
Abstract
Key message
Water availability and soil pH seem to be major constraints for enzyme activities in calcareous soils under Pinus halepensis and acidic soils under Pinus sylvestris plantations respectively. Proposals for improving enzyme activities may include the promotion of broadleaf species to increase soil pH and the modulation of stand density or the implementation of soil preparation techniques to facilitate water infiltration.
Context
Soil enzymes play a key role in nutrient turnover in forest ecosystems, as they are responsible for the transformation of organic matter into available nutrients for plants. Enzyme activities are commonly influenced by temperature, humidity, nutrient availability, pH, and organic matter content.
Aims
To assess the differences between enzyme activities in calcareous soils below Pinus halepensis and acidic soils below Pinus sylvestris plantations in Spain and to trace those differences back to edapho-climatic parameters to answer the questions: Which environmental factors drive enzyme activities in these soils? How can forest management improve them?
Methods
The differences in climatic, soil physical, chemical, and biochemical parameters and the correlations between these parameters and enzyme activities in soils were assessed.
Results
Low pH and high level of phenols in acidic soils under Pinus sylvestris and water deficit in calcareous soils under Pinus halepensis plantations appeared to be the most limiting factors for enzyme activities.
Conclusion
Options such as the promotion of native broadleaf species in the Pinus sylvestris stands and the modulation of Pinus halepensis stand density or the implementation of soil preparation techniques may improve enzyme activities and, therefore, nutrient availability.
Similar content being viewed by others


1 Introduction
Research on nutrient dynamics is vital to understanding forest ecosystem dynamics (Muscolo et al. 2007). Soil enzyme activities play a key role in the catalytic reactions involved in organic matter decomposition and nutrient cycling by regulating carbon, nitrogen, and phosphorous turnover, which then regulates the availability of these nutrients to plants. They are also responsible for the humification of the organic matter in soils. Enzymes in soils are mainly produced by bacteria and fungi (Burns 1978; Sinsabaugh 1994), but also by plant roots and animals (Bloem et al. 2006; Tabatabai 1994). Soil microbial biomass is considered the “eye of the needle” through which all organic material in soil must pass (Jenkinson 1977). Soil enzyme activities are positively related to soil organic matter and responsible for soil quality, crop productivity, and energy transfer (Tabatabai 1994). Soil enzymes have the potential to respond rapidly to environmental changes and serve as indicators of health and quality in planted and natural ecosystems (Bloem et al. 2006).
As an active component of soil organic matter, microbial biomass participates in the transformations and accumulation of nutrients in soil and serves as a good measurement of organic matter turnover and biological activity in forest and agricultural ecosystems (Bueis et al. 2016; Gartzia-Bengoetxea et al. 2009). Enzyme activities and soil microbial biomass have been proposed as suitable indicators of nutrient availability (Yang et al. 2012) and soil quality (Muscolo et al. 2015) because of their relation to soil biology and their quick reaction to changes (Bandick and Dick 1999; Das and Varma 2011; Llorente and Turrion 2010; Muscolo et al. 2007). Consequently, biochemical parameters have been included in models to predict Pinus halepensis and Pinus sylvestris productivity in Spanish plantations (Bueis et al. 2017b; Bueis et al. 2016).
Decomposition of organic matter is an important process through which nutrients are released into soil. It affects ecosystem productivity, particularly in forests and nutrient-poor ecosystems (Muscolo et al. 2007), such as those of the Mediterranean. Nutrient release from plant litter takes place through the enzymatic activities of microorganisms in the soil and depends on several complex and interacting mechanisms. Besides, enzymatic activities are also responsible for the organic matter stabilization through the humification process. Humus improves the soil water and nutrient retention, presents a hormonal role for plants, acts as a filter for contaminants, and also presents soil pH buffering capacity.
Soil enzymes catalyze the main reactions involved in nutrient cycles and then serve as sensitive indicators of soil fertility (Burns 1978; Nannipieri et al. 2002) and provide integrated information about soil function as they catalyze a wide variety of reactions involved in biological processes, especially dehydrogenase, urease, and phosphatase (Nannipieri et al. 2002). The individual enzymes most often used in bibliography as soil fertility indicators are dehydrogenase, fluorescein diacetate hydrolysis reaction, urease, and phosphatase activities (Gil-Sotres et al. 2005). Therefore, these enzyme activities have been included in this study. Additionally, catalase, an intracellular enzyme with detoxifying activity, has been included as a useful indicator of detoxifying needs in soils (Garcia and Hernandez 1997).
Most enzymes catalyze reactions inside living cells (intracellular enzymes), but many others function outside the cell (extracellular enzymes). Intracellular enzymes are also released when cells die and, together with extracellular enzymes, can be stabilized with organo-mineral complexes, enabling them to remain active for long periods of time (Bloem et al. 2006).
The fluorescein diacetate hydrolysis reaction reflects all hydrolytic activity in soil (Alef and Nannipieri 1995). It has been used as an indicator of general microbial activity in soil (Bandick and Dick 1999) and may provide detailed information about changes in soil organic matter dynamics.
Dehydrogenases are the main oxidoreductase enzymes that oxidize organic compounds in soils by transferring electrons between substrates and acceptors (Das and Varma 2011). They are used as indicators of microbial redox systems and considered a measure of microbial oxidative activities in soils (Burns 1978; Tabatabai 1994). Dehydrogenase activity is broadly used as an indicator of biological activity in soils (Casida et al. 1964). It reflects the activity of a group of enzymes that are present inside cells and do not accumulate outside the cell (Tabatabai 1994).
