
Abstract
The rich early Miocene small mammal assemblage from Montalvos2, collected from lacustrine deposits directly overlying the basement, is unique within the Teruel Basin, a basin that is otherwise well known for its late Miocene/Pliocene mammal faunas. The presence of Democricetodon decipiens, Megacricetodon primitivus, Eumyarion and Ligerimys ellipticus enables correlation with the local biozone Ca (approx. 16.3 Ma, MN4). The high percentage of ochotonids in Montalvos2 is remarkable, a phenomenon that is known to occur in older assemblages from the adjacent Calatayud-Montalbán Basin. The association in Montalvos2 includes species indicative of an arid biotope, such as the ochotonid Lagopsis penai and the rodents Simplomys simplicidens, Armantomys aragonensis, Melissiodon dominans and the xerine squirrel Heteroxerus, and species that are supposed to have lived in a more humid forested biotope, such as the Talpidae and the flying squirrel Albanensia. This wide range of species indicates that the environment in the Teruel Basin was highly diverse during the early Miocene.
Similar content being viewed by others


Introduction
Spain is well known for its continental sedimentary basins with rich fossil faunas of vertebrates, particularly smaller mammals (rodents, insectivores and lagomorphs). One of these basins is the Teruel Basin, from which numerous mammal assemblages, mostly of late Miocene age, have been recovered (Van de Weerd 1976; Van Dam et al. 2001). The small mammal assemblage of Montalvos2 represents the only known find in the Teruel Basin of a late early Miocene age, and it is therefore of great interest.
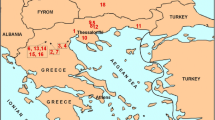
The locality Montalvos2 is situated in the northern part of the Teruel Basin, 3 km ESE of the village of Cuevas Labradas (Fig. 1) and has yielded a rich small mammal assemblage, which we here summarily describe and place within a stratigraphical and palaeoecological framework. It contains 775 dental elements of small mammals: rodents, lagomorphs, insectivores and marsupials. The assemblage has already been given an estimated age of approximately 16.4 Ma by Van der Meulen et al. (2012), based on the interpolation of two comparable assemblages from the Aragonian type area in the nearby Calatayud-Montalbán Basin (Artesilla, dated 16.49 Ma; San Roque 3, dated 16.33 Ma).

Location of the Calatayud-Montalbán Basin and the Teruel Basin in Spain (Modified from García-Paredes et al. 2010); Montalvos2 is situated in the Teruel Basin
Sediments of the Calatayud-Montalbán Basin (Fig. 1) have provided the data needed to define both the Aragonian and Ramblian stages and their associated small mammal biozones (Daams et al. 1977; Daams et al. 1987). The Aragonian stage and the included biozones have undergone significant revisions and updates since their original definition (Daams and Freudenthal 1981; Freudenthal and Daams 1988; Daams et al. 1999a, b; Van der Meulen et al. 2012). The combination of the Aragonian biozones, based on a densely sampled record of, in many cases, superposed localities with high-resolution magnetostratigraphical data, has resulted in a well-established chronological framework for this time slice and region (Krijgsman et al. 1994, 1996; Daams et al. 1999a; Alcalá et al. 2000; Van der Meulen et al. 2012). However, in the absence of magnetostratigraphical data from the Olmo-Redondo–San Roque sections, the dating of the early Aragonian (late early Miocene) mammal faunas in the type area, which includes zone A to Ca assemblages, is inferred from assumed sedimentary rates (Daams et al. 1999a; Van Dam et al. 2006).
The late early Aragonian is characterised by a mild turnover and species enrichment phase in the mammal succession from the Calatayud-Montalbán Basin (Van der Meulen et al. 2005; Hordijk 2010). The mammal assemblage from Montalvos2 is located outside this basin, and its mammal assemblage comprises an uncommon combination of species. We used the presence of species, taxon-free characteristics (i.e. a ridge index), relative abundances and a method to estimate mean annual precipitation to interprete the Montalvos2 assemblage and thereby provide additional and new insights in the biostratigraphy and palaeoecology of the region.
Locality, material and methods
The geological and stratigraphical position of Montalvos2 is special, as it is situated precisely at the northernmost extension of the NNE–SSW-running fault bordering the basin. The small lacustrine unit that contains the site of Montalvos2 directly overlies the Jurassic limestone that constitutes the local basement (approx. 1 m below the site). The fossiliferous bed consists of grey marl and is situated between limestone beds. Higher up in the sequence, lacustrine deposits are replaced by reddish clastic sediments. A thick series of dominantly conglomeratic sediments represents the proximal basin infill that is ultimately overlain by the “classical” late Miocene distal fluvio-lacustrine sediments that are well-known for their rich mammal record of the Teruel Basin (Van de Weerd 1976; Van Dam et al. 2001).
Montalvos2 contains four mammalian orders, as represented by 490 dental elements of rodents, 247 elements of lagomorphs, 37 elements of insectivores and one tooth of a marsupial (numbers excluding the incisors). The material was collected by the late Remmert Daams, and Albert van der Meulen, and the sediments were wet-screened by using a set of screens, the finest of which had a 0.5-mm mesh. The fossils are stored in the collection of the National Museum of Natural History Madrid (Spain).
The measurements of the dental elements were taken with a reflex microscope (Prior S2000, ocular 25×; Prior Scientific, Cambridge, UK) and with the Leitz Ortholux measuring microscope equipped with mechanical stage and measuring clocks (Esselte Leitz GmbH & Co KG). All measurements (Appendix 1) are given in millimetres. Tooth size measurements and morphology of the ochotonids were taken following the procedure of Hordijk (2010).
The Lagomorpha are described here in detail because lagomorphs have received considerable less attention in the literature. Prolagus lopezmartinezae is only known from a few localities and is not abundant in fossil assemblages. For the following reasons there is a strong focus on the p3, P2 and P3 in the descriptions, elements which are usually the most distinctive in pika taxonomy. First, the other cheek teeth, the p4–m1 in particular, are generally less informative in taxon identifications, although these elements may possess taxon specific traits. Secondly, many specimens in the available pika material of Montalvos2 are damaged to a variable degree. The P4–M2 and p4–m2-3 are very poorly preserved, if at all. Thirdly, the available D3 and D4 material in the Montalvos2 assemblage is fragmentary and is very likely a mix of both species. Both the level of damage and the worn state of the specimens prevent reliable identification of these elements.
We follow the taxonomic interpretation of Van der Meulen et al. (2003) and consider Fahlbuschia, Pseudofahlbuschia and Renzimys to be synonyms of Democricetodon. In the family assignment of the genus Prolagus, we prefer the use of the family name Ochotonidae over the family name Prolagidae Gureev, 1960, as has been proposed by Erbajeva (1988, 1994) and endorsed by Hoffmann and Smith (2005), following the argumentation of Hordijk (2010) and Angelone et al. (2014).
The nomenclature of the tooth morphology used is that of Rössner and Heissig (1999) for the Rodentia, Van den Hoek Ostende (2003) for the Eulipotyphla and Hordijk (2010; which is generally in line with López-Martínez 1974; 1989) for the Lagomorpha. Assuming that similarity in dental morphology indicates similarity in diet (Mayr 1979), the dentition types of extinct and extant glirid genera can be grouped, assuming that the diet of the genera within each group was similar to that of the extant member of that group (Van der Meulen and De Bruijn 1982; Daams et al. 1984). Since the extant dormice are a relict showing very specialised traits, such as hibernation (an adaptation to seasonality), the loss of the caecum (an adaptation to a protein-rich diet) and arboreal life style (except for Myomimus), it seems most unlikely that all were shared by the diverse array of extinct genera. This means that the actualistic approach may have a limited result for the Gliridae.
In order to avoid the effect caused by over-splitting on the generic level, as well as the difficulty of allocating extinct genera to a specific type of dentition, we use here the method of De Bruijn (1998), who introduced a ridge index that is based on the total number of ridges counted on all upper and lower first and second molars in an association divided by the number specimens. This method has the disadvantage that it disguises the distribution over the various taxonomic classes because the number of ridges on the molars of different species within the same genus may vary (García-Paredes 2006).
Quantitative data in fossil assemblages provide a valuable source of information. Relative abundances of species in fossil mammal assemblages can be determined in various ways. For quantitative analysis of the Montalvos2 material, we follow the method adopted in several previous studies (e.g. Daams et al. 1988; Van Dam and Weltje 1999; Van der Meulen et al. 2005) and base relative abundance estimates of rodent and insectivore species on the sum of the first two upper and lower molars. In the Gliridae and Sciuridae, the M1 and M2 are not separated. Because of the high diagnostic value of ochotonid premolars in taxonomy and the lack of properly preserved ochotonid molars in the Montalvos2 assemblage, the relative abundance of ochotonid species is determined by the number of upper and lower third premolars multiplied by two, following Hordijk (2010). Although based on different elements, this relative abundance value for ochotonids is equivalent to that used for the rodents. Specifications on number of elements, measurement and collection numbers are provided in Appendix 1.
To estimate mean annual precipitation (MAP) we used a method based on the percentages of invertivore species (PI) and arboreal species (PA) in the total fauna of Rodentia, Eulipotyphla and Lagomorpha [Van Dam 2006; MAP for the NW Old World (i.e. Europe, Middle East and North Africa)]. The formula employed is MAP (mm) = 0.179 + 14.134 PI + 18.066 PA (R 2 = 0.69), where PI = 100 × NI/N (with NI being the number of invertivore species and N being the total number of species) and PA = 100 × NA/N (with NA being the number of arboreal species).
Systematic palaeontology
In the faunal list (Table 1) all species are listed, including their taxonomic placement and authorities. Here we briefly introduce the most important taxa.
Order Rodentia Bowdich, 1821
The order Rodentia is represented by 15 species from four families, of which the Muridae is quantitatively the best represented. Here we also report on species of the Gliridae, Sciuridae and Eomyidae families.
Family Muridae Illiger, 1811
Subfamily Melissiodontinae Schaub, 1920
Melissiodon dominans (Fig. 2) has teeth that fall in size and morphology within the range of the type material (Wintershof-West, Germany; MN3); however, apart from the m2 and m3, the molars are distinctly larger than those from the localities Forsthart and Rembach in the lower part of the Freshwater Molasse of Bavaria (Germany) that otherwise contain the same murid association (Ziegler and Fahlbusch 1986). Melissiodon is known from two localities in the Calatayud-Montalbán Basin (biozone A, approx. 17 Ma), from Buñol (MN4; Prov. Valencia; Daams 1976) and from a MN4 locality in the Ribesalbes-Alcora Basin (Crespo et al. 2014).

Melissiodon dominans. a M3 dex (MNT2 354), b M2 dex (MNT2 351), c M1 dex (MNT2 346), d m3 dex (MNT2 361), e m1 dex (MNT2 357). Scale bar: 1 mm
Subfamily Cricetinae Fischer, 1817
Democricetodon decipiens from Montalvos2 is of the same size as D. decipiens from Buñol (Freudenthal and Daams 1988) and D. decipiens from several other localities in the Aragonian type area (Calatayud-Montalbán Basin, OR4A, SR3, ART1; in Van der Meulen et al. 2003). An exception is the M2, which is slightly shorter and overlaps the size range of D. hispanicus Freudenthal, 1967. The mesolophs and mesolophids are never long, and the metalophule is always directed forwards.
Megacricetodon primitivus from Montalvos2 is of about the same size as the material from the type-locality, Valtorres (early Aragonian, zones C and D; Freudenthal 1963). The m1s from Valtorres, however, are more elongate than our m1s (L/W ratio is 1.49 in Montalvos2 and 1.56 in Valtorres), and the width of the M1 from Montalvos2 is slightly larger.
Eumyarion weinfurteri from Montalvos2 has somewhat larger teeth than E. weinfurteri from Dolnice 1–3 and Orechov (Czechoslovakia; Fejfar 1974). They are about the same size as E. weinfurteri molars from Aliveri (Greece; Klein Hofmeijer and De Bruijn 1988), although in Montalvos2 the teeth are slightly wider. A simple anterocone, no mesostyl and a poorly developed or absent anterolophulid of the m1 are all characteristics of E. weinfurteri.
