
Abstract
Miscanthus is a perennial C4 crop whose lignocellulose can be used as an alternative to the production of biosourced material. Miscanthus x giganteus (M. x giganteus) has demonstrated high maximum yields but also high yield variability across farmers’ fields. Miscanthus sinensis (M. sinensis) can be an alternative to M. x giganteus because it is considered to be more tolerant to water stress and to produce more stable yields. This study aimed to identify the main factors explaining the variability of yields across site-years for M. x giganteus and M. sinensis. A multi-local and multi-year trial network was set up in France (Ile de France and Center regions). Four treatments were established on seven sites, from spring 2013 to winter 2019: at each site, two treatments of M. x giganteus (a treatment from rhizome and a treatment from rhizome-derived plantlets) and two treatments of M. sinensis (a treatment from seed-derived plantlets established in single density and a treatment from seed-derived plantlets established in double density). We experienced 5 years of harvest because miscanthus was not harvested in 2014. First, we characterized yield variations across site-years for both genotypes. Second, we defined and calculated a set of indicators (e.g., water stress indicator, sum of degree-days of the previous year, number of frost days) that could affect miscanthus yields. Finally, we performed a mixed model with re-sampling to identify the main indicators that explained yield variability for each genotype specifically. Results showed that water stress and crop age mainly explained yield variability for both genotypes. M. sinensis yields were also affected by the sum of degree-days of the previous year of growth. Hence, genotype choice must take into account environmental characteristics. M. sinensis could indeed achieve higher and more stable yields than those of M. x giganteus in shallow sandy soils or in locations with a higher risk of low rainfall.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Miscanthus x giganteus (hereafter named M. x giganteus) is a perennial C4 grass with a growing period of 15–20 years and yields progressively increasing every year during the first years of growth [1]. Introduced into Europe from Japan in 1930, it is now cultivated across the continent [2]; it reaches a high potential biomass production, from 10 to 25 t ha−1 of dry matter, in experimental conditions [1]. M. x giganteus is therefore the only genotype now cultivated in European farmers’ fields [2]. However, using a single clone presents some disadvantages, such as loss of genetic diversity, or risks of pest and disease development in the future. To date, no miscanthus pest or disease control is required for miscanthus production in Europe [3].
Yield variability of M. x giganteus that takes sites and years into account together has reached 73%: Alexopoulou et al. [4] indeed obtained a yield variability with a cv amounting to 31% of M. x giganteus at a site in Italy over a period of 24 years, and Ouattara et al. [5] showed that yields of M. x giganteus were variable (with a coefficient of variation ranging from 64 to 73%) between 2013 and 2019 on six sites in France. Yield variability has been shown to be greater in clayey soils than in sandy and loamy soils (37% vs. 10% in 22 sites field experiments of M. x giganteus with growth years greater than or equal to 5 years) because M. x giganteus is noticeably slower to establish in clayey soils [6]. In addition to soil effect, climatic conditions have a significant effect on M. x giganteus [7], which is sensitive to low temperatures and water availability [1, 8].
Miscanthus sinensis (hereafter named M. sinensis) is more tolerant to water stress and frost [1, 7, 8], which allows it to be suited to a broader range of soil and climatic conditions. M. sinensis achieves lower yields than M. x giganteus but is less variable across different sites and years [5, 6]. It could therefore be an alternative to M. x giganteus on less favorable sites.
Several studies have been conducted on factors that could impact M. x giganteus yields [1, 9]. To our knowledge, M. sinensis has received little attention in the literature, while research is mostly focused on M. x giganteus because of its higher yield. Yet under stressed environmental conditions, M. sinensis could achieve higher yields [9]. Today, almost all commercial production of miscanthus is based on the M. x giganteus genotype alone [3, 10]. In addition, there are not many studies that have been conducted under field conditions to determine which genotype would be best suited to contrasting environmental conditions. Few studies have been carried out to identify some factors affecting the yields of M. x giganteus and, to a lesser extent, M. sinensis [11,12,13,14].
It is therefore necessary to specifically identify the factors that could explain the yield variability of each genotype across site-years. Factors explaining yield variability could differ between M. x giganteus and M. sinensis [7], as M. sinensis is more tolerant to water stress and to frost. Mineral nutrition could also explain this variability in yields of the two genotypes. It is known that miscanthus is a crop with low nitrogen requirements [10] but nutrients like P and K could be interesting to assess for explaining yield variability. This last aspect will not be studied in this paper.
An agronomic diagnosis [15,16,17,18] was used in this study to identify and assess the main limiting factors explaining the variability of M. sinensis and M. x giganteus yield under field conditions, and across several pedo-climatic conditions in France. This would in turn support the identification of potential actionable levers to overcome the limiting factors.
Material and Methods
A Multi-Environment Experimental Network
A multi-environment experimental network was set up on seven sites in France from 2013 to 2019: six sites in the Ile de France region and one site in the Center region. The trial network was characterized by different types of soils, with various textures, depths, and stoniness (Table 1). Trials were either located on agricultural lands, i.e., lands that were cultivated with annual crops before carrying out the experiment (Bioferme and La Bondue sites), or on marginal lands, i.e., lands that were previously maintained as set-aside land, mainly because they were located between several roads/highways and were therefore difficult to get to with agricultural equipment, such as the Marne et Gondoire site. The range of maximum soil water content (SWCmax) of these sites is wide, from 72 mm for La Bondue site to 317 mm for the Bioferme site (Table 1). Lastly, annual mean temperatures from March 25th to October 15th ranged from 14.7 to 18.4 °C across years.
The preceding crop before setting up the trials was a set-aside field at the Subdray site. Since 1992, La Bondue and Episy sites had been maintained as set-aside lands. The Marne et Gondoire and Evry sites had also been maintained as set-aside lands since 2003 and 2005, respectively. The preceding crop was winter barley (Hordeum vulgare) for the Bioferme and Chanteloup sites (Table 1).
We located the growth cycle of miscanthus between March 25 (regrowth or emergence in the first year) and October 15 (beginning of senescence). Climatic parameters were assessed over the period corresponding to the cycle before its senescence.
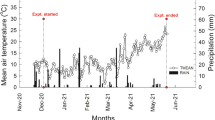
Climatic conditions followed the same trend throughout the trial network with little variation across sites, except for the rainfall recorded at Chanteloup (between 300 and 400 mm), which was lower than for the other sites in the Ile de France region and less variable across years compared to the other sites of the trial network (Fig. 1). In 2015 and 2018, rainfall was lower over the entire network, except for the Chanteloup and Marne et Gondoire sites in 2018. The climatic water balances (rainfall minus potential evapotranspiration) of these two years, i.e., 2015 and 2018, were also the lowest ones. Lastly, the Subdray sites recorded the highest climatic water deficits in 2015 (− 435 mm) and 2018 (− 582 mm).

Rainfall (mm) (a) and climatic water balance, i.e., rainfall minus potential evapotranspiration (mm) (b), covering the growth cycle of miscanthus (i.e., from March 1 to October 15) from 2013 to 2018. For the Bioferme, Episy, and La Bondue sites, we used the meteorological data of Nangis because these sites are close to one another. The weather data used originated from meteo France and from the French National Institute for Agriculture, Food, and Environment (INRAE) on-line platform CLIMATIK (https:// intranet.inra.fr/climatik_v2/ClimatikGwt.html)
Treatments
Each trial of the multi-environment network was composed of four strips, with an area of 141.4 m2 (27.2 m \(\times\) 5.2 m) for each strip (Fig. 2). Four treatments were established (one per strip): two treatments with M. x giganteus established from rhizomes (G_r-sd) or from plantlets (G_p-sd) and two treatments with M. sinensis K1399 (population variety whose seeds were provided by Wageningen University & Research) from plantlets in single (S_p-sd) and double (S_p-dd) density, i.e., 3.33 plants per m2 instead of 1.56 plants per m2. Seedlings were raised in greenhouses before transplanting [5]. To summarize, 4 treatments of M. x giganteus and M. sinensis were carried out under 7 sites × 5 years of harvest. However, the G_p-sd treatment was not successfully established at the Chanteloup site, which led to the loss of this treatment from the start of the experiment.

Experimental design setup on each site. G_r-sd: M. x giganteus established from rhizomes. G_p-sd: M. x giganteus established from plantlets. S_p-sd: M. sinensis K1399 established from plantlets at single density. S_p-dd: M. sinensis K1399 established from plantlets at double density
Three sub-plots were positioned on each treatment after the establishment phase. For single density (G_r-sd, S_p-sd, G_p-sd), five rows and five plants per row were selected for a plot area of 16 m2 (distance between plants and between rows: 0.8 m). For the double density (S_p-dd), the plot area was 15 m2 with five rows and ten plants per row (distance between plants: 0.5 m; distance between rows: 0.6 m) (Fig. 2).
Yield Measurements
Yields had been measured every year in February–March for the three sub-plots of each treatment, from 2015 to 2019, before harvesting the whole trial. Miscanthus was not harvested the first year (2014); the aboveground biomass was chopped and left on the soil surface. The following years, miscanthus was harvested in late winter (i.e., in February or March) either using a manual cutting tool (brushcutter) (Chanteloup, Subdray, Marne et Gondoire, and Evry sites) or with a silage harvester (Bioferme and Episy sites). To assess yields, we considered only shoots higher than 1 m. Shoots were cut 15 cm from the ground. The fresh matter of the biomass (FM) was immediately weighed using a scale. A sub-sample of 1.5 to 2 kg of fresh matter (FW) was taken and then put in the oven for 48 h at 80 °C and weighed to get its dry weight (DW). The following equation was used to calculate the yield in t of DM per ha:
where Y is the yield (t.ha−1 of DM), FM is the fresh matter of the sub-plot (kg), FW is the fresh weight of the sub-sample (kg), DW is the dry weight of the sub-sample (kg), and A is the sub-plot area (m2).
These measurements were fully described in [5].
Identification of Candidate Indicators Explaining Yield Variations Across Sites
Choice and Calculation Method of Candidate Indicators
Age
Since miscanthus is a perennial crop with yields that increase every year during the first years of growth, variability in yields could be related to the age of the crop. This indicator is expressed in years.
Initial Aerial Biomass (IAB)
The amount of aerial biomass after the first year of growth provides information on the initial conditions for crop startup, which could affect the evolution of yields in the following years. The initial aerial biomass was chopped the first year (in February–March 2014) and left on the soil as a mulch. To assess initial aerial biomass (IAB), 10 plants inside each strip (treatment) but outside the sub-plots were selected to avoid disturbing the sub-plots, cut at 15 cm from the ground, and weighed (SWs). Shoot numbers have been counted (SNs) for these 10 plants. A 2–3-kg sample was put in the oven at 80 °C for 48 h to determine the dry matter (DM). The number of plants on each sub-plot was counted (SNp) and the yields measured in kg.m−2. The sub-plot area (Ap) was 15 or 16 m2. We multiplied by 10 the formula described below to express the aerial biomass in t.ha−1. The aerial biomass produced during the first year was obtained as follows:
with IAB: initial aerial biomass the first year of production (t.ha−1 of DM)
SNs: shoot number per m2 of the sample
SWs: shoot weight of the sample (kg)
SNp: shoot number of the sub-plot (kg)
Ap: area of the sub-plot (m2)
DM: dry matter (%)
Water Stress Indicator (Ks)
The calculation of the water stress indicator (Ks) was carried out from March 25 to October 15. This indicator was previously assessed and used by [17]. The water stress indicator (Ks) ranged from 0 to 1. When the crop was not water-stressed, Ks was equal to 1 and when it was stressed, it was lower than 1. The maximum soil water content (SWCmax) was calculated by the Gras method [19] and was a function of soil texture and rooting depth of M. x giganteus and M. sinensis for each site. Water balance was initialized on March 1 of the growth year, assuming that SWCd on that date was equal to SWCmax (i.e., the soil was at field capacity). SWCd is calculated as follow:
where SWCd is the daily soil water content (mm), SWCd−1 is the previous day soil water content (mm), Pd is the daily precipitation (mm/day), and AETd is the actual daily evapotranspiration (mm/day).
where PETd is the daily potential evapotranspiration, Kcd is the daily crop coefficient defined as a function of degree-days [20] and using data from the French multisite experimental network REGIX and information about Kc of sugarcane as defined by the FAO [21] and ranged from 0.4 to 1.18, and Ksd is the daily water stress indicator.
If SWCd > 2/3 \(\times\) SWCmax, then Ksd = 1, else Ksd = SWCd/SWCmax
where n is the number of days and Ks is the average water stress indicator.
The Number of Frost Days (FD)
This indicator aims to identify the effect of frost on crop regrowth and therefore on yields. Miscanthus is sensitive to frost during crop regrowth. Miscanthus base temperature for regrowth is 6 °C for M. x giganteus [22] and 5 °C for M. sinensis [23]. The number of frost days (FD) corresponds to the number of days spent by the crop under minimum temperatures below 0 °C. The choice of 0 °C is based on the work of [24]. FD is calculated from April 1 to May 30 for M. x giganteus and from March 15 to May 30 for M. sinensis.
The Sum of Degree-Days (DD) of the Previous Year
Miscanthus has a “memory effect” between two successive years of growth: when this crop reaches the end of its development cycle for a given year of growth, it transfers nitrogen from the aerial parts to its rhizomes. It then reuses the nitrogen stored in the rhizomes to start its regrowth the following year [25, 26]. Hence, a sufficient sum of degree-days must be accumulated by the crop to complete its cycle. The following year, the vegetation regrowth could then benefit from a satisfactory N remobilization from the aerial biomass to the rhizome [26]. The sum of the degree-days was calculated as the difference between the average daily temperature and the base temperature of the crop. The calculation of the sum of degree-days was obtained as follows:
where n is the number of days, DD is the sum of degree-days (°C), Td is the daily mean temperature (°C), and T0 is the base temperature of the crop (°C).
The base temperature (T0) of M. x giganteus is 6 °C [22]. A base temperature of 5 °C was used for M. sinensis [23].
Statistical Analysis
First, we performed a principal component analysis (PCA) using the software R version 4.0.2 [27] between the different candidate indicators to detect possible correlations that could lead to confounding effects between candidate indicators.
Second, we used the mixed-model approach [28] to identify indicators that explain variability in yields across sites. This method assesses all possible linear combinations between the yield and candidate indicators. It allowed us to rank the importance of each indicator on yield variability through the Akaïke weight. For each genotype (M. x giganteus and M. sinensis), all the possible linear combinations were performed as follows:
where the explained variable Yi is the yield of the miscanthus (t.ha−1 of DM) for the ith site, x1,i, …, xp,i are the explanatory variables (i.e., Age, IAB, Ks, FD, DD) for the ith site,\(\mu\) is the global average of the explained variables, and α1, …, αp are the parameters associated with the explanatory variables. Yi is assumed to follow independent Gaussian distribution with mean and constant variance σ2. ei ~ N (0, σ2). The mixed-model method [28] was used to select the explanatory variables × 1,..., xp and to estimate the model parameters α1, …,αp. We used the package MMIX [29] to implement this method.
The calculation of the Akaïke weight (wi) was based on the models which have a high predictive value, i.e., with the lowest AIC (Akaïke Information Criteria) [30]. It was calculated as follows:
where wi is the Akaïke weight obtained for the ith combination explanatory variables, AICi is the AIC value for the corresponding model, AICmin is the smallest AIC obtained for all possible model combinations, and n is the number of possible models.
The Akaïke weight wi is the probability that, given a set of models, model i is the best AIC model. The relative importance of each explanatory variable is estimated, i.e., the sum of the Akaïke weights of all models where this explanatory variable is present. The higher the sum of the Akaïke weight was, the more important the considered explanatory variable was [28]. All the explanatory variables can be ranked according to their importance following the sum of the Akaïke weight. The Akaïke weight ranged from 0 to 1.
The next step was to assess the stability of the mixed-model results using the bootstrapping method, as described by [17]. The bootstrap allows us to see if a change in our initial data set leads to a change in our results. If the results obtained after the bootstrap are the same or vary very little from the initial results, our results will be robust. Throughout the bootstrap, we generated new data sets based on the initial data set, i.e., the database with all indicators for each treatment, each species, and for all sites. A total of 10,000 bootstrap samples were generated by random sampling with replacement. We then recalculated the Akaïke weight to assess the uncertainty of the selection method. For each explanatory variable, the frequency of selection, the mean of the estimated parameter values, and the standard deviation of the estimated parameter values were calculated. For a given variable, a high selection frequency combined with a low standard deviation indicates that results are stable. A selection frequency close to 0 or 1 means that the results are stable, while a frequency close to 0.5 indicates an instability in the selection of variables.
Finally, we performed a simple linear regression between yields and the candidate indicators that had been identified by the mixed model as indicators explaining the variability of yields for M. sinensis and M. x giganteus. This regression was performed according to the following formula:
where Yi is the yield in t.ha−1, for the ith site, Xi is the indicators (Age, Ks, FD, IAB, and/or DD depending on mixed-model results) for the ith site, n is the number of explanatory variables considered and is between 1 to 5, \(\mu\) is the global average of the explained variable, and α is the parameter associated with the explanatory variables. Yi is assumed to follow independent Gaussian distribution with mean and constant variance. This regression allowed us to estimate which part of the total yield variability the selected indicators explained (through the R2 value).
Results
Yield Variability Across Site-Years
Yields of M. x giganteus and M. sinensis varied across site-years (Fig. 3). Site-year variability was greater for M. x giganteus (coefficient of variation amounted to 54% and 40% respectively for G_r-sd and G_p-sd) than for M. sinensis (coefficient of variation amounted to 30% and 27% respectively for S_p-sd and S_p-dd). Yields tended to increase with the age of the crop, especially for M. x giganteus (Fig. 3a, b). Yield trends of G_p-sd treatment in Chanteloup site are missing because the establishment of this treatment did not succeed, as mentioned above (Fig. 3b).

Yields trends across site-years for M. x giganteus (a, b) and M. sinensis (c, d). G_r-sd: M. x giganteus established from rhizomes. G_p-sd: M. x giganteus established from plantlets. S_p-sd: M. sinensis K1399 established from plantlets at single density. S_p-dd: M. sinensis K1399 established from plantlets at double density
For M. x giganteus, G_r-sd treatment yields ranged from 0.8 to 20.5 t.ha−1 with an average of 9.9 t.ha−1 and yields of G_p-sd treatment ranged from 3.8 to 19.5 t.ha−1 with an average of 10.9 t.ha−1 (Fig. 3a, b). Average yields of M. sinensis treatments were respectively of 6.7 t.ha−1 and 7.1 t.ha−1 for S_p-sd and S_p-dd. S_p-sd yields ranged from 2.27 to 11.01 t.ha−1 and that of S_p-dd from 4 to 11.9 t.ha−1 (Fig. 3c, d).
Yields and yields of the preceding year of both genotypes were correlated, except for the growth year 2 of M. x giganteus (Fig. 4). For a perennial crop such as Miscanthus, an interpretation of that correlation could be that yields could be significantly affected by yields of the previous year. Another interpretation could be that miscanthus experienced the same limiting factors on the same site (due to soil characteristics for instance) from a year to another. This correlation was found to be stronger for growth year 2 (R2 = 0.51), growth year 3 (R2 = 0.40), and growth year 4 (R2 = 0.72) (Fig. 4b) for M. sinensis and growth year 4 (R2 = 0.82), growth year 5 (R2 = 0.84), and growth year 6 (R2 = 0.66) for M. x giganteus (Fig. 4a).

Relationship between the yields of the preceding growth year and the yields of the current growth year for M. x giganteus (a) and M. sinensis (b) from the second growth year to the fifth growth year. RSS: residual sum of squares. ***p-value < 0.001, **p-value < 0.01, *p-value = 0.05
Relationships Between All Candidate Indicators
The principal component analysis showed that the candidate indicators were not correlated with each other: the risk of confounding effects between indicators is therefore low (Fig. 5).

Principal components analysis between the candidate indicators. FD: frost days, Age: age of cultivation, DD: sum of degree-days of previous year, IAB: initial aerial biomass, Ks: water stress indicator
Identification of Indicators Explaining Yield Variability for Each Genotype
Indicators Explaining M. x giganteus Yield Variability
The sum of Akaïke weight (wi) before the bootstrap showed that age (0.99) and water stress indicator Ks (0.99) mostly explained the variability in yields across site-years. This result was robust after implementing the bootstrap procedure, i.e., when the data set was modified. The sum of Akaïke weight did not change or changed slightly (it was equal to 0.99 and 0.93 for age and Ks respectively, and the other candidate indicators still had a lower sum of Akaïke weights).
Parameter values (Table 2) showed that yields increased with Ks. Years 3 and 6 experienced the highest water stress, with Ks values lower than 0.7 (Fig. 6). Once we take into account this effect of Ks, we also observed in Fig. 6 that yield of miscanthus increased with the age of the crop, as shown in the parameter values in Table 2. When we performed a simple linear model with the crop age and the water stress indicator, we explained 31.9% yield variation across site-years (R2 = 0.32).

M. x giganteus yields as a function of crop age for different values of water stress indicators (Ks). The black triangle is the average yield
Indicators Explaining M. sinensis Yield Variability
The age of the crop (Age), the sum of degree-days in the previous growth year (DD), and the water stress indicator (Ks) best explained the inter-site and interannual variability of M. sinensis yields (Table 3). The sum of Akaïke weight values was similar (for Age and Ks) or only slightly changed after the bootstrap (for DD), which indicated that the results were robust when the data set was modified.
Our results showed that M. sinensis was sensitive to water stress. The lowest yields of M. sinensis are obtained for low Ks values (Fig. 7a). The years that experienced lower average yield were year 3 (5.82 t.ha−1) and year 6 (7.3 t.ha−1), which were also the years with the highest water stress (Fig. 7a).

M. sinensis yield trends as a function of age for different values of water stress indicator (a) and of sum of degree-days of the previous year (b). The black triangle is the average yield
The sum of the degree-days of the previous growth year also explained yield variations of M. sinensis: yields of M. sinensis increased when the sum of degree-days of the previous year was high (Fig. 7b). The lowest yields were obtained for DD values ranging from 2100 and 2300 °C.
A yield can be high while being considered in a stressed condition for an indicator. At this point, it is necessary to relate it to other indicators to explain its position. For example, when we look at the red dots at the top for DD (Fig. 7b), the same dots are green on the Ks figure (Fig. 7a). Therefore, these yields were negatively impacted by DD but these dots were under good water conditions which explains their presence at the top of the DD figure.
When we implemented a simple linear model with the crop age, the water stress indicator, and the sum of degree-days, we explained 49.9% of yield variations across site-years (R2 = 49.9).
Finally, under stress conditions (i.e., for years 3 and 6 that were impacted by Ks and DD) yield losses of M. x giganteus (17% and 37% respectively for years 6 and 3) were higher than for M. sinensis (5% and 15% respectively for yields 6 and 3). The parameter estimations (and the associated standard deviations allowing us to estimate a confidence interval) described in Tables 2 and 3 showed that M. sinensis (with an estimate of the parameter associated with Ks equal to 11.67) was less sensitive to water stress than M. x giganteus (with an estimate of the parameter associated with Ks equal to 15.25) (Tables 2 and 3). The parameter estimation associated with the age effect was also higher for M. x giganteus (2.38) compared to M. sinensis (0.65) (see Tables 2 and 3).
Discussion
The Origin of the Variability in Yields Across Site-Years Is Partially Common for M. x giganteus and M. sinensis
Age is a common indicator explaining yield variability of M. x giganteus and M. sinensis. During the juvenile phase of miscanthus cultivation, yields increase before reaching a plateau [5, 10, 31, 32]. The effect of crop age on yield is more pronounced for M. x giganteus than M. sinensis because of its longer juvenile period [5]. Moreover, the higher the previous year’s yields, the higher the current year’s yields, at least for the juvenile phase. Similar results were obtained by Clifton-Brown et al. [33] who showed that there was a significant R2 of 0.81 between the third and second year and 0.56 between the third year and the first year for several genotypes, including M. x giganteus and M. sinensis during the first 3 years.
M. x giganteus and M. sinensis also shared another common indicator explaining yield variability across site-years: the water stress indicator (Ks). Both M. x giganteus and M. sinensis yields were negatively affected by water stress. These results are consistent with those of da Costa et al. [11] who found that M. sinensis and M. x giganteus were sensitive to water stress after 15 days from the onset of the stress in controlled conditions in the UK. Ings et al. [34] also showed that yields of M. x giganteus were negatively impacted by water stress with a significant reduction of the stem elongation rate after the 20th day of drought as the most sensitive response to water stress, followed by a reduction of chlorophyll content by 42% after the 32nd day of drought, resulting in lower yields. van der Weijde et al. [35] showed that for 50 genotypes of miscanthus (including 39 genotypes of M. sinensis and 7 genotypes of M. x giganteus) in drought conditions; the average loss in plant weight was 45%. However, our results differ from those of Stavridou et al. [12] who did not find a significant decrease in yields of M. x giganteus and M. sinensis under water stress conditions. This result could be explained by the small magnitude of water stress and the low duration of the experiment (8 weeks), which did not allow taking into account the whole growth cycle of miscanthus.
Water deficit affected stem weight and stem height of miscanthus [34, 35]. According to Ouattara et al. and Christian and Haase [5, 36], stems (number, weight, height, and elongation rate) are strongly correlated to miscanthus yields. Hence, these relationships could explain the yield loss in drought conditions, in addition to the effect of water stress on crop physiology as the decrease of chlorophyll content for photosynthesis and stomatal conductance [34, 36].
Our results showed that M. x giganteus was less tolerant to water stress than M. sinensis. Clifton-Brown et al. [1, 8] also showed that M. x giganteus was less tolerant to water stress than M. sinensis, particularly with M. x giganteus senescing more rapidly than M. sinensis in water stress conditions. Yield losses due to water stress of M. sinensis ranged from 9.2 to 12.9 t.ha−1, which is lower than yield losses of M. x giganteus, which ranged from 12.1 to 26.4 t.ha−1 [8]. Similar results were obtained by da Costa et al. [11], who showed that M. sinensis was more tolerant to water stress than M. x giganteus. In water stress conditions, the water use efficiency of M. sinensis was increased by 2% while that of M. x giganteus was reduced by 7% [37]. The morphology of the plant could explain these results. M. sinensis is not as tall as M. x giganteus, and small plants need less water and are less likely to lose water through evapotranspiration due to a proportionally smaller leaf area [38]. According to van der Weijde et al. [35], the miscanthus plants that had the highest biomass production under drought conditions were genotypes with relatively small morphologies (i.e., small height and/or small leaves) and low stem weight under both normal and drought conditions.
Only M. sinensis yields were affected by the sum of degree-days of the previous year. Our results could be explained by the fact that there is a linear relationship between the sum of degree-days of the previous year and the aboveground biomass produced by miscanthus [24]. When the crop has been able to accumulate enough degree-days, it is able to complete its cycle by transferring nitrogen from the aerial parts of the crop (leaves and stems) to the rhizomes. When this transfer has been carried out, the yield of the following year could be higher [22]. The maximum yield achieved by M. sinensis in a growth year is earlier than that of M. x giganteus; however, its senescence period is longer [39], as several stem cohorts appearing as M. sinensis growth cycle progresses (Zapater, pers. comm.). As a result, M. sinensis remains functional somewhat later than M. x giganteus [39], which may explain the fact that M. sinensis is more sensitive to the sum of degree-days in the previous year.
What Could be the Origin of the Unexplained Yield Variability?
The indicators we identified explained 31.9% and 49.9% of the variability in yields for M. x giganteus and M. sinensis respectively. Other factors which were not taken into account in our study could also explain yield variability.
We did not observe an effect of frost on yield variability. However, many authors have shown that frost during regrowth could lead to stem senescence that reduces the number of stems at maturity, thus impacting yields [40,41,42]. Temperatures below 0 °C would kill young miscanthus shoots in winter during plant regrowth [24]. We did not observe this effect, probably because six of our sites were located in the same region and did not experience very low temperatures during the regrowth period of miscanthus (frost days ranged from 0 to 11 days for M. x giganteus and from 1 to 13 days for M. sinensis across the years 2014 to 2018).
Besides, a previous paper on the same experiment showed that the establishment mode of M. x giganteus (from rhizome vs. from rhizome-derived plantlets) and that doubling the density of M. sinensis did not have any effect on yield [5].
Solar radiation was an indispensable factor in photosynthesis. The intercepted photosynthetically active radiation had an effect on the interannual variation in yields. All the more so as the two genotypes did not have the same light utilization efficiency, as M. x giganteus had a light utilization efficiency of 2.4 g.MJ−1 of MS compared to 1.66 g.MJ−1 for M. sinensis [43]. An indicator which takes into account this effect would capture some of the variability specific to each genotype [43].
Miscanthus is a crop with low nitrogen requirements because it stores its nitrogen in the rhizomes at the end of each cycle and re-mobilizes it at the beginning of each cycle [10]. However, P and K inputs are necessary because these elements are not re-mobilized. Therefore, part of the variability could be associated with mineral nutrition [31, 44, 45]. In our study, we estimated the nitrogen nutrition index (NNI) using the critical nitrogen curve of miscanthus [46] in order to use it as a candidate indicator explaining yield variability. Unfortunately, we could not use the NNI because of errors in its measurement. As a perspective, it would be interesting to deepen the analysis of the factors explaining yield variability by identifying indicators related to PK nutrition. K is taken up more by miscanthus than N, and could therefore be a good nutrition indicator [45, 47]. In our analysis, root depth could have been used as an additional indicator to assess the capacity of the crop to uptake soil nutrients (NPK). However, root depth needs to be combined with soil measurements on N, P, or K. Besides, we have already used root depth in the calculation of our indicator of water stress (Ks), which could have increased the risk of confounding effects.
What Perspectives for Genotype Choice for a Given Environment, Crop Management, and Plant Breeding?
As M. sinensis is more tolerant to water stress than M. x giganteus, it could be grown over a wider range of environmental conditions than M. x giganteus, including soil with low water capacity, climatic conditions with low rainfall, and high potential evapotranspiration [48]. This tolerance would allow M. sinensis to achieve yields that vary little across sites and years compared to M. x giganteus [5]. This is consistent with another study suggesting that M. sinensis could thrive in stressful environments, and would likely explain its wide distribution across Asia [49]. To establish miscanthus it would then be necessary to assess the maximum soil water capacity (using information on granulometry and soil depth), and in case of low maximum soil water capacity (especially associated with a low climatic water balance) M. sinensis could be better suited to this environment compared to M. x giganteus.
M. x giganteus achieved lower yields than M. sinensis in some pedo-climatic conditions [5]. On sandy soils, yields of M. x giganteus were lower than 5 t.ha−1 [47], Scharwz and J. Greef, pers. comm reported by Vermerris [50]. This was the finding at the Chanteloup site (shallow sandy soil with a maximum soil water capacity of 149 mm and rainfall lower than 400 mm) where M. sinensis obtained higher yields than M. x giganteus. Scordia et al. [37] found similar results and showed a low yielding of M. x giganteus compared to M. sinensis established in sandy soil in Italy. In a context of climate change, characterized in several regions by a higher rainfall variability, the choice of genotypes for a perennial crop should be for the most stable yields across years [37] and even better if they are high and stable. For this purpose, on sites with low water availability and sandy soils, M. sinensis would be a better choice than M. x giganteus. In 2016, Lewandowski et al. [51] recommended the use of some genotypes of M. sinensis instead of M. x giganteus in Southern Europe because of their resistance to drought, hot summers, and the advantage of having a homogeneous crop in the fields. But so far, M. sinensis seeds are not sterile. To enable their commercial establishment, research on the breeding of sterile M. sinensis seeds is needed to avoid the risk of invasiveness [52].
Hence, miscanthus breeding programs could be developed to provide a range of miscanthus genotypes able to produce high (and/or regular) amounts of biomass per hectare under contrasting pedo-climatic conditions. Studies of 15 miscanthus genotypes at several sites in Europe from 2013 to 2015 identified important genotypes of M. sinensis and M. x giganteus. These studies based on the variability of yields and phenology of the genotypes have issued recommendations of genotypes according to the major climatic zones of Europe [39, 51]. Other species of miscanthus could also be sought for breeding, as the genus miscanthus contains more than 20 species that originate from a large geographical area [53].
Data Availability
The data and material that support the findings of this study are available from the corresponding author upon reasonable request.
Code Availability
The data analysis script is available from the corresponding author upon reasonable request. The script was written on the R software.
References
Clifton-Brown J (2000) Water use efficiency and biomass partitioning of three different Miscanthus genotypes with limited and unlimited water supply. Ann Bot 86:191–200. https://doi.org/10.1006/anbo.2000.1183
Lewandowski I, Scurlock JMO, Lindvall E, Christou M (2003) The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe. Biomass Bioenerg 25:335–361. https://doi.org/10.1016/S0961-9534(03)00030-8
Lewandowski I, Clifton-Brown J, Kiesel A, Hastings A, Yasir I (2018) Miscanthus. In: Perennial grasses for bioenergy and bioproducts. Elsevier, pp 35–59. https://doi.org/10.1016/B978-0-12-812900-5.00002-3
Alexopoulou E, Zanetti F, Scordia D et al (2015) Long-term yields of switchgrass, giant reed, and Miscanthus in the Mediterranean Basin. Bioenerg Res 8:1492–1499. https://doi.org/10.1007/s12155-015-9687-x
Ouattara MS, Laurent A, Barbu C et al (2020) Effects of several establishment modes of Miscanthus × giganteus and Miscanthus sinensis on yields and yield trends. GCB Bioenergy 12:524–538. https://doi.org/10.1111/gcbb.12692
Richter GM, Agostini F, Barker A et al (2016) Assessing on-farm productivity of Miscanthus crops by combining soil mapping, yield modelling and remote sensing. Biomass Bioenerg 85:252–261. https://doi.org/10.1016/j.biombioe.2015.12.024
Zub HW, Brancourt-Hulmel M (2010) Agronomic and physiological performances of different species of Miscanthus, a major energy crop. A review Agron Sustain Dev 30:201–214. https://doi.org/10.1051/agro/2009034
Clifton-Brown JC, Lewandowski I, Bangerth F, Jones MB (2002) Comparative responses to water stress in stay-green, rapid- and slow senescing genotypes of the biomass crop, Miscanthus. New Phytol 154:335–345. https://doi.org/10.1046/j.1469-8137.2002.00381.x
Farrell AD, Clifton-Brown JC, Lewandowski I, Jones MB (2006) Genotypic variation in cold tolerance influences the yield of Miscanthus. Ann Applied Biology 149:337–345. https://doi.org/10.1111/j.1744-7348.2006.00099.x
Lewandowski I, Clifton-Brown JC, Scurlock JMO, Huisman W (2000) Miscanthus: European experience with a novel energy crop. Biomass Bioenerg 19:209–227. https://doi.org/10.1016/S0961-9534(00)00032-5
da Costa RMF, Simister R, Roberts LA et al (2019) Nutrient and drought stress: implications for phenology and biomass quality in miscanthus. Ann Bot 124:553–566. https://doi.org/10.1093/aob/mcy155
Stavridou E, Webster RJ, Robson PRH (2019) Novel Miscanthus genotypes selected for different drought tolerance phenotypes show enhanced tolerance across combinations of salinity and drought treatments. Ann Bot 124:653–674. https://doi.org/10.1093/aob/mcz009
Malinowska M, Donnison I, Robson P (2020) Morphological and physiological traits that explain yield response to drought stress in miscanthus. Agronomy 10:1194. https://doi.org/10.3390/agronomy10081194
De Vega JJ, Teshome A, Klaas M et al (2021) Physiological and transcriptional response to drought stress among bioenergy grass Miscanthus species. Biotechnol Biofuels 14:60. https://doi.org/10.1186/s13068-021-01915-z
Doré T, Sebillotte M, Meynard JM (1997) A diagnostic method for assessing regional variations in crop yield. Agric Syst 54:169–188. https://doi.org/10.1016/S0308-521X(96)00084-4
Doré T, Clermont-Dauphin C, Crozat Y et al (2008) Methodological progress in on-farm regional agronomic diagnosis. A review Agronomy for Sustainable Development 28:151–161. https://doi.org/10.1051/agro:2007031
Lesur-Dumoulin C, Lorin M, Bazot M et al (2016) Analysis of young Miscanthus × giganteus yield variability: a survey of farmers’ fields in east central France. GCB Bioenergy 8:122–135. https://doi.org/10.1111/gcbb.12247
Leclère M, Lorent A-R, Jeuffroy M-H et al (2021) Diagnosis of camelina seed yield and quality across an on-farm experimental network. Eur J Agron 122:126190. https://doi.org/10.1016/j.eja.2020.126190
Chiaverini J, Gras R (1977) Comparaison de quelques methodes d’evaluation au laboratoire de la capacite au champ. Annales Agronomiques 28:445–461
Audoire S (2011) Etude des déterminants agronomiques de la production de biomasse du miscanthus
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration - guidelines for computing crop water requirements - FAO Irrigation and drainage paper 56
Price L, Bullard M, Lyons H et al (2004) Identifying the yield potential of Miscanthus x giganteus: an assessment of the spatial and temporal variability of M. x giganteus biomass productivity across England and Wales. Biomass Bioenerg 26:3–13. https://doi.org/10.1016/S0961-9534(03)00062-X
Kandel TP, Hastings A, Jørgensen U, Olesen JE (2016) Simulation of biomass yield of regular and chilling tolerant Miscanthus cultivars and reed canary grass in different climates of Europe. Ind Crops Prod 86:329–333. https://doi.org/10.1016/j.indcrop.2016.04.007
Clifton-Brown JC, Neilson B, Lewandowski I, Jones MB (2000) The modelled productivity of Miscanthus×giganteus (GREEF et DEU) in Ireland. Ind Crops Prod 12:97–109. https://doi.org/10.1016/S0926-6690(00)00042-X
Christian D, Poulton P, Riche A et al (2006) The recovery over several seasons of 15N-labelled fertilizer applied to Miscanthus×giganteus ranging from 1 to 3 years old. Biomass Bioenerg 30:125–133. https://doi.org/10.1016/j.biombioe.2005.11.002
Strullu L, Cadoux S, Preudhomme M et al (2011) Biomass production and nitrogen accumulation and remobilisation by Miscanthus×giganteus as influenced by nitrogen stocks in belowground organs. Field Crop Res 121:381–391. https://doi.org/10.1016/j.fcr.2011.01.005
R Core Team (2020). — European Environment Agency. https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006. Accessed 1 Feb 2021
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York
Morfin M, Makowski D (2010) MMIX : un package R pour combiner des modèles en agronomie. Cah Tech Inra 41–49. https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwiurOTIkJ_zAhWGyYUKHYNZD4QQFnoECAsQAQ&url=https%3A%2F%2Fwww6.inrae.fr%2Fcahier_des_techniques%2Fcontent%2Fdownload%2F3427%2F32278%2Fversion%2F2%2Ffile%2F41_Makowski_Morfin69.pdf&usg=AOvVaw0yC2F5FjoltFzLI5wYhmHA
Akaike H (1974) A new look at the statistical model identification. IEEE Trans Autom Control 19:716–723. https://doi.org/10.1109/TAC.1974.1100705
Miguez FE, Villamil MB, Long SP, Bollero GA (2008) Meta-analysis of the effects of management factors on Miscanthus×giganteus growth and biomass production. Agric For Meteorol 148:1280–1292. https://doi.org/10.1016/j.agrformet.2008.03.010
Lesur C, Jeuffroy M-H, Makowski D et al (2013) Modeling long-term yield trends of Miscanthus×giganteus using experimental data from across Europe. Field Crop Res 149:252–260. https://doi.org/10.1016/j.fcr.2013.05.004
Clifton-Brown JC, Lewandowski I, Andersson B et al (2001) Performance of 15 Miscanthus Genotypes at Five Sites in Europe. Agron J 93:1013–1019. https://doi.org/10.2134/agronj2001.9351013x
Ings J, Mur LAJ, Robson PRH, Bosch M (2013) Physiological and growth responses to water deficit in the bioenergy crop Miscanthus x giganteus. Front Plant Sci 4:. https://doi.org/10.3389/fpls.2013.00468
van der Weijde T, Huxley LM, Hawkins S et al (2017) Impact of drought stress on growth and quality of Miscanthus for biofuel production. GCB Bioenergy 9:770–782. https://doi.org/10.1111/gcbb.12382
Christian DG, Haase E (2001) Agronomy of Miscanthus. In: Jones M, Walsh M (eds), Miscanthus for energy and fibre, James and James, p 21–45. https