Phosphatases have an essential function in the P cycle (Burns 1978), as they catalyze the hydrolysis of organic forms of P into inorganic forms that are available to plants (Alef and Nannipieri 1995). A negative correlation is usually found between P availability and phosphatase activity due to the negative feedback of phosphorus concentration on enzyme activity (Olander and Vitousek 2000). Phosphatase activity plays a crucial role in the P cycle (Burns 1978) and is correlated to P stress and plant growth. In P-deficient soils, acid phosphatase secretion from plant roots increases to enhance phosphate availability to plants (Nannipieri et al. 2011).
Urease, an extracellular enzyme that catalyzes the hydrolysis of urea to NH3 and CO2 (Das and Varma 2011), accounts for up to 63% of total enzyme activity in soil (Martínez-Salgado et al. 2010) and is often used as a soil biological indicator because of its high sensitivity to changes induced by external factors.
The relationships between microbial activities and environmental factors such as soil chemistry, climate, or vegetation are crucial to understanding how ecosystems function. The aims of this study were (1) to assess the differences in the fluorescein diacetate hydrolysis reaction and dehydrogenase, acid and alkaline phosphatase, urease (most often used as soil fertility indicators), and catalase activities (indicative of detoxifying needs) between two contrasting soil types in Spain (calcareous soils under Pinus halepensis Mill. and acidic soils under Pinus sylvestris L. plantations) and (2) to trace those differences back to edapho-climatic parameters by means of the study of the correlations of these enzyme activities and environmental parameters on each of the contrasting soils studied separately to answer the questions: Which environmental factors drive enzyme activities in these soils? How can forest management improve those limiting factors?
2 Material and methods
2.1 Study area
The study took place in the forests of the Castile and Leon region of northern Spain. Specifically, the research was carried out in 32 plots on Pinus halepensis plantations belonging to the Spanish National Forest Inventory (located in the center of Castilla y León; Table 1) and 35 plots on Pinus sylvestris plantations belonging to the Sustainable Forest Management Research Institute (located in the north of Castilla y León; Table 2). Stand ages ranged from 28 to 54 years in Pinus sylvestris plots and from 45 to 61 years in Pinus halepensis plots. Stand density data (trees ha−1) was extracted from the 3rd Spanish National Forest Inventory for Pinus halepensis and was determined in the field for Pinus sylvestris plots. Pinus sylvestris plots are located at an average altitude of 1067 m above sea level and Pinus halepensis plots at 821 m.a.s.l.
Lithology in Pinus sylvestris plots is composed of quarzitic gravels with a sandy-clay matrix while in Pinus halepensis plots is composed of limestone and marl (IGME 1975). Soils in the areas studied are classified as inceptisols but present contrasting characteristics. Under the Pinus sylvestris plantations, soils are highly acidic (pH ranging from 3.7 to 5.6) and P availability is very low because it has been immobilized in insoluble Fe and Al compounds (Bueis et al. 2016). Under Pinus halepensis plantations, soils are calcareous (pH ranges from 8.0 to 8.9) and P availability is even lower because P forms insoluble calcium phosphates in these soils. These soils present a high amount of organic matter as it is usual in forest soils and total nitrogen contents are low. Texture of acidic soils under Pinus sylvestris plots is mainly sandy-clay-loam while calcareous soils under Pinus halepensis plots mainly present a clay-loam textural class.
Both areas suffer from summer drought (mean hydric deficit of 254 mm for Pinus halepensis stands and 133 mm for Pinus sylvestris stands); mean annual temperature was 9.6 °C and mean annual precipitation 734 mm in Pinus sylvestris stands, while Pinus halepensis stands presented a mean annual temperature of 11.7 °C and mean annual precipitation of 456 mm.
2.2 Climatic data
The Digital Climatic Atlas for the Iberian Peninsula provided precipitation and temperature data for each plot (Ninyerola et al. 2005), which was used to calculate annual total precipitation, mean annual temperature, and hydric deficit for the plots. Hydric deficit was calculated as the sum of the differences between monthly potential evapotranspiration and precipitation for the months when potential evapotranspiration was higher than actual precipitation.
2.3 Sampling and soil analysis
Sampling points were established 5 m N, S, E, and W from the center of each plot. A composite sample of about 1 kg of mineral soil until 10 cm depth was collected from the four sampling points per plot after removing the organic horizon on top of it.
Soil samples were air-dried and sieved (2 mm mesh) to determine physical, chemical, and biochemical parameters. The preferred soil storage method prior to biochemical soil analyses is the conservation at field moist condition for less than 15 days at 4 °C (Alef and Nannipieri 1995). However, this preference varies among different countries depending on climate. Thus, Sweden and Finland usually freeze samples at − 20 °C, the UK, Denmark, Germany, and Switzerland prefer to store samples at 4 °C while in Italy, samples are air-dried (Bloem et al. 2006). Air-drying of soil samples provokes the death of soil organisms not adapted to hydric deficit (Mondini et al. 2002) but may be a proper alternative in areas subjected to summer drought, where microorganisms may be physiologically adapted to seasonal dryness (Zornoza et al. 2007). Moreover, several studies have found that microbial biomass content and enzyme activities determined on air-dried samples were representative of those detected using field-moist soil (Bandick and Dick 1999; Zornoza et al. 2007).
