
Abstract
The activities of the antioxidant enzymes superoxide dismutase (SOD), catalase (CAT) and glutathione S-transferase (GST), the incidence of DNA damage, the activation of poly (ADP-ribose) polymerase-1 (PARP-1), a marker of DNA repair, and connective tissue growth factor (CTGF), a marker of tissue fibrosis, were examined in the hearts of rats for 16 weeks after diabetes induction by streptozotocin (STZ) administration. A 150% increase in CAT activity was detected at the end of the 2nd week post-STZ administration, and CAT activity remained 80% above the control level throughout 16 weeks. While total SOD and CuZn-SOD exhibited progressively decreasing activities, those of Mn-SOD and GST were elevated. Neither DNA strand breaks (apoptosis or necrosis) nor changes in PARP-1 activity and in CTGF levels (fibrosis) were observed in the diabetic heart. The absence of cardiomyopathy is accompanied with increased activities of CAT, MnSOD and GST.
Similar content being viewed by others

Introduction
The hyperglycemia that accompanies diabetes mellitus is responsible for inducing oxidative stress that leads to modifications of intracellular structures, proteins and DNA damage, with serious consequences for proper cell functioning [1, 2]. The overproduction of different reactive oxygen species such as O2, OH· and H2O2, which are collectively referred to as ROS, is the initiating event leading to long-term development of diabetic complications, expressed primarily in the cardiovascular system and kidneys [3, 4]. Diabetes alters free radical metabolism in blood and tissues; however, these alterations are diverse, and a clear picture of how they are involved in organ-specific complications has yet to emerge [5, 6].
ROS are produced in cells under physiological conditions. To prevent damage by ROS, cells have developed an antioxidant defense system [4]. The central function of antioxidant enzymes is to maintain the concentrations of ROS in the cell at a stationary level. The resistance of the organism and its individual organs, as well as their capability to adapt to the effects of the environment, are largely determined by the activity of superoxide dismutase (SOD) enzymes found in the cytosol (CuZn-SOD, Sod 1) and mitochondria (Mn-SOD, Sod 2), and catalase (CAT) [7, 8]. Although CAT and SOD protect against ROS, they are ineffective against the cumulative toxicity of lipid peroxidation products [9]. Enzymes involved in detoxification of lipid peroxidation products are important in the hierarchy of antioxidant defense mechanisms as well as glutathione S-transferases (GST) [9]. These enzymes play an important role in the decomposition of hydrogen peroxide and/or other organic hydroperoxides and are abundantly expressed in liver tissue [10]. Information concerning the presence of GST in the heart is contradictory.
When ROS production exceeds the cell’s capacity to neutralize them, the ensuing oxidative stress leads to tissue damage. Diabetic cardiomyopathy is the result of physiological adaptation to metabolic alterations and degenerative changes of the myocardium, which has a limited capacity for repair. In general, many factors can affect the process of development of diabetic cardiomyopathy, and all diabetic persons are not affected by the same factors to the same degree. As a result, marked variability in the manifestations of diabetic cardiomyopathy are observed. Diabetic cardiomyopathy initiated by hyperglycemia at an early stage is characterized by metabolic disturbances (increased free fatty acids, insulin resistance), without changes in myocardial structure. An increase in myocyte apoptosis and necrosis is observed in the middle stage of cardiomyopathy, with resulting myocardial fibrosis and hypertrophy [11].
Oxidative stress has long been implicated in the progression of diabetes complications [12]. Angiotensin II stimulates ROS generation via NADPH oxidase activity, whereas antioxidants provide renal protection from oxidative stress [13, 14]. While such data strongly point to a link between ROS, activation of the renin-angiotensin system (RAS) and renal cell apoptosis in diabetes, no data suggest that there is a connection between the development of cardiomyopathy and antioxidant levels.
