
Abstract
Key message
A set of models of bark thickness at breast height and bark volume are now available for six species in France. A common model suitable for predicting bark volume was proposed for all species. A small but significant altitude effect on bark thickness at breast height was detected for three species.
Context
The growing demand for wood energy and bio-molecules requires a thorough evaluation of forest biomass, particularly bark.
Aims
The objective of this study is to have statistical models of bark volumes for the six main forest species present in North-Eastern France and to be able to estimate regional bark biomasses and quantities of chemical extractives at regional scale.
Methods
A large databank gathering bark thickness measured at different heights in France was used for selecting literature or new alternative models of tree bark volume. These models were applied to the available forest inventory data from North-Eastern France to estimate the regional bark volume. Secondly, by multiplying these volumes by basic density data and extractive content recently obtained, bark biomasses and extractives quantities were deduced.
Results
The first results consist in a set of species-specific models of bark thickness at breast height with R2 around 0.70 and a relative RMSE around 30% which is an improvement of 0.1 for R2 and of 1–2% for relative RMSE depending on the species compared to the best models from the literature. The second results consist in a set of species-specific models of tree bark volumes with R2 of 0.90 and a relative RMSE which varies between 22% when bark thickness at breast height is included and 40% when it is predicted. A significant relationship between bark thickness at breast height and altitude was also observed. The bark resources of Grand Est and Bourgogne-Franche-Comté regions were estimated at 558 000 m3/year and 611 000 m3/year respectively representing between 5.5% and 15% of the stem volume depending on the species. The propagation of the measurement error of bark gauge was estimated at 5% for model of bark thickness at breast height and 24% for bark volume model.
Conclusion
These results constitute an important contribution for a better knowledge of the bark resource at a regional scale and may help to optimise bark valuation by the forest-wood sector.
Similar content being viewed by others

1 Introduction
Bark is a multifunctional structure absolutely necessary for the tree life (Rosell 2019). It protects living tissues, i.e., sapwood, phloem, dormant buds (Charles-Dominique et al. 2015), from high temperatures (Pausas 2015; Rosell 2016) or very low ones (De Antonio et al. 2020), superficial injuries due to rock falls, harvesting, herbivores and insects (Theander 1985; Harun and Labosky 1985), or pathogenic micro-organisms (Franceschi et al. 2005). The bark also has less known functions such as storage of water (Levia and Herwitz 2005) or mineral reserves (Schowalter and Morrell 2002), and is involved in tree biomechanics (Clair et al. 2019). Consequently, bark participates through its multiple functions to the adaptation of trees to different ecosystems (Rosell et al. 2014).
To fulfil these different functions, bark can be described by different variables: Its thickness at different heights (Gordon 1983), its total volume in a tree (Wehenkel et al. 2012; Rosell et al. 2017), its proportion of the volume of the stem (Cellini et al. 2012), its density (Miles and Smith 2009), its structure, particularly different in young or old bark (Dedrie et al. 2015), its yields and quantities of different chemical compounds (Jyske et al. 2014; Trivelato et al. 2016; Feng et al. 2013; Brennan et al. 2020), especially carbon (Jones and O’Hara 2018; Castaño-Santamaría and Bravo 2012) and minerals (Buamscha et al. 2007). All these variables not only participate in relating structures and functions in living trees but also are critical in determining the value of harvested timber.
To fulfil these different functions, bark can be described by different variables: Its thickness at different heights, its total volume in a tree, its proportion of the volume of the stem, its density, its structure, particularly different in young or old bark, its yields and quantities of different chemical compounds, especially carbon and minerals. All these variables not only participate in relating structures and functions in living trees but also are critical in determining the value of harvested timber.
To fulfil these different functions, bark can be described by different variables: Its thickness at different heights, its total volume in a tree, its proportion of the volume of the stem, its density, its structure, particularly different in young or old bark, its yields and quantities of different chemical compounds, especially carbon and minerals. All these variables not only participate in relating structures and functions in living trees but also are critical in determining the value of harvested timber.
The bark is thus a rich and very accessible raw material and human has exploited it for a very long time (Pasztory et al. 2016; Harkin and Rowe 1971) to produce for instance medicines (Turner and Hebda 1990; Anderson 1955; Rastogi et al. 2015), perfume (e.g., cinnamon), latex corks (Thomas et al. 1995) or insulating material (Gil 2014). In the current economy of the forest-wood sectors, bark is a by-product of the primary wood processing industries which contributes very little to their total turnover. Its economic value, mostly coming as a horticultural substrate or as a fuel (Lu et al. 2006), may reach 15% for all by-products pooled together (Chalayer 2015).
Not all softwood bark is suitable for use as horticultural substrate and, compared to wood, bark is a second choice fuel due to its high humidity and high mineral composition (Adler 2007) despite its high extractive content which improves its calorific value (Telmo and Lousada 2011; Tenorio and Moya 2013; Fuwape 1989). Only boilers specifically designed to burn biomass with a high content of water, impurities (soil, gravel, etc.), and mineral matter (and therefore ash) can burn it efficiently (Martin 2015).
It turns out that society is increasingly demanding biomolecules that are considered less harmful to health than petro-sourced products and are produced in a more environmentally friendly manner. Industries want to take advantage of this trend. They need consequently to answer questions about the volume of the different markets, the availability of raw materials rich in these biomolecules and the industrial processing of the raw material.
To estimate the available raw material, it is possible to make advantage of data and information already available. Bouvet and Deleuze (2013) have provided values of bark percentage to calculate stem bark volume from stem volume. There are also data on basic density to convert fresh wood volumes into dry biomasses (Billard et al. 2020). Moreover, the rate of extractable chemical compounds (Brennan et al. 2020) expressed in grams of extractives/gram of dry matter can be used to obtain the mass of extractives.
Depending on the intended purpose, bark quantity has been estimated either from bark thickness (Stängle et al. 2017; Muhairwe 2000; Laasasenaho et al. 2005), bark area (or proportion in bark area) or from whole stem bark biomass (e.g., Zianis et al. 2005). There are numerous studies that have developed bark thickness models (e.g., Gordon 1983; Muhairwe 2000; Hannrup 2004; Van Laar 2007). However, few studies have provided models to directly predict the volume of bark from tree variables such as diameter at breast height, tree height, and bark thickness at breast height when it is available (e.g., Kozak and Yang (1981)). Often cited, Meyer (1946) computed the bark volume of a tree on the basis of a constant proportionality between over-bark and under-bark stem diameters, generally measured at breast height. More recently, Wehenkel et al. (2012) and Liepiņš et al. (2015) have modelled the proportion of bark volume.