Family Gliridae Muirhead, 1819
The family Gliridae is represented in Montalvos2 by five species groups, differentiated by their dental morphology.
The Peridyromys/Pseudodryomys group:
In the Peridyromys/Pseudodryomys group the genera share the basic glirid pattern, consisting of the four main ridges plus the two centrolophs and sometimes an extra ridge between the protoloph and the anterior centroloph in the upper molars (M1-2), and the four main ridges plus a centrolophid of medium length and sometimes an extra ridge in the valley between the mesolophid and the posterolophid in the lower molars (m1-2).
In this group, a larger part of the ‘genera’ is included in the ‘subfamily’ Myomiminae sensu Daams and De Bruijn (1995). However, disregarding endemic island forms, the group contains Peridyromys, Pseudodryomys, Myomimus, Miodyromys, Prodryomys and Ramys. The differences in dental pattern shown by these ‘genera’ are recognised as being very subtle due to descriptions of generic type species that are intermediate in size and morphology. This phenomenon is especially obvious in the case of Pseudodryomys and Peridyromys. When one of us (HdB) first defined the genus Pseudodryomys (type P. ibericus), it included the species that are now correctly separated into the genus Simplomys García-Paredes, 2009 (in García-Paredes et al. 2009). Moreover, Peridyromys was at the time known by its type species only, P. murinus, a small animal with a simple basic glirid dental pattern, which is clearly different from the much larger Pseudodryomys ibericus occurring in the same sites. With our present knowledge, separating the species P. ibericus from P. murinus on the generic level seems a case of over-splitting, so we believe it would be a good idea to synonymise these genera. Obviously, there is a need for a revision of this group, an action that is well beyond the scope of this paper.
The cheek teeth with a Pseudodryomys/Peridyromys morphology from Montalvos2 constitute a homogeneous sample (Fig. 3) and resemble those of Pseudodryomys ibericus and Peridyromys darocensis. However, they are intermediate in size between these species and have less frequently an extra ridge in the valley between the protoloph and anterior centroloph in the M1-2 (4/13) and between the mesolophid and the posterolophid (3/18) than in either of these species (García-Paredes 2006). Since our association contains specimens that are identical to ones from the type localities of these two species, we arbitrarily allocate the material from Montalvos2 to Pseudodryomys aff. ibericus De Bruijn, 1966.

Scatter plot of Gliridae (MNT2), M1-2. Open circle Pseudodryomus ibericus, ☓ Simplomys julii, open triangle Simplomys sp., open square Simplomys simplicidens
The Simplomys group:
In the Simplomys group the morphology of the M1-2 and the m1-2 consists of the four main ridges only. Other dental characteristics of the species included into the Simplomys group are the strong reduction of the P4/p4 and M3/m3.
The Simplomys group currently contains five species: S. simplicidens (type species), S. robustus, S. julii, S. meulenorum and S. aljaphi. These species have very similar dental morphologies and differ primarily in size and, in the case of S. meulenorum, in the degree of reduction of the premolars and of the m3. The bulk of the material from Montalvos2 has the size of S. simplicidens (Fig. 3), but there are a few outliers, which are assigned to S. julii and Simplomys sp. (dental elements that differentiate between S. meulenorum and S. robustus are absent). The S. simplicidens teeth from localities assigned to the early/middle Aragonian (MN4) are somewhat narrower than the ones from the middle Miocene (García-Paredes 2006; García-Paredes et al. 2009).
The Microdyromys group:
In the Microdyromys group, the M1-2 share the presence of a complete endoloph, a protoloph and metaloph that do not strongly converge lingually, while both centrolophs plus a species-specific number of extra ridges may be developed. The m1-2 of Microdyromys are not characteristic and may be easily confused with those of the smaller, more complex species of the Peridyromys/Pseudodryomys group (Daams and De Bruijn 1995).
In Montalvos2, the Microdyromys group is represented by a small species that may have an anterior extra ridge in the M1-2 and/or a posterior extra ridge in the m1-2. Four species with these dental characteristics have been formally named: Microdyromys koenigswaldi, M. legidensis, M. monspeliensis and M. remmerti. Among these, M. remmerti is the largest in comparison to the others (García-Paredes et al. 2010).
The type material of M. koenigswaldi is slightly larger than that of M. legidensis, and that of M. legidensis is slightly larger than that of M. monspeliensis. García-Paredes (2006), who studied the sequence of samples of the Gliridae from the type area of the Aragonian, suggested synonymising M. legidensis from the early Miocene with the somewhat larger M. koenigswaldi from the middle Miocene, whilst identifying the smaller specimens from the middle Miocene as M. monspeliensis. Van der Meulen et al. (2012) rehabilitate M. legidensis and include all in the lineage M. legidensis-koenigswaldi. The teeth of Microdyromys legidensis from Montalvos2 are identical to those of the type material of M. legidensis (= M. koenigswaldi sensu García-Paredes 2006).
The Glirudinus group:
In the Glirudinus group, the occlusal surface of the cheek teeth is (almost) flat and bears a large number of very fine parallel ridges that make an angle with the longitudinal axis of the teeth. Records of the genus Glirudinus in Spain are known from late Oligocene until late Miocene, where species diversity is low and the number of specimens per locality is usually limited. Other than from Montalvos2, G. gracilis is known in Spain from the locality of Buñol (Province of Valencia; Daams 1976) and from the Vallès-Penedès Basin (Agustí et al. 1985; Casanovas-Vilar et al. 2010). Here, the material of this genus assigned to Glirudinus gracilis (Fig. 4) is also quite poor. Species diversity in Glirudinus seems to have been largest in Anatolia and in Central Europe (Ünay et al. 2003).

Glirudinus gracilis. a M3 dex (MNT2 529), b M1-2 sin (MNT2 525 c m3 dex (MNT2 524). Scale bar: 0.5 mm. Underlined letter indicates inverse specimen
The Armantomys (= Quercomys ) group:
Finally, in the Armantomys (= Quercomys sensu Daams 1991) group the dental pattern of the M1-2 and m1-2 consists of the four main ridges. Other than in Simplomys, these teeth show a tendency to develop a detached metaloph in the M1-2 and are hypsodont. In the Montalvos2 association, the Armantomys (= Quercomys) group is represented by very few and poorly preserved teeth. The genera Armantomys and Praearmantomys are unique within the family Gliridae in having cheek teeth with ridges that are high relative to the crown basis and in being geographically restricted to the Iberian peninsula. Two evolutionary lineages have been reconstructed within Armantomys (Daams 1991). The material from Montalvos2 represents one of the larger species, Armantomys aragonensis.
Family Eomyidae Winge, 1887
In Montalvos2, the Eomyidae are represented by a few teeth only, here assigned to Ligerimys ellipticus. These teeth are morphologically similar to those of L. ellipticus from Buñol (typelocality). Some of the M1-2, M3 and m3 from Montalvos2 are slightly wider than the molars from the typelocality, but they fall within the size ranges of L. ellipticus from Vargas 1A, Olmo Rodondo 8 and San Roque 3 (Álvarez-Sierra 1987).
Family Sciuridae Fischer, 1817
The Sciuridae are represented in Montalvos2 by two species of the subfamily Sciurinae Fischer de Waldheim, 1817 and one species of the subfamily Pteromyinae Brandt, 1855.
The Heteroxerus rubricati teeth correspond in size with teeth of H. rubricati from several Spanish localities (Cuenca Bescós 1988).
One xerine tooth, an m2 (Fig. 5a), is identified as Heteroxerus cf. grivensis. The tooth clearly exceeds the size of the H. rubricati teeth. In terms of size and morphology, it compares well with the m2 of H. grivensis from several Spanish localities (Cuenca Bescós 1988) and it is similar in morphology but smaller than m2 of Atlantoxerus blacki (De Bruijn 1967) from Spain (Peláez-Campomanes 2001). The presence of a species close to H. grivensis in the Montalvos2 fauna is unexpected because H. grivensis is only known from younger faunas (Van der Meulen et al. 2012). Van der Meulen et al. (2012) indicate two new but undescribed Heteroxerus species in their Figs. 2 and 6. Apparently, the development of Heteroxerus in early and middle Miocene times is more complex than has been thought earlier.

Heteroxerus cf. grivensis. a m2 sin (MNT2 625), Albanensia cf. sansaniensis, b m3 dex (MNT2 623). Scale bar: 1 mm. Underlined letter indicates inverse specimen

Lagopsis penai, a p3 dex (MNT2 817B), b p3 dex (MNT2 812A), c p3 sin (MNT2 803A), d p3 sin (MNT2 804A), e P3 dex (MNT2 847B), f P3 dex (MNT2 851B), g P3 dex (MNT2 856A), h D2 sin (MNT2 932A), i D2 dex (MNT2 936A), j P2 sin (MNT2 892B), k P2 dex (MNT2 896B), l d3 sin(MNT2 922A), m d3 dex (MNT2 927A), n d4 dex (MNT2 928B). Scale bar: 1 mm. Underlined letter Inverse specimen
Albanensia sansaniensis is represented by an m3 (Fig. 5b). The tooth appears rather short, as compared with m3s from the type locality Sansan (France; Ginsburg and Mein 2012).
Order Lagomorpha Brandt, 1855
Family Ochotonidae Thomas, 1897
The ontogenetic stage of individual specimens plays a role in determining the presence or absence of particular morphological features. Chronospecies subdivisions in various ochotonid lineages are based on shifts in the occurrence frequency of particular tooth characteristics within an assemblage through time (López-Martínez 1984, 1988; Hordijk 2010).
Lagopsis penai
In the P2 the paraflexus and mesoflexus are narrow and oblique, with their apices oriented in postero-labial direction. The mesoflexus is mostly straight to lightly curved. The lagicone is well developed, usually intermediate in length and size between the smaller postlobus and larger hypercone. A mesial hyperloph is absent. The lingual side of the hypercone is smooth in most specimens, but several show a weak to distinct indentation or incipient hypoflexus.
In general, the P3 has a trapezoidal-like shape in occlusal view, in which the mesial hyperloph lies almost parallel to the distal hyperloph and is usually thickest in its central part. The mesoloph length relative to the tooth width depends on the stage of wear. The postlobus usually extends widely on the labial side. The tip of the precone generally reaches to the position of the mesoflexus. The paraflexus has a distinct J-shape, which is narrowest at the anterior side of the lagicone. The lagicone is large and oval and is oriented in a antero-lingual direction. In ontogenetically younger specimens, a straight mesoflexus is present, which is distinctly more shallow or absent in more deeply worn specimens. A distinct shallow hypoflexus is visible in all specimens that have this part preserved.
In the p3, the anteroconid is generally triangular, wide and well developed. In a few specimens, however, a more simple and smaller diamond-shaped anteroconid is observed. The anteroconid is larger than the metaconid in all but two adult specimens. Proto- and paraflexid are always present, but may vary in depth. The metaflexid is distinctly developed toward the midsection of the occlusal surface of the tooth in all specimens, but its shape is variable. The majority of specimens (27) show a distinct anterior extension, while the outline of the the rest (12) is more symmetrical. The shape of the mesial side of the anteroconid is variable. An anteroflexid is absent or incipient in the majority of specimens, while in a few, a more distinctly developed undulation is visible. Two specimens have crown cement preserved in the anteroflexid. The metaconid is relatively small and has a slightly oval to somewhat rounded rectangular shape. Its width is usually larger than its length, and this conid represents the most lingual extension of the tooth. The posterior lobe, formed by the entoconid and hypoconid has a more or equal thickness and is slightly curved labially, placing the hypoconid in a more anterior position.
The morphology of the D2 strongly resembles that of the P3 in having a very similarly shaped lagicone, postlobus and paraflexus. The paraflexus is deep and strongly curved, nearly reaching the posterior wall of the tooth. The mesoflexus is variably present and is usually straight. The mesial hypercone is distinctly more developed than the distal hypercone.
The d3s are slightly more robust and wider than the specimens provisionally assigned to P. lopezmartinezae.
