
Abstract
Siderophore facilitates iron availability in soil, but its assistance in iron transportation to different plant parts is not reported till date. Therefore, it is worthwhile to study the effect of siderophore produced by Pseudomonas for iron acquisition in the presence and absence of iron. To study these effects, two siderophore-producing Pseudomonas aeruginosa strains RSP5 and RSP8 were selected. RSP5 and RSP8 produced the highest and lowest amounts of siderophore, respectively. Iron (Fe) concentration of stem, leaf, seed, and shoot length, root length, cob length, and number of grains parameters were analysed. It was observed that the plants treated with RSP5 were sturdier and taller than RSP5 + Fe > RSP8 > RSP8 + Fe > Fe > Control plants. Iron content of RSP8 vs. RSP8 + Fe, RSP8 + Fe vs. Control, and RSP8 + Fe vs. RSP5 + Fe was significantly different (P < 0.01). Analysis of variance (ANOVA) proves that RSP5 was able to transport higher amount of iron to maize plant than other treatments. Increase in shoot length, root length, cob length, grain number and iron content of stem, and leaf and seed of maize plant inoculated with RSP5 suggests that the strain can be used as an inoculant for increasing iron transportation in maize plant. (Indian Patent Filed: 40163/DEL/2016).
Similar content being viewed by others


Introduction
Seventeen essential plant food nutrients or minerals are required for optimal growth and development of crops. All are equally important to the plant, though required in different amounts. This difference has led to the grouping of these essential minerals into three categories; primary (macro) nutrients (nitrogen, phosphorus, potassium, carbon, hydrogen, and oxygen), secondary nutrients (calcium, magnesium, and sulphur) and micronutrients (boron, chlorine, cooper, iron, manganese, molybdenum, and zinc). Inadequate balance of these minerals with other nutrients may lead to growth suppression or even complete inhibition (Mengel et al. 2001). Micronutrients are involved in the key physiological processes of photosynthesis and respiration (Marschner 1995; Mengel et al. 2001) and their deficiency can impede these vital physiological processes thus limiting yield gain.
Iron (Fe) is an essential element for all living microorganisms and human beings, and for plant growth (Kobayashi and Nishizawa 2015), as it plays a major role in oxygen transport, oxidative metabolism, cellular proliferation, oxygen metabolism, electron transfer, DNA, and RNA synthesis, and acts as a catalyst in enzymatic processes (Aguado-Santacruz et al. 2012). Although iron is present in abundance, it is unavailable to plants due to its prevalence as insoluble iron oxyhydroxide polymers under aerobic conditions. Bacteria present in soil tend to produce siderophores under iron limiting conditions to enhance the bioavailability of Fe in the environment. The existence of siderophore-producing microorganisms in all the rhizosphere has led to the hypothesis that plants would become Fe deficient in the absence of siderophore as siderophore chelate the insoluble iron (Fe3+) in different pH condition and make it available for plants (Kloepper et al. 1980; Crowley et al. 1991).
Iron in human and animal occurs in two forms (dietary iron): heme and non-heme. The primary sources of heme iron are hemoglobin and myoglobin from consumption of meat, poultry, and fish, whereas non-heme iron is obtained from cereals, pulses, legumes, fruits, and vegetables (Abbaspoura et al. 2015). In agriculture, Maize occupies a prominent position and each part of the maize plant is put to one or the other use and nothing goes as waste. Maize as a crop has multiple uses but is chiefly grown for human and Livestock consumption. In most of the developing countries, maize is consumed directly as food but is iron deficient. Therefore, the purpose of the study was to utilize the siderophore-producing ability of Pseudomonas strains to transport iron to the seeds of maize plant from soil. The research could be beneficial to enhance iron content in the plant/seed to enhance the dietary heme in food and feed components.
Material and method
Isolation of siderophore-producing Pseudomonas
Presumptive Pseudomonas were selected from lab culture isolated from Pine and Oak forest and Agricultural land of Uttarakhand, India (Sah and Singh 2016).