Modelling bark volume (Bv) and bark thickness at breast height (BTBH) for France is the central objective of this article. On the one hand, when models exist but are not adapted to the French resource they must be readjusted as advised by Jenkins et al. (2003) and Stängle et al. (2017). On the other hand, if the models do not exist, new ones must be developed. In addition, models of BTBH will also be designed since BTBH can be used as an input variable in bark volume models, yet they are not always measured in the field. Numerous bark data have been collected by French research and development organisations: INRAE (Institut National de Recherche pour l’Agriculture, l’Alimentation et l’Environnement), FCBA (institut technologique Forêt Cellulose Bois-construction, Ameublement), ONF (Office National des Forêts), IGN (Institut National de l’Information Géographique et Forestière). This database is managed by the FCBA (Bouvet and Deleuze 2013).
As a secondary objective, we applied the best models to forest inventory data to provide an initial estimate of the available resource, e.g. for the chemical industry. The resource must be assessed in terms of bark volume, bark biomass, and quantity of extractives. We provide this estimate at the scale of two French regions: Grand-Est and Bourgogne Franche-Comté. For the sake of clarity, we will consider only the total content of extractives and not the content of specific chemical families.
This study concerns six temperate tree species that are present and industrially processed in Eastern France: silver fir (Abies alba Mill.), Norway spruce (Picea abies L.), Douglas fir (Pseudotsuga menziesii Mirb.), European beech (Fagus sylvatica L.), sessile oak (Quercus petraea Matt.), and pedunculate oak (Quercus robur L.).
2 Materials and methods
BTBH models were developed from the data recorded during several campaigns of the IGN. Since 2008, these measurements are no longer carried out as the database has been considered complete. Models for total bark volume (Bv) were developed from a “research” dataset called EMERGE. Both datasets are described in detail in Section 2.1.
Table 1 summarises the abbreviations used in this paper.
2.1 Datasets
In this work, three different databases were used namely EMERGE, IGN, and French NFI.
2.1.1 EMERGE dataset
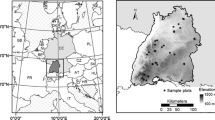
EMERGE was a project led by the French National Forest Office (ONF) and supported by the French National Research Agency (ANR) (Deleuze et al. 2013). Its purpose was to estimate the available biomass in French forests. Eight French research and development organisations worked together on this project. The EMERGE dataset combined several subsets of data collected by the various partners. This dataset contained bark thickness (Bt) measurements performed in several French regions (Fig. 1), statistically representative of the French resource. The measurements were made using a Swedish bark gauge at several heights along the stem but often not at breast height. In the latter case, a linear interpolation between the two values collected from closest values to 1.3 m allowed for an estimation at this height.

Location of the sites where the EMERGE data were recorded
Usual tree variables (total tree height, Htot and diameter at breast height, DBH) were measured and additional information about the corresponding plots such as geographic coordinates and altitude were recorded. The tree DBH distribution is given in Appendix Fig. 5.
The composition of EMERGE database is given in Table 2.
2.1.2 IGN dataset
The second dataset, called IGN is a “resource” dataset. BTBH was measured on a large number of trees everywhere in France. This dataset includes only one measurement at breast height made with a Swedish bark gauge as well as DBH, Htot and location of the trees (latitude, longitude and altitude). Table 3 shows the composition of the IGN dataset.
2.1.3 French NFI data in grand Est and Bourgogne-Franche-Comté
The National Forest Inventory (NFI) is a continuous statistical survey of French metropolitan forests, undertaken by the IGN. The NFI is carried out in public and private forests, regardless of whether they are available for wood supply. The NFI design features a systematic sampling grid with squared cells of 1 km side (Colin et al. 2017; Hervé 2016). Each year, 10% of the cells are sampled according to two phases. In the first phase, approximately 80 000 photo plots are interpreted to assess land cover and land use. In the second phase, approximately 7 000 temporary ground plots are established at a sub-sample of first phase photo plot locations that have forest land use. On each plot, every tree with a DBH of at least 7.5 cm is measured. Measurements include DBH and Htot. Typically, five annual NFI samples are combined to calculate statistics for the forest resource. Moreover, since 2010, the French NFI provides direct measurements of logging based on a re-inventory of temporary plots placed in the inventory 5 years earlier. For example, in 2014, the NFI returned data on plots for the inventory in 2009, which can serve as an objective for giving an estimate of the mean annual harvest between 2009 and 2014.
French NFI visits approximately 1 650 new inventory plots each year in the Grand Est and Bourgogne-Franche-Comté regions (870 plots and 780 plots, respectively). In this paper, we used the observations performed on cut trees during the 2014–2018 periods to infer the amount of bark volumes harvested each year in the two regions between 2009 and 2018. For that, the best BTBH models developed for each species were applied on all felling trees recorded by the NFI to estimate BTBH from tree DBH and altitude whenever significant. Then, the best Bv models were applied to estimate stem bark volume of felling trees based on estimated BTBH, tree DBH and tree Htot. Finally, the five annual NFI samples (2014–2018) were combined to estimate the volume of bark harvested each year, between 2009 and 2018, from the forest resource in the two regions.
2.2 Calculating the bark volume
First, all Bt measurements from the EMERGE dataset were used to compute over- and under-bark cross-sectional areas at different heights. Over- and under-bark stem area, respectively Aob and Aub at a given height were calculated with Eqs. 1 and 2.
where Dob is the over-bark stem diameter and Bt the bark thickness.
To calculate the bark volume of each tree, over- or under-trunk portions were considered as a stack of truncated cones with a volume given by Eq. 3 with the tree top portion considered as a cone (thus with A2 = 0) and the stump portion calculated by extrapolation from the areas calculated from the two lowest measures:
where Hc is the truncated cone height and A1 and A2 the area of, respectively, its lower section and upper sections.
The sum of over- and under-bark volumes of all truncated cones gives the total over- and under-bark stem volumes for the tree (Fig. 2a). And as a consequence, their difference gives the bark volume.

a Sequence of truncated cones modelling a stem. b Simplified geometric calculation of bark volume. BTBH: Bark thickness at breast height; DBH: Diameter at breast height; D0ob and D0ub: Diameters at ground level over and under bark; Htot: total tree height
2.3 BTBH modelling
We first adjusted literature models on the FCBA database and then design alternative models. Finally, we selected the most relevant models based on the lowest Akaike Information Criterion (AIC) and relative root mean square error (RMSErel).
