the epithelial tumor cells, which will successfully traverse the vasculature [2], exiting to form metastases [3], but this hypothesis needs additional experimental support (Color figure online)
Conclusions
Recent work has established the critical link between EMT and stemness features of CSCs. This work has identified novel mechanisms regulating the induction and maintenance of EMT and CSCs that will pave the way to finding novel solutions for targeting CSCs that are resistant to traditional cancer therapy [23, 24]. Clearly, better treatment of patients with CSCs could be achieved if CTC isolation methods could detect CTCs that have undergone EMT. Currently, many clinical trials are in progress to examine either EMT or CTCs (ClinicalTrials.gov). There are, however, only two trials, one of metastatic colorectal cancer (NCT01286883) and one of breast cancer (NCT01866202), that are examining EMT markers in CTCs and CSCs, with the idea of using these as biomarkers and/or prognostic markers of treatment. Defining molecular mechanisms controlling EMT in cancer cells will aid the treatment of patient cancers. The resistance of CSCs with EMT to traditional therapy clearly demonstrates the importance of targeting CSCs and/or the conversion of non-CSCs to CSCs. Recent high-throughput, small-molecule screening has focused on identification of treatment which specifically targets CSCs and/or EMT [25]. Additional work has focused on identification and use of approved drugs to reduce CSCs and/or EMT. One such proposed treatment is use of the broad-acting tyrosine kinase inhibitor sunitinib against claudin-low breast cancer and other EMT-dependent cancers [26]. The biotechnology industry is attempting to develop novel therapeutic strategies to specifically target EMT and CSCs. As we learn more about the genomic and/or proteomic properties of these cells, therapeutic targeting will become a real possibility for developing new therapy for treatment of cancer.
References
Recently published papers of particular interest have been highlighted as: ∙ Of importance
Kalluri R, Weinberg RA (2009) The basics of epithelial–mesenchymal transition. J Clin Investig 119:1420–1428
Zeisberg M, Neilson EG (2009) Biomarkers for epithelial–mesenchymal transitions. J Clin Investig 119:1429–1437
Hay ED (2005) The mesenchymal cell, its role in the embryo, and the remarkable signaling mechanisms that create it developmental dynamics. Dev Dyn 233:706–720
Thiery JP, Acloque H, Huang RY, Nieto MA (2009) Epithelial–mesenchymal transitions in development and disease. Cell 139:871–890
Visvader JE, Lindeman GJ (2012) Cancer stem cells: current status and evolving complexities. Cell Stem Cell 10:717–728
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, Yang J, Weinberg RA (2008) The epithelial–mesenchymal transition generates cells with properties of stem cells. Cell 133:704–715
Radisky DC, LaBarge MA (2008) Epithelial–mesenchymal transition and the stem cell phenotype. Cell Stem Cell 2:511–512
Creighton CJ, Li X, Landis M, Dixon JM, Neumeister VM, Sjolund A, Rimm DL, Wong H, Rodriguez A, Herschkowitz JI, Fan C, Zhang X, He X, Pavlick A, Gutierrez MC, Renshaw L, Larionov AA, Faratian D, Hilsenbeck SG, Perou CM, Lewis MT, Rosen JM, Chang JC (2009) Residual breast cancers after conventional therapy display mesenchymal as well as tumor-initiating features. Proc Natl Acad Sci USA 106:13820–13825
∙ Rhim AD, Mirek ET, Aiello NM, Maitra A, Bailey JM, McAllister F, Reichert M, Beatty GL, Rustgi AK, Vonderheide RH, Leach SD, Stanger BZ (2012) EMT and dissemination precede pancreatic tumor formation. Cell 148:349–361. One of the few in vivo demonstrations of metastasis showing pancreatic cancer cells with features of EMT and stemness. Some of these cancer stem cell like cells were in an “intermediate EMT”, simultaneously expressing both epithelial and mesenchymal markers
Abell AN, Jordan NV, Huang W, Prat A, Midland AA, Johnson NL, Granger DA, Mieczkowski PA, Perou CM, Gomez SM, Li L, Johnson GL (2011) MAP3K4/CBP-regulated H2B acetylation controls epithelial-mesenchymal transition in trophoblast stem cells. Cell Stem Cell 8:525–537
Jordan NV, Johnson GL, Abell AN (2011) Tracking the intermediate stages of epithelial–mesenchymal transition in epithelial stem cells and cancer. Cell Cycle 10:2865–2873
Jordan NV, Prat A, Abell AN, Zawistowski JS, Sciaky N, Karginova OA, Zhou B, Golitz BT, Perou CM, Johnson GL (2013) SWI/SNF chromatin-remodeling factor Smarcd3/Baf60c controls epithelial–mesenchymal transition by inducing Wnt5a signaling. Mol Cell Biol 33:3011–3025