All of the d4 specimens are provisionally assigned to L. penai on the basis of size, which fits that of the d3 assigned to this species. The tooth consists of two lobes that are isolated during most stages of wear but which are connecting in strongly worn specimens. The talonid lobe is wider than the trigonid lobe in relatively worn specimens. The general shape of the occlusal surface of the trigonid and talonid lobes strongly resembles that of the p4.
Prolagus lopezmartinezae
In the P2, the mesoflexus and paraflexus are straight and oriented in a posterior direction (Fig. 7). One slightly worn specimen shows an additional enamel loop and protrusions, which would disappear with wear. The hypercone forms the largest cone of the tooth. The mesial hyperloph varies slightly in thickness and length, but is generally short. It does not cover the paraflexus in any of the specimens, but does represent the most anterior part of the tooth. The width of the distal hyperloph decreases from lingual to labial, which is primarily due to the mesoflexus almost reaching the posterior wall. A hypoflexus is absent (Fig. 7).
In the P3 specimens, the postlobus is well developed in specimens where it has been preserved, but represents a smaller portion of the width of the tooth in comparison to the width covered by the mesial hyperloph, which extends over 50 % of the tooth width. A precone is present in all but one specimen where this feature can be observed. The mesoflexus is deep and can vary in shape from being curved in various degrees to completely straight. In the latter case, the shape of the lagicone is somewhat triangular, and a lagiloph is lacking. In specimens with a curved mesoflexus, the lagicone usually has a short, but discernable lagiloph that does not reach the outline of the tooth. One incomplete specimen has a relatively long lagiloph, and the lagicone is compressed lingually, resulting in a lingual bulge. A shallow cement-filled hypoflexus is present in all specimens in which this part of the tooth has been preserved. The mesial hypercone tends to be less developed and placed more labially than the distal hypercone.

Prolagus lopezmartinezae. a p3 dex (MNT2 837A), b p3 dex (MNT2 833A), c p3 sin (MNT2 826B), d p3 dex (MNT2 832A), e P3 dex (MNT2 878A), f P3 sin (MNT2 864B), g P3 sin (MNT2 862B), h P3 sin (MNT2 861B), i P2 sin (MNT2 913A), j P2 sin (MNT2 914A), k P2 dex (MNT2 917A), l D2 dex (MNT2 940A), m d3 dex (MNT2 921B). Scale bar: 1 mm. Underlined letter indicates inverse specimen
The P4-M2 are very fragmentary and poorly preserved. One fragment showing a hypoflexus that spans roughly one half to two thirds of the tooth width and has a small curved parafossette which resembles the morphology of the M1 known from San Marcos (Hordijk 2010). A very small fragment containing only a parafossette that is somewhat larger and placed more lingually than that of the surmised M1 is tentatively identified as a worn P4 fragment.
The p3 has a triangular outline of the occlusal surface. The anteroconid is somewhat sub-round to more or less ellipsoid in two specimens while being slightly anteriorly flattened on two sides, vaguely resembling a triangle. A third specimen shows a more clear triangular shape. The anteroconid is isolated in all specimens in which this feature can be verified. One of the anteroconids shows a weak anteroflexid along the crown. The protoistmid is present in all specimens, while a metaistmid is absent. A metaflexid is well developed and usually follows a path parallel to the width of the tooth, curving sharply toward the posterior side at the centre of the tooth. Three specimens also show a very shallow anterior extension. The entoconid shows one or more small ‘crochet’-like structures at, or close to, the curve of the metaflexid in half the number of the specimens. The protoconid and metaconid are broadly connected. A shallow centroflexid is present in all specimens, ranging from a very shallow indentation between the protoconid and metaconid to a distinct narrow short inflexion that is slightly narrowing the protoconid–metaconid connection. The protoconid is smaller than the metaconid; it is truly very small or incipient and clearly visible in all but two specimens. The entoconid has a relatively rectangular shape, while the hypoconid is more rounded and drop-like. The hypoflexid is deep, ending close to the posterior wall of the tooth, similar to the metaflexid.
The damaged D2 has a morphology which strongly resembles that of the P3 in terms of shape of the lagicone, metaflexus and paraflexus.
The three d3 have been provisionally assigned to P. lopezmartinezae because they are smaller than the specimens provisionally assigned to L. penai. The shape of these teeth is more slender and narrower than those assigned to L. penai. Unfortunately, two of the specimens show considerable damage.
The morphology and size of the material of P. lopezmartinezae strongly resembles that from the type locality (San Marcos) of the species with which it has been directly compared. The combination of the primitive and derived morphologies of P3, p3 and P2 as observed in P. lopezmartinezae represents an intermediate stage in the evolutionary lineage of P. vasconiensis Viret, 1930–P. lopezmartinezae–P. tobieni López-Martínez, 1977 (Hordijk 2010).
Order Eulipotyphla Waddell, Okada and Hasegawa, 1999
The Eulipotyphla (formerly Insectivora) are represented by five species, belonging to four families.
Family Erinaceidae Fisher, 1814
According to Van den Hoek Ostende and Doukas (2003), two species of Galerix were identified in the Calatayud-Montalbán Basin in biozones B-Da. The small Galerix symeonidisi would have gradually been replaced by the middle-sized G. exilis. This assumption is based on the gradual decrease of P3 bearing a hypocone and the size variation that was too large for a single species. However, since the size ranges of the two species overlap, it is difficult to assign individual elements to the one or the other. The same problem arises in the small Galerix assemblage from Montalvos2. Here, we tentatively assign all elements to G. symeonidisi. The only P3 present bears a hypocone, which is indicative of G. symeonidisi. Furthermore, the elements from Montalvos2 fall within the lower part of the size ranges found for this species in the Calatayud-Montalbán Basin area. Other symeonidisi-like characters are the well-developed trigonid of the p4 and the absence of a protocone–metaconule connection in the M1 and M2. These characters are variable, however, and can only be used in large assemblages. Therefore, we can not entirely exclude the possibility that G. exilis was also present in Montalvos2.
Family Talpidae Fischer, 1817
The mole species cf. Desmanella sp., a representative of the Uropsilinae, has a symmetrical P4, which features a large paracone and a well-developed, conical protocone. This well-developed conical protocone of the P4 identifies this small talpid as an Uropsilinae. Van den Hoek Ostende (2001) showed that this subfamily was quite diverse in Europe during late Oligocene/early Miocene times, represented by such genera as Asthenoscaptor, Desmanella and Mygatalpa. Without the characteristic upper molars, the genus cannot be unequivocally determined. We tentatively assign the Montalvos2 material to Desmanella, which to date is the only uropsiline found in Iberia. The only early Miocene record of the genus is D. fejfari from Rubielos de Mora (Gibert 1975) and from Alto de Ballester (Van den Hoek Ostende et al. submitted). The premolars are comparable in size to the material from these two MN3 localities, but the m3 are about 10 % shorter.
The second talpid species from Montalvos2 is Desmanodon sp. The upper molars of this species feature a relatively well-developed hypocone, particularly on the M2 and M3. These characters are in agreement with Desmanodon from the Daroca area (Van den Hoek Ostende 1997).
The presence of two talpid species in Montalvos2 is remarkable. In localities belonging to zones B and C in the Calatayud-Montalbán Basin only Desmanodon has been found.
Family Heterosoricidae Viret and Zapfe, 1951
The taxonomy of the Heterosoricidae is largely based on mandible characters (Engesser 1975; Ziegler 2009). However, there are also a number of useful characters to distinguish the various genera on the basis of dental characteristics (Engesser 1980; Ziegler 1998). One of the differences between the Miocene genera Dinosorex and Heterosorex is the weaker development of the lingual cusps of the upper molars in the latter. Since these cusps are weakly developed in Montalvos2 (the hypocone is not even discernible as a separate cusp), this heterosoricid is referable to Heterosorex. This is corroborated by the outline of the P4, which has a triangular shape and is not square as is usual in Dinosorex. The Montalvos2 material agrees in size and morphology well with Heterosorex neumayrianus from various German MN 3/ MN 4 localities (Doben-Florin 1964; Ziegler and Fahlbusch 1986; Ziegler 1989; Klietmann et al. 2014b).
Family Soricidae Fischer, 1814
The unidentifiable soricid (Soricid gen. et sp. indet.) has lower molars with a well-developed entoconid and which are entocristed, barring identification as Paenelimnoecus. The talonid of the only m3 is strongly reduced. The damaged M2 is characterised by a moderate posterior emargination and a ridge-shaped hypocone.
Order Didelphimorpha Gill, 1872
Family Didelphidae Gray, 1821
Subfamily Herpetotheriinae Trouessart, 1879
Marsupials are represented in Montalvos2 by a worn labial fragment of an upper molar only.
It cannot be further identified to the genus or species level and is assigned to Amphiperatherium, the only genus present at this time (Furió et al. 2012). The small marsupial Amphiperatherium is quite common in contemporary faunas from southern Germany (Ziegler and Fahlbusch 1986; Klietmann et al. 2014a). Marsupials have also been found in faunas from the Calatayud-Montalbán Basin (L. van den Hoek Ostende, personal observation), but unfortunately, the material was lost.
Biostratigraphy
The presence of Democricetodon decipiens places Montalvos2 in local zone Ca, the Megacricetodon primitivus–Democricetodon decipiens zone, with an age of 16.6–16.2 Ma, as defined by Van der Meulen et al. (2012) in the neighbouring Calatayud-Montalbán Basin (Fig. 8). According to these authors, zone Ca (together with zones B and Cb) is correlated to the European MN4 (Fig. 8). In the Calatayud-Montalbán Basin, zone Ca is represented by three sites: Artesilla (16.49 Ma), San Roque 3 (16.33 Ma) and Olmo Redondo 4A (16.30 Ma). We regard the presence of Ligerimys ellipticus in Montalvos2 as an additional constraint, implying an age younger than that of Artesilla (16.49 Ma), which still contains L. florancei. This inference is further confirmed by the absence in Montalvos2 of Praearmantomys crusafonti, one of the most abundant forms (21 %) in the rodent fauna of Artesilla (Van der Meulen and Daams 1992).

The biostratigraphic position of Montalvos2. Indicated are a selection of key events of rodents and lagomorphs in the Calatayud-Montalbán Basin and the FO of several chronospecies in the time slice of interest. The correlation of Aragonian mammal sites to the Astromonically Tuned Neogenic Time Scale of Lourens et al. (2005) (ATNTS 2004) is after Van der Meulen et al. (2012). FO: First occurrence; LO: Last occurrence
Furthermore, we note that San Roque 3 and Olmo Redondo 4A lack Eumyarion, a form present in Montalvos2 and in the next younger locality in the Calatayud-Montalbán sequence Vargas 4A, at the base of next zone Cb (16.15 Ma). Another similarity between Montalvos2 and Vargas 4A is the presence of Petauristinae, although these are very rare. Based on these similarities and differences, a placement of Montalvos2 between Olmo Redondo 4A and Vargas 4A seems probable (thus between 16.30 Ma and 16.15 Ma).
In terms of the most abundant family, the Gliridae, a placement of Montalvos2 in MN4 is not inconsistent with the stratigraphical ranges of the glirid species present. Although these are long and usually cover several MN-zones, the ages of three of seven type-localities of the species recognised in the assemblage from Montalvos2 correlate to MN4, whereas one correlates to MN2, two to MN3 and one to MN5. The concurrent range zone of Simplomys simplicidens, S. robustus and S. julii on the Iberian peninsula covers the local zones A, B and the lower part of C, which are correlated with the upper part of MN3 plus the lower part of MN4 (García-Paredes et al. 2009).
The ranges of the ochotonid species provide only limited biostratigraphical evidence for Montalvos2. Both Lagopsis penai (zone A-Dd) and Prolagus lopezmartinezae (zone A-Cb certain, Da-G1 lack of data) first appear in the Late Ramblian and continue into the middle Aragonian (MN3–5) (Hordijk 2010). Nevertheless, the available ochotonid material fits well with a correlation to MN4.