The aim of our study was to correlate the presence of antioxidative enzymes to the level of cardiomyocyte damage in rat heart during long-term streptozotocin (STZ)-induced diabetes. Since the indicators for cardiomyopathy development are necrosis and/or apoptosis and subsequent fibrosis and hypertrophy, we examined whether the diabetic state was associated with an increase in DNA damage and resulting activation of poly (ADP-ribose) polymerase-1 (PARP-1). PARP-1 plays an important role in DNA repair and as a result of its overactivation it leads the cell to its demise. As every tissue possesses specific combinations of different antioxidant enzymes and different capacities for their activity, we examined SOD, CAT and GST activities during long-term STZ-induced diabetes in rats.
Methods
Animals
All animal procedures were approved by the Committee for the Ethical Care and Use of Laboratory Animals of the Institute for Biological Research, Belgrade, and were in accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication no. 85/23, revised 1986). Ten-week-old male Wistar rats were maintained at 22 ± 1°C, 50 ± 5% humidity at 12-h light/day intervals and were provided with rat chow and drinking water ad libitum. In a single experimental cycle, the following groups of animals were used: (1) a control group consisting of 10 rats that received saline (ml/kg); (2) a group of 40 rats that was injected with streptozotocin (STZ). The rats were anesthetized by an intraperitoneal injection of sodium pentobarbital (50 mg/kg) and killed at the indicated times. Diabetes was induced by a single intraperitoneal injection of STZ at a dose of 60 mg/kg. Administration of insulin during the early stages of diabetes can potentially delay or impede the progression of more permanent tissue damage. For that reason experimental rats were not treated with insulin. Diabetes was confirmed by glucosuria that was measured using glucose test strips (BM-Test-5L, Boehringer Mannheim, Germany). After administration of a single dose of STZ, the blood glucose concentration was raised 400–500% above the normal value and remained elevated throughout the follow-up period of 16 weeks. Blood glucose levels were also measured using a commercial kit (Gluco-quant Glucose/HK, Boehringer Mannheim, Germany) based on the hexokinase/G6P-DH enzymatic method. Rats with sustained diabetes (glucosuria of at least 20 mmol/l 2 weeks after injection of STZ) (n = 34) were randomly divided and killed 2, 4, 8 and 16 weeks after STZ administration.
Isolation of heart nuclei
The procedure for the isolation of heart nuclei was carried out as described by Liew et al. [15]. The nuclei were stored in sterile glycerol (v/v = 1/1) at −20°C. For the PARP-1 activity assay the nuclei were lysed in lysis buffer [50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% Triton X-100, 0.2 M PMSF, aprotinin, leupeptin and pepstatin (each at 1 μg/ml)]. For Western blot analysis for the presence of CTGF, crude nuclear extracts were used. Hearts that were deep-frozen were ground to a powder using a mortar and pestle and suspended in extraction buffer (6 M urea, 10% glycerol, 2% SDS, 2% bovine serum albumin, β-mercaptoethanol, protease inhibitor coctail; Protease Inhibitor Mix G, Serva). The suspension was sonicated for 15 s (Bandelin Sonopuls MS73 sonicator). The entire procedure was performed at 0–4°C.
Western immunoblot analysis
Western immunoblot blot analysis was performed according to the procedure recommended by the supplier of the ECL Western analysis kit (Amersham Pharmacia Biotech). Twenty micrograms of proteins were subjected to 9 or 12% sodium dodecyl sulfate-polyacrylamide-gel electrophoresis (SDS-PAGE) and transferred to PVDF membranes (Hybond-P, Amersham Pharmacia Biotech). Western immunoblot analysis was performed using polyclonal antibodies for PARP-1 and CTGF (Santa Cruz Biochemical Inc., Santa Cruz, CA).
