
Abstract
Mammalian target of rapamycin (mTOR) plays essential roles in cell proliferation, survival and metabolism by forming at least two functional distinct multi-protein complexes, mTORC1 and mTORC2. External growth signals can be received and interpreted by mTORC2 and further transduced to mTORC1. On the other hand, mTORC1 can sense inner-cellular physiological cues such as amino acids and energy states and can indirectly suppress mTORC2 activity in part through phosphorylation of its upstream adaptors, IRS-1 or Grb10, under insulin or IGF-1 stimulation conditions. To date, upstream signaling pathways governing mTORC1 activation have been studied extensively, while the mechanisms modulating mTORC2 activity remain largely elusive. We recently reported that Sin1, an essential mTORC2 subunit, was phosphorylated by either Akt or S6K in a cellular context-dependent manner. More importantly, phosphorylation of Sin1 at T86 and T398 led to a dissociation of Sin1 from the functional mTORC2 holo-enzyme, resulting in reduced Akt activity and sensitizing cells to various apoptotic challenges. Notably, an ovarian cancer patient-derived Sin1-R81T mutation abolished Sin1-T86 phosphorylation by disrupting the canonical S6K-phoshorylation motif, thereby bypassing Sin1-phosphorylation-mediated suppression of mTORC2 and leading to sustained Akt signaling to promote tumorigenesis. Our work therefore provided physiological and pathological evidence to reveal the biological significance of Sin1 phosphorylation-mediated suppression of the mTOR/Akt oncogenic signaling, and further suggested that misregulation of this process might contribute to Akt hyper-activation that is frequently observed in human cancers.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
mTORC1 and mTORC2 are functionally distinct mTOR-containing protein kinase complexes
Mammalian target of rapamycin (mTOR) is a highly conserved protein kinase (van Dam et al., 2011) belonging to the phosphatidylinositol-3-kinase-related protein kinase (PIKK) super-family. Biologically, mTOR is a master regulator of cellular homeostasis, cell growth and proliferation as well as metabolism in a broad range of physiological (Wullschleger et al., 2006; Zoncu et al., 2011b) and pathological settings, including diabetes and cancer (Guertin and Sabatini, 2007; Inoki et al., 2005). In doing so, mTOR serves as a central sensor for cellular physiological cues such as growth signals and nutrients, by functioning as an essential catalytic subunit in two functionally distinct complexes, mTOR complex 1 (mTORC1) (Kim et al., 2002) and mTOR complex 2 (mTORC2) (See (Sabatini, 2006) and (Wullschleger et al., 2006) for review). These two complexes are distinguished by their accessory protein composition, localization and function. Specifically, both mTORC1 and mTORC2 share the common components mTOR and GβL/mLST8, while the unique subunit Raptor defines mTORC1 (Kim et al., 2002) whereas Rictor (Sarbassov et al., 2004) and Sin1 (Frias et al., 2006; Guertin et al., 2006; Jacinto et al., 2006; Wullschleger et al., 2005; Yang et al., 2006) define mTORC2. mTORC1 was reported to localize on the outer membrane of lysosome, which is critical for its activation and physiological function (Bar-Peled and Sabatini, 2012; Thoreen et al., 2012), while mTORC2 association with ribosomes was recently demonstrated to be critical for mTORC2 activity (Zinzalla et al., 2011).