Total C and total N were ascertained through dry combustion using a LECO CHN 2000 analyzer. In soils with carbonates, total organic carbon was calculated as the difference between total and carbonate carbon. Soil total carbonates were determined by using of 1 M HCl titrated with 0.5 M NaOH (Bashour and Sayegh 2007). The Walkley and Black (1934) method was applied to analyze easily oxidizable C. Exchangeable cations (Ca, Mg, K, and Na) were analyzed by extraction with 1 M ammonium acetate (pH 7) and an atomic absorption/emission spectrometer. Water-soluble phenols were identified using the method developed by Box (1983). Soil pH was determined in a 1:2.5 (soil:water) suspension. Available P was extracted with anion exchange membranes (Turrión et al. 1997) and P content in the HCl extracts was determined through colorimetry, following the Murphy and Riley (1962) method.
The Isermeyer (1952) method was applied to determine the mineralizable C which is a quantification of the soil microbial metabolic activities (Nannipieri et al. 2002). It is estimated through the incubation of soil in closed jars and the mensuration of the amount of CO2 produced during organic matter oxidation at optimal conditions (Alef and Nannipieri 1995). Microbial biomass constitutes the most active part of soil organic matter. It is composed of living microorganisms that take part in nutrient mineralization and nutrient immobilization and act as a nutrient pool which are released and become available for plants and other microorganisms after their death (Duchaufour 1984; Jenkinson and Ladd 1981). Microbial biomass C, N, and P were determined using the fumigation-extraction method described by Vance et al. (1987). Fumigation was followed by extraction for P with 0.5 M NaHCO3 (pH 8.5) and P content was determined in both fumigated and unfumigated samples through colorimetry following the method described by Murphy and Riley (1962). Both fumigated and unfumigated samples also underwent extraction with 0.5 M K2SO4 for N and C and a TOC SKALAR autoanalyzer was used to determine C and N concentrations. The calculation of microbial biomass C, N, and P was done as the difference between the contents of these elements in the fumigated sample minus the content in the unfumigated sample. As some of the phosphorous released during fumigation is adsorbed by the soil colloid, a correction was done basing on the addition of a known quantity of phosphorous during the extraction and correcting for recovery (Brookes et al. 1982). Metabolic quotient was also calculated as the ratio between microbial activity and microbial biomass (qCO2 = mineralizable C/microbial biomass C). Metabolic quotient expresses the metabolic efficiency of soil microorganisms (Anderson and Domsch 1993).
Soil enzyme activities involved in C, N, and P biogeochemical cycles were also determined, including fluorescein diacetate hydrolysis reaction (Alef and Nannipieri 1995), dehydrogenase activity (Casida et al. 1964), and alkaline phosphatase, acid phosphatase (Tabatabai and Bremner 1969), urease (Hofmann 1963), and catalase (Beck 1971) activities.
2.4 Statistics
Some variables were not normally distributed after the removal of the outliers observed in the boxplots and even transformed (see supporting information 1 and 2), and then, a non-parametric test, the Wilcoxon-Mann-Whitney test, was performed to all the variables studied (wilcox.test in R) to assess whether they differed significantly in the soils under the two species studied. The Wilcoxon-Mann-Whitney test compares the medians of the studied groups. Median absolute deviation (MAD) was also calculated (in R: mad) as a measure of data dispersion:
Correlations between variables were studied using Spearman’s correlation coefficient (in R: cor.test, method = “spearman”) and principal component analysis (in R: princomp). Statistical analyses were performed with R software.
3 Results
Table 3 shows climatic and stand density data for the forest plots studied. Pinus halepensis plots had significantly higher mean annual temperature and hydric deficit than Pinus sylvestris plots, which presented significantly higher total precipitation and stand density.
No significant difference was detected between the two soil types in cation exchange capacity and total nitrogen (see Table 4). In the acidic soils, pH, K, Ca, and Mg were higher than in the calcareous soils. In contrast, easily oxidizable C, available P, total organic C to total N ratio, and water-soluble phenols were significantly higher in the acidic soils than in the calcareous soils.
Table 5 shows that no significant differences in fluorescein diacetate hydrolysis reaction were found in the soils under the two species. However, microbial biomass C and N and dehydrogenase, urease, and catalase activities were significantly higher in the calcareous soils under Pinus halepensis. We found no significant differences in alkaline phosphatase activity for the soils under each species, but acid phosphatase activity, mineralizable C, microbial biomass P, and qCO2 were significantly higher in the acidic soils under Pinus sylvestris.
The correlations between enzyme activities and microbial-related parameters are reported in Table 6 for Pinus halepensis plots and Table 8 for Pinus sylvestris plots.
In the calcareous soils under Pinus halepensis, microbial biomass C and N and mineralizable C were significantly correlated to all the enzyme activities studied except acid phosphatase. Only catalase and dehydrogenase activities were significantly correlated to microbial biomass P. No significant correlation was found between enzyme activities and Mg, K, pH, and mean annual temperature (Table 6) and acid phosphatase only correlated with easily oxidizable C, total precipitation, and hydric deficit. Other enzyme activities (fluorescein diacetate hydrolysis reaction and dehydrogenase, alkaline phosphatase, urease, and catalase) were significantly correlated with organic matter and nutrient-related parameters such as easily oxidizable C, total organic C to total N ratio, available P, total N, cation exchange capacity, and Ca. Significant correlations were also found between total precipitation and all enzyme activities except catalase. All enzyme activities except urease were significantly correlated to hydric deficit, but only alkaline phosphatase and catalase were significantly correlated to water-soluble phenols.