2.3.1 BTBH models from the literature
We selected three models from Wilhelmsson et al. (2002) for Norway spruce and (Pinus sylvestris L.) (Eq. 4); Cao and Pepper (1986) for North-American species (Pinus echinata Mill., Pinus taeda L. and Pinus palustris Mill.) (Eq. 5) and Gordon (1983) for Pinus radiata D. Don in New-Zealand (Eq. 6).
2.3.2 Alternative BTBH models
After carefully analysing several equations we selected Eq. 7 as a base for our modelling, reflecting the strong relationship with DBH.
The addition of an intercept to Eq. 7 was studied but we preferred to remove it, even if this parameter was significant, because we considered that a tree with DBH = 0 should logically have BTBH = 0.
For assessing whether a relation between altitude and BTBH could be introduced, quantitative altitude values were transformed into ten increasing altitude classes and their mean was assigned to each tree belonging to their corresponding class. To make sure the classes were approximatively of comparable size, we merged classes representing less than 5% of our data with their closest class. The parameters a and b in Eq. 7 were then adjusted according to altitude class. We thus obtained around ten values of a and b, for each species, that were analysed in relation with the mean altitude of trees from each class. Sometimes and only on a, we observed a significant altitude effect. In this case Eq. 7 becomes Eq. 8.
with alt as the altitude
2.4 Bark volume models
2.4.1 Bark volume models from the literature
Meyer (1946) proposed a method to calculate Bv using the a coefficient according to Eq. 9.
where the sum is calculated on data from a pool of trees of the same species.
Bark volume for individual tree may therefore be computed using Eq. 10.
We determined the a coefficient from the IGN dataset for each species. We then calculated the bark volume of the trees from the EMERGE dataset using the a coefficient for this species and Eq. 10. This method was developed by Meyer (1946) for Cinchona but he also proposed an application for other species such as Tsuga canadensis, Pinus strobus, Quercus alba and Acer rubrum.
Kozak and Yang (1981) proposed a model of bark volume following Eq. 11. This model was applied on 32 species including both hardwood species and softwood species.
2.4.2 Alternative model of bark volume
We built a new model of Bv by considering the difference between two cones Vob and Vub which are the over-bark and under-bark stem volumes assuming the trunk is cone-shaped (Eq. 12 and Fig. 2 b).
where D0ob and D0ub are the over bark and under bark diameters at the ground level.
D0ob and D0ub can be calculated from DBH and BTBH with Eqs. 13 and 14, respectively, on the basis of the Thales’ theorem.
We obtained the theoretical Eq. 15.
For mature trees, by assuming that Htot ≫ 1.3 m and DBH ≫ BTBH, Eq. 15 can be simplified to give Eq. 16.
Models following Eqs. 17 and 18 were designed from Eqs. 15 and 16, respectively.
Nevertheless, Eqs. 15 and 16 are only exact for a perfectly cone shaped stem, both over bark and under bark. To account for the geometric difference with a real stem the two models following Eqs. 17 and 18, in which parameters a and b can be adjusted statistically, were designed.
Finally, we obtained the model following Eq. 19 by replacing BTBH by its prediction \(\widehat {BTBH}\).
\(\widehat {BTBH}\) is the predicted value obtained with the best BTBH models depending on the species.
2.5 Statistical methods
2.5.1 Modelling
All the regressions were carried out using the R software (R Core Team 2018). Two methods have been applied, depending on the modelled variable.
For BTBH, the R function nls was used to fit the model. A total of 13 outliers were removed from the datapool (0 observation for silver fir and for Norway spruce, 7 for Douglas fir, 2 for European beech, 2 for sessile oak, and 2 for pedunculate oak). These observations were removed based on graphical analysis.
For Bv, bark volume measurements showed strong heteroscedasticity with variance increasing with tree size. For handling heteroscedasticity in the non linear regression analyses, we used the gnls function, provided by the nlme package (Pinheiro et al. 2019) with a variance structure described by the function varPower (Eq. 20):
The Breusch-Pagan test (BP-test) was used to verify the efficiency of this formulation by analysing the heteroscedasticity of the model residuals.
The model has been considered without heteroscedasticity if the obtained p-value is above 0.1.
In order to assess if species-specific models were required especially for distinguishing the two oak species, the R function nlsList provided by the nlme package (Pinheiro et al. 2019) was used.
This function partitions the data according to the levels of a grouping factor, in our case, the species, and gives specific fits for each data partition, using same models for different factors.
2.5.2 Cross-validation
All models, either from the literature or alternative ones, were validated through a cross-validation method as follows: The data were separated according to their plot of origin. Then these parts were merged randomly to form ten groups of approximatively the same size. Each group was used as validation set, the remaining observations being used as training set. A fitted value was thus obtained from a validation set for each measure made. These two datasets were used to calculate RMSE and relative RMSE (RMSErel) in%, as recommended by Mayer and Butler (1993), using Eqs. 21 and 22, respectively.
where y is the measured value, \(\widehat {y}\) the fitted value, \(\bar {y}\) the mean of observed values and n the number of observations.
The R2 value of the model was obtained by fitting a linear regression between observed and fitted value, as recommended by Piñeiro et al. (2008). This R2 was taken to characterize the quality of this modelling.
Then our data were split again, calibration and validation performed twice more. Three values of the indicators RMSE, RMSErel, and R2 were thus obtained. Finally, these three values were used to calculate their mean value, which are presented in Section 3. However, the parameter values are issued from the model adjustment made on all datasets.
We also calculated the AIC (Akaike 1973) for each model. We used for this a model adjusted on all data, without cross-validation.
2.6 Propagation of measurement error (PME) for Norway spruce
According to the analysis done by Stängle et al. (2016), measuring Bt using a Swedish bark gauge is subject to an error. This measurement error was identified for Norway spruce species in the same analysis as an overestimation of the Bt by 13.6% ± 28.4% (mean ± standard deviation) relative to its true value. The relevance of this systematic error calls for considering it within the fitted model. To this end, we use a “toy” Monte Carlo method based approach. Its design is based on the rationale found in different references (Rocha and Nogueira 2012; Clarkson 2014; Mahmoud and Hegazy 2017). The method can be summarized in four steps.