In echoing their unique subcellular localizations, mTORC1 and mTORC2 display distinct cellular functions by targeting different sets of downstream effectors for phosphorylation and function. Notably, mTORC1 promotes cap-dependent mRNA translation (Thoreen et al., 2012) and protein synthesis through direct phosphorylation and activation of its bona fide substrates S6K1, 4EBP1 and TFEB1 (Ma and Blenis, 2009; Pena-Llopis et al., 2011), inhibits autophagy through phosphorylation of Ulk1 (Chan, 2009) and establishes Treg-cell function by facilitating cholesterol and lipid metabolism through yet undefined phosphorylation targets (Zeng et al., 2013). On the other hand, mTORC2 was firstly identified to regulate cellular skeletal organization (Jacinto et al., 2004) and later shown to be indispensible in governing cell growth, proliferation, survival and anabolism, which are mainly through direct phosphorylation and activation of its physiological downstream targets including Akt (S473) (Sarbassov et al., 2005), SGK (S422) (Garcia-Martinez and Alessi, 2008) and PKCα (Ikenoue et al., 2008). Moreover, as mTOR serves as an oncogenic pathway to promote cellular growth and survival, deregulation of many components of the mTOR pathway has been implicated in human cancer and metabolic diseases (Weber and Gutmann, 2012; Zoncu et al., 2011b).
mTORC1 and mTORC2 are differentially regulated in cells
Previous work clearly established that both activation of mTORC1 and mTORC2 are tightly, yet differentially controlled (Laplante and Sabatini, 2012; Weber and Gutmann, 2012). Mechanistically, when stimulated by extra-cellular growth signals, mTORC2 receives activation signals from the Ras/PI3K signaling pathway through undefined mechanisms, and activates mTORC1 by Akt-dependent phosphorylation of TSC2 (Inoki et al., 2002; Manning et al., 2002) or PRAS40 (Vander Haar et al., 2007), releasing their repression on mTORC1. In addition, mTORC1 can also sense cellular energy states and amino acid levels independent of mTORC2. Specifically, when cellular AMP levels are high, which indicates low energy status in cells, the LKB1/AMPK (AMP-activated serine/threonine protein kinase) pathway becomes activated, leading to inhibition of mTORC1 (Shaw, 2009), either by phosphorylation of TSC2 (Inoki et al., 2003) or by phosphorylation of the essential mTORC1 component Raptor (Gwinn et al., 2008). Furthermore, mTORC1, but not mTORC2, serves as an inner cellular amino acid sensor (Efeyan et al., 2012) dependent on its recruitment to lysosome surface by Rag (Sancak et al., 2010; Sancak et al., 2008), Ragulator (Bar-Peled et al., 2012) and the lysosome vacuolar H+-ATPase (Zoncu et al., 2011a) through an inside-out mechanism to control the timely activation of mTORC1.
The differential regulatory mechanisms of these two mTOR-containing complexes extend further to their altered responses to rapamycin. mTORC1 is sensitive to low-dose rapamycin treatment in both cell culture and mouse models, where rapamycin directly binds to FKBP12 and disrupts the interaction between Raptor and mTOR, suppressing mTORC1 assembly and activation (Oshiro et al., 2004; Yip et al., 2010). On the other hand, mTORC2 only responds to prolonged and chronic rapamycin treatment, in part because rapamycin cannot directly interfere with existing mTORC2 complex, but rather only blocks the assembly of mTORC2 from newly synthesized Rictor and mTOR (Sarbassov et al., 2006).
More importantly, mTORC1 could also be activated independent of mTORC2. Mice with deleted S6K1 display elevated resistance to high-fat diet and weight gain, in part due to deficiency in adipocytes generation (Carnevalli et al., 2010). Adipose specific Raptor knockout mice phenocopied the S6K1 knockout mice with regard to adipocyte generation (Polak et al., 2008), highlighting the critical role of mTORC1 in adipogenesis in vivo. However, Rictor knockout mice showed no defects in adipogenesis (Kumar et al., 2010), although an mTORC2 substrate, BTSA (a BSD domain-containing protein) has been characterized to be indispensible for adipogenesis (Kumar et al., 2010). Taken together, mTORC1 and mTORC2 could function synergistically or independently to maintain cell physiology.