The Lagopsis penai from Montalvos2 is indistinguishable from L. penai from the nearby Calatayud-Montalbán Basin. Lagopsis penai is known to have a fairly stable size range through time, as well as a relatively high variability range in both size and morphology within assemblages (Hordijk 2010). Specific primitive or derived morphological features occur in assemblages throughout the Lagopsis lineage during the Miocene and thus do not provide a strong argument for precise biostratigraphical correlations. This is illustrated by the occurrence of two p3 with a cement-filled anteroflexid in the Montalvos2 assemblage, a feature that is associated with Lagopsis verus (Hensel 1856) when dominantly present in an assemblage (FO in zone Dd; Hordijk and Van der Meulen 2010a).
The absence of Prolagus vargasensis Hordijk and Van der Meulen, 2010 (Hordijk and Van der Meulen 2010b) in Montalvos2, known in the Aragonian type area from the base of Cb and consistently present in mammal assemblages of this zone (Hordijk 2010), suggests the likelihood that the fauna from Montalvos2 is older than zone Cb.
Independent of the ranges of the ochotonid species, the relative high abundance of Lagopsis penai in the Montalvos2 assemblage is very similar to that of mammal assemblages in zone B known from the nearby Calatayud-Montalbán Basin (Hordijk 2010; Van der Meulen et al. 2012), suggesting a potentially similar age. However, this feature strongly depends on the local environment. The relatively close proximity of the Calatayud-Montalbán Basin and Teruel Basin does not guarantee similar environments and mammal abundance patterns at the scale of biozones.
The insectivore Desmanella has been found in biozone A localities from the Rubielos de Mora basin (Van den Hoek Ostende et al. submitted). Desmanodon is a quite common element in the late early/earliest middle Miocene faunas of Spain. In terms of size and morphology, the Montalvos2 Desmanodon agrees well with the finds from the Calatayud-Montalbán Basin. Heterosorex is found in various zone C localities in the Aragonian of the Calatayud-Montalbán Basin (Van der Meulen et al. 2012), and fragments referable to Heterosorex are also known from the Ramblian (Ramblar 1; Van den Hoek Ostende 2003).
Some of the faunal elements of the Montalvos2 assemblage point to a correlation with biozone B (e.g. lagomorph abundance, petauristines, some glirids); however, the presence of these species could also be an indication of differences in environmental conditions. That such differences played a role is also evident from the presence of some faunal elements [e.g. Heteroxerus (cf.) grivensis] that occur much later in the Calatayud-Montalbán Basin, although these latter species are usually present in low abundances which could have limited the chance of being found.
Here, we consider the presence of the cricetines Democricetodon decipiens, Megacricetodon primitivus and Eumyarion, combined with the presence of Ligerimys ellipticus, as the key stratigraphical information and constrict the biostratigraphical position of Montalvos2 to biozone Ca. Thus, the best estimate of the age of Montalvos2 lies between 16.4 and 16.2 Ma (Fig. 8).
It is also interesting to compare the fauna of Montalvos2 to that of Buñol in the Province of Valencia (Daams 1976; Robles et al. 1991; Hoek et al. 2005). Both faunas are time-equivalent and can be correlated to zone Ca, as defined in the Calatayud-Montalbán Basin. Assuming an isochronous presence of Ligerimys ellipticus, the age of Buñol can be estimated to lie between approximately 16.4 Ma (entry between Artesilla and San Roque 3), and 15.94 (Cb-D boundary). A specific parallel with Montalvos2 constitutes the presence of Melissiodon.
Palaeoecology and palaeoclimate
The relative abundances of the various small mammal species in Montalvos2 (Fig. 9; Table 2) show that rodents, despite comprising 15 species, are only slightly more abundant than the lagomorphs, represented by only two taxa (53 vs. 43 %, respectively). Two species are quite dominant, Lagopsis penai by approximately 32 % and Megacricetodon primitivus by approximately 22 %, followed by Prolagus lopezmartinez (approx. 11 %). The relative abundance of the small mammals in Montalvos2 is very similar to the abundance pattern in biozone B in the Calatayud-Montalbán Basin and differs from the abundances in biozones Ca and Cb.

The abundance distribution of the main small mammal orders present in Montalvos2 (Teruel Basin) compared to small mammal faunas of the Calatayud-Montalbán Basin
The relative abundance of ochotonids in the Calatayud-Montalbán Basin is highly variable in the time interval between zone B and H (MN4–9), comprising on average 19 % of the small mammal primary consumers in fossil assemblages (Hordijk 2010). Despite the overall variability, the relative abundances observed in zones B to Cb are fairly consistent within these zones, where those of zone B are clearly different from those of Ca and Cb. The average abundance of ochotonids in zone B is particularly high relative to that of the rodents, ranging between 22 and 57 % (average approx. 44 %), while there is a distinct decrease in ochotonid abundance going into zone Ca, ranging between 13 and 18 % (average approx. 16 %), as well as into zone Cb, ranging between 0 and 13 % (average approx. 7 %) (Hordijk 2010). The markedly high abundance of the ochotonids in the Montalvos2 assemblage, and of Lagopsis penai in particular, suggests similar environmental conditions as in zone B assemblages of the Calatayud-Montalbán Basin.
Despite the known environmental flexibility of Lagopsis penai, which was fairly widespread in Western Europe and has even been found in the context of a subtropical and fairly humid environment with a mix of open to more closely vegetated habitats (Angelone 2009; Tütken and Vennemann 2009), this species is usually associated with relatively warm and drier conditions (Mein 1984; López-Martínez et al. 1987; López-Martínez 1989, 2001). The high abundance of Lagopsis penai in particular and of the ochotonids in general suggest the availability of a relatively open and (semi)-arid landscape in the vicinity of the locality of Montalvos2.
The Eulipotyphla constitute 3.4 % of the smaller mammal assemblage from Montalvos2. This is comparable with the relative abundance of the insectivores in biozones A, B and C in the Calatayud-Montalbán Basin, where they usually make up less than 5 % of the total assemblage. Only in the top of zone Cb (Vargas 2A) is the insectivore percentage around 8 %.
A reconstruction of the diet and biotope requirements of the various Gliridae is necessarily based on tooth morphology; therefore, we use the ridge index of De Bruijn (1998).
The percentages of glirid genera from five MN4 assemblages from Greece (Aliveri, Karydia), Austria (Oberdorf) and Spain (Villafeliche 2A, Montalvos2) and the distribution of the classes of the number of ridges on the M1-2 and m1-2 are given in Table 3. The choice of the localities used for comparison is largely the result of availability in the Utrecht University collection. To ensure a degree of uniformity within this comparison, endemic genera have been omitted (Armantomys and Seorsumuscardinus; De Bruijn 1998; García-Paredes 2006). We realise that results based on such limited material from insufficiently calibrated localities can, at best, only roughly suggest whether this approach has potential. Moreover, the dental pattern of Peridyromys and Pseudodryomys is basically the same, and here we combine the count of ridges for these two genera.
The differences in taxon composition, as well as in the distribution of the ridge-classes (Table 3) between the glirid associations analysed suggest differences in habitat. Remarkably, the generally common genus Microdyromys is absent or rare in the Greek localities (Karydia and Aliveri respectively), and Peridyromys is absent in the association from Montalvos2, while the Spanish associations (Villafeliche 2A and Montalvos2) are very similar otherwise. We believe that these differences may well have a different origin. The absence of Microdyromys in Karydia could to be due to unfavourable circumstances, while the absence of Peridyromys in Montalvos2 may be either due to the unfavourable circumstances or to over-splitting on the generic level.
The data on the classes of ridge numbers suggest that within the range of ridge numbers chosen for study, there is a shift towards species with fewer ridges from SE to SW Europe. Van der Meulen and De Bruijn (1982) and Daams and Van der Meulen (1984) concluded that a vegetarian diet constitutes a smaller portion of the total diet of species with a limited number of ridges compared to those with many ridges. Moreover, these authors concluded that Microdyromys has a preference for a warm climate (and wet), whereas Peridyromys prefers a cool climate (and dry). Following these suggestions, it seems that the conditions may have been wetter in SE Europe than in SW Europe in the early part of MN4. The glirids from Montalvos2 thus suggest a semi-arid warm biotope.
The highly aberrant shape of the weak and slender mandible of Melissiodon was already noted by Schaub (1920) when he defined the genus. Subsequent workers in the field have commented on its unusual long tubular diastema with weak lower incisors that end below the m2 in combination with the complex fragile high-crested cheek teeth. In this context, it is of interest that the extant endemic shrew rats from Sulawesi and the Philippines which feed on soft-bodied invertebrates have mandibles that are very similar to those of Melissiodon (Musser 1990; Esselstyn et al. 2012). The combination of a long and weak mandible and high-crested cheek teeth in Melissiodon suggests that these murids were also feeding on insects (beetles?), thus making the members of this taxon, ecologically speaking, an insectivore. The continuous availability of this very specific food source may well be the reason that these murids survived the decline of the family in Europe during the early Miocene. Since Melissiodon is invariably rare in lacustrine deposits and more common in fissure infills, we interpret Melissiodon species to be inhabitants of dry limestone plateaus.
The overall percentage of eulipotyphlan insectivores can be used as a crude indicator of humidity (Van den Hoek Ostende 2001) as the percentages of insectivores present in Montalvos2 are not much different from those in the Calatayud-Montalbán Basin, suggesting similar humidity conditions. However, the presence of a second talpid suggests that the environment in Montalvos2 may have been somewhat more humid than in contemporaneous localities in the Calatayud-Montalbán Basin.
To estimate mean annual precipitation (MAP), the method of Van Dam (2006) is used, where MAP = 0.179 + 14.134 PI + 18.066 PA (R 2 = 0.69). The percentages of invertivore species (PI) and arboreal species (PA) are based on the fauna of Rodentia, Eulipotiphla and Lagomorpha (the marsupial Amphiperatherium was not included in the calculations), and thus the total number of species considered (N) amounts to 22. The large majority of the species recorded are considered to be ground-dwelling. The glirid Glirudinus and the flying squirrel Albanensia, however, can be considered to have had an arboreal life style (NA = 2). Melissiodon and all Eulipotyphla are assumed to have been insectivorous (NI = 6). PA and PI have respective values of 9.1 and 27.3 %, and the resulting best estimate for MAP is 550 mm/year. The 95 % prediction interval, which is a combination of the both uncertainty in the regression parameters and individual variation (i.e. the residuals), is 204–897 mm/year. Estimates for the MAP from time-equivalent (zone Ca) sites in the Calatayud-Montalbán Basin are lower at 283 mm/year (Artesilla), 404 mm/year (San Roque 3) and 250 mm/year (Olmo Redondo 4A); this decrease is caused by the lower number of arboreal (0–1) and/or invertivore species (3–5). It is therefore likely that annual rainfall levels could have been some 150–300 mm/year lower at these three sites than at Montalvos2. The next younger locality Vargas 4A (base zone Cb, NI = 8, NA = 1) would have had a MAP value only somewhat lower than that of Montalvos2 (estimated at 505 mm/year). The precipitation (MAP) estimate for Buñol amounts to 875 mm/year, i.e. much higher than that of Montalvos2 and sites near Daroca. This is mainly due to a much higher number of arboreal species (NA = 7, PA = 24 %, both dormice and flying squirrel species), pointing to a more closed environment. It is possible that both geographic and small temporal differences in climate can explain the differences in relative abundances, composition and relative humidity between Montalvos2 fauna in the Teruel Basin and the time-equivalent faunas from the more NNW-situated Calatayud-Montalbán Basin.
References
Agustí, J., Cabrera, L., & Moyà-Solà, S. (1985). Sinopsis estratigráfica del Neógeno de la fosa del Vallès-Penedès. Paleontologia i Evolució, 18, 57–81.
Alcalá, L., Alonso-Zarza, A. M., Alvarez-Sierra, M. A., Azanza, B., Calvo, J., Cañaveras, J. C., Dam, A. J. van, Garcés, M., Krijgsman, W., Meulen, A. J. van der, Morales, J., Peláez- Campomanes, P., Pérez-González, A., Sánchez-Moral, S., Sancho, R., & Sanz Rubio, E. (2000). El registro sedimentario y faunístico de las cuencas de Calatayud-Daroca y Teruel. Evolución paleoambiental y paleoclimática durante el Neógeno. Revista de la Sociedad Geológica de España, 13(2), 323–343.