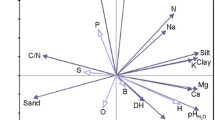
In the principal component analysis performed to the variables studied in calcareous soils under Pinus halepensis, the first two principal components accounted for more than 50% cumulated variance of the data. Table 7 presents the loadings of the variables in relation to the first two principal components. As can be seen in Table 7, the first principal component is negatively correlated to most biochemical parameters (enzyme activities such as fluorescein diacetate, dehydrogenase, urease, and catalase; microbial biomass C and N; mineralizable C) and nutrient-related parameters (cation exchange capacity, easily oxidizable C, available P, total N, and Ca). According to Fig. 1, which represents the biplot, biochemical parameters (enzyme activities, microbial biomass, and mineralizable C) and nutrient-related parameters are strongly correlated. The first principal component is correlated as well to hydric deficit and mean annual temperature and, therefore, biochemical and nutrient-related parameters are negatively correlated to mean annual temperature and hydric deficit, as previously observed in Spearman’s correlations.

Biplot of the principal component analysis performed on Pinus halepensis plots (calcareous soils) [CEC, cation exchange capacity; EOC, easily oxidizable C; MAT, mean annual temperature; qCO2, metabolic quotient; Cmic, Nmic, Pmic, microbial biomass C, N, and P; AcPhos, AlkPhos, acid and alkaline phosphatase; AP, available P; FDA, fluorescein diacetate hydrolysis reaction; DHA, dehydrogenase activity; C.N, total organic C to total N ratio; Deficit, hydric deficit]
With the exception of fluorescein diacetate hydrolysis reaction, a different trend was observed in the enzyme activities in acidic soils under Pinus sylvestris (Table 8), where dehydrogenase, acid phosphatase, and catalase were not significantly correlated to organic matter or nutrient-related parameters (easily oxidizable C, total organic C to total N ratio, available P, total N, cation exchange capacity, Ca, and K). Significant correlations between fluorescein diacetate hydrolysis reaction and microbial biomass C, N, and P and qCO2 were found, but catalase, urease, and dehydrogenase were significantly correlated to mineralizable C. Significant correlations were also found between alkaline phosphatase and microbial biomass P. Urease was significantly correlated to microbial biomass C and N and mineralizable C, and we found significant correlations between water-soluble phenols and fluorescein diacetate hydrolysis reaction and dehydrogenase and catalase activities. Catalase and urease activities were also significantly correlated to pH.
In the principal component analysis of variables studied in acidic soils under Pinus sylvestris plantations, the first two principal components accounted for almost 60% of cumulated variance. Table 9 presents the loadings of the variables studied in relation to the first two principal components. According to Table 9, the first principal component is negatively correlated to some biochemical parameters related to microbial biomass and activity (microbial biomass C, N, P, and mineralizable C), to some nutrient-related parameters (total N, available P, K, Ca, Mg), and to pH. The second principal component is positively correlated to the fluorescein diacetate hydrolysis reaction and some nutrient-related parameters (cation exchange capacity, easily oxidizable C) and negatively correlated to water-soluble phenols and hydric deficit. Figure 2 presents the biplot of the first two principal components in acidic soils under Pinus sylvestris plantations. According to Fig. 2, fluorescein diacetate hydrolysis reaction, which represent the total microbial hydrolytic activity in soils, is positively correlated to microbial biomass C, N, and P and negatively correlated to the amount of water-soluble phenols and metabolic quotient, as previously observed in Spearman’s correlations.

Biplot of the principal component analysis performed on Pinus sylvestris plots (acidic soils) [CEC, cation exchange capacity; EOC, easily oxidizable C; MAT, mean annual temperature; qCO2, metabolic quotient; Cmic, Nmic, Pmic, microbial biomass C, N, and P; AcPhos, AlkPhos, acid and alkaline phosphatase; AP, available P; FDA, fluorescein diacetate hydrolysis reaction; DHA, dehydrogenase activity; C.N, total organic C to total N ratio; Deficit, hydric deficit]
Data availability
The datasets generated during and/or analyzed during the current study are available in the Zenodo repository, URL: http://zenodo.org/record/345791#.WdymSDDtaUl (Bueis et al 2017c).
4 Discussion
4.1 Enzymatic and microbial activities
Our results indicate that soil enzymatic activities (dehydrogenase, urease, and catalase) and microbial biomass C and N tended to be higher in the calcareous soils under Pinus halepensis than in the acidic soils under Pinus sylvestris. Each forest species has different nutrient release and humification patterns that depend on litter quality and environmental factors. In our case, litter quality is similar: both forest ecosystems presented Pinus species with similar chemical composition in leaves, similar strategies for nutrient conservation in their tissues, and similar decomposition rates when other conditions affecting the process remained constant (Berg 2014; Bueis et al. 2017a; Carrasco et al. 2017; Cobertera 1993). Litter C/N ratio is very high in coniferous species and such is in these Pinus species (Cobertera 1993). The litter half-life of the studied species is about 24 months (Bueis et al. 2017a) and then, litter accumulates in the forest floor with an average thickness of 4 cm (unpublished results). Environmental factors must therefore be responsible for the differences in nutrient release and humification processes between the two types of soil. Our results indicated differences in climatic and edaphic properties between the two soil types. Correlations between enzymatic activities, microbial parameters, and environmental factors (soil and climatic characteristics) can help us in understanding the behavior of these soils.