SOD, CAT and GST activity in tissue
Total heart was homogenized in 0.25 M sucrose, 0.1 M EDTA and 0.05 M Tris-HCl, pH 7.4. After sonication, the homogenate was centrifuged for 90 min at 100,000×g. Aliquots of the supernatant were stored at −80°C. Total SOD (totSOD) activity was measured by the epinephrine method [16], which is based on the capacity of SOD to inhibit the auto-oxidation of epinephrine to adrenochrome, and was expressed as U/g wet mass. Mn-SOD activity was performed after preincubation with 8 mmol/l KCN, while CuZnSOD activity was calculated from the difference between total SOD and MnSOD activities. CAT activity was measured by the rate of hydrogen peroxide decomposition and expressed as μmol H2O2/min/g wet mass [17]. GST activity was determined by the method of Habig et al. [18] based on the reaction of 1-chloro-2,4-dinitrobenzene (CDNB) with –SH groups of glutathione catalyzed by GST contained in the samples (nmol GSH/min/g wet mass).
DNA extraction and separation
Heart tissue homogenized in buffer (20 mM EDTA, 50 mM Tris-HCl, pH 8.0, 0.5% SDS and 100 μg/ml proteinase K) was incubated for 3 h at 50°C and treated with RNaseA (final concentration 150 μg/ml) for 1 h at 37°C. DNA was phenol/chloroform extracted and resuspended in TE (Tris-EDTA) buffer. DNA (1 μg) was separated in a 1.5% agarose gel in 0.5 × TBE (Tris-Borate-EDTA) buffer at 90 V.
Poly (ADP-ribose) polymerase-1 (PARP-1) activity assay
PARP-1 activity was measured in heart nuclear lysates using a commercial colorimetric PARP-1 assay kit (Trevigen) according to the manufacturer’s instructions.
Immunohistochemistry
Tissue sections were incubated with polyclonal PARP-1 and CTGF antibodies (Santa Cruz Biochemical Inc., Santa Cruz, CA) at 4°C overnight at 1:100 dilution. This was followed by incubation with biotinylated horse anti-mouse IgG (Vector Laboratories, Burlingame, CA) for 30 min and thereafter with the avidin-biotin-peroxidase complex using a Vectastain ABC kit (Vector Laboratories). The reaction was visualized using a 3,3′-diaminobenzidine tetrahydrochloride (DAB) solution. After counterstaining with hematoxylin, the slides were dehydrated and mounted in Permount. Photomicrographs were taken with a Zeiss Axiolab microscope equipped with a Fuji HC-300C digital camera. Controls were incubated with PBS in place of the primary antibody, and no positive staining was observed.
TBARS assay
Plasma concentrations of thiobarbituric acid reactive substances (TBARS) are an index of lipid peroxidation and oxidative stress. The basic principle of the TBARS assay is based on the reaction of malonalaldehyde bis (MDA) with thiobarbituric acid (TBA) at 95°C. Briefly, an aliquot of the homogenate (0.1 ml) was mixed with 0.2 ml of 8.1% SDS, 1.5 ml 20% acetic acid (pH 3.5), 1.5 ml 0.8% TBA and 0.7 ml water and heated at 95°C for 60 min. After cooling to room temperature, 1 ml of water and 5 ml of n-butanol/pyridine (15:1, v/v) were added, mixed and centrifuged at 3,000×g for 10 min. The red pigment in the supernatant fractions was estimated by absorbance at 532 nm. A calibration curve was prepared with an MDA standard (MDA concentrations ranged from 25 nmol/ml to 1 μmol/ml) [19].
Statistical analysis
The results were analyzed by one-way ANOVA followed by the Mann-Whitney U-test (post hoc comparison was performed between intact control and treated groups). The results were considered to be statistically significant at p < 0.05.
Results
Essential biological and biochemical parameters of rats relevant to diabetes are presented in Table 1. STZ administration led to a significant 4.5–5-fold increase in blood glucose concentration in rats and to a decrease in body weight (by about 20% in 2 weeks). The increase in blood glucose correlated with a significant decrease (3–1.8-fold) in plasma insulin levels during the analyzed period. The heart mass to body mass ratio serves as an estimate of hypertrophy of the heart that accompanies cardiomyopathy and was measured after induction of diabetes. No changes in the heart mass to body mass ratio were observed in diabetic rats during the analyzed period (Table 1).