Quantification of siderophore production with Fe
Quantification of siderophore was done for 14 isolates on sodium succinate medium (SSM). SSM supplemented with 20 µM Fe was inoculated with overnight grown culture of Pseudomonas and incubated for 24 h at 28 °C in incubator shaker. Sample was withdrawn after 24 h from SSM and centrifuged at 8000 rpm for 8 min. Absorbance of supernatant was measured at 400 nm (Meyer and Abdallah 1978; Sah and Singh 2016)
Plant growth experiment
On the basis of siderophore production, RSP5 and RSP8 were selected to observe the iron transport in maize plant. Seeds of maize (Zea mays L.) were obtained from G. B. Pant University of Agriculture and Technology, Pantnagar, India. These seeds were surface sterilized in 0.1% HgCl2, followed by thorough washing with autoclaved distilled water. Bacterization of seed (seed load; 105 CFU per seed) was performed using carboxy methyl cellulose (CMC) as matrix (Sharma and Johri 2003). This was followed by sowing seeds in pots (500 g soil volume/pot). Standard Hoagland’s solution was used to irrigate the pots once a week; autoclaved double distilled water was used during rest of growth period. Standard Hoagland’s solution supplemented with 20 µM Fe was used to irrigate the pots for observation to stop siderophore production. The observations for root/shoot length, number of leaves, length of corn, number of grains per corn, number of rows per corn, and iron content in leaf, stem, and seed, and were taken after 70 days of sowing. Different combinations were designed in triplicates as: (a) soil with Bacteria; (b) soil with Fe (20 µM); (c) Soil with Bacteria and Fe (20 µM); and (d) Control: Without bacteria and Fe (20 µM). The results were analysed biometrically using SPSS Statistics for Windows, Version 20.0. (Armonk, NY: IBM Corp.).
Atomic absorption spectroscopy (AAS) of the plant parts (stem, leaf, and seed)
Stem, leaf, and seeds were collected separately for all the sample plants and labeled. Equal weight of each sample was weighed and burnt to ash in hot oven. Aqua regia was prepared taking HCl and HNO3 in the ratio of 3:1.15 ml of aqua regia; 10 ml water and sample were mixed in separate beakers followed by boiling on hot plate. The mixture was boiled till the volume reduced to 8 ml followed by filtration. The filtrate was then properly diluted and iron content was measured by AAS (Analytical Iena vario-6 cold Vapour-HG-AAS) at NPL, Delhi. One-way repeated-measures analysis of variance (ANOVA) and post hoc analysis (Holm–Sidak method) were performed for further exploration of data through pairwise multiple comparisons through professional statistical software, i.e., Sigma Plot (Systat Software, San Jose, California, USA) and IBM SPSS Statistics for Windows, Version 20.0. (Armonk, NY: IBM Corp.).
Result and discussion
Isolation of siderophore-producing Pseudomonas
On the basis of molecular studies, the Pseudomonas strains isolated from Pine and Oak forest and Agricultural land of Uttarakhand, India were identified as a P. aeruginosa. The GenBank/NCBI accession number of the strain P. aeruginosa strains was as RSP5–KR051490 and RSP8–KR051488 (Sah et. al 2017).
Quantification of siderophore-producing Pseudomonas with and without Fe
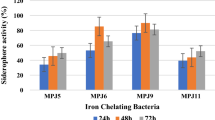
Siderophore production for 14 isolates having a zone size more than 2 cm was quantified with and without adding Fe (20 µM) to SSM. Supernatant of SSM medium after 24 h of inoculation with Pseudomonas was scanned spectrophotometrically and clear peak was observed in the range of 367–400 nm confirming production of siderophore (Meyer and Abdallah 1978). SSM medium without addition of iron was accompanied by siderophore production in the range of 75.00–210 µg mL−1, while after supplementing SSM medium with iron, siderophore production was repressed with a range between 10.00 and 160 µg mL−1. The highest siderophore producer was identified as P. aeruginosa, RSP5 (134, 210 µg mL−1 with and without iron, respectively) and the lowest siderophore producer was identified as P. aeruginosa RSP8 (10, 75 µg mL−1 with and without iron, respectively). Colquhoun and Sorum (2001) confirm that P. fluorescens growing in iron-limited conditions (less than 160 gL−1) should induce siderophore production. It was observed that siderophore production and iron concentration are inversely proportional. Threshold level of iron which repressed siderophore production was found to be 20 µM. It was reported earlier also that Siderophore production impedes at a concentration ≥20 µM (Nair et al. 2007; Bholay et al. 2012).