mTORC1 negatively regulates mTORC2 activation through indirect mechanisms mediated by phosphorylation of IRS-1 and GRB10
mTORC1 has also been shown to indirectly suppress mTORC2 signaling that in TSC2-/- cells with elevated mTORC1/S6K activity, Akt activation was significantly reduced (Manning et al., 2005). Further studies revealed that elevated mTORC1/S6K could suppress the activation of the PI3K pathway through multiple negative feedback loops (Gual et al., 2005). Specifically, mTORC1 and S6K can phosphorylate IRS-1 to block its interaction with the p85 regulatory subunit of PI3K to negatively regulate the insulin-signaling pathway (Gual et al., 2005). More recently, both the Blennis (Yu et al., 2011) and Sabatini (Hsu et al., 2011) groups independently showed that Grb10 is phosphorylated and stabilized by mTORC1 to block the PI3K signaling. However, in response to certain stimuli such as PDGF and EGF, activation of mTORC2/Akt was not affected by depletion of Grb10, suggesting the presence of an additional mechanism for mTORC1 to suppress mTORC2. As genetic disruption of either Rictor or Sin1 results in the inactivation of mTORC2 (Guertin et al., 2006; Jacinto et al., 2006), we reasoned that mTORC1/S6K-dependent regulation of mTORC2 might occur through these unique mTORC2 components in a direct manner. However, we and others previously showed that AGC kinases-mediated phosphorylation of Rictor at T1135 does not significantly affect mTORC2 complex integrity and its kinase activity (Dibble et al., 2009; Gao et al., 2010), which urged us to further examine whether the other mTORC2 essential component, Sin1, is a major target to mediate mTORC1/S6K’s negative regulation of mTORC2.
mTORC1 negatively regulates mTORC2 activation through direct mechanisms mediated by phosphorylation of SIN1
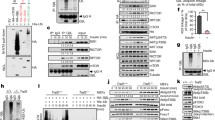
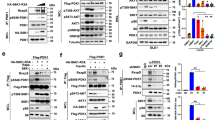
To test if modulation of Sin1 is important for mTORC2 activity, we and others tested if Sin1 could be targeted for phosphorylation by Akt or S6K at either T86 and T398, or both sites, in adipocytes (Humphrey et al., 2013) or epithelial cells (Liu et al., 2013), respectively. Importantly, we found that phosphorylation of Sin1 on both the T86 and T398 sites led to a disassociation of Sin1 from other mTORC2 components, revealing a direct negative regulatory mechanism for mTORC2 governed by mTORC1 (Liu et al., 2013). Mechanistically, we observed that Sin1 utilized various domains to interact with mTORC2 components (Fig. 1), with its N-terminus binding to Rictor and GβL, RBD (Ras binding domain) binding to GβL, and the PH domain interacting with GβL and mTOR-KD (kinase domain). Interestingly, only the N and PH domains were capable of interacting with S6K1 (Fig. 1). Considering that the identified T86 and T398 phosphorylation sites are located in the N- and PH domains, respectively, it is therefore plausible that S6K could directly phosphorylate these two sites to influence mTORC2 integrity. In keeping with this notion, phosphorylation at T86 interfered with Sin1-N-terminus binding to Rictor, while phosphorylation at T398 impaired Sin1-PH domain interaction with the mTOR kinase domain. Importantly, Sin1 dissociation from the mTORC2 complex requires both phosphorylation events (Fig. 2). In addition, we demonstrated that Sin1 phosphorylation at both sites occurred in response to various external cellular stimuli including insulin, IGF1, PDGF and EGF, which significantly blocked the activation of Akt induced by these stimuli, proposing a novel negative feedback loop independent of IRS-1 and Grb10 to suppress mTORC2.

mTORC2 components and S6K interact with different domains of Sin1

Phosphorylation of Sin1 at both T86 and T398 sites dissociates Sin1 from functional mTORC2 complex and subsequently suppresses Akt activation
Physiologically, we demonstrated that Sin1 phosphorylation-mediated mTORC2 suppression on Akt led to increased levels of cleaved caspase 3 and subsequently sensitized cells to apoptosis-initiating agents such as etoposide or cisplatin. More importantly, we found that in freshly isolated splenic B cells, rapamycin treatment significantly reduced Sin1-T86 phosphorylation accompanied by an increase in Akt-S473 phosphorylation. A similar inverse correlation between phosphorylation of Sin1-T86 and Akt-S473 was also observed in mouse liver lysates, and rapamycin administration to whole mice led to reduced Sin1-pT86 concomitant with increased Akt-pS473 signals, further confirming the physiological significance of mTORC1-mediated negative regulation on mTORC2 function.