The first step is to define a bound for each measurement we have of the Bt, based on the identified measurement error. The second step is to take random values of errors (according to a Gaussian generation) at each measurement we have of Bt within the defined bounds. The third step is to fit the model according to these random values. Then, we repeat for a sufficient number of times (> 1000), where at each simulation we get different values for the regression parameters, and eventually determine their mean and standard error. Finally, the fourth step is to use the formula (Eq. 23) of error propagation (Ku 1966) to calculate the propagation from each regression parameter onto the model.
where f is a function, which in our case, is the model we are interested in.
2.7 Consistency of our model predictions with proportions of bark on over-bark trunk volume
In order to check the relevance of our modelling results, we translated results from the IGN dataset into bark proportions (Bp, calculated with Eq. 24) using bark volume estimated by our selected models and over-bark stem volume estimated following (Tran-Ha et al. 2007) (Eq. 25). We translated the results from Emerge dataset into Bp using either the bark volume and over-bark trunk volume calculated according to the procedure found in Section 2.2 and Fig. 2 (Method 1), or the bark volume estimated by our selected models with BTBH measured and over-bark stem volume calculated as previously (Method 2). We then compared the results obtained according to these procedures with the results published in (FCBA 2019) and adapted them to our dataset (Emerge or IGN) according to the repartition of tree’s DBH and obtained by Meyer (1946) where, according to Eq. 10, Bp corresponds to (1 − a2). To calculate the Bp, (FCBA 2019) used the Emerge dataset to build a relation between the ratio of bark thickness to log radius at 1.3 m and Bp on the tree, then applied it to the IGN dataset. These relations were built for different DBH classes for each species.
The a and b coefficients were estimated by Tran-Ha et al. (2007) for several species including our six targeted species.
For all these comparisons, we used Eq. 26 in order to calculate a mean Bp of all our trees.
where n is the number of observations.
3 Results
3.1 BTBH modelling
The detailed plots showing data, trend lines and models for each species are given in Appendix Figs. 6 and 7.
3.1.1 Fit of literature models
Table 4 summarises the results obtained for models following Eqs. 4, 5, 6, 7 and 8.
With regard to the model proposed by Cao and Pepper (1986) for the sessile oak species, d parameter was found insignificant. The model was thus recalibrated without this parameter. This new model is the one presented here. In terms of R2, RMSErel and AIC, all models presented in Table 4 are quite similar. It can be observed that except for pedunculate oak and Norway spruce, the model proposed by Cao and Pepper (1986) is slightly better than others. For pedunculate oak, the three models are really equivalent. The PME calculation, which is possible in the case of the Norway spruce species, showed a 5.12%, 4.49%, and 5.67% error propagation in the case of Cao and Pepper (1986), Wilhelmsson et al. (2002), and Gordon (1983), respectively. For models following Eqs. 7 and 8, PME were 0.84% and 0.19%.
Models following Eq. 7 appeared to be rather good, with R2 of 0.65 for European beech, 0.67 for sessile oak, 0.68 for pedunculate oak, 0.69 for Norway spruce, 0.71 for silver fir, and 0.77 for Douglas fir. However, the RMSErel around 30% (up to 37.3% for European beech) shows that the natural variation of Bt cannot be perfectly described by a simple model. For all species, the model according to Eq. 7 was thus not better than the best models found in the literature except for Douglas fir, pedunculate oak and European beech. The new models had an accuracy close to that of the literature models but had the advantage of having only two parameters and better results in terms of propagation of measurement error for the Norway spruce species.
After adjusting the model on altitude classes, we observed a significant relation between a and altitude only for silver fir, Norway spruce and European beech. This relations are given in Appendix Fig. 8. Furthermore, it can be seen that models following Eq. 8 are better than models of the literature. At the end we selected the model following Eq. 8 for silver fir, Norway spruce, and European beech, models following Eq. 7 for Douglas fir and pedunculate oak, and model following Eq. 5 for sessile oak (Table 4). Figure 3 shows the form of the selected models predicting BTBH for each species.

Prediction of bark thickness at breast height (BTBH) from diameter at breast height (DBH) according to models selected in Table 4. a Models depending on altitude for silver fir, Norway spruce and European beech. b Models without altitude for Douglas fir, pedunculate oak and sessile oak. The dotted lines show the extrapolated parts of the BTBH
3.2 Models of B v
Table 5 shows the modelling results for all species.
First, by considering Breusch-Pagan test (BP-test) results, we observed that weights argument of gnls function handled correctly heteroscedasticity for all species except for the European beech models.
It can be observed that there is only a small difference between model following Eq. 17 and model following Eq. 18, in terms of AIC, RMSErel and R2. Consequently, the approximation we made by assuming, DBH >> BTBH and Htot >> 1.3 seems to be proper.
It can also be observed that results obtained with Eq. 18 are much better than the ones obtained by Meyers’ method (Eq. 10). RMSErel is decreased by 13% for silver fir, 12% for Norway spruce, over 9% for Douglas fir, 5% for sessile oak, 1.5% for pedunculate oak and 15% for European beech.
By comparing models following Eqs. 18 and 11 (Kozak and Yang 1981), it can be observed that, even if in terms of R2 both models seem rather good, in terms of relative RMSE and AIC, the model proposed by Kozak and Yang (1981) is significantly better. We select this model even if the one following Eq. 18 is simpler.
The PME is, in the case of Norway spruce, approximately 24.5% for model following Eq. 18, while 65.73% in the case of Kozak and Yang (1981). As Meyer’s is not a model, PME can not be calculated.
Table 6 presents the results obtained with models following Eqs. 19 and 11 with parameters summarised in Table 5 and BTBH estimated with selected models presented in Table 4.
By comparing models following Eq. 19, in Tables 6 and 5, several differences can be observed. For silver fir, Douglas fir, and European beech, a large increase of RMSErel can be detected when instead of using the model with BTBH (Eq. 18) we use the model with \(\widehat {BTBH}\). For Norway spruce, pedunculate oak, and sessile oak, a small increase of RMSErel can also be observed between the two models respectively of 6%, 4%, and 6%.
The same accuracy loss can be observed for the model proposed by Kozak and Yang (1981). It can be nevertheless pointed out that for sessile oak, model following Eq. 19 became better than models proposed by Kozak and Yang (1981).