Plant growth experiment
Biometrical analysis
Maize seeds were sown and different treatments were given, according to the experiment design. After 70 days, the shoot length, root length, cob length, and number of seeds of maize plants were analysed (Table 1). It was observed that the plants treated with RSP5 were sturdier and taller than the other combinations (RSP5 + Fe > RSP8 > RSP8 + Fe > Fe > Control plants). The shoot length (SL) and root length (RL) of control plant were observed to be 103 and 7 cm, respectively. An increase of 51 and 44% was observed in the shoot length of maize plants inoculated with RSP5 and RSP5 + Fe, respectively. The shoot enhancement was recorded to be 19.35 and 9.99% by RSP8 and RSP8 + Fe respectively. A negligible increase of 3% was observed in shoot length of plants uninoculated by any bacteria. A comparable difference of 21% was observed in shoot length of plants inoculated with RSP5 and RSP8. The SL and RL of plants inoculated with RSP5 > RSP8 as RSP5 produce more siderophore than RSP8. Siderophore produced by RSP5 and RSP8 was lowered when 20 µM Fe was added, and hence the SL and RL of RSP5 + Fe < RSP5 and RSP8 + Fe < RSP8 treated plants.
Some species of Pseudomonas are considered as the plant growth promoting rhizobacteria (PGPR). Pseudomonas bacteria improve plant growth and yield through direct and indirect mechanisms (Crowley et al. 1991). Pseudomonas produce siderophore under Fe deficiency conditions which act like chelates and transport Fe into plant roots and help in growth of plants (Sharma and Johri 2003).
Corn data analysis
Figure 1 and Table 1 represent the variation in Cob length and grains per cob from soil inoculated with RSP5, RSP8, RSP5 + Fe and RSP8 + Fe. The cob length (CL): 21 cm, rows per cob (RC): 18 and grains per cob (GC): 395 were highest when plants were treated with RSP5, while the CL and GC of Control plants were 10 cm and 120. The CL and GC of RSP5 + Fe treated plants was 12 cm and 156, respectively; the values were less than RSP5 but more than Control. The CL and GC of plants inoculated with RSP8 (18 cm and 344 respectively) was more than the control plant but less than the plants treated with RSP5. The CL and GC of the various treatments was observed to be highest in RSP5 followed by RSP8 > RSP5 + Fe > RSP8 + Fe ≥ Fe > Control. The reason for the above results may be higher siderophore production by RSP5 than RSP8. The absence of RSP5 and RSP8 in Fe treated and Control plants leads to least CL and GC as no siderophore is produced in either of the treatments.

Effect of different bacterial treatments on cob length, number of rows/cob, and number of grains/row
Iron transportation with the help of siderophore
The AAS results revealed highest level of iron in stem, leaf, and seed (SLS) (3.59, 3.65, and 2.81 ppm, respectively) of plants treated with RSP5. The iron transportation in plants treated with Fe and RSP8 + Fe was almost equal. Iron content in SLS of plants uninoculated with bacteria and supplied with Fe (20 µM) was recorded to be 0.40, 0.75, and 0.24 ppm, respectively, and for the plants treated with RSP8 + Fe (1.17, 0.09, and 0.14 ppm, respectively). The iron content of SLS increased by 301.6, 328, and 487.5%, respectively, when treated with RSP5 and 71.87, 63.56, and 92.89% when plants treated with RSP8 (Fig. 2). The graph clearly depicts the relation between siderophore and iron in seed.

Iron transported to stem, leaf, and seed with the help of siderophore using different treatments
Iron transportation in plant through leaf to seeds represented in Fig. 3 and was better understood when analysed statistically.