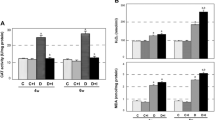
Changes in the level of lipid peroxides that serve as an indicator of the presence of oxidative stress were examined by the TBARS assay in diabetic hearts. The level of lipid peroxides changed significantly (40–60%) in the heart during 2–16 weeks after STZ injection (Fig. 1), showing the presence of oxidative stress. It is generally accepted that long-term diabetes exposes an organism to a state of chronic systemic oxidative stress that causes tissue damage and subsequent multiple organ dysfunction. Comparison of the basal antioxidative enzyme status between the heart and the liver, the central detoxification organ characterized by relatively high endogenous antioxidant levels revealed lower activities of the examined enzymes in the heart (CAT, SOD and GST were 60-, 2.5- and 10-fold lower, respectively, in the heart than in the liver; results not shown). The lower levels of SOD and CAT in the control heart suggest that this organ could be more susceptible to oxidative damage by the action of ROS. To examine the status of antioxidative defenses in the heart during long-term illness, we examined antioxidant enzyme activities 2, 4, 8 and 16 weeks after STZ administration. At all of the examined times, total SOD and CuZn-SOD activities in the diabetic heart were lower than in the controls. Two weeks after induction of diabetes (Fig. 2a), total SOD activity was 25.7% lower. A similar trend was observed for CuZn-SOD. At the end of the 2nd week after diabetes induction, 40.1% lower values compared to control activity were measured (Fig. 2a). At the same time, a slight increase of Mn-SOD activity was observed, with a maximal 33.7% increase observed at the end of the 4th week. Sixteen weeks after induction of diabetes, the levels of totSOD and CuZn-SOD remained lower compared to the control (22 and 42%, respectively), while at the same time the level of Mn-SOD activity was increased by 24.5% relative to the control (Fig. 2a).

Thiobarbituric acid-reactive substance (TBARS) levels in the heart. TBARS, an indicator of oxidative stress at the end of the 2nd, 4th, 8th and 16th weeks after diabetes induction compared to control level. C intact control; 2 W, 4 W, 8 W and 16 W 1st, 2nd, 4th, 8th and 16th weeks after STZ treatment. Values represent means ± SD from nine separate experiments. *p < 0.5 statistically significant difference versus intact control

Antioxidant enzymes in the hearts of control and diabetic rats. a SOD activity, b CAT activity, c GST activity at the end of the 2nd, 4th, 8th and 16th weeks after diabetes induction compared to respective control levels. Values represent means ± SEM from five separate experiments. The minimum level of significance was *p < 0.05
Unlike these antioxidant enzymes, CAT activity was significantly increased in the diabetic heart at all of the examined times (Fig. 2b). At the end of the 2nd week after induction of diabetes, CAT activity exhibited a maximal increase of 150.7% relative to the control. CAT activity persisted at the elevated level up to the end of the follow-up period. At 16 weeks after induction of diabetes, the activity of CAT was 82.8% higher than in controls. These results suggest that CAT plays an important role in protection against oxidative stress-related damage in the heart. Increased GST activity was detected at all analyzed times, except at the end of the 8th week. By the end of the 16th week after induction of diabetes, GST activity was raised by 34.4% above control enzyme activity (Fig. 2c).
To assess whether long-term diabetes leads to the development of cardiomyopathy as a result of cell death after DNA damage, we examined the integrity of cardiomyocyte DNA by agarose gel electrophoresis and by measuring PARP-1 activity. As can be seen from Fig. 3, double-strand DNA breaks were not detected at any of the examined time points. No relevant changes in PARP-1 activity in cardiomyocyte nuclear lysates prepared from hearts at 2, 4, 8 and 16 weeks after the induction of diabetes were observed (Fig. 4). In correlation with these results, Western blot analysis of nuclear lysates prepared from heart cells (Fig. 5c) revealed the presence of only native full length PARP-1 in both control and diabetic hearts. Immunohistochemical analysis showed consistent presence of PARP-1 in control and diabetic (16th week) hearts, despite severe architectural alterations of the diabetic myocardium (Fig. 5a, b).