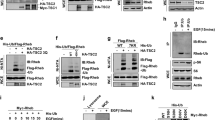
Pathologically, we screened a panel of human ovarian patient clinical samples by IHC and an inverse correlation between Sin1-pT86 and Akt-pS473 was observed in a certain number of cases. However, due to the limited number of patient samples available, the inverse correlation did not reach statistical significance, which warrants further larger-scale examinations. This inverse correlation was also observed in a panel of T-ALL cancer cell lines. Interestingly, two Sin1 mutations (R81T and S84L) were identified in ovarian cancer patients. Using biochemical assays we demonstrated that both of these mutations led to significantly reduced Sin1 phosphorylation at the T86 site, as these mutations impair the canonical AGC kinase consensus recognition motif “RxRxxpS/pT” (Alessi et al., 1996; Manning and Cantley, 2007). As phosphorylation at both T86 and T398 is necessary to dissociate Sin1 from the functional mTORC2 complex, substitutions at R81T or S84L diminished the possibility of co-occurrence of pT86 and pT398, resulting in stabilized mTORC2 integrity and function (Fig. 3). Consistent with this notion, compared to Sin1-WT, relatively sustained Akt phosphorylation was observed in Sin1-R81T expressing cells. At the cellular levels, expressing Sin1-R81T, opposing to the phospho-mimetic Sin1-T86E/T398E mutant, led to comparable levels of cleaved caspase 3 to cells expressing Sin1-WT upon etoposide or cisplatin challenge, subsequently conferring cellular resistance to these agents. More importantly, compared to Sin1-WT, Sin1-R81T expressing cells gained oncogenic ability to promote ovarian cancer cell growth in soft agar as well as in a xenograft mouse model. Together, these data consistently support a model that Sin1 phosphorylation plays critical roles in inhibiting mTORC2 oncogenic function in both physiological and pathological settings.

The cancer patient-derived Sin1-R81T mutation leads to stabilized mTORC2 integrity and sustained Akt activation to facilitate tumorigenesis
Implications of mTORC1-mediated suppression of mTORC2
Compared to the extensively studied regulations of the mTORC1 complex, the upstream regulatory signaling pathways governing mTORC2 activity remained largely to be determined. So far, the only defined upstream regulation of mTORC2 is the requirement for ribosome association for its activation, where the mechanisms remain largely uncharacterized (Zinzalla et al., 2011). As S6K and 4EBP-1, two characterized mTORC1 substrates, are indispensible for translation of capped mRNAs by ribosome, there might be some yet-to-be identified positive regulatory events governed by S6K or 4EBP-1, which are vital for mTORC2 activation on ribosome. Furthermore, given that both mTORC1 and mTOCR2 contain mTOR, it is not surprising that many layers of crosstalks might exist between these two complexes.