Schematic representation of iron transportation to stem, leaf, and seed with the help of siderophore
One-way repeated-measures analysis of variance (ANOVA) exhibited highly significant differences in iron content among various treatments of stems, leaves, and seeds of maize plant (df = 5; SS = 19.216; MS = 3.843; F = 55.475; P < 0.001). A post hoc analysis (Holm–Sidak Method) was performed for further exploration of data through pairwise multiple comparison (Table 2) which showed a significant effect of various treatments on the transportation of iron on the plant through leaf to seed. Analysis of variance proves that RSP5 was able to transport higher amount of iron to maize plant than other treatments (RSP8, Fe, RSP5 + Fe, RSP8 + Fe, Control). A highly significant difference P < 0.001 was observed when iron content of stem, leaf, and seed of plants supplied with RSP5 was compared to Fe, RSP5 + Fe, RSP8, RSP8 + Fe, and Control with a mean difference of 2.746, 2.504, 2.161, 3.288, and 2.234, respectively (Table 2). The statistical difference observed in iron content of plants treated with RSP5 and RSP8 represents the difference in iron transported with both the treatments. Significant difference (P < 0.01) was observed when iron content of RSP8 vs. RSP8 + Fe, RSP8 + Fe vs. Control, and RSP8 + Fe vs. RSP5 + Fe were compared. Highly significant difference in iron level, P < 0.001, and a difference of mean 2.504 in RSP5 vs. RSP5 + Fe suggest that there is a comparable difference in iron content of both the treatments and excess of iron hindered production of siderophore as well as transport. A highly significant difference was observed when soil supplemented with RSP5 and Control was compared. The iron content in RSP8 and Control was not significantly different (P > 0.05) with a minor difference of mean (0.740), while iron content of RSP5 vs. RSP8 + Fe have a high difference of mean (3.288). The iron content in treatments Control vs. Fe, RSP5 + Fe, RSP8 and Fe vs. RSP5 + Fe, and RSP8 + Fe was not significantly different suggesting that the difference in their iron content was not comparable. The statistical analysis results also prove that the iron content in stem, leaf, and seeds of maize plant grown in uninoculated iron sufficient and iron-deficient soil were not significantly different (P > 0.05).
RSP5 produces higher amount of siderophore than RSP8; therefore, the iron transported with the help of siderophore is more in seeds coated with RSP5 than RSP8. Siderophore production reduced after inoculating the soil with Fe and it was interesting to note that iron transportation was also reduced when inoculated with RSP5 and RSP8 along with iron supplementation. The absence of Pseudomonas strains (RSP5 and RSP8) in Control plant resulted in low iron transportation and plant growth.
Soil microbes play an important role in favoring plant iron (Fe) uptake under Fe-limiting conditions (Jin et al. 2010); similarly, siderophore released by RSP5 was capable of solubilising the insoluble iron present in soil making it available to plant and transporting it to the plant (stem, leaf, and seed). Masalha et al. (2000) suggested that the microbial community present in rhizosphere plays an important role in Fe acquisition, affecting the composition of siderophore secreting microbes in the rhizosphere (Jin et al. 2010; Camejo et al. 2013). Under Fe-deficient conditions, it has been reported that the pyoverdines (siderophores secreted by Pseudomonas) show a high affinity for Fe(III) (Meyer and Abdallah 1978; Boukhalfa and Crumbliss 2002), and thus, when the Fe(III) content in soil was increased more than 20 µM, the siderophore production by (P. aeruginosa) RSP5 and RSP8 was negligible affecting the iron transportation to the maize plant (stem, leaf, and seed). Bacterial siderophore is not absorbed by the plant, but Fe released by siderophore through a reduction-based mechanism (Cesco et al. 2002; Hordt et al. 2000; Romheld and Marschner 1986) is absorbed by the plant.
Conclusion
Siderophore production by different Pseudomonas species has widely been studied and is found to be dependent on iron content present in soil. Our experimental findings reveal that Pseudomonas isolates RSP5 and RSP8 yield different amount of siderophore in iron deficient (≤20 µM) and iron sufficient (≥20 µM) soil. RSP5 produces higher amount of siderophore than RSP8 in iron sufficient as well as iron-deficient soil. The plant growth experiment suggests high amount of siderophore produced by RSP5 solubilised more iron from soil and transported iron more efficiently to various plant parts of Maize (dietary food) than RSP8. The increased iron content in stem, leaf, and seed of Maize will be beneficial for supplementation of dietary iron (non-heme) in human and animal food and feed.