3.3 Bark proportion
In Table 7, column Emerge Method 1 represents the measured Bp and column Emerge Method 2 the Bp predicted with the bark volume model of Eq. 11 (Kozak and Yang 1981, best fit model see Table 5). Column (FCBA 2019) adapted for IGN, (FCBA 2019) adapted for Emerge and Meyers’Coefficient, (1 − a2) are added to simplified the comparison with the results and will be discuss in the discussion section.
Between methods 1 and 2, the only change is the prediction of bark volume. The prediction over-estimates the bark proportion for all species.
3.4 Application of BTBH and B v models to NFI data to estimate bark resources at regional scale
Applying the best BTBH and Bv models to NFI data, as well as specific bark densities (Billard et al. 2020) and extractive content (Brennan et al. 2020), we were able to provide estimates of the volume, biomass and amount of extractives in the bark of all trees cut annually in Grand Est and Bourgogne-Franche-Comté regions. We will consider the whole stem, stump and tree top included. In total for the two regions, about 1.2 million m3 of bark were cut each year between 2009 and 2018. This represents a total bark biomass of 600 000 t/year and about 160 000 t/year of bark extractives. For each metric (volume, biomass, amount of extractives), the annual cutting was slightly higher in Bourgogne-Franche-Comté than in Grand Est (52% vs 48% of the total for the two regions, respectively). In addition, they were slight differences in harvest profiles in terms of species composition between the two regions. In Bourgogne-Franche-Comté, the annual cutting was dominated by pedunculate oak (22% of the regional bark volume cut each year) followed by Norway spruce (21%), Douglas fir and silver fir (18% each). By contrast in Grand Est, the annual cutting was dominated by Norway spruce (27% of the regional bark volume cut each year) followed by silver fir (20%), pedunculate oak (18%) and European beech (15%).
Table 8 and Fig. 4 show the ressources calculated for the six species in the two regions, Grand-Est and Bourgogne-Franche-Comté.

a Repartition of bark volume removals between species. b Removals depending on species for two regions. Error bars represent the 95% confidence interval around means
4 Discussion
4.1 Accuracy of measurements
Since a Swedish bark gauge has a high degree of inaccuracy (Stängle et al. 2016; Althen 1964), it is noteworthy to identify the different sources of errors that are propagated to the target models. To achieve this, we have used a method based on a Monte Carlo technique (see Section 2.6). At this stage, it is appropriate to mention that the amount of error propagated onto the model does not speak for the goodness of the model, but rather determines its accuracy and prediction power. Determining whether the model is good or not would require an instrumental variable method in the case of non-linear models (Pearl 2009), or a dedicated Monte Carlo simulations with pseudo-data and a detailed study of the bias and variance that could be generated on model parameters; this is outside the scope of this paper. In this article, we have applied this method on two kinds of selected models, BTBH and Bv.
For the BTBH model, we have seen that the error propagation (when altitude is unknown) for Eq. 5 is fairly small (5.12%). However, when altitude is known (Eq. 8) the error propagation is relatively smaller (0.19%). For the Bv models, Kozak and Yang’s 1981 model incurred a 64.8% error propagation. This can only be explained by the number of Bt measurements that are used to calculate Bv in the first place (several truncated cones with different Bt measurements), each measurement possessing an error on its own, eventually affecting its propagation within the model. It is expected that a better measurement technique could induce a higher inherent model accuracy.
4.2 Model application
In this work we applied models found in the literature to our targeted species. The Bv models following Eqs. 9 and 11 were initially designed for softwood and hardwood species. The differences in stem shape between both types of species were not taken into account. However, the models seem generic enough to fit both types of species in an equivalent way.
It can nevertheless be pointed out that for hardwood species, total bark volume may have been seriously underestimated as we do not consider the bark of branches. Ver Planck and MacFarlane (2014) estimated that, on average, branches of hardwoods account for 41% of the total wood volume in a tree.
4.3 Comparison of BTBH models
In spite of their variable forms, none of the tested models in this study provided RMSErel lower than 20%. This may be due to the natural variability of the bark, the measurement error described above, and/or the fact that no independant variables complementary to DBH and altitude were identified. Nevertheless, an effect of altitude was evidenced. The models that we recommend to use are shown in bold in Table 4.
4.4 Altitude effect
An effect of altitude was observed for three species (silver fir, Norway spruce, European beech). This effect is positive: the higher the altitude, the larger the parameter a and the more Bt increases with DBH. One of the reasons could be a different allocation of biomass between bark and the other parts of the tree, with the increasing altitude. No effect was observed for the other species (Douglas fir, sessile oak and pedunculate oak).
The difference between species can be first explained by a statistical reason: For some species data are available over a wide range of altitudes, while for others this range is more restricted. The altitude effect can be tested only in the first case. This hypothesis is confirmed because it turns out that for species for which the altitude effect is significant, i.e. silver fir, Norway spruce and European beech, the altitude ranges are respectively 0–1800 m, 0–2000 m and 0–1600 m. On the other hand, the range of altitudes is much more restricted for the other three species, respectively 0–1400 m, 0–1100 m and 0–1000 m for Douglas fir, sessile oak and pedunculate oak. To be exact it should be verified if in all cases, and particularly for Douglas fir, sessile oak, and pedunculate oak, this altitude range reflects the real distribution of trees elevation or represents only a part of it.
With regard to the effect of environmental factors, many studies have showed relationships between Bt and fire (Pausas 2015; Schafer et al. 2015; Bauer et al. 2010). Although fire is not a common issue for these species in France, bark thermal insulation properties are worth considering. Indeed, De Antonio et al. (2020) showed that bark properties, especially low bark density, protected buds against frost while Molina et al. (2016) showed a link between resistance to frost and Bt. They have worked on species of Brazilian savannah and in Patagonia. It can be assumed that this protection strategy also applied to our species. Further investigations may be required to analyse the actual effect of frost on increasing Bt. Moreover, it is questionable whether the protection strategy of Douglas fir, sessile oak, and pedunculate oak against frost may be based more on decreasing bark density than on increasing Bt.
Another factor influencing relative bark thickness is growth rate (Stängle et al. 2017; Stängle and Dormann 2018; Laasasenaho et al. 2005). Growth rate has a negative influence on relative bark thickness, meaning that the slower a tree grows, the bigger Bt will be compared to DBH. The fact that trees grow slower at higher altitudes, given the more difficult environmental conditions, may explain the observed relationship. However, we are not able to separate the influence of growth rate and the influence of altitude since the age of the trees was not recorded.