Recent studies have begun to reveal a complicated cross-communication between these two mTOR-containing complexes while the exact molecular mechanism(s) remain largely elusive. In this effort, we have defined a novel and independent negative feedback loop through which either S6K1 or Akt directly phosphorylates Sin1 to repress mTORC2 activation in epithelial cells or adipocytes, respectively, providing a possible molecular mechanism for mTORC1 to balance the extra- and intra- cellular signals. Moreover, systematic analyses of many signaling pathways revealed a general oscillation pattern of activation/inactivation signaling dynamics (Purvis et al., 2012; Purvis and Lahav, 2012, 2013). This also applied to mTORC2-mediated growth factor signaling pathways, as suggested by the periodic Akt activation pattern upon stimulation by insulin or growth factors (Purvis and Lahav, 2013). To this end, mTORC1-mediated inhibition of mTORC2 through Sin1 phosphorylation might be one of such mechanisms, in addition to de-phosphorylation of Akt and multiple other negative feedback mechanism, to ensure that mTORC2 is only activated in a “pulse” manner (Chen et al., 2012). Therefore, between these two mTOR-containing complexes, it is plausible that mTORC1 exhibits constant basal activity whereas the mTORC2 complex is only transiently activated following external stimuli. This may partially explain the fact that hyper-activation of the critical signaling pathways including mTORC2/Akt is a hallmark for majority of human cancers (Testa and Tsichlis, 2005).
Finally, the identification of this mTORC1/S6K feedback suppression of mTORC2 expanded the critical role of mTORC1 in regulating and coordinating various growth factor signaling pathways. Compared to the previously identified negative feedback regulatory loops via IRS-1 (Harrington et al., 2004; Shah and Hunter, 2006) or Grb10 (Hsu et al., 2011; Yu et al., 2011), which nicely explained the IGF-1 and insulin but not the PDGF or EGF signaling regulatory pathways, the mTORC1/S6K1-mediated phosphorylation of Sin1 could function to balance the mTORC2/Akt activation in response to a wider range of growth stimuli beyond insulin and IGF-1, including but not limited to PDGF and EGF. Moreover, by targeting the mTORC2-specific component Sin1, instead of IRS-1 or Grb10 that are upstream of both mTORC1 and mTORC2, for phosphorylation and inhibition refines the suppression effects mainly on mTORC2, suggesting that this newly identified mechanism might be a specific mTORC2-targeted negative regulation. Taken together, our study unravels a novel IRS-1/Grb10-independent feedback mechanism of the tightly regulated PI3K/mTORC2/Akt pathway. More significantly, an ovarian cancer patient-derived R81T mutation of Sin1 was demonstrated to gain oncogenic capacity by bypassing this negative regulation due to the lack of Sin1-T86 phosphorylation, providing a molecular mechanism for the elevated mTORC2/Akt activation that could potently promote tumorigenesis at least in this cancer patient. In summary, our work points to the prospect of targeting Sin1 phosphorylation signaling as an effective therapeutic strategy in treating human disorders such as diabetes and cancer.
References
Alessi DR, Caudwell FB, Andjelkovic M, Hemmings BA, Cohen P (1996) Molecular basis for the substrate specificity of protein kinase B; comparison with MAPKAP kinase-1 and p70 S6 kinase. FEBS Lett 399:333–338