Álvarez-Sierra, M. Á. (1987). Estudio sistematico y bioestratigrafico de los Eomyidae (Rodentia) del Oligoceno superior y Mioceno inferior Espanol. Scripta Geologica, 86, 207.
Angelone, C. (2009). Lagomorphs from the Miocene of Sandelzhausen (southern Germany). Paläontologische Zeitschrift, 83(1), 67–75.
Angelone, C., Prieto, J., & Gross, M. (2014). Complement to the study of the pikas (Lagomorpha, Ochotonidae) from the middle Miocene of Gratkorn, Austria. In M. Böhme, M. Gross, J. Prieto (eds) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobiodiversity and Palaeoenvironments, 94(1), 125–134.
Bowdich, T. E. (1821). An analysis of the natural classifications of Mammalia for the use of students and travellers (p. 115). Paris: J. Smith.
Brandt, J. F. von (1855). Beiträge zur nähern Kenntniss der Säugethiere Russlands. Mémoires de l’Académie Impériale des Sciences de Saint-Pétersbourg. 6e Série: Sciences Mathématiques, Physiques et Naturelles; 2e Partie: Sciences Naturelles, 9, 1–375.
Bruijn, H. de (1966). Some new Miocene Gliridae (Rod. Mam.) from the Calatayud area (province of Zaragoza, Spain). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B, 69, 1–21.
Bruijn, H. de (1967). Gliridae, Sciuridae y Eomyidae (Rodentia, Mammalia) miocenos de Calatayud (provincia de Zaragoza, España) y su relación con la bioestratigrafía del área. Boletín del Instituto Geológico y Minero de España, 78, 187–373.
Bruijn, H. de (1998). Vertebrates from the early Miocene lignite deposits of the opencast mine Oberdorf (Western Styrian Basin, Austria): 6. Rodentia I (Mammalia). Annalen des Naturhistorischen Museums Wien, 99A, 99–137.
Casanovas-Vilar, I., García-Paredes, I., Alba, D. M., Hoek Ostende, L. W. van den, & Moyà-Solà, S. (2010). The European Far West: Miocene mammal isolation, diversity and turnover in the Iberian peninsula. Journal of Biogeography, 37, 1079–1093.
Crespo, V. D., Ruiz-Sánchez, F. J., Freudenthal, M., & Montoya, P. (2014). Primer registro del género Melissiodon (Mammalia, Cricetidae) en la cuenca de Ribesalbes-Alcora (Castelló, España). Jornadas de Paleontología de la Sociedad Española de Paleontología, 24, 53–55.
Cuenca Bescós, G. (1988). Revisión de los Sciuridae del Aragoniense y del Rambliense en la fosa de Calatayud-Montalbán. Scripta Geologica, 87, 1–116.
Crusafont, M., Villalta, J. F., & de Truyols, J. (1955). El Burdigaliense continental de la cuenca del Vallés-Penedés. Memorias y Comunicaciones del Instituto Geológico, Diputación provincial de Barcelona, 12, 11–272.
Daams, R. (1976). Miocene rodents (Mammalia) from Cetina de Aragon (Prov. Zaragoza) and Buñol (Prov. Valencia), Spain. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B, 79(3), 152–182.
Daams, R. (1981). The dental pattern of the dormice Dryomys, Myomimus, Microdyromys and Peridyromys. Utrecht Micropalaentological Bulletin Special Issue, 3, 1–115.
Daams, R. (1989). Miscellaneous Gliridae from the Miocene of the Calatayud-Teruel Basin, Aragon, Spain. Scripta Geologica, 89, 13–26.
Daams, R. (1991). Hypsodont Myomiminae (Gliridae, Rodentia) from the Miocene and the Oligocene-Miocene boundary interval of Spain. Scripta Geologica, 95, 1–63.
Daams, R., & Bruijn, H. de (1995). A classification of the Gliridae (Rodentia) on the basis of dental morphology. Hystrix, 6(1–2), 3–50.
Daams, R., & Freudenthal, M. (1981). Aragonian: the stage concept versus Neogene Mammal Zones. Scripta Geologica, 62, 1–17.
Daams, R., & Freudenthal, M. (1988). Megacricetodon (Cricetidae) from the Aragonian and the Lower Vallesian of the Calatayud-Teruel Basin. In M. Freudenthal (Ed.), Biostratigraphy and paleoecology of the Neogene micromammalian faunas from the Calatayud-Teruel Basin (Spain). Scripta Geologica Special Issue 1, 39–132.
Daams, R., & Meulen, A. J. van der (1984). Paleoenvironmental and paleoclimatic interpretation of micromammal faunal successions in the upper Oligocene and Miocene of north central Spain. In J. Meulenkamp (Ed.), Paleoenvironnements continentaux en Méditerrannée au Néogène et évolution paléoclimatique (pp. 241–257). Paleobiologie Continentale, 14, Montpellier.
Daams, R., Freudenthal, M., & Weerd, A. van de (1977). Aragonian, a new stage for continental deposits of Miocene age. Newsletters on Stratigraphy, 6(1), 42–55.
Daams, R., Freudenthal, M., & Meulen, A. J. van der (1984). Ecostratigraphy of micromammal faunas from the Neogene of Spain. Scripta Geologica Special Issue, 1, 287–302.
Daams, R., Freudenthal, M., & Álvarez-Sierra, M. Á. (1987). Ramblian: a new stage for continental deposits of early Miocene age. Geologie en Mijnbouw, 65, 297–308.
Daams, R., Freudenthal, M., & Meulen, A. J. van der (1988). Ecostratigraphy of micromammal faunas from the Neogene of Spain. In M. Freudenthal (Ed.), Biostratigraphy and paleoecology of the Neogene micromammalian faunas from the Calatayud-Teruel Basin (Spain). Scripta Geologica Special Issue 1, 287–302.
Daams, R., Meulen, A. J. van der, Álvarez-Sierra, M. Á., Peláez-Campomanes, P., Calvo, J. P., Alonso Zarza, M. A., & Krijgsman, W. (1999a). Stratigraphy and sedimentology of the Aragonian (early to middle Miocene) in its type area (North-Central Spain). Newsletters on Stratigraphy, 37(3), 103–139.
Daams, R., Meulen, A. J. van der, Álvarez-Sierra, M. Á., Peláez-Campomanes, P., & Krijgsman, W. (1999b). Aragonian stratigraphy reconsidered, and a re-evaluation of the middle Miocene mammal biochronology in Europe. Earth and Planetary Science Letters, 165, 287–294.
Dam, A. J. van (2006). Geographic and temporal patterns in the late Neogene (12–3 Ma) aridification of Europe: The use of small mammals paleoprecipitation proxies. Palaeogeography, Palaeoclimatology, Palaeoecology, 238, 190–218.
Dam, A. J. van, & Weltje, G. J. (1999). Reconstruction of the Late Miocene climate of Spain using rodent palaeocommunity successions: an application of end-member modelling. Palaeogeography, Palaeoclimatology, Palaeoecology, 151, 267–305.
Dam, A. J. van, Alcalá, L., Alonso Zarza, A., Calvo, J. P., Garcés, M., & Krijgsman, W. (2001). The upper Miocene mammal record from the Teruel-Alfambra region (Spain). The MN system and continental stage/age concepts discussed. Journal of Vertebrate Paleontology, 21(2), 367–385.
Dam, A. J. van, Abdul Aziz, H., Álvarez-Sierra, M. Á., Hilgen, F. J., Hoek Ostende, L. W. van den, Lourens, L. J., Mein, P., Meulen, A. J. van der, & Peláez-Campomanes, P. (2006). Long-period astronomical forcing of mammal turnover. Nature, 443, 687–691.
Dehm, R. (1950). Die Nagetiere aus dem Mittel-Miozän (Burdigalium) von Wintershof-West bei Eichstätt in Bayern. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie Stuttgart, 91, 321–428.
Doben-Florin, U. (1964). Die Spitzmäuse aus dem Alt-Burdigalium von Wintershof-West bei Eichstätt in Bayern. Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematische-Naturwissenschaftliche Klasse NF, 117, 1–82.
Dobson, G. E. (1883). A monograph of the Insectivora, systematic and anatomical, Part II. Including the families Potamogalidae, Chrysochloridae, and Talpidae. John Van Voorst: London.
Doukas, C. S. (1986). The mammals of the lower Miocene of Aliveri (Island of Evia, Greece). Part 5. The insectivores. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B, 89(1), 15–38.
Erbajeva, M. (1988). Piščuhi Kajnozoja: taksonimija, sistematika, filogenija. Moscow: Akademija Nauk SSSR, Sibirskoe otdelenie, Burjatskij filial, Geologičeskij institut.
Erbajeva, M. A. (1994). Phylogeny and evolution of Ochotonidae with emphasis on Asian Ochotonids. In Y. Tomida, C. K. Li, & T. Setoguchi (Eds.), Rodent and lagomorph families of Asian origins and diversification (pp. 1–13). Tokyo: National Science Museum.
Engesser, B. (1975). Revision der europäischen Heterosoricinae (Insectivora, Mammalia). Eclogae Geologicae Helvetiae, 68(3), 649–672.
Engesser, B. (1980). Insectivora und Chiroptera (Mammalia) aus dem Neogen der Türkei. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B, 102, 37–363.
Esselstyn, J. A., Achmadi, A. S., & Rowe, K. C. (2012). Evolutionary novelty in a rat with no molars. Biology Letters, 8, 990–993.
Fejfar, O. (1974). Die Eomyiden und Cricetiden (Rodentia, Mammalia) des Miozäns der Tschechoslowakei. Palaeontographica, A146, 100–180.
Fisher Von Waldheim, G. (1814). Zoognosia Tabulis synopticis illustrata (Vol. 3). Moscow: Nicolai S. Vsevolozsky.
Fischer Von Waldheim, G. (1817). Adversaria Zoologica. Mémoires de la Societé Impériale des Naturalistes de Moscou, 5, 357–446.
Forsyth Major, C. J. (1893). On some Miocene squirrels, with remarks on the dentition and classification of the Sciurinae. Proceedings of the Zoological Society of London, 1893, 179–215.
Freudenthal, M. (1963). Entwicklungsstufen der miozänen Cricetodontinae (Mammalia, Rodentia) Mittelspaniens und ihre stratigraphische Bedeutung (pp. 1–107). Wageningen: Ponsen & Looijen.
Freudenthal, M. (1967). On the mammalian fauna of the Hipparion-beds in the Calatayud-Teruel Basin (prov. Zaragoza, Spain). Part 3: Democricetodon and Rotundomys (Rodentia). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B, 70(3), 298–315.
Freudenthal, M., & Daams, R. (1988). Democricetodon, Fahlbuschia, Pseudofahlbuschia nov. gen., and Renzimys from the Aragonian and the lower Vallesian of the Calatayud-Teruel Basin. In M. Freudenthal (Ed.), Biostratigraphy and paleoecology of the Neogene micromammalian faunas from the Calatayud-Teruel Basin (Spain). Scripta Geologica Special Issue 1, 133–252.
Furió, M., Ruiz-Sánchez, F. J., Crespo, V. D., Freudenthal, M., & Montoya, P. (2012). The southernmost Miocene occurrence of the last European herpetotheriid Amphiperatherium frequens (Metatheria, Mammalia). Comptes Rendus Palevol, 11(5), 371–377.
García-Paredes, I. (2006). Patrones evolutivos de los Gliridae (Rodentia, Mammalia) del Mioceno inferior y medio del área tipo del Aragoniense (Cuenca de Calatayud-Montalbán). Unpublished PhD thesis. Madrid: Universidad Complutense de Madrid.
García-Paredes, I., Peláez-Campomanes, P., & Álvarez-Sierra, M. Á. (2009). Gliridae (Rodentia, Mammalia) with simple dental pattern: a new genus and new species from the European Lower and middle Miocene. Zoological Journal of the Linnean Society, 157, 622–652.