Rosell et al. (2014) showed that bark also has a function of water storage. Water storage mainly increases with Bt. As studied by Antoni et al. (2011), French mountains have a smaller usable reserve of water than lowland areas which could influence Bt. However, usable reserve is not only dependent on altitude, and thus further research is needed to validate this hypothesis.
If one of these reasons can be validated, it is reasonable to think that using these values (temperature or usable reserve of water) will enable a better model to be built, that would be better able to represent the actual effect that is linked to the variation of Bt.
Other variables such as latitude, longitude, and type of forest (coppice or high forest), and Htot were tested but no significant influence was found.
4.5 Comparison of B v models
In the Results section, we have selected the model following Eq. 11 taking into account RMSErel and R2 values. However, it can be reasonably argued that the model following Eq. 18 is preferable since it has fewer parameters and thus is more robust and simpler to use. Moreover, the error associated to the parameters is smaller for the model following Eq. 18 than for the one following Eq. 11 (Appendix Table 10). Thus the model following Eq. 18, although slightly less accurate than model following Eq. 11, could be a good alternative.
Comparing Tables 5 and 6 it appears that, for silver fir, Douglas fir, and European beech, the accuracy losses by replacing BTBH by \(\widehat {BTBH}\) are respectively of 23%, 14%, and 25%. Although BTBH measurements are found to be helpful when predicting Bv for these species, the models predicting BTBH for these species are not enough accurate to predict correctly Bv.
4.6 Comparison of B p values predicted by the models
As shown by Table 7, compared to Emerge Method 1, the values predicted using the Meyer’s coefficients all over-estimated Bp except for European beech.
The difference with FCBA (2019) may be due to the method applied to the provided data. Indeed, FCBA (2019) provided Bp for different ranges of DBH starting from 25 cm. However, for computing the mean proportion we averaged these values weighed by the number of trees in each class and we applied the value of the lower class (25–30 cm) to all trees smaller than 25 cm.
The IGN dataset (column IGN) is closer to (FCBA 2019) (column (FCBA 2019) adapted for IGN) except for sessile oak, pedunculate oak, and Norway spruce. The high proportion of trees with a DBH smaller than 20cm can explain this difference, especially for Norway spruce. One may wonder if, for pedunculate oak and sessile oak, there is an important variation of Bp with respect to DBH for small trees. It can also be observed that the assumption made by Meyer (1946) of a constant Bp along the stem is rightful for silver fir, Norway spruce, and European beech. Indeed, the Meyers’ coefficient is close to the bark ratio measured for our trees except for Douglas fir and the two oak species.
5 Conclusion
To assess regional bark availability in terms of volume, biomass and quantities of extractive, we built several models to predict bark thickness at breast height and tree bark volume, from usual tree measurements such as total height and diameter at breast height and a site variable, altitude. This modelling was achieved for six temperate species: Silver fir, Norway spruce, Douglas fir, sessile oak, pedunculate oak, and European beech. We observed a statistical influence of altitude on bark thickness at breast height for three species, silver fir, Norway spruce, and European beech which opens the door to more ecological studies on bark. These models have been fitted on a pool of data collected in France. Considering the number of trees studied, the diversity of measurements made, and the number of bark thickness measurements made, these data are particularly rich and unique. The model set includes models developed elsewhere for other species or new ones created in this study. In this paper we have been able to valorise national forest inventory data, newly collected basic density and extractive rate data.
Change history
30 June 2023
A Correction to this paper has been published: https://doi.org/10.1186/s13595-023-01195-7
References
Adler A (2007) Accumulation of elements in Salix and other species used in vegetation filters with focus on wood fuel quality. Ph.D thesis. Swedish University ofAgricultural Sciences, Uppsala
Akaike (1973) Information theory and an extension of the maximum likelihood principle. Springer, New York
Althen FV (1964) Accuracy of the Swedish bark measuring gauge. For Chron 40:257–258
Anderson AB (1955) Recovery and utilization of tree extractives. Econ Bot 9:108
Antoni V, Arrouays D, Bispo A, Brossard M, Le Bas C, Stengel P, Villanneau E et al (2011) L’état des sols de France. Groupement d’intérêt scientifique sur les sols
Bauer G, Speck T, Blömer J, Bertling J, Speck O (2010) Insulation capability of the bark of trees with different fire adaptation. J Mater Sci 45:5950–5959
Billard A, Bauer R, Mothe F, Jonard M, Colin F, Longuetaud F (2020) Improving aboveground biomass estimates by taking into account density variations between tree components. Ann For Sci 77:103
Bouvet A, Deleuze C (2013) Taux d’écorce pour les principales essences forestières françaises. Rendez-Vous Techniques 39-40:60–67
Brennan M, Fritsch C, Cosgun S, Dumarcay S, Colin F, Gérardin P (2020) Quantitative and qualitative composition of bark polyphenols changes longitudinally with bark maturity in Abies alba Mill. Ann For Sci 77:1–14
Buamscha MG, Altland JE, Sullivan DM, Horneck DA, Cassidy J (2007) Chemical and physical properties of Douglas fir bark relevant to the production of container plants. HortScience 42:1281–1286
Cao QV, Pepper WD (1986) Predicting inside bark diameter for shortleaf, loblolly, and longleaf pines. South J Appl For 10:220–224
Castaño-Santamaría J, Bravo F (2012) Variation in carbon concentration and basic density along stems of sessile oak (Quercus petraea (Matt.) Liebl.) and Pyrenean oak (Quercus pyrenaica Willd.) in the Cantabrian Range (NW Spain). Ann For Sci 69:663–672
Cellini JM, Galarza M, Burns SL, Martinez-Pastur G, Lencinas MV (2012) Equations of bark thickness and volume profiles at different heights with easy-measurement variables. Forest Syst 21:23–30. http://sedici.unlp.edu.ar/handle/10915/42330
Chalayer M (2015) L’état de grâce des produits connexes de scieries. Forêt privée 345:75–83