Bar-Peled L, Sabatini DM (2012) SnapShot: mTORC1 signaling at the lysosomal surface. Cell 151(1390–1390):e1391
Bar-Peled L, Schweitzer LD, Zoncu R, Sabatini DM (2012) Ragulator is a GEF for the rag GTPases that signal amino acid levels to mTORC1. Cell 150:1196–1208
Carnevalli LS, Masuda K, Frigerio F, Le Bacquer O, Um SH, Gandin V, Topisirovic I, Sonenberg N, Thomas G, Kozma SC (2010) S6K1 plays a critical role in early adipocyte differentiation. Dev Cell 18:763–774
Chan EY (2009) mTORC1 phosphorylates the ULK1–mAtg13–FIP200 autophagy regulatory complex. Sci Signal 2:pe51
Chen JY, Lin JR, Cimprich KA, Meyer T (2012) A two-dimensional ERK–AKT signaling code for an NGF-triggered cell-fate decision. Mol Cell 45:196–209
Dibble CC, Asara JM, Manning BD (2009) Characterization of Rictor phosphorylation sites reveals direct regulation of mTOR complex 2 by S6K1. Mol Cell Biol 29:5657–5670
Efeyan A, Zoncu R, Sabatini DM (2012) Amino acids and mTORC1: from lysosomes to disease. Trends Mol Med 18:524–533
Frias MA, Thoreen CC, Jaffe JD, Schroder W, Sculley T, Carr SA, Sabatini DM (2006) mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s. Curr Biol 16:1865–1870
Gao D, Wan L, Inuzuka H, Berg AH, Tseng A, Zhai B, Shaik S, Bennett E, Tron AE, Gasser JA et al (2010) Rictor forms a complex with Cullin-1 to promote SGK1 ubiquitination and destruction. Mol Cell 39:797–808
Garcia-Martinez JM, Alessi DR (2008) mTOR complex 2 (mTORC2) controls hydrophobic motif phosphorylation and activation of serum- and glucocorticoid-induced protein kinase 1 (SGK1). Biochem J 416:375–385
Gual P, Le Marchand-Brustel Y, Tanti JF (2005) Positive and negative regulation of insulin signaling through IRS-1 phosphorylation. Biochimie 87:99–109
Guertin DA, Sabatini DM (2007) Defining the role of mTOR in cancer. Cancer Cell 12:9–22
Guertin DA, Stevens DM, Thoreen CC, Burds AA, Kalaany NY, Moffat J, Brown M, Fitzgerald KJ, Sabatini DM (2006) Ablation in mice of the mTORC components raptor, Rictor, or mLST8 reveals that mTORC2 is required for signaling to Akt–FOXO and PKCalpha, but not S6K1. Dev Cell 11:859–871
Gwinn DM, Shackelford DB, Egan DF, Mihaylova MM, Mery A, Vasquez DS, Turk BE, Shaw RJ (2008) AMPK phosphorylation of raptor mediates a metabolic checkpoint. Mol Cell 30:214–226
Harrington LS, Findlay GM, Gray A, Tolkacheva T, Wigfield S, Rebholz H, Barnett J, Leslie NR, Cheng S, Shepherd PR et al (2004) The TSC1-2 tumor suppressor controls insulin-PI3K signaling via regulation of IRS proteins. J Cell Biol 166:213–223
Hsu PP, Kang SA, Rameseder J, Zhang Y, Ottina KA, Lim D, Peterson TR, Choi Y, Gray NS, Yaffe MB et al (2011) The mTOR-regulated phosphoproteome reveals a mechanism of mTORC1-mediated inhibition of growth factor signaling. Science 332:1317–1322
Humphrey SJ, Yang G, Yang P, Fazakerley DJ, Stockli J, Yang JY, James DE (2013) Dynamic adipocyte phosphoproteome reveals that Akt directly regulates mTORC2. Cell Metab 17:1009–1020
Ikenoue T, Inoki K, Yang Q, Zhou X, Guan KL (2008) Essential function of TORC2 in PKC and Akt turn motif phosphorylation, maturation and signalling. EMBO J 27:1919–1931
Inoki K, Li Y, Zhu T, Wu J, Guan KL (2002) TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat Cell Biol 4:648–657
Inoki K, Zhu T, Guan KL (2003) TSC2 mediates cellular energy response to control cell growth and survival. Cell 115(5):577–590
Inoki K, Corradetti MN, Guan KL (2005) Dysregulation of the TSC–mTOR pathway in human disease. Nat Genet 37:19–24