DNA from the hearts of control and diabetic rats. DNA was stained with ethidium bromide and loaded on a 1.5% agarose gel. M standard DNA ladder; C DNA from control hearts

PARP-1 activity in heart nuclei. Nuclei isolated from hearts were lysed and analyzed for PARP-1 activity. C control heart; 2 W, 4 W and 16 W 2, 4 and 16 weeks after induction of diabetes, respectively

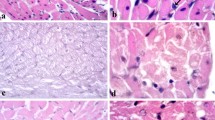
PARP-1 in the heart of control and diabetic rats. Immunohistochemical staining for PARP-1 in whole heart sections (indirect immunoperoxidase technique; hematoxylin diaminobenzidine counterstaining); a control and b 16 weeks after diabetes induction (a, b ×20). c Western blot analysis of heart nuclei with anti-PARP-1 antibody. C control heart; 2 W, 4 W and 16 W 2, 4 and 16 weeks after induction of diabetes, respectively. Lamin B served as an internal standard
The middle stage of diabetic cardiomyopathy is characterized by hypertrophy and myocardial fibrosis. To determine whether rat hearts after long-term diabetes displayed fibrous changes, we followed the expression of connective tissue growth factor (CTGF) in the heart after induction of diabetes. The expression of CTGF in the heart is normally repressed during postnatal life, but is dramatically induced during heart failure. Except for minor architectural alterations of the myocardium, immunohistochemical staining for CTGF in heart sections of control and diabetic rats (Fig. 6a, b) suggests the absence of fibrosis at the end of the 16th week after diabetes induction. This result was confirmed by Western blot analysis with anti-CTGF antibody of cardiomyocyte proteins prepared from control and diabetic hearts (Fig. 6c) that did not reveal an increase of CTGF.

Detection of CTGF in the heart of control and diabetic rats. Immunohistochemical staining for CTGF in whole heart sections (indirect immunoperoxidase technique; hematoxylin diaminobenzidine counterstaining), a control, b 16 weeks after diabetes induction (a, b ×20). c Western blot analysis of whole heart tissue homogenates with anti-CTGF antibody; C control heart; 2 W, 4 W and 16 W 2, 4 and 16 weeks after induction of diabetes. The lysate of NIH 3T3 mouse fibroblasts (lysF) served as a positive control. Actin served as an internal standard
Discussion
Streptozotocin-induced diabetes and hyperglycemia resulting from insulinopenia increase lipid peroxidation in the heart, which may provoke cardiomyopathy. Diabetes-associated cardiac dysfunction is associated with mitochondrial dysfunction and oxidative stress. Impaired insulin signaling predisposes cardiac mitochondria to oxidative stress, which not only damages mitochondria but can also impair cellular energetics through mitochondrial uncoupling. Insulin signaling plays an essential role in the protection of mitochondrial homeostasis in the heart [20]. Since the development of cardiomyopathy is a slow process, we analyzed a long period after diabetes induction. Throughout the examined 16-week period, despite persistent hyperglycemia we did not observe DNA damage or increased PARP-1 activity. The presence of CTGF in the heart was unchanged, suggesting the absence of fibrosis and hypertrophy of the heart. At the same time, elevated levels of CAT, GST and Mn-SOD in the heart were observed. Although long-term elevated glucose levels have been reported to induce damage and abnormalities in the hearts of experimental animals, we were unable to detect any damage.