García-Paredes, I., Peláez-Campomanes, P., & Álvarez-Sierra, M. Á. (2010). Microdyromys remmerti, sp. nov., a new Gliridae (Rodentia, Mammalia) from the Aragonian type area (Miocene, Calatayud–Montalbán Basin, Spain). Journal of Vertebrate Paleontology, 30(5), 1594–1609.
Gibert, J. (1975). New insectivores from the Miocene of Spain I–II. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen B, 78(2), 108–132.
Gill, T. N. (1872). Arrangement of the families of mammals, with analytical tables. Prepared for the Smithsonian Institution. Smithsonian Miscellaneous Collections, 230, 1–198.
Ginsburg, L., & Mein, P. (2012). Les Sciuridae (Rodentia) de Sansan. Mémoires du Muséum National d’Histoire Naturelle, 203, 81–94.
Gray, J. E. (1821). On the natural arrangement of vertebrose animals. London Medical Repository, 15, 296–310.
Gureev, A. A. (1960). Zajtseobraznye (Lagomorpha) Oligotsena Mongolii i Kazakhstana. Trudy Paleontologičeskogo Instituta, 77, 5–34.
Hensel, R. (1856). Beiträge zur Kenntniss fossiler Säugetiere. Zeitschrift der Deutschen Geologischen Gesellschaft, 8, 660–704.
Hoek Ostende, L. W. van den (1997). Insectivore faunas from the Lower Miocene of Anatolia. Part 4: The genus Desmanodon (Talpidae) with the description of a new species from the Lower Miocene of Spain. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 100(1/2), 27–65.
Hoek Ostende, L. W. van den (2001). Insectivore faunas from the Lower Miocene of Anatolia. Part 5: Talpidae. Scripta Geologica, 122, 1–45.
Hoek Ostende, L. W. van den (2003). Insectivores (Erinaceomorpha, Soricomorpha, Mammalia) from the Ramblian of the Daroca-Calamochaarea. In N. López-Martínez, P. Peláez-Campomanes, & M. Hernández Fernández (Eds.), En torno a fósiles de mamíferos: datacíon, evolucióny paleoambiente. Coloquios de Paleontología Volume extraordinaire 1, 281–310.
Hoek Ostende, L. W. van den, & Doukas, C. D. (2003). Distribution of the early Miocene erinaceid Galerix symeonidisi Doukas, 1986. In J. W. F. Reumer, & W. Wessels (Eds.), Distribution and migration of tertiary mammals in Eurasia. Deinsea 10, 287–303.
Hoek Ostende, L. W. van den, & Furió, M. (2005). Spain. In L. W. van den Hoek Ostende, C. S. Doukas & J. W. F. Reumer (Eds.), The fossil record of the Eurasian Neogene insectivores (Erinaceomorpha, Soricomorpha, Mammalia), (Part I. pp. 149–284) Nationaal Natuurhistorisch Museum Naturalis, Leiden.
Hoffmann, R. S., & Smith, A. T. (2005). Order Lagomorpha. In D. E. Wilson & D. M. Reeder (Eds.), Mammal species of the world: A taxonomic and geographic reference (pp. 185–211). Baltimore: The Johns Hopkins University Press.
Hordijk, K. (2010). Perseverance of pikas in the Miocene. Interplay of climate and competition in the evolution of Spanish Ochotonidae (Lagopmorpha, Mammalia). Geologica Ultraiectina, 333, 1–232.
Hordijk, K., & Meulen, A. J. van der (2010a). Systematics of resident species of Lagopsis and Prolagus (Ochotonidae, Lagomorpha, Mammalia) from the late early and middle Miocene of northeastern Central Spain. Geologica Ultraiectina, 333, 17–85.
Hordijk, K., & Meulen, A. J. van der (2010b). Systematics of immigrant species of Prolagus (Ochotonidae, Lagomorpha, Mammalia) from the late early and middle Miocene of north-eastern Central Spain. Geologica Ultraiectina, 333, 87–141.
Illiger, C. (1811). Prodromus systematis mammalium et avium; additis terminis zoographicis utriusque classis, eorumque versione germanic. Berlin: C. Salfeld.
Klein Hofmeijer, G., & Bruijn, H. de (1988). The mammals from the Lower Miocene of Aliveri (Island of Evia, Greece). Part 8. The Cricetidae. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen Series B, 91(2), 185–204.
Klietmann, J., Nagel, D., Rummel, M., & Hoek Ostende, L. W. van den (2014a). Amphiperatherium and Erinaceidae of Petersbuch 28. Bulletin of Geosciences, 89(1), 1–20.
Klietmann, J., Nagel, D., Rummel, M., & Hoek Ostende, L. W. van den (2014b). Heterosorex and Soricidae (Eulipotyphla, Mammalia) of the fissure Petersbuch 28; micro-evolution as indicator of temporal mixing? Comptes Rendus Palevol, 13, 157–181.
Krijgsman, W., Garcés, M., Langereis, C. G., Daams, R., Dam, J. van, Meulen, A.J. van der, Agustí, J., & Cabrera, L. (1996). A new chronology for the middle to late Miocene continental record in Spain. Earth and Planetary Science Letters, 142, 367–380.
Krijgsman, W., Langereis, C. G., Daams, R., & Meulen, A. J. van der (1994). Magnetostratigraphic dating of the middle Miocene climate change in the continental deposits of the Aragonian type area in the Calatayud-Teruel Basin (Central Spain). Earth and Planetary Science Letters, 128, 513–526.
Lartet, E. (1851). Notice sur la colline de Sansan. Annuaire du Département du Gers 1851, 1–47. Auch, France.
López-Martínez, N. (1974). Evolution de la lignée Piezodus-Prolagus (Lagomorpha, Ochotonidae) dans le Cénozoïque d’Europe Sud-Occidentale. Montpellier: Université des Sciences et Techniques du Languedoc, Académie de Montpellier.
López-Martínez, N. (1977). Nuevos lagomorfos (Mammalia) del Neogeno y Cuaternario Español. Trabajos sobre Neógeno-Cuaternario, 8, 7–45.
López-Martínez, N. (1984). Los Lagomorfos (Mammalia) de la sucesión del Mioceno inferior de Calamocha (prov. de Teruel). Coloquios de Paleontología, 39, 27–44.
López-Martínez, N. (1988). Contribution à l’étude du gisement Miocène supérieur de Montredon (Hérault). Les grands mammifères. Les lagomorphes. Palaeovertebrata, Mémoire Extraordinaire, 1988, 3–14.
López-Martínez, N. (1989). Revision sistematica y biostratigrafica de los Lagomorpha (Mammalia) de Terciario y Cuaternario de España. Memorias del Museo Paleontologico de la Universidad de Zaragoza, 3(3), 1–350.
López-Martínez, N. (2001). Paleobiogeographical history of Prolagus, an European ochotonid (Lagomorpha). Lynx, 32, 215–231.
López-Martínez, N., Sesé-Benito, C., & Sanz-García, J. L. (1977). La microfauna (Rodentia, Insectívora, Lagomorpha y Reptilia) de las fisuras del Mioceno medio de Escobosa de Calatañazor (Soria, España). Acta Geologica Hispánica, 12, 60–68.
López-Martínez, N., Sesé Benito, C., & Herraez, E. (1987). Los yacimientos de micromamíferos del área de Madrid. Boletín Geológico y Minero, 98(2), 159–176.
Lourens, L., Hilgen, F., Shackleton, N. J., Laskar, J., & Wilson, D. (2005). The Neogene period. In F. M. Gradstein, J. G. Ogg, & A. G. Smith (Eds.), A geologic time scale 2004 (pp. 409–440). Cambridge: Cambridge University Press.
Mayr, H. (1979). Gebißmorphologische Untersuchungen an miozänen Gliriden (Mammalia, Rodentia) Süddeutschlands. Inaugural-Dissertation (pp. 1–380). Munich: Fakultät für Geowissenschaften der Ludwig-Maximilians-Universität zu München.
Mein, P. (1984). Composition quantitative des faunes de Mammifères du Miocène moyen et supérieur de la région lyonnaise. Paléobiologie Continentale, 14(2), 339–346.
Meulen, A. J. van der, & Bruijn, H. de (1982). The mammals from the lower Miocene of Aliveri (Island of Evia, Greece). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, 85(4), 485–524.
Meulen, A. J. van der, & Daams, R. (1992). Evolution of early-middle Miocene rodent faunas in relation to long-term palaeoenvironmental changes. Palaeogeography, Palaeoclimatology, Palaeoecology, 93, 227–253.
Meulen, A. J. van der, Peláez-Campomanes, P., & Daams, R. (2003). Revision of medium-sized Cricetidae from the Miocene of the Daroca-Villafeliche area in the Calatayud-Teruel basin (Zaragoza, Spain). Coloquios de Paleontología, 1, 385–441.
Meulen, A. J. van der, Peláez-Campomanes, P., & Levin, S. A. (2005). Age structure, residents, and transients of Miocene rodent communities. The American Naturalist, 165(4), E108–E125.
Meulen, A. J. van der, García-Parédes, I., Álvarez-Sierra, M. A., Hoek Ostende, L. W. van den, Hordijk, K., Oliver, A., & Peláez-Campomanes, P. (2012). Updated Aragonian biostratigraphy: small mammal distribution and its implication for the Miocene European Chronology. Geologica Acta, 10(2), 159–179.
Muirhead, L. (1819). Mazology [sic], pp. 393–486 [pls. 353–358] In D. Brewster (Ed.), The Edinburgh Encyclopaedia. Fourth ed. 13, 1–744, pls. 347–371, 1830. Edinburgh: William Blackwood.
Musser, G. G. (1990). Sulawesi rodents: species traits and chromosomes of Haeromys minahassae and Echiothrix leucura (Muridae, Murinae). American Museum Novitates, 2989, 1–18.
Peláez-Campomanes, P. (2001). Revision of the Aragonian (Miocene) Atlantoxerus (Sciuridae). Journal of Paleontology, 75(2), 418–426.
Robles, F., Belinchón, M., García-Flor, J., & Morales, J. (1991). El Neógeno continental de Buñol y del valle del río Cabriel. In M. De Renzi, A. Márquez-Aliaga, & J. Usera (Eds.), El estudio de la Forma Orgánica y sus consecuencias en Paleontología Sistemática, Paleoecología y Paleontología Evolutiva. Revista Española de Paleontología. Volumen Extraordinario, 205–215.
Pomel, A. N. (1848). Etudes sur les carnassiers insectivores (extrait). Seconde partie, Classification des insectivores. Archives des sciences physiques et naturelles Genève, 9, 244–251.
Rössner, G. E., & Heissig, K. (Eds.). (1999). The Miocene Land Mammals of Europe. München: Verlag Dr. Friedrich Pfeil.
Royo Gómez, J. (1928). Memoria explicativa del Mapa Geológico, Hoja 560 de Alcalá de Henares. Madrid: Instituto Geológico y Minero de España.
Schaub, S. (1920). Melissiodon n. gen., ein bisher übersehener oligocäner Muride. Senckenbergiana, 2, 43–47.
Schaub, S., & Zapfe, H. (1953). Die Fauna der miozänen Spaltenfüllung von Neudorf an der March (CSR) Simplicidentata. Sitzungsberichte Österreichische Akademie der Wissenschaften, Mathematisch-naturwissenschafliche Klasse, 162(3), 181–215.
Schlosser, M. (1887): Die Affen, Lemuren, Chiropteren, Insectivoren, Marsupialier, Creodonten und Carnivoren des Europäischen Tertiärs und deren Beziehungen zu ihren lebenden und fossilen außereuropäischen Verwandten. I. Beiträge zur Paläontologie Österreich-Ungarns und des Orients, 6(1,2), 1–244.
Thomas, O. (1897). On the genera of rodents: an attempt to bring up to date the current arrangement of the order. Proceedings of the Zoological Society of London, 1896, 1012–1028.
Trouessart, E. L. (1879). Catalogue des mammifères vivants et fossiles. Insectivores. Revue et Magasin de Zoologie Pure et Appliquée, Paris, 3(7), 219–285.
Tütken, T., & Vennemann, T. (2009). Stable isotope ecology of Miocene large mammals from Sandelzhausen, southern Germany. Paläontologische Zeitschrift, 83(1), 207–226.