Song SJ, Poliseno L, Song MS, Ala U, Webster K, Ng C, Beringer G, Brikbak NJ, Yuan X, Cantley LC, Richardson AL, Pandolfi PP (2013) MicroRNA-antagonism regulates breast cancer stemness and metastasis via TET-family-dependent chromatin remodeling. Cell 154:311–324
Taube JH, Malouf GG, Lu E, Sphyris N, Vijay V, Ramachandran PP, Ueno KR, Gaur S, Nicoloso MS, Rossi S, Herschkowitz JI, Rosen JM, Issa JP, Calin GA, Chang JT, Mani SA (2013) Epigenetic silencing of microRNA-203 is required for EMT and cancer stem cell properties. Sci Rep 3:2687
Vermeulen L, De Sousa EMF, van der Heijden M, Cameron K, de Jong JH, Borovski T, Tuynman JB, Todaro M, Merz C, Rodermond H, Sprick MR, Kemper K, Richel DJ, Stassi G, Medema JP (2010) Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat Cell Biol 12:468–476
Scheel C, Eaton EN, Li SH, Chaffer CL, Reinhardt F, Kah KJ, Bell G, Guo W, Rubin J, Richardson AL, Weinberg RA (2011) Paracrine and autocrine signals induce and maintain mesenchymal and stem cell states in the breast. Cell 145:926–940
∙ Chaffer CL, Brueckmann I, Scheel C, Kaestli AJ, Wiggins PA, Rodrigues LO, Brooks M, Reinhardt F, Su Y, Polyak K, Arendt LM, Kuperwasser C, Bierie B, Weinberg RA (2011) Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. In Proceedings of the national academy of sciences of the United States of America, vol 108, pp. 7950–7955. Demonstration of the plasticity of non-CSCs to become CSCs. This dedifferentiation is dependent on both signals from the microenvironment and the configuration of the ZEB1 promoter
Armstrong AJ, Marengo MS, Oltean S, Kemeny G, Bitting RL, Turnbull JD, Herold CI, Marcom PK, George DJ, Garcia-Blanco MA (2011) Circulating tumor cells from patients with advanced prostate and breast cancer display both epithelial and mesenchymal markers. Mol Cancer Res 9:997–1007
Aktas B, Tewes M, Fehm T, Hauch S, Kimmig R, Kasimir-Bauer S (2009) Stem cell and epithelial–mesenchymal transition markers are frequently overexpressed in circulating tumor cells of metastatic breast cancer patients. Breast Cancer Res 11:R46
Barriere G, Riouallon A, Renaudie J, Tartary M, Rigaud M (2012) Mesenchymal and stemness circulating tumor cells in early breast cancer diagnosis. BMC Cancer 12:114
Barriere G, Tartary M, Rigaud M (2012) Epithelial mesenchymal transition: a new insight into the detection of circulating tumor cells. ISRN Oncol 2012:382010
∙ Yu M, Bardia A, Wittner BS, Stott SL, Smas ME, Ting DT, Isakoff SJ, Ciciliano JC, Wells MN, Shah AM, Concannon KF, Donaldson MC, Sequist LV, Brachtel E, Sgroi D, Baselga J, Ramaswamy S, Toner M, Haber DA, Maheswaran S (2013) Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science 339:580–584. Show EMT in CTCs that correlates with breast cancer treatment responses and relapse. Many of the mesenchymal CTCs circulated as multicellular clusters as compared to individual cells. This study clearly suggests the importance of EMT as a potential biomarker for therapeutic resistance and as a drug target in breast cancer
Dave B, Mittal V, Tan NM, Chang JC (2012) Epithelial–mesenchymal transition, cancer stem cells and treatment resistance. Breast Cancer Res 14:202
Singh A, Settleman J (2010) EMT, cancer stem cells and drug resistance: an emerging axis of evil in the war on cancer. Oncogene 29:4741–4751
Gupta PB, Onder TT, Jiang G, Tao K, Kuperwasser C, Weinberg RA, Lander ES (2009) Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell 138:645–659
Hollier BG, Tinnirello AA, Werden SJ, Evans KW, Taube JH, Sarkar TR, Sphyris N, Shariati M, Kumar SV, Battula VL, Herschkowitz JI, Guerra R, Chang JT, Miura N, Rosen JM, Mani SA (2013) FOXC2 expression links epithelial–mesenchymal transition and stem cell properties in breast cancer. Cancer Res 73:1981–1992
Acknowledgments
Amy N. Abell is supported by funding from the Memphis Research Consortium. Gary L. Johnson is supported by National Institutes of Health grants GM101141, GM68820, and NCI Breast SPORE CA58223, the Komen Foundation, the Lustgarten Foundation, and the University Cancer Research Fund. The authors thank Betsy Clarke for her assistance in graphical work.
Compliance with Ethics Guidelines
Conflict of Interest
Amy N. Abell declares that she has no conflict of interest. Gary L. Johnson is cofounder of KinoDyn and inventor on a patent application for defining kinome dynamics in drug-resistant cancer.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Abell, A.N., Johnson, G.L. Implications of Mesenchymal Cells in Cancer Stem Cell Populations: Relevance to EMT. Curr Pathobiol Rep 2, 21–26 (2014). https://doi.org/10.1007/s40139-013-0034-7
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40139-013-0034-7