Charles-Dominique T, Beckett H, Midgley GF, Bond WJ (2015) Bud protection: a key trait for species sorting in a forest–savanna mosaic. New Phytol 207:1052–1060
Clair B, Ghislain B, Prunier J, Lehnebach R, Beauchêne J, Alméras T (2019) Mechanical contribution of secondary phloem to postural control in trees: the bark side of the force. New Phytol 221:209–217
Clarkson W (2014) HOWTO Estimate parameter-errors by Monte-Carlo. http://www-personal.umd.umich.edu/wiclarks/AstroLab/HOWTOs/NotebookStuff/MonteCarloHOWTO.html
Colin A, Wernsdörfer H, Thivolle-Cazat A, Bontemps J-D (2017) National woody biomass projection systems based on forest inventory, Fance. In: Barreiro S, Schelhaas M-J, McRoberts RE, Ländler G (eds) Forest inventory-based projection systems for wood and biomass availability
De Antonio AC, Scalon MC, Rossatto DR (2020) The role of bud protection and bark density in frost resistance of savanna trees. Plant Biol 22:55–61
Dedrie M, Jacquet N, Bombeck P. -L., Hébert J., Richel A (2015) Oak barks as raw materials for the extraction of polyphenols for the chemical and pharmaceutical sectors: a regional case study. Ind Crop Prod 70:316–321
Deleuze C, Morneau F, Constant T, Saint André L, Bouvet A, Colin A, Vallet P, Gauthier A, Jaeger M (2013) Le projet EMERGE pour des tarifs cohérents de volumes et biomasses des essences forestières françaises métropolitaines. Rendez-vous Techniques ONF 39-40:32–36
FCBA (2019) Memento 2019. Institut Technologique Forêt Cellulose Bois-construction Ameublement, Champs-sur-Marne, France https://www.fcba.fr/content/memento
Feng S, Cheng S, Yuan Z, Leitch M, Xu CC (2013) Valorization of bark for chemicals and materials: A review. Renew Sustain Energy Rev 26:560–578
Franceschi VR, Krokene P, Christiansen E, Krekling T (2005) Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol 167:353–376
Fuwape JA (1989) Gross heat of combustion of Gmelina (Gmelina arborea (Roxb)) chemical components. Biomass 19:281–287
Gil L (2014) Cork: a strategic material. Front Chem 2:16
Gordon A (1983) Estimating bark thickness of pinus radiata. NZJ For Sci 13:340–348
Hannrup B (2004) Funktioner för skattning av barkens tjocklek hos tall och gran vid avverkning med skördare. Skogforsk
Harkin JM, Rowe JW (1971) Bark and its possible uses. U.S. Department of Agriculture, Forest Service Forest products laboratoyr. Madison, Wis
Harun J, Labosky P (1985) Antitermitic and antifungal properties of selected bark extractives. Wood Fiber Sci 17:327–335
Hervé J-C (2016) National forest inventories reports. In: Vidal C, Alberdi I, Hernández L, Redmond J (eds) National forest inventories. Springer, France
Jenkins JC, Chojnacky DC, Heath LS, Bsey RA (2003) National-scale biomass estimators for United States tree species. For Sci 49:12–35
Jones DA, O’Hara KL (2018) Variation in carbon fraction, density, and carbon density in conifer tree tissues. For 9:430
Jyske T, Laakso T, Latva-Mäenpää H, Tapanila T, Saranpää P (2014) Yield of stilbene glucosides from the bark of young and old Norway spruce stems. Biomass Bioenergy 71:216–227
Kozak A, Yang R (1981) Equations for estimating bark volume and thickness of commercial trees in British Columbia. For Chron 57:112–115
Ku HH (1966) Notes on the use of propagation of error formulas. Eng Instrum 70:263–273
Laasasenaho J, Melkas T, Alden S (2005) Modelling bark thickness of Picea abies with taper curves. For Ecol Manag 206:35–47
Levia D, Herwitz S (2005) Interspecific variation of bark water storage capacity of three deciduous tree species in relation to stemflow yield and solute flux to forest soils. Catena 64:117–137
Liepiņš J, Liepiņš K, et al. (2015) Evaluation of bark volume of four tree species in Latvia. Res Rural Develop 2:22–28
Lu W, Sibley JL, Gilliam CH, Bannon JS, Zhang Y (2006) Estimation of US bark generation and implications for horticultural industries. J Environ Hortic 24:29–34
Mahmoud GM, Hegazy RS (2017) Comparison of GUM and Monte Carlo methods for the uncertainty estimation in hardness measurements. Int J Metrol Qual Eng 8:14. publisher: EDP Sciences. https://www.metrology-journal.org/articles/ijmqe/abs/2017/01/ijmqe170002/ijmqe170002.html
Martin P (2015) Les Combustibles Bois. Tech rep. VAlBiom, Service public de Wallonie
Mayer D, Butler D (1993) Statistical validation. Ecol Modell 68:21–32
Meyer (1946) Bark volume Determination in Tree. J For 1061–1070
Miles PD, Smith WB (2009) Specific gravity and other properties of wood and bark for 156 tree species found in North America US Department of Agriculture. Forest Service, Northern Research Station
Molina JGA, Hadad MA, Domínguez DP, Roig FA (2016) Tree age and bark thickness as traits linked to frost ring probability on Araucaria araucana trees in northern Patagonia. Dendrochronologia 37:116–125
Muhairwe CK (2000) Bark thickness equations for five commercial tree species in regrowth For of Northern New South Wales. Australian forestry 63:34–43
Pasztory Z, Mohácsiné IR, Gorbacheva G, Börcsök Z (2016) The utilization of tree bark. BioResources 11:7859–7888
Pausas JG (2015) Bark thickness and fire regime. Funct Ecol 29:315–327
Pearl J (2009) Models Reasoning, and Inference, 3nd edn. Causality
Piñeiro G., Perelman S, Guerschman JP, Paruelo JM (2008) How to evaluate models: observed vs. predicted or predicted vs. observed? Ecol Modell 216:316–322
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2019) Nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-139. https://CRAN.R-project.org/package=nlme
R Core Team (2018) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rastogi S, Pandey MM, Rawat AKS (2015) Medicinal plants of the genus Betula—Traditional uses and a phytochemical–pharmacological review. J Ethnopharmacol 159:62–83
Rocha WFC, Nogueira R (2012) Monte Carlo simulation for the evaluation of measurement uncertainty of pharmaceutical certified reference materials. J Braz Chem Soc 23:385–391
Rosell JA (2016) Bark thickness across the angiosperms: more than just fire. New Phytol 211:90–102
Rosell JA (2019) Bark in woody plants: understanding the diversity of a multifunctional structure. Integr Comp Biol 59:535–547
Rosell JA, Gleason S, Méndez-Alonzo R., Chang Y, Westoby M (2014) Bark functional ecology: evidence for tradeoffs, functional coordination, and environment producing bark diversity. New Phytol 201:486–497
Rosell JA, Wehenkel C, Pérez-martínez A, Arreola Palacios JA, García-jácome SP, Olguín M (2017) Updating bark proportions for the estimation of tropical timber volumes by indigenous community-based forest enterprises in Quintana Roo, Mexico. For 8:338