Dehydrogenase activity was almost fifteen times lower in the acidic soils than in the calcareous soils. Since these enzymes intervene in soil processes that create metabolic pathways for soil microorganisms, they may give some idea of the potential of the soil to harbor biochemical activities which are crucial to soil fertility and health (Das and Varma 2011). The dehydrogenase enzyme is also frequently used as a direct measure of soil microbial activity in relation to mineralization and the formation of humic substances.
Many factors affect enzymatic activities in soils (Tabatabai 1994). Our results showed significant positive correlations of dehydrogenase, urease, and catalase activities with mineralizable C for both soil types when studied separately (Tables 6 and 8). Several authors have found significant correlations between enzyme activities and microbial biomass C and N and mineralizable C (García et al. 1994; Muscolo et al. 2015). In laboratory conditions, mineralizable C values were significantly higher in acidic soil than in calcareous soil which may be due to the significantly higher easily oxidizable C in these stands. Enzyme activities are also highly correlated to soil organic matter, which constitutes the energy source for microorganisms and can also contain stabilized enzymes (Alef and Nannipieri 1995; Lucas-Borja et al. 2012). We found significant positive correlations of dehydrogenase, urease, and catalase activities with easily oxidizable C in the calcareous soils, but not in the acidic soils studied. As easily oxidizable C increases in soil, so does microorganism activity and the decomposition rate of organic matter. This is reflected in soil respiration, indicating that dehydrogenase is positively correlated with easily oxidizable C content. The quality of organic matter, represented by total organic C to total N ratio, correlated significantly with urease activity in the acidic soils under Pinus sylvestris. Urease activity in this soil is therefore determined by N availability, as indicated by total N and total organic C to total N ratio, but not by the amount of organic matter (Alef and Nannipieri 1995; McCarty et al. 1992).
Acid phosphatase was significantly higher in the acidic soils under Pinus sylvestris, but alkaline phosphatase did not differ significantly between the two soil types. Acid phosphatase is usually higher in acidic soils, while alkaline phosphatase prevails in alkaline soil, but they can coexist (Burns 1978). Our results showed very low available P concentration in both soils. Phosphorus availability limits microbial biomass in some forest ecosystems (Scheu 1990) because P is immobilized by Al and Fe sesquioxides at low pH and by Ca at high pH (Gallardo and Schlesinger 1994). We found microbial biomass P to be three times higher than available P in both soils. Available P and microbial biomass P were also significantly higher in the acidic soils under Pinus sylvestris than in the calcareous soils studied. This suggests a high degree of P immobilization by microorganisms, which limits available P to plants but could be important for organic P mineralization in these Mediterranean forest soils.
In the acidic soils under Pinus sylvestris, significant correlations were found between urease and catalase activities and pH. Significant negative correlations were also found between fluorescein diacetate hydrolysis reaction, which is used as an indicator of general microbial activity in soil (Bandick and Dick 1999), and the concentration of water-soluble phenols. Low pH is known to limit bacterial communities (Blagodatskaya and Anderson 1998) and high water-soluble phenols can inhibit enzymatic activities. In the acidic soils under Pinus sylvestris, no significant correlations were found between dehydrogenase activity and microbial biomass, indicating that part of the soil microbial biomass is inactive due to inhibitory factors such as low pH (which limits bacterial forms) or high water-soluble phenols, which affects the amount and activity of soil microbial decomposers (Hattenschwiler and Vitousek 2000). It might also be related to the presence of enzymes of plant rather than microbial origin; plant roots can also exude enzymes and plant density was significantly higher in Pinus sylvestris stands (see Table 3).
Significant and negative correlations have also been found between enzyme activities and hydric deficit in calcareous soils, but not in acidic soils. Our results indicated that the higher the hydric deficit, the lower the enzyme activity in Mediterranean ecosystems under dry conditions such as those of the calcareous soils studied. In the long term, decreased soil enzyme activities will affect soil nutrient availability by reducing the nutrient supply to plants. Hydric deficit was not significantly correlated with any enzyme activity in the acidic soils under Pinus sylvestris, where the mean annual rainfall was around 750 mm, so hydric deficit was not a limiting factor for enzyme activity there. Lucas-Borja et al. (2012) also found higher microbial biomass and activities in areas with higher precipitations in pine forests in central Spain. No significant correlation was detected between enzyme activities and mean annual temperature, indicating that temperature did not influence enzyme activities in the soils under either Pinus species.
The metabolic quotient (qCO2), which expresses the amount of C released as CO2 by microbial respiration per unit microbial biomass, was significantly higher in the soils under Pinus sylvestris. Microorganisms use only part of the C contained in the substrates for growth and the maintenance of microbial structures; the rest is released into the atmosphere as CO2. Thus, qCO2 reflects microbial efficiency and can be interpreted as a measure of stress, because greater amounts of CO2 are produced under stressed conditions (Gonzalez-Quinones et al. 2011). The stressful soil conditions under Pinus sylvestris are likely related to low pH (3.7 to 5.6), a condition known to inhibit microbial activities.