References
Abbaspoura KC, Rouholahnejada E, Vaghefia S, Srinivasan BR, Yanga HC, Kloved BA (2015) Continental-scale hydrology and water quality model for Europe: calibration and uncertainty of a high-resolution large-scale SWAT model. J Hydrol 524:733–752
Aguado-Santacruz GAA, Moreno-Gómez BA, Jiménez-Francisco BB, García-Moya EB, Preciado-Ortiz RE (2012) Impact of the microbial siderophores and phytosiderophores on the iron assimilation by plants: a synthesis. Rev Fitotec Mex 35:9–21
Bholay AD, Priyanka JU, Borkhataria BV, Mayuri VD (2012) Fluorescent pseudomonads as plant growth promoting rhizobacteria and their siderophoregenesis. J Pharm Biol Sci 3:27–32
Boukhalfa H, Crumbliss AL (2002) Chemical aspects of siderophore mediated iron transport. Biometals 15(4):325–339
Camejo D, Romero-Puertas MDC, Rodriguez-Serrano M, Sandalio LM, Lazaro JJ, Jimenez A (2013) Salinity-induced changes in S-nitrosylation of pea mitochondrial proteins. J Proteom 79:87–99
Cesco S, Nikolic M, Romheld V, Varanini Z, Pinton R (2002) Uptake of Fe from soluble Fe-humate complexes by cucumber and barley plants. Plant Soil 241:121–128
Crowley DE, Wang YC, Reid CPP, Szansiszlo PJ (1991) Mechanism of iron acquisition from siderophores by microorganisms and plants. Plant Soil 130:179–198
Colquhoun DJ, Sorum H (2001) Temperature dependent siderophore production in Vibrio salmonicida. Microb Pathog 31:213–219
Hordt W, Romheld V, Winkelmann G (2000) Fusarinines and dimerum acid, mono and dihydroxamate siderophores from Penicillium chrysogenum, improve iron utilization by strategy I and strategy II plants. Biometals 13:37–46
Jin L, Ravella R, Ketchum B, Bierman PR, Heaney P, White TS, Brantley SL (2010) Mineral weathering and elemental transport during hill slope evolution at the susquehanna/shale hills critical zone observatory. Geochim Cosmochim Acta 74(13):3669–3691
Kloepper JW, Leong J, Teintze M, Schroth NM (1980) Enhanced plant growth by siderophores produced by plant growth promoting rhizobacteria. Nature 286:885–886
Kobayashi T, Nishizawa NK (2015) Intracellular iron sensing by the direct binding of iron to regulators. Front Plant Sci 6:155
Marschner H (1995) Mineral nutrition of higher plants, 2nd edn. Academic Press, London
Masalha J, KosegartenH ElmaciO, Mengal K (2000) The central role of microbial activity for iron acquisition in maize and sunflower. Biol Fertil Soils 30:433–439
Mengel K, Kirkby EA, Kosegarten H, Appel T (2001) Principles of plant nutrition. Kluwer Academic, Dordrecht
Meyer JM, Abdallah MA (1978) The fluorescent pigment of P. fluorescens: biosynthesis, purification and physiochemical properties. J Gen Microbiol 107:319–328
Nair A, Juwarkar AA, Singh SK (2007) Production and characterization of siderophores and its application in arsenic removal from contaminated soil. Water Air Soil Pollut 1:199–212
Romheld V, Marschner H (1986) Evidence for a specific uptake system for iron phytosiderophores in roots of grasses. Plant Physiol 80:175–180
Sah S, Singh R (2016) Effect of Pseudomonas on micronutrient status of forest and agricultural soil of Uttarakhand. Ecol Env Conserv 22:S279–S284
Sah S, Sahgal M, Singh R (2017) Concomitant ability of siderophore against iron paucity and fusarium wilt in lycopersicon esculentum. Biosci Biotech Res Asia 14:319–328
Sharma A, Johri BN (2003) Growth promoting influence of siderophore-producing Pseudomonas strains GRP3A and PRS9 in maize (Zea mays L.) under iron limiting conditions. Microbiol Res 158:243–248
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sah, S., Singh, N. & Singh, R. Iron acquisition in maize (Zea mays L.) using Pseudomonas siderophore. 3 Biotech 7, 121 (2017). https://doi.org/10.1007/s13205-017-0772-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-017-0772-z