Myocyte cell death results from apoptosis and/or necrosis [12]. Apoptosis does not cause scar formation or significant interstitial collagen accumulation. Conversely, myocyte necrosis causes a widening of the extra-cellular compartments between myocytes and increased deposition of collagen in a diffuse or scattered manner resulting from both replacement fibrosis because of myocyte necrosis and connective tissue cell proliferation [12]. Myocytes are highly differentiated cells with a very limited ability to proliferate and with a very low level of baseline apoptosis [21]. PARP-1 (113 kDa) is cleaved to 89- and 24-kDa fragments during apoptosis and is processed during necrosis to a 50-kDa fragment [21]. Neither apoptotic nor necrotic cleavage products of PARP-1 were observed. Western blot analyses of heart nuclear lysates showed only the presence of native full-length PARP-1 in both control and diabetic hearts, indicating that cardiac myocytes were not undergoing apoptosis or necrosis. These results are in correlation with the observed absence of DNA degradation at all analyzed time points.
Immunohistochemical staining of PARP-1 in whole heart sections from control and diabetic rats did not expose significant differences in PARP-1 staining. At the same time, we detected slightly decreased PARP-1 activity at 16-week post-diabetes induction, which correlates with the undetected development of cardiomyopathy [22, 23]. Increased activity of PARP-1 is usually related to the development of cardiomyopathy [24, 25]. Overactivation of PARP-1 in response to ROS-induced DNA breaks leads to cell dysfunction and a necrotic-type of cell death.
Data indicate that hyperglycemia directly induces myocyte necrosis in the myocardium, which is triggered by reactive oxygen species derived from high levels of glucose [12]. High glucose induces ROS generation, which also contributes to apoptosis in both primary neonatal and adult cardiomyocytes from the Nrf2 wild-type mouse heart. In Nrf2 knockout cells ROS was significantly higher, suggesting that Nrf2 is a critical regulator of defense against ROS in normal and diabetic hearts [12]. Up-regulation of local RAS in diabetes may enhance oxidative damage, activating cardiac cell apoptosis and necrosis [13]. Data have directly linked the overexpression of CAT with prevention of apoptosis [26]. Thus, there is evidence that ROS generation in RPTCs of db/db mice actually induces tubular apoptosis and that CAT overexpression can prevent it. We demonstrated that although hyperglycemia persisted at all examined times and oxidative stress and elevated lipid peroxidation were present, the heart of diabetic rats was protected. At the same time, elevated levels of CAT, GST and Mn-SOD in the heart were observed. We speculated that CAT overexpression prevented damage to the DNA. Our data can be linked with the data on RPTCs transgenic db/db mice with experimentally induced CAT overexpression [26] that showed that CAT has a potential for protecting and arresting the development of apoptosis. The absence of fibrosis at all examined periods can also be connected to increased CAT activity, as it was shown that CAT and metallothionein protect sarcomere functioning in diabetic myocytes [27].
We found that long-term hyperglycemia caused a reduction of totSOD and CuZn-SOD enzymatic activities in the heart while Mn-SOD was slightly increased. The decreasing trend of total SOD activity that was observed in the heart with progression of diabetes could be due to glycation of CuZnSOD, which caused its inactivation [28]. The Cu2+ that is released from the glycated CuZnSOD facilitates a Fenton reaction. Thus, the hydroxy radical produced by a Fenton reaction between Cu2+ and H2O2 released from the glycated CuZnSOD can participate in nuclear DNA cleavage, thereby contributing to the complications that accompany diabetes [29]. We assume that the H2O2 formed in this way and the H2O2 that was formed by oxidative stress (detected by oxidative peroxidation) were removed by CAT, whose activity was significantly elevated in diabetes.
The increase in Mn-SOD activity correlated with the progression of diabetes. Overexpression of Mn-SOD was shown to alter mitochondrial morphology and protect the morphology of the diabetic heart and completely normalize contractility in diabetic cardiomyocytes [30]. Mn-SOD overexpression also elevates myocyte CAT activity and GSH levels, and these changes can act together against oxidative insults [30].