Microbial biomass C was found to be significantly higher in the calcareous soils under Pinus halepensis even when easily oxidizable C and available P were significantly higher in the acidic soils under Pinus sylvestris. Correlations between organic matter and nutrient-related parameters in soil indicate microbial biomass dependence on an energy source and association with organic matter (Muscolo et al. 2015). Soil pH strongly influences microbial biomass, activity, and composition. These results suggest that pH limits both P availability and the bacterial community in the highly acidic soils under the Pinus sylvestris plantations.
4.2 Forest management
Forest management for sustainability must assess measures to improve ecosystem functioning, which crucially involves nutrient cycling and humification processes and the soil enzymes responsible for them. This work provides knowledge that can inform managerial alternatives which may improve soil nutrient conditions by enhancing enzyme performance. Low pH significantly limits enzyme activities in the acidic soils under Pinus sylvestris plantations, but soil pH may be modified in several ways. The most natural proposal consists of transitioning to mixed stands by promoting the growth of native broadleaf species such as Quercus pyrenaica Willd., as the litter inputs from this species may increase soil pH (Marcos et al. 2010). However, further studies are needed to confirm whether broadleaf species in these stands would actually or sufficiently increase soil pH and enzyme activities.
Enzyme activities in the calcareous soils under Pinus halepensis plantations seem to be mainly limited by hydric deficit. Forest management alternatives may involve modulating stand density to minimize tree competition for water (Ameztegui et al. 2017). However, the idea should be weighed carefully, as these stands primarily serve as protection against erosion. Extremely low densities may threaten soil retention and increase evaporation as more radiation reaches the soil. Soil preparation in new afforestation projects may also improve soil water availability (Querejeta et al. 2001). Creating suitable micro-topography, especially on steep slopes, may help water percolate into the soil where it is available for plant roots. Again, additional studies are required to determine optimal stand densities and soil preparation techniques for maximizing water availability.
5 Conclusion
Significant differences in enzyme activities were found between the calcareous soils under Pinus halepensis plantations and the acidic soils under Pinus sylvestris plantations. Soil organic matter and nutrient-related parameters were significantly correlated to most microbial parameters. The soils under Pinus sylvestris presented low pH and high amounts of water-soluble phenols, both of which limit the activity and composition of the microbial community. However, hydric deficit seemed to be the most limiting factor for enzyme activities in the calcareous soils under Pinus halepensis. Over time, decreased soil enzyme activity will affect mineralization and humification processes and, therefore, soil nutrient availability.
The findings from this research have informed the following managerial proposals for improving enzyme activities in these soils. For acidic soils under Pinus sylvestris plantations, we suggest promoting the growth of native broadleaf species such as Quercus pyrenaica in the stands, as litter from this species trees may improve soil pH and thereby encourage increased enzyme activities. For calcareous soils under Pinus halepensis plantations, we propose taking enzyme activities into account in decision-making for new afforestation projects, which may involve adjusting stand density and improving soil preparation to facilitate water filtration into the soil, especially in areas with steep slopes.
References
Alef K, Nannipieri P (1995) Methods in applied soil microbiology and biochemistry. Academic Press, London
Ameztegui A, Cabon A, De Cáceres M, Coll L (2017) Managing stand density to enhance the adaptability of Scots pine stands to climate change: a modelling approach. Ecol Model 356:141–150. https://doi.org/10.1016/j.ecolmodel.2017.04.006
Anderson TH, Domsch KH (1993) The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental-conditions, such as pH, on the microbial biomass of forest soils. Soil Biol Biochem 25:393–395. https://doi.org/10.1016/0038-0717(93)90140-7
Bandick AK, Dick RP (1999) Field management effects on soil enzyme activities. Soil Biol Biochem 31:1471–1479. https://doi.org/10.1016/s0038-0717(99)00051-6
Bashour I I, Sayegh A H (2007) Methods of analysis for soils of arid and semi-arid regions. Food and Agriculture Organization of the United Nations, Rome
Beck T (1971) Die Messung der Katalaseaktivität von Böden. Z Pflanzenernahrung dungung und bodenkunde 130:68–81. https://doi.org/10.1002/jpln.19711300108
Berg B (2014) Foliar litter decomposition: a conceptual model with focus on pine (Pinus) litter—a genus with global distribution. ISRN Forestry 2014:22–22. https://doi.org/10.1155/2014/838169
Blagodatskaya EV, Anderson TH (1998) Interactive effects of pH and substrate quality on the fungal-to-bacterial ratio and QCO(2) of microbial communities in forest soils. Soil Biol Biochem 30:1269–1274. https://doi.org/10.1016/s0038-0717(98)00050-9