Jacinto E, Loewith R, Schmidt A, Lin S, Ruegg MA, Hall A, Hall MN (2004) Mammalian TOR complex 2 controls the actin cytoskeleton and is rapamycin insensitive. Nat Cell Biol 6:1122–1128
Jacinto E, Facchinetti V, Liu D, Soto N, Wei S, Jung SY, Huang Q, Qin J, Su B (2006) SIN1/MIP1 maintains Rictor–mTOR complex integrity and regulates Akt phosphorylation and substrate specificity. Cell 127:125–137
Kim DH, Sarbassov DD, Ali SM, King JE, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2002) mTOR interacts with raptor to form a nutrient–sensitive complex that signals to the cell growth machinery. Cell 110:163–175
Kumar A, Lawrence JC Jr, Jung DY, Ko HJ, Keller SR, Kim JK, Magnuson MA, Harris TE (2010) Fat cell-specific ablation of Rictor in mice impairs insulin-regulated fat cell and whole-body glucose and lipid metabolism. Diabetes 59:1397–1406
Laplante M, Sabatini DM (2012) mTOR signaling in growth control and disease. Cell 149:274–293
Liu P, Gan W, Inuzuka H, Lazorchak AS, Gao D, Arojo O, Liu D, Wan L, Zhai B, Yu Y et al (2013) Sin1 phosphorylation impairs mTORC2 complex integrity and inhibits downstream Akt signalling to suppress tumorigenesis. Nat Cell Biol 15:1340–1350
Ma XM, Blenis J (2009) Molecular mechanisms of mTOR-mediated translational control. Nat Rev Mol Cell Biol 10:307–318
Manning BD, Cantley LC (2007) AKT/PKB signaling: navigating downstream. Cell 129:1261–1274
Manning BD, Tee AR, Logsdon MN, Blenis J, Cantley LC (2002) Identification of the tuberous sclerosis complex-2 tumor suppressor gene product tuberin as a target of the phosphoinositide 3-kinase/akt pathway. Mol Cell 10:151–162
Manning BD, Logsdon MN, Lipovsky AI, Abbott D, Kwiatkowski DJ, Cantley LC (2005) Feedback inhibition of Akt signaling limits the growth of tumors lacking Tsc2. Genes Dev 19:1773–1778
Oshiro N, Yoshino K, Hidayat S, Tokunaga C, Hara K, Eguchi S, Avruch J, Yonezawa K (2004) Dissociation of raptor from mTOR is a mechanism of rapamycin-induced inhibition of mTOR function. Genes Cells 9:359–366
Pena-Llopis S, Vega-Rubin-de-Celis S, Schwartz JC, Wolff NC, Tran TA, Zou L, Xie XJ, Corey DR, Brugarolas J (2011) Regulation of TFEB and V-ATPases by mTORC1. EMBO J 30:3242–3258
Polak P, Cybulski N, Feige JN, Auwerx J, Ruegg MA, Hall MN (2008) Adipose-specific knockout of raptor results in lean mice with enhanced mitochondrial respiration. Cell Metab 8:399–410
Purvis JE, Lahav G (2012) Decoding the insulin signal. Mol Cell 46:715–716
Purvis JE, Lahav G (2013) Encoding and decoding cellular information through signaling dynamics. Cell 152:945–956
Purvis JE, Karhohs KW, Mock C, Batchelor E, Loewer A, Lahav G (2012) p53 dynamics control cell fate. Science 336:1440–1444
Sabatini DM (2006) mTOR and cancer: insights into a complex relationship. Nat Rev Cancer 6:729–734
Sancak Y, Peterson TR, Shaul YD, Lindquist RA, Thoreen CC, Bar-Peled L, Sabatini DM (2008) The Rag GTPases bind raptor and mediate amino acid signaling to mTORC1. Science 320:1496–1501
Sancak Y, Bar-Peled L, Zoncu R, Markhard AL, Nada S, Sabatini DM (2010) Ragulator–Rag complex targets mTORC1 to the lysosomal surface and is necessary for its activation by amino acids. Cell 141:290–303
Sarbassov DD, Ali SM, Kim DH, Guertin DA, Latek RR, Erdjument-Bromage H, Tempst P, Sabatini DM (2004) Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr Biol 14:1296–1302
Sarbassov DD, Guertin DA, Ali SM, Sabatini DM (2005) Phosphorylation and regulation of Akt/PKB by the Rictor–mTOR complex. Science 307:1098–1101