Clearly, CAT, GST and Mn-SOD have a protective potential against oxidative stress in the rat heart. The GST level was increased after diabetes induction. Overexpression of GST in culture attenuates lipid peroxidation under normal conditions as well as during oxidative stress [31]. We detected increased peroxidation in the heart of diabetic rats that could be the reason for increased amounts of GST. It could protect against DNA damage at two levels: by preventing the formation of DNA damage as a result of the removal of lipid hydroperoxides and by directly repairing damaged DNA [32]. We must point out that increases in GST at 16 weeks could also represent an early marker for heart damage that was undetectable due to the high level of CAT. On the other hand, the absence of PARP-1 apoptotic and necrotic fragments may be connected to elevated GST activity since its overexpression was shown to protect from apoptosis in culture [11]. In conclusion, we suggest that increased antioxidant enzyme activities, especially of CAT, GST and Mn-SOD, play a role in heart protection and assist it in handling oxidative stress during diabetes in rats.
References
King GL, Loeken MR (2004) Hyperglycemia-induced oxidative stress in diabetic complications. Histochem Cell Biol 122:333–338
Wohaieb SA, Godin DV (1987) Alterations in free radicals tissue-defense mechanisms in streptozotocin-induced diabetes in rat: effect of insulin treatment. Diabetes 36:1014–1018
Baynes JW (1991) Role of oxidative stress in development of complications in diabetes. Diabetes 40:405–412
Boudina S, Abel ED (2007) Diabetic cardiomyopathy revisited. Circulation 115:3213–3223
Gupta BL, Baquer NZ (1998) Hexokinase, glucose-6-phosphate dehydrogenase and antioxidant enzymes in diabetic reticulocytes. Effects of insulin and vanadate. Biochem Mol Biol Int 46:1145–1152
Kakar R, Kalra J, Mantha SV, Prasad K (1995) Lipid peroxidation and activity of antioxidant enzymes in diabetic rats. Mol Cell Biochem 151:113–119
Perez-Campo R, López-Torez M, Rajas C, Cadenas S, Barja GA (1993) comparative study of free radicals in vertebrates. I. Antioxidant enzymes. Comp Biochem Physiol 105B:749–755
Maritim AC, Sanders RA, Watkins JB (2003) Diabetes, oxidative stress, and antioxidants a review. J Biochem Mol Toxicol 17:24–38
Hayes JD, Flanagan JU, Jowsey IR (2005) Glutathione transferases. Annu Rev Pharmacol Toxicol 45:51–88
Yang Y, Sharma R, Zimniak P, Awasthi YC (2002) Role of alpha class glutathione S-transferases as antioxidant enzymes in rodent tissues. Toxicol Appl Pharmacol 182:105–115
Fang ZY, Prins JB, Marwick T (2004) Diabetic cardiomyopathy: evidence, mechanisms, and therapeutic implications. Endocr Rev 25:543–567
He X, Kan H, Ca L, Ma Q (2009) Nrf2 is critical in defense against high glucose-induced oxidative damage in cardiomyocytes. J Mol Cell Cardiol 46:47–58
Lim HS, Robert J, MacFadyen F, Gregory Y, Lip H (2004) Diabetes mellitus, the renin–angiotensin–aldosterone system, and the heart. Arch Intern Med 164:1737–1748
Singh VP, Le B, Khode R, Baker KM, Kumar R (2008) Intracellular angiotensin II production in diabetic rats is correlated with cardiomyocyte apoptosis, oxidative stress, and cardiac fibrosis. Diabetes 57:3297–3306
Liew CC, Liu DK, Gornall AG (1972) Effects of aldosterone on RNA polymerase in rat heart and kidney nuclei. Endocrinology 90:488–495
Misra HP, Fridovich I (1972) The role of superoxide anion in the autooxidation of epinephrine and simple assay for superoxide dismutase. J Biol Chem 247:3170–3175