Bloem J, Hopkins DW, Benedetti A (2006) Microbiological methods for assessing soil quality. CABI Publishing, Wallingford
Box JD (1983) Investigation of the Folin-Ciocalteau phenol reagent for the determination of polyphenolic substances in natural-waters. Water Res 17:511–525. https://doi.org/10.1016/0043-1354(83)90111-2
Brookes PC, Powlson DS, Jenkinson DS (1982) Measurement of microbial biomass phosphorus in soil. Soil Biol Biochem 14:319–329. https://doi.org/10.1016/0038-0717(82)90001-3
Bueis T, Bravo F, Pando V, Turrión MB (2016) Relationship between environmental parameters and Pinus sylvestris L. site index in forest plantations in northern Spain acidic plateau. Iforest-Biogeosci Forestry 9:394–401. https://doi.org/10.3832/ifor1600-008
Bueis T, Bravo F, Pando V, Turrion MB (2017a) Influencia de la densidad del arbolado sobre el desfronde y su reciclado en pinares de repoblación del norte de España. Bosque 38:401–407. https://doi.org/10.4067/S0717-92002017000200017
Bueis T, Bravo F, Pando V, Turrion MB (2017b) Site factors as predictors for Pinus halepensis Mill. productivity in Spanish plantations. Ann For Sci 74:6. https://doi.org/10.1007/s13595-016-0609-7
Bueis T, Turrión MB, Bravo F, Pando V, Muscolo A (2017c) Dataset of soil, climatic and stand variables in Pinus sylvestris and Pinus halepensis plantations in Spain [Dataset]. Zenodo. https://doi.org/10.5281/zenodo.345791
Burns RG (1978) Soil enzymes. Academic Press, London
Carrasco B, Cabaneiro A, Fernandez I (2017) Exploring potential pine litter biodegradability as a natural tool for low-carbon forestry. For Ecol Manag 401:166–176. https://doi.org/10.1016/j.foreco.2017.07.003
Casida LE Jr, Klein DA, Santoro T (1964) Soil dehydrogenase activity. Soil Sci 98:371–376
Cobertera E (1993) Edafología aplicada: suelos, producción agraria, planificación territorial e impactos ambientales
Das SK, Varma A (2011) Role of enzymes in maintaining soil health. In: Shukla G (ed) Soil enzymology. Soil Biology, vol 22. Springer, Berlin Heidelberg, pp 25–42
Duchaufour P (1984) Edafología 1. Edafogénesis y Clasificación. Masson, Barcelona, Spain
Gallardo A, Schlesinger WH (1994) Factors limiting microbial biomass in the mineral soil and forest floor of a warm-temperate forest. Soil Biol Biochem 26:1409–1415. https://doi.org/10.1016/0038-0717(94)90225-9
Garcia C, Hernandez T (1997) Biological and biochemical indicators in derelict soils subject to erosion. Soil Biol Biochem 29:171–177. https://doi.org/10.1016/s0038-0717(96)00294-5
García C, Hernández T, Costa F (1994) Microbial activity in soils under Mediterranean environmental conditions. Soil Biol Biochem 26:1185–1191. https://doi.org/10.1016/0038-0717(94)90142-2
Gartzia-Bengoetxea N, Gonzalez-Arias A, Kandeler E, de Arano IM (2009) Potential indicators of soil quality in temperate forest ecosystems: a case study in the Basque Country. Ann For Sci 66:303. https://doi.org/10.1051/forest/2009008
Gil-Sotres F, Trasar-Cepeda C, Leirós MC, Seoane S (2005) Different approaches to evaluating soil quality using biochemical properties. Soil Biol Biochem 37:877–887. https://doi.org/10.1016/j.soilbio.2004.10.003
Gonzalez-Quinones V, Stockdale EA, Banning NC, Hoyle FC, Sawada Y, Wherrett AD, Jones DL, Murphy DV (2011) Soil microbial biomass—interpretation and consideration for soil monitoring. Soil Res 49:287–304. https://doi.org/10.1071/sr10203
Hattenschwiler S, Vitousek PM (2000) The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ecol Evol 15:238–243. https://doi.org/10.1016/s0169-5347(00)01861-9
Hofmann E (1963) Urease. In: Bergmeyer H-U (ed) Methods of enzymatic analysis. Academic Press, New York
IGME (1975) Mapa Geológico de España. Escala 1/50 000. Instituto Geológico y Minero de España,
Isermeyer H (1952) Eine einfache Methode sur Bestimmung der Bodenatmung und der Carbonate im Boden. Z Pflanzenernähr Bodenkd 56:26–38
Jenkinson D S (1977) The soil biomass New Zealand Soil News 25:213–218
Jenkinson DS, Ladd JN (1981) Microbial biomass in soils: measurement and turnover. In: Paul EA, Ladd JN (eds) Soil biochemistry, vol 5. Marcel Dekker, New York, pp 415–417
Llorente M, Turrion MB (2010) Microbiological parameters as indicators of soil organic carbon dynamics in relation to different land use management. Eur J For Res 129:73–81. https://doi.org/10.1007/s10342-008-0249-z
Lucas-Borja ME, Candel Pérez D, López Serrano FR, Andrés M, Bastida F (2012) Altitude-related factors but not Pinus community exert a dominant role over chemical and microbiological properties of a Mediterranean humid soil. Eur J Soil Sci 63:541–549. https://doi.org/10.1111/j.1365-2389.2012.01438.x
Marcos E, Calvo L, Marcos JA, Taboada A, Tarrega R (2010) Tree effects on the chemical topsoil features of oak, beech and pine forests. Eur J For Res 129:25–30. https://doi.org/10.1007/s10342-008-0248-0
Martínez-Salgado MM, Gutiérrez-Romero V, Jannsens M, Ortega-Blu R (2010) Biological soil quality indicators: a review. In: Mendez-Vilas A (ed) Current research technology and education topics in applied microbiology and microbial biotechnology. Formatex Research Center, Spain, pp 319–328
McCarty GW, Shorgen DR, Bremner JM (1992) Regulation of urease production in soil by microbial assimilation of nitrogen. Biol Fertil Soils 12:261–264. https://doi.org/10.1007/BF00336041