Ünay, E., Bruijn, H. de, & Saraç, G. (2003). A preliminary zonation of the continental Neogene of Anatolia based on rodents. In J. W. F. Reumer, & W. Wessels (Eds). Distribution and migration of tertiary mammals in Eurasia. Deinsea 10, 539–547.
Viret, J., & Zapfe, H. (1951). Sur quelques soricidés miocènes. Eclogae Geologicae Helvetiae, 44, 411–426.
Waddell, P. J., Okada, N., & Hasegawa, M. (1999). Towards resolving the interordinal relationships of placental mammals. Systematic Biology, 48, 1–5.
Weerd, A. van de (1976). Rodent faunas of the Mio-Pliocene continental sediments of the Teruel-Alfambra region, Spain. Utrecht Micropaleontological Bulletins Special Publication, 2, 1–217.
Winge, H. (1887). Jordfundne og nulevende Gnavere (Rodentia) fra Lagoa Santa, Minas Geraes, Brasilien. Med Udsigt over Gnavernes indbyrdes Slaegtskab. Volume 1, Part 3. Copenhagen: Museo Lundii.
Ziegler, R. (1989). Heterosoricidae und Soricidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Naturkunde B, 154, 1–73.
Ziegler, R. (1998). Marsupialia und Insectivora (Mammalia) aus den oberoligozänen Spaltenfüllungen Harrlingen 8 und Herrlingen 9 bei Ulm (Baden-Württemberg). Senckenbergiana lethaea, 77(1/2), 101–143.
Ziegler, R. (2009). Soricids (Soricidae, Mammalia) from early Oligocene fissure fillings in South Germany, with remarks to the phylogeny of Heterosoricinae. Palaeodiversity, 2, 321–342.
Ziegler, R., & Fahlbusch, V. (1986). Kleinsäuger-Faunen aus der basalen Oberen Süsswasser-Molasse Niederbayerns. Abhandlungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie, Zitteliana, 14, 3–80.
Acknowledgements
This paper is in honour of Albert van der Meulen, friend, colleague, supervisor, tutor and mentor and main participant of this research project. Tilly Bouten and the late Tom van Hinte assisted with the SEM and figures. The constructive comments of guest editor Pablo Peláez-Campomanes and the reviewers, Israel García-Paredes and Jérôme Prieto, are highly appreciated.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is a contribution to the special issue “Old worlds, new ideas. A tribute to Albert van der Meulen”.
Appendix 1
Appendix 1
Material and measurements of all the Montalvos2 small mammal cheek teeth elements, including collection numbers
Element | Length (mm) | Width (mm) | Collection number | ||||||
|---|---|---|---|---|---|---|---|---|---|
Mean | Range | SD | N | Mean | Range | SD | Locality code | Number | |
Melissiodon dominans | |||||||||
m1 | 2.68 | 1 | 1.94 | MNT2 | 356–357 | ||||
m2 | 2.50 | 1 | 2.09 | MNT2 | 358–360 | ||||
m3 | 2.37 | 2.29–2.44 | 2/1 | 1.67 | MNT2 | 361–362 | |||
M1 | 3.49 | 3.37–3.56 | 3 | 2.27 | 2.25–2.28 | MNT2 | 344–348, 350 | ||
M2 | 2.35 | 2.33–2.37 | 2 | 1.99 | 1.98–2.00 | MNT2 | 349, 351 | ||
M3 | 1.78 | 1.68–1.86 | 2/1 | 2.11 | MNT2 | 352–354 | |||
Democricetodon decipiens | |||||||||
m1 | 1.50 | 1.31–1.67 | 0.099 | 12 | 1.06 | 0.96–1.16 | 0.061 | MNT2 | 126–139 |
m2 | 1.37 | 1.31–1.40 | 0.035 | 8 | 1.12 | 1.01–1.17 | 0.057 | MNT2 | 140–148 |
m3 | 1.18 | 1.13–1.23 | 0.073 | 2 | 0.96 | 0.87–1.04 | 0.118 | MNT2 | 149–150 |
M1 | 1.76 | 1.68–1.80 | 0.050 | 12 | 1.20 | 1.15–1.25 | 0.036 | MNT2 | 101–107 |
M2 | 1.25 | 1.20–1.30 | 0.037 | 9 | 1.17 | 1.13–1.26 | 0.042 | MNT2 | 108–116 |
M3 | 0.91 | 0.87–0.97 | 0.041 | 5 | 1.00 | 0.96–1.06 | 0.037 | MNT2 | 118–123 |
Megacricetodon primitivus | |||||||||
m1 | 1.30 | 1.19–1.37 | 0.052 | 22 | 0.87 | 0.77–0.93 | 0.055 | MNT2 | 241–270 |
m2 | 1.37 | 1.31–1.40 | 0.035 | 33 | 1.12 | 1.01–1.17 | 0.057 | MNT2 | 271–288, 290–304 |
m3 | 1.07 | 0.97–1.15 | 0.046 | 6 | 0.90 | 0.77–0.99 | 0.055 | MNT2 | 306–310 |
M1 | 1.47 | 1.37–1.60 | 0.055 | 26 | 0.97 | 0.90–1.03 | 0.032 | MNT2 | 151–186 |
M2 | 1.01 | 0.91–1.09 | 0.040 | 41 | 0.93 | 0.82–1.02 | 0.047 | MNT2 | 187–209, 211–231, 289 |
M3 | 0.88 | 0.85–0.92 | 0.038 | 5 | 0.77 | 0.74–0.82 | 0.036 | MNT2 | 233–237 |
Eumyarion weinfurteri | |||||||||
m1 | 1.73 | 2 | 1.13 | 1.13-1.13 | 0.001 | MNT2 | 331–334 | ||
m2 | 1.48 | 1.47–1.49 | 0.008 | 4 | 1.28 | 1.24–1.31 | 0.034 | MNT2 | 335–340 |
m3 | 1.45 | 1.37–1.54 | 0.120 | 3 | 1.14 | 1.13–1.16 | 0.015 | MNT2 | 341–343 |
M1 | 1.96 | 1.84–2.03 | 0.072 | 7 | 1.34 | 1.21–1.40 | 0.069 | MNT2 | 311–319 |
M2 | 1.39 | 1.33–1.46 | 0.043 | 5 | 1.4 | 1.34–1.48 | 0.056 | MNT2 | 320–327 |
M3 | 1.10 | 1.02–1.16 | 0.073 | 3 | 1.18 | 1.13–1.23 | 0.051 | MNT2 | 328–330 |
Length (mm) | Width (mm) | Collection number | |||||||
|---|---|---|---|---|---|---|---|---|---|
Element | Mean | Range | SD | N | Mean | Range | SD | Locality code | Number |
Pseudodryomys aff. ibericus | |||||||||
p4 | 0.82 | 0.80–0.92 | 4 | 0.89 | 0.69–1.06 | MNT2 | 451–452, 464, 515 | ||
m1 | 1.16 | 1.07–1.32 | 0.070 | 11/12 | 1.06 | 0.98–1.20 | 0.059 | MNT2 | 461–462, 484–493 |
m2 | 1.13 | 1.01–1.28 | 12 | 1.07 | 0.99–1.30 | MNT2 | 463, 494–506 | ||
m3 | 0.97 | 0.77–1.07 | 0.108 | 7/8 | 0.93 | 0.87–1.00 | 0.050 | MNT2 | 508–513, 521 |
P4 | 0.92 | 0.88–0.96 | 2 | 1.05 | 1.04–1.07 | MNT2 | 466–467 | ||
M1-2 | 1.12 | 1.00–1.25 | 1/11 | 1.24 | 1.13–1.34 | 0.022 | MNT2 | 454–458, 471–479 | |
M3 | 0.89 | 0.74–0.98 | 3 | 1.10 | 0.96–1.19 | MNT2 | 481–482, 522 | ||
Simplomys simplicidens | |||||||||
m1 | 1.35 | 1.22–1.47 | 5/4 | 1.30 | 1.17–1.22 | MNT2 | 415–420 | ||
m2 | 1.39 | 1.33–1.43 | 3/4 | 1.40 | 1.40–1.56 | MNT2 | 421–432 | ||
m3 | 1.08 | 0.95–1.08 | 0.040 | 13 | 1.17 | 1.06–1.28 | 0.076 | MNT2 | 434–448 |
D4 | 0.77 | 0.75–0.78 | 2 | 0.90 | 0.89–0.91 | MNT2 | 652, 653 | ||
P4 | 0.82 | 0.72–0.92 | 0.027 | 10 | 0.97 | 0.83–1.12 | 0.065 | MNT2 | 641–651 |
M1-2 | 1.22 | 1.12–1.43 | 0.083 | 16/22 | 1.49 | 1.36–1.57 | 0.066 | MNT2 | 371–391, 93–398 |
M3 | 0.83 | 0.72–0.91 | 0.057 | 12 | 1.21 | 1.11–1.32 | 0.067 | MNT2 | 399–413 |
Simplomys sp. | |||||||||
Length | Width | Collection number | |||||||
M1-2 | 1.27 | 1.77 | MNT2 | 468 | |||||
Simplomys julii | |||||||||
M1-2 | 1.05 | 1.27 | MNT2 | 392 | |||||
M1-2 | 1.08 | 1.21 | MNT2 | 470 | |||||
Microdyromys legidensis | |||||||||
m1 | 0.93 | 0.87–0.96 | 5 | 0.89 | 0.77–0.97 | MNT2 | 543–548 | ||
m2 | 1.08 | 1 | 1.04 | MNT2 | 549–550 | ||||
P4 | 0.72 | 1 | 0.70 | MNT2 | 625 | ||||
M1-2 | 0.91 | 0.84–1.00 | 10 | 1.01 | 0.94–1.00 | MNT2 | 531–541 | ||
Glirudinus gracilis | |||||||||
Length | Width | Collection number | |||||||
m2 | 1.24 | 1.22 | MNT2 | 525 | |||||
m2 | 1.21 | 1.19 | MNT2 | 527 | |||||
m2 | fragm. | fragm. | MNT2 | 528 | |||||
m3 | 1.03 | 0.94 | MNT2 | 530 | |||||
m3 | 1.01 | 1.03 | MNT2 | 529 | |||||
m3 | fragm. | fragm. | MNT2 | 526 | |||||
M3 | 0.95 | fragm. | MNT2 | 524 | |||||
Armantomys aragonensis | |||||||||
P4 | 1.37 | 2.03 | MNT2 | 556 | |||||
M2 | 1.27 | 1.99 | MNT2 | 553 | |||||
M3 | 1.26 | 1.86 | MNT2 | 554 | |||||
M3 | 1.12 | 1.81 | MNT2 | 555 | |||||
10 fragments | MNT2 | 551–552, 558–567 | |||||||
Element | Length (mm) | Width (mm) | Collection number | ||||||
|---|---|---|---|---|---|---|---|---|---|
Mean | Range | SD | N | Mean | Range | SD | Locality code | Number | |
Ligerimys ellipticus | |||||||||
D4 | 1.22 | 1.15–1.29 | 2 | 0.8 | 0.80–0.80 | MNT2 | 595–596 | ||
P4 | 0.98 | 0.96–1.00 | 2 | 0.93 | 0.92–0.94 | MNT2 | 597–598 | ||
M1-2 | 1.14 | 1.02–1.25 | 0.069 | 8/9 | 1.04 | 0.93–1.13 | 0.060 | MNT2 | 581–591 |
M3 | 1.02 | 0.97–1.07 | 3 | 0.89 | 0.84–0.93 | MNT2 | 593 | ||
M1-2 | 1.11 | 1.03–1.17 | 0.048 | 7 | 1.19 | 1.14–1.28 | 0.053 | MNT2 | 571–579 |
M3 | 0.84 | 1 | 1.08 | MNT2 | 580 | ||||
Heteroxerus rubricati | |||||||||