Sarbassov DD, Ali SM, Sengupta S, Sheen JH, Hsu PP, Bagley AF, Markhard AL, Sabatini DM (2006) Prolonged rapamycin treatment inhibits mTORC2 assembly and Akt/PKB. Mol Cell 22:159–168
Shah OJ, Hunter T (2006) Turnover of the active fraction of IRS1 involves raptor-mTOR- and S6K1-dependent serine phosphorylation in cell culture models of tuberous sclerosis. Mol Cell Biol 26:6425–6434
Shaw RJ (2009) LKB1 and AMP-activated protein kinase control of mTOR signalling and growth. Acta Physiol 196:65–80
Testa JR, Tsichlis PN (2005) AKT signaling in normal and malignant cells. Oncogene 24:7391–7393
Thoreen CC, Chantranupong L, Keys HR, Wang T, Gray NS, Sabatini DM (2012) A unifying model for mTORC1-mediated regulation of mRNA translation. Nature 485:109–113
van Dam TJ, Zwartkruis FJ, Bos JL, Snel B (2011) Evolution of the TOR pathway. J Mol Evol 73:209–220
Vander Haar E, Lee SI, Bandhakavi S, Griffin TJ, Kim DH (2007) Insulin signalling to mTOR mediated by the Akt/PKB substrate PRAS40. Nat Cell Biol 9:316–323
Weber JD, Gutmann DH (2012) Deconvoluting mTOR biology. Cell Cycle 11:236–248
Wullschleger S, Loewith R, Oppliger W, Hall MN (2005) Molecular organization of target of rapamycin complex 2. J Biol Chem 280:30697–30704
Wullschleger S, Loewith R, Hall MN (2006) TOR signaling in growth and metabolism. Cell 124:471–484
Yang Q, Inoki K, Ikenoue T, Guan KL (2006) Identification of Sin1 as an essential TORC2 component required for complex formation and kinase activity. Genes Dev 20:2820–2832
Yip CK, Murata K, Walz T, Sabatini DM, Kang SA (2010) Structure of the human mTOR complex I and its implications for rapamycin inhibition. Mol Cell 38:768–774
Yu Y, Yoon SO, Poulogiannis G, Yang Q, Ma XM, Villen J, Kubica N, Hoffman GR, Cantley LC, Gygi SP et al (2011) Phosphoproteomic analysis identifies Grb10 as an mTORC1 substrate that negatively regulates insulin signaling. Science 332:1322–1326
Zeng H, Yang K, Cloer C, Neale G, Vogel P, Chi H (2013) mTORC1 couples immune signals and metabolic programming to establish T(reg)-cell function. Nature 499:485–490
Zinzalla V, Stracka D, Oppliger W, Hall MN (2011) Activation of mTORC2 by association with the ribosome. Cell 144:757–768
Zoncu R, Bar-Peled L, Efeyan A, Wang S, Sancak Y, Sabatini DM (2011a) mTORC1 senses lysosomal amino acids through an inside-out mechanism that requires the vacuolar H(+)-ATPase. Science 334:678–683
Zoncu R, Efeyan A, Sabatini DM (2011b) mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol 12:21–35
Acknowledgements
We thank Hiroyuki Inuzuka, Kai Xu, Alan Lau, Brian North, Lixin Wan, Xiangpeng Dai, Yueyong Liu, Kohei Ogura, Shavali Shaik and Jifang Zhang for critical reading of the manuscript and helpful discussions. This work was supported by grants from the National Institute of General Medicine, NIH (GM089763, GM094777, CA177910) to W.W. W.W. is an American Cancer Society Scholar and Leukemia and Lymphoma Society Research Scholar. P.L. is supported by 5T32HL007893.
Compliance With Ethics Guidelines
Pengda Liu, Jianping Guo, Wenjian Gan and Wenyi Wei declare that they have no conflict of interest.
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Liu, P., Guo, J., Gan, W. et al. Dual phosphorylation of Sin1 at T86 and T398 negatively regulates mTORC2 complex integrity and activity. Protein Cell 5, 171–177 (2014). https://doi.org/10.1007/s13238-014-0021-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13238-014-0021-8