Beutler E (1982) Catalase. In: Beutler E (ed) Red cell metabolism, a manual of biochemical methods. Grune and Stratton Inc., New York
Habig WH, Pubst MJ, Jakoby WB (1974) Glutathione S-transferase. J Biol Chem 249:7130–7139
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95:351–358
Boudina S, Bugger H, Sena S, O’Neill BT, Zaha VG, Ilkun O, Wright JJ, Mazumder PK, Palfreyman E, Tidwell TJ, Theobald H, Khalimonchuk O, Wayment B, Sheng X, Rodnick KJ, Centini R, Chen D, Litwin SE, Weimer BE, Abel D, Phil D (2009) Contribution of impaired myocardial insulin signaling to mitochondrial dysfunction and oxidative stress in the heart. Circulation 119:1272–1283
Gobeil S, Boucher CC, Nadeau D, Poirier GG (2001) Characterization of the necrotic cleavage of poly(ADP-ribose) polymerase (PARP-1): implication of lysosomal proteases. Cell Death Differ 8:588–594
Pieper AA, Verma A, Zhang J, Snyder SH (1999) Poly (ADP-ribose) polymerase, nitric oxide and cell death. Trends Pharmacol Sci 20:171–181
Pieper AA, Brat DJ, Krug DK, Watkins CC, Gupta A, Blackshaw S, Verma A, Wang ZQ, Snyder SH (1999) Poly(ADP-ribose) polymerase-deficient mice are protected from streptozotocin-induced diabetes. Proc Natl Acad Sci U S A 96:3059–3064
Chiu J, Farhangkhoee H, Xu BY, Chen S, George B, Chakrabarti S (2008) PARP mediates structural alterations in diabetic cardiomyopathy. J Mol Cell Cardiol 45:385–393
Pacher P, Liaudet L, Soriano FG, Mabley JG, Szabó E, Szabó C (2002) The role of poly (ADP-ribose) polymerase activation in the development of myocardial and endothelial dysfunction in diabetes. Diabetes 51:514–521
Brezniceanu M-L, Liu F, Wei C-C, Che′nier I, Godin N, Zhang S-L, Filep JG, Ingelfinger JR, Chan JSD (2008) Attenuation of interstitial fibrosis and tubular apoptosis in db/db transgenic mice overexpressing catalase in renal proximal tubular cells. Diabetes 57:451–459
Ye G, Metreveli NS, Donthi RV, Xia S, Xu M, Carlson EC, Epstein PN (2004) Catalase protects cardiomyocyte function in models of type 1 and type 2 diabetes. Diabetes 53:1336–1343
Hunt J, Smith C, Wolff S (1990) Autoxidative glycosylation and possible involvement of peroxides and free radicals in LDL modification by glucose. Diabetes 39:1420–1424
Kaneto H, Fujii J, Suzuki K, Kasai H, Kawamori R, Kamada T, Taniguchi N (1994) DNA cleavage induced by glycation of Cu, Zn-superoxide dismutase. Biochem J 304:219–225
Shen X, Zheng S, Metreveli NS, Epstein PN (2006) Protection of cardiac mitochondria by overexpression of MnSOD reduces diabetic cardiomyopathy. Diabetes 55:798–805
Singhal SS, Godley BF, Chandra A, Pandya U, Jin GF, Saini MK, Awasthi S, Awasthi YC (1999) Induction of glutathione S-transferase hGST 5.8 is an early response to oxidative stress in RPE cells. Invest Ophthalmol Vis Sci 40:2652–2659
Bao Y, Jemth P, Mannervik B, Williamson G (1997) Reduction of thymine hydroperoxide by phospholipids hydroperoxide glutathione peroxidase and glutathione transferase. FEBS Lett 410:210–212
Acknowledgments
This work was supported by the Ministry of Science and Environmental Protection, Republic of Serbia, grant 143002B.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Ivanović-Matić, S., Mihailović, M., Dinić, S. et al. The absence of cardiomyopathy is accompanied by increased activities of CAT, MnSOD and GST in long-term diabetes in rats. J Physiol Sci 60, 259–266 (2010). https://doi.org/10.1007/s12576-010-0093-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12576-010-0093-z