Schafer JL, Breslow BP, Hohmann MG, Hoffmann WA (2015) Relative bark thickness is correlated with tree species distributions along a fire frequency gradient. Fire Ecology 11:74–87
Schowalter TD, Morrell JJ (2002) Nutritional quality of Douglas-fir wood: effect of vertical and horizontal position on nutrient levels. Wood Fiber Sci 34(1). https://ir.library.oregonstate.edu/concern/articles/s7526c877
Stängle SM, Dormann CF (2018) Modelling the variation of bark thickness within and between European silver fir (Abies alba Mill.) trees in southwest Germany. Int J For Res 91:283–294
Stängle SM, Sauter UH, Dormann CF (2017) Comparison of models for estimating bark thickness of Picea abies in southwest Germany: the role of tree, stand, and environmental factors. Ann For Sci 74:16
Stängle SM, Weiskittel AR, Dormann CF, Brüchert F (2016) Measurement and prediction of bark thickness in Picea Abies: assessment of accuracy, precision, and sample size requirements. Can J For Res 46:39–47
Telmo C, Lousada J (2011) The explained variation by lignin and extractive contents on higher heating value of wood. Biomass Bioenergy 35:1663–1667
Tenorio C, Moya R (2013) Thermogravimetric characteristics, its relation with extractives and chemical properties and combustion characteristics of ten fast-growth species in Costa Rica. Thermochim Acta 563:12–21
Theander O (1985) Cellulose, hemicellulose and extractives. In: Overend R.P., Milne T.A., M.L. (eds) Fundam Thermochem Biomass Convers. Springer, Dordrecht, pp 35–60
Thomas V, Premakumari D, Reghu C, Panikkar A, Saraswathy CA (1995) Anatomical and histochemical aspects of bark regeneration in Hevea brasiliensis. Ann Bot 75:421–426
Tran-Ha M, Perotte G, Cordonnier T, Duplat P (2007) Volume tige d’un arbre ou d’une collection d’arbres pour six essences principales en France. Revue Forestiè,re Française 59:609–624
Trivelato P, Mayer C, Barakat A, Fulcrand H, Aouf C (2016) Douglas bark dry fractionation for polyphenols isolation: From forestry waste to added value products. Ind Crop Prod 86:12–15
Turner NJ, Hebda RJ (1990) Contemporary use of bark for medicine by two Salishan native elders of southeast Vancouver Island, Canada. J Ethnopharmacol 29:59–72
Van Laar A (2007) Bark thickness and bark volume of pinus patula in south africa. Southern Hemisphere Forestry Journal 69:165–168
Ver Planck NR, MacFarlane DW (2014) Modelling vertical allocation of tree stem and branch volume for hardwoods. For: Int J For Res 87:459–469
Wehenkel C, Cruz-Cobos F, Carrillo A, Lujan-Soto JE (2012) Estimating bark volumes for 16 native tree species on the Sierra Madre Occidental, Mexico. Scand J For Res 27:578–585
Wilhelmsson L, Arlinger J, Spångberg K, Lundqvist S-O, Grahn T, Hedenberg Ö, Olsson L (2002) Models for predicting wood properties in stems of Picea abies and Pinus sylvestris in Sweden. Scand J For Res 17:330–350
Zianis D, Muukkonen P, Mäkipää R, Mencuccini M (2005) Biomass and stem volume equations for tree species in Europe. Silva Fennica Monographs
Acknowledgements
We would like to thanks Christine Deleuze and Philippe Lejeune for theirs advices to the modelling part, Philippe Saintenoise for his help on the statistical test, Mathieu Fortin for his assistance on the error propagation part and Pierre Montpied for his help on the handling of the heteroscedasticity of our data.
Funding
This thesis work was made possible in terms of salaries by funding from the Ministry of Agriculture and Agri-Food (MAA) represented by the DRAAF Grand Est and from the ERDF Lorraine. Operating funds were brought by the French National Research Agency (ANR) as part of the Investissements d’Avenir program (ANR-11- LABX-0002-01, Lab of Excellence ARBRE). The EMERGE dataset was collected within the frame of the EMERGE ANR project leaded by Christine Deleuze. IGN dataset was collected by Institut Geographique National.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Handling Editor: John Moore
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contribution of the co-authors Rodolphe Bauer performed the data analysis, wrote the original draft of this paper and was the main writer. Antoine Billard helped with the data analysis and contributed to the review and editing. Frédéric Mothe helped with the data analysis, contributed to the creation of alternatives model and contributed to the writing. Fleur Longuetaud helped with the data analysis, contributed to the creation of alternatives model and contributed to the writing. Mojtaba Houballah worked on the error propagation measurements part. Alain Bouvet collected the data, helped on the comparison with FCBA bark proportion and participated on the discussion and data analysing. Henri Cuny performed the estimating on Bourgogne-FrancheComté and Grand Est bark resources and review the article. Antoine Colin performed the estimating on Bourgogne-Franche-Comté and Grand Est bark resources and review the article. Francis Colin, supervised the work, coordinated the ExtraFor_Est project, helped with the data analysis and contributed to the discussion, review and editing.
Appendix
Appendix

Diameter at breast height (DBH) distribution of the trees from Emerge and IGN datasets, for silver fir, Norway spruce, Douglas fir, pedunculate oak, sessile oak and European beech

Relationship between standardized residuals and fitted values for silver fir, Norway spruce, Douglas fir, pedunculate oak, sessile oak and European beech for the models selected in Table 4. The red lines correspond to the horizontal axes, the green lines give the trend of the residuals

Relationship between standardized residuals and fitted values for silver fir, Norway spruce, Douglas fir, pedunculate oak, sessile oak and European beech for the models selected in Table 5. The red lines correspond to the horizontal axes, the green lines give the trend of the residuals

Relationship between altitude and the value of a parameter in model following Eq. 7 for silver fir, Norway spruce and European beech. Red line corresponds to the regression done
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bauer, R., Billard, A., Mothe, F. et al. Modelling bark volume for six commercially important tree species in France: assessment of models and application at regional scale. Annals of Forest Science 78, 104 (2021). https://doi.org/10.1007/s13595-021-01096-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-021-01096-7