Mondini C, Contin M, Leita L, De Nobili M (2002) Response of microbial biomass to air-drying and rewetting in soils and compost. Geoderma 105:111–124. https://doi.org/10.1016/s0016-7061(01)00095-7
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphorus in natural waters. Anal Chim Acta 27:31–36. https://doi.org/10.1016/s0003-2670(00)88444-5
Muscolo A, Sidari M, Mercurio R (2007) Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine (Pinus laricio, Poiret) stands. For Ecol Manag 242:412–418. https://doi.org/10.1016/j.foreco.2007.01.058
Muscolo A, Settineri G, Attina E (2015) Early warning indicators of changes in soil ecosystem functioning. Ecol Indic 48:542–549. https://doi.org/10.1016/j.ecolind.2014.09.017
Nannipieri P, Kandeler E, Ruggiero P (2002) Enzyme activities and microbiological and biochemical processes in soil. In: Burns RG, Dick RP (eds) Enzymes in the environment. Marcel Dekker, New York, pp 1–33
Nannipieri P, Giagnoni L, Landi L, Renella G (2011) Role of phosphatase enzymes in soil. In: Bünemann E, Oberson A, Frossard E (eds) Phosphorus in action: biological processes in soil phosphorus cycling. Springer Berlin Heidelberg, Berlin, Heidelberg, pp 215–243. https://doi.org/10.1007/978-3-642-15271-9_9
Ninyerola M, Pons i Fernández X, Roure J M (2005) Atlas climático digital de la Península Ibérica: metodología y aplicaciones en bioclimatología y geobotánica. Universidad Autónoma de Barcelona. Bellaterra, Barcelona, Spain
Olander LP, Vitousek PM (2000) Regulation of soil phosphatase and chitinase activity by N and P availability. Biogeochemistry 49:175–190. https://doi.org/10.1023/a:1006316117817
Querejeta JI, Roldán A, Albaladejo J, V c C (2001) Soil water availability improved by site preparation in a Pinus halepensis afforestation under semiarid climate. For Ecol Manag 149:115–128. https://doi.org/10.1016/S0378-1127(00)00549-1
Scheu S (1990) Changes in microbial nutrient status during secondary succession and its modification by earthworms. Oecologia 84:351–358. https://doi.org/10.1007/BF00329758
Sinsabaugh RL (1994) Enzymic analysis of microbial pattern and process. Biol Fertil Soils 17:69–74. https://doi.org/10.1007/bf00418675
Tabatabai MA (1994) Soil enzymes. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis. American Society of Agronomy, Madison, pp 775–833
Tabatabai MA, Bremner JM (1969) Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol Biochem 1:301–307. https://doi.org/10.1016/0038-0717(69)90012-1
Turrión MB, Gallardo JF, González MI (1997) Nutrient availability in forest soils as measured with anion exchange membranes. Geomicrobiol J 14:51–64. https://doi.org/10.1080/01490459709378033
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707. https://doi.org/10.1016/0038-0717(87)90052-6
Walkley A, Black IA (1934) An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci 37:29–38. https://doi.org/10.1097/00010694-193401000-00003
Yang K, Zhu JJ, Yan QL, Zhang JX (2012) Soil enzyme activities as potential indicators of soluble organic nitrogen pools in forest ecosystems of Northeast China. Ann For Sci 69:795–803. https://doi.org/10.1007/s13595-012-0198-z
Zornoza R, Guerrero C, Mataix-Solera J, Arcenegui V, Garcia-Renes F, Mataix-Beneyto J (2007) Assessing the effects of air-drying and rewetting pre-treatment on soil microbial biomass, basal respiration, metabolic quotient and soluble carbon under Mediterranean conditions. Eur J Soil Biol 43:120–129. https://doi.org/10.1016/j.ejsobi.2006.11.004
Acknowledgements
The authors thank Elisa Mellado, Temesgen Desalegn, Olga López, and Carlos Alejandro Mendoza for their assistance in the field and Carmen Blanco, Juan Carlos Arranz, Carmelo Malamaci, and Maria Sidari for their advice in laboratory analysis.
Funding
This work was supported by the University of Valladolid and Banco Santander (predoctoral grant to T. Bueis), the Mediterranean Regional Office of the European Forest Institute (EFIMED; “Short Scientific Visit” grant to T. Bueis), and the Ministry of Economy and Competitiveness of the Spanish Government (AGL2011-29701-C02-02 and AGL2014-51964-C2-1-R).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Erwin Dreyer
Contribution of the co-authors
Teresa Bueis: designed the experiment, carried out the field and laboratory work, run the data analysis, discussed the results, and wrote the paper
Felipe Bravo: designed the experiment, coordinated the research project, and corrected the manuscript
Valentín Pando: supported the statistical analysis and corrected the manuscript
María-Belén Turrión: designed the experiment, supervised the laboratory analysis, and corrected the manuscript
Adele Muscolo: supervised the laboratory analysis and corrected the manuscript
This article is part of the Topical Collection on Mediterranean pines
Rights and permissions
About this article

Cite this article
Bueis, T., Turrión, M.B., Bravo, F. et al. Factors determining enzyme activities in soils under Pinus halepensis and Pinus sylvestris plantations in Spain: a basis for establishing sustainable forest management strategies. Annals of Forest Science 75, 34 (2018). https://doi.org/10.1007/s13595-018-0720-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-018-0720-z