D4 | 1.23 | 1 | 0.94 | MNT2 | 614–615 | ||||
P4 | 0 | MNT2 | |||||||
M1-2 | 1.49 | 1.45–1.53 | 2 | 1.52 | 1.49–1.55 | MNT2 | 616–617 | ||
m3 | 1.57 | 1 | 1.43 | MNT2 | 618–620 | ||||
D4 | 1.23 | 1.19–1.27 | 2 | 1.25 | 1.17–1.33 | MNT2 | 611–612 | ||
P4 | 1.23 | 1.20–1.25 | 3/2 | 1.55 | 1.52–1.57 | MNT2 | 603–604, 609 | ||
M1-2 | 1.41 | 1.40–1.42 | 3/5 | 1.63 | 1.60–1.68 | MNT2 | 601–602, 605–608, 610 | ||
M3 | 1.45 | 1 | 1.52 | MNT2 | 621 | ||||
Heteroxerus cf. grivensis | |||||||||
Mean length | Mean width | Collection number | |||||||
m2 | 1.90 | 1.91 | MNT2 | 625 | |||||
Albanensia cf. sansaniensis | |||||||||
m3 | 3.20 | 3.08 | MNT2 | 623 | |||||
Element | Length (mm) | Width (mm) | Collection number | ||||||
|---|---|---|---|---|---|---|---|---|---|
Mean | Range | SD | N | Mean | Range | SD | Locality code | Number | |
Lagopsis penai | |||||||||
d3 | 1.61 | 1/0 | MNT2 | 921A–924A, 926A–928A | |||||
p3 | 1.56 | 1.25–1.74 | 0.13 | 21/19 | 1.65 | 1.44–1.86 | 0.12 | MNT2 | 801A–810A, 801B–809B |
MNT2 | 811A–820A, 811B820B | ||||||||
MNT2 | 821A–827A, 911B–913B | ||||||||
d4 | 1.20 | 1.14–1.28 | 3/1 | 1.54 | MNT2 | 926B–928B | |||
D2 | 1.04 | 0.96–1.10 | 0.05 | 6/1 | 1.81 | MNT2 | 931A–939A | ||
P2 | 0.80 | 0.69–0.99 | 0.07 | 35/29 | 1.53 | 1.29–1.78 | 0.13 | MNT2 | 876B–880B, 881A–890A |
MNT2 | 881B–890B, 891A–900A | ||||||||
MNT2 | 891B–892B, 896B–900B | ||||||||
MNT2 | 901A–910A, 901B–904B | ||||||||
MNT2 | 909B–910B | ||||||||
P3 | 1.24 | 1.07–1.39 | 0.07 | 43/9 | 2.46 | 2.32–2.64 | 0.11 | MNT2 | 831B–840B, 841A–850A, |
MNT2 | 841B–850B, 851A–860A, | ||||||||
MNT2 | 851B–860B, 861A–868A, 915B | ||||||||
Prolagus lopezmartinezae | |||||||||
ᅟ | ᅟ | ᅟ | ᅟ | ᅟ | ᅟ | ᅟ | ᅟ | ᅟ | |
d3 | 1.63 | 1 | 1.27 | MNT2 | 921B–923B | ||||
p3 | 1.47 | 1.47–1.48 | 2/9 | 1.41 | 1.21–1.59 | 0.12 | MNT2 | 821B–829B, 831A–838A, 917B | |
D2 | 1.01 | 0 | MNT2 | 940A | |||||
P2 | 0.78 | 0.69–0.88 | 0.07 | 5 | 1.53 | 1.40–1.61 | 0.08 | MNT2 | 911A–919A |
P3 | 1.35 | 1.31–1.42 | 0.04 | 8/2 | 2.24 | 2.22–2.25 | MNT2 | 861B–870B, 871A–880A, | |
MNT2 | 871B–872B, 918B, 919B | ||||||||
Species | Element | Length anteroconid (mm) | Width anteroconid (mm) | ||||||
|---|---|---|---|---|---|---|---|---|---|
N | Mean | Range | SD | N | Mean | Range | SD | ||
L. penai | p3 | 28 | 0.50 | 0.35–0.62 | 0.07 | 23 | 0.89 | 0.64–1.19 | 0.15 |
P. lopezmartinezae | p3 | 3 | 0.41 | 0.37–0.44 | 3 | 0.55 | 0.46–0.61 | ||
L. penai | d3 | 3 | 0.66 | 0.64–0.71 | 2 | 0.91 | 0.90–0.92 | ||
P. lopezmartinezae | d3 | 2 | 0.60 | 0.58–0.62 | 1 | 0.69 | |||
Length metaconid-entoconid (mm) | Width protoconid-metaconid (mm) | ||||||||
L. penai | p3 | 22 | 1.10 | 1.01–1.18 | 0.05 | 28 | 1.19 | 0.89–1.41 | 0.12 |
P. lopezmartinezae | p3 | 10 | 0.95 | 0.85–1.03 | 0.06 | 14 | 0.95 | 0.69–1.22 | 0.13 |
Length protoconulid-entoconid (mm) | |||||||||
P. lopezmartinezae | p3 | 10 | 0.97 | 0.80–1.10 | 0.095 | ||||
Width distal hyperloph (mm) | Length postlobe (mm) | ||||||||
L. penai | P3 | 19 | 1.74 | 1.37–2.06 | 0.22 | 27 | 0.77 | 0.62–0.95 | 0.10 |
P. lopezmartinezae | P3 | 5 | 1.60 | 1.34–1.86 | 0.25 | 11 | 0.89 | 0.75–1.00 | 0.09 |
L. penai | P2 | 33 | 0.43 | 0.28–0.67 | 0.08 | 32 | 0.68 | 0.50–0.82 | 0.08 |
P. lopezmartinezae | P2 | 5 | 0.61 | 0.52–0.68 | 0.07 | 5 | 0.63 | 0.55–0.68 | 0.05 |
L. penai | D2 | 4 | 1.21 | 0.93–1.34 | 4 | 0.74 | 0.65–0.84 | ||
P. lopezmartinezae | D2 | 1 | 0.93 | 1 | 0.74 | ||||
Width mesial hyperloph (mm) | Width mesial hyperloph/width | ||||||||
L. penai | P3 | 26 | 1.52 | 1.19–1.80 | 0.18 | 5 | 0.61 | 0.52–0.67 | 0.05 |
P. lopezmartinezae | P3 | 6 | 1.35 | 1.10–1.66 | 0.25 | 2 | 0.67 | 0.60 | |
p3 | Anteroconid | Anteroflexid | Protoconulid | Centroflexid | ||||||||||
Species | Ntotal (excl. juveniles) | (Sub)round | Ellipsoid | Flattened/broad/triangular | Diamond shaped | Distinct with cement | Distinct inflexion | Absent/weak/incipient inflexion | Absent | Incipient/small | Developed | Absent | Incipient/superficial | Deep |
L. penai | 43 | 0 | 0 | 28 | 5 | 2 | 8 | 23 | 0 | 0 | 0 | 43 | 0 | 0 |
P. lopezmartinezae | 16 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 2 | 13 | 0 | 0 | 15 | 0 |
p3 | Crochet | Connections | Relative Size | |||||||||||
Species | Ntotal (excl. juveniles) | Absent | Incipient/small | Developed | Metaconid isolated | Anterolophid | Metalophid | Protoisthmid present | Protoisthmid absent | Metaisthmid | Anteroisthmid | Anteroconid < metaconid | Anteroconid = metaconid | Anteroconid > metaconid |
L. penai | 43 | 41 | 0 | 0 | 0 | 43 | 43 | 43 | 0 | 0 | 0 | 0 | 2 | 41 |
P. lopezmartinezae | 16 | 7 | 5 | 2 | 0 | 0 | 16 | 16 | 0 | 0 | 0 | 1 | 1 | 1 |
P3 | Mesoflexus | Precone | Lagilophe | P2 | Hypoflexus | |||||||||
Species | Ntotal (excl. juveniles) | absent | incipient | developed | indistinct (mhl = pr) | distinct (precone thicker than mhl) | long | short | absent (only lagicone) | Ntotal (excl. juveniles) | absent | incipient/superficial | distinct | with cement |
L. penai | 57 | 7 | 16 | 7 | 40 | 5 | 0 | 1 | 56 | 52 | 32 | 9 | 1 | 2 |
P. lopezmartinezae | 19 | 0 | 0 | 17 | 1 | 9 | 1 | 8 | 8 | 6 | 5 | 0 | 0 | 0 |
Element | Length (mm) | Width 1 (mm) | Width 2 (mm) | Collection number | |
|---|---|---|---|---|---|
Amphiperatherium sp. | |||||
1 labial fragment of an upper molar | MNT2 | 1058 | |||
Galerix symeonidisi | |||||
p4 | 1.76 | 1.18 | MNT2 | 1001 | |
m1 | fragm. | fragm. | MNT2 | 1003 | |
m2 | fragm. | fragm. | MNT2 | 1004 | |
m3 | 1.67 | 1.10 | 0.93 | MNT2 | 1005 |
P3 | 1.88 | 1.57 | MNT2 | 1007 | |
P4 | fragm. | fragm. | MNT2 | 1008 | |
M1 | fragm. | fragm. | MNT2 | 1009 | |
M1 | fragm. | fragm. | MNT2 | 1010 | |
M2 | 1.88 | 2.46 | MNT2 | 1011 | |
M2 | fragm. | fragm. | MNT2 | 1012 | |
M3 | 1.17 | 1.75 | MNT2 | 1014 | |
M3 | 1.02 | 1.47 | MNT2 | 1015 | |
M3 | 1.07 | 1.00 | MNT2 | 1016 | |
cf. Desmanella sp. | |||||
p4 | 0.93 | 0.64 | MNT2 | 1021 | |
m3 | 1.12 | 0.74 | 0.64 | MNT2 | 1023 |
m3 | 1.05 | 0.78 | 0.64 | MNT2 | 1024 |
P4 | 1.28 | 1.10 | MNT2 | 1026 | |
Element | Length (mm) | Width 1 (mm) | Width 2 (mm) | Collection number | |
|---|---|---|---|---|---|
Desmanodon sp. | |||||
p4 | 1.21 | 0.78 | MNT2 | 1028 | |
m1 | 1.86 | 1.15 | 1.29 | MNT2 | 1029 |
m1 | 2.04 | 1.18 | 1.34 | MNT2 | 1030 |
m2 | 2.18 | 1.32 | 1.31 | MNT2 | 1031 |
m3 | 1.63 | 1.10 | 0.96 | MNT2 | 1033 |
M1 | fragm. | fragm. | MNT2 | 1036 | |
M1 | 2.54 | 1.57 | MNT2 | 1035 | |
M2 | 1.89 | 2.26 | MNT2 | 1037 | |
M2 | fragm. | fragm. | MNT2 | 1038 | |
M3 | 1.17 | 1.68 | MNT2 | 1041 | |
M3 | 1.06 | 1.59 | MNT2 | 1043 | |
M3 | fragm. | fragm. | MNT2 | 1044 | |
M3 | fragm. | fragm. | MNT2 | 1042 | |
Heterosorex cf. neumayrianus | |||||
m1 | fragm. | fragm. | MNT2 | 1046 | |
m2 | fragm. | fragm. | MNT2 | 1047 | |
m3 | 1.39 | 0.95 | 0.91 | MNT2 | 1048 |
P4 | 1.60 | 1.72 | MNT2 | 1051 | |
M1 | 1.79 | 1.94 | MNT2 | 1053 | |
M1 | fragm. | fragm. | MNT2 | 1054 | |
Soricid gen. et sp. indet. | |||||
m1 | 1.15 | 0.70 | 0.80 | MNT2 | 1061 |
m1 | 1.20 | 0.74 | 0.80 | MNT2 | 1062 |
m2 | 1.21 | 0.74 | 0.74 | MNT2 | 1064 |
m3 | 1.00 | 0.57 | 0.46 | MNT2 | 1066 |
M2 | 1.13 | 1.41 | MNT2 | 1068 | |
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hordijk, K., Bosma, A., de Bruijn, H. et al. Biostratigraphical and palaeoecological implications of the small mammal assemblage from the late early Miocene of Montalvos2, Teruel Basin, Spain. Palaeobio Palaeoenv 95, 321–346 (2015). https://doi.org/10.1007/s12549-015-0203-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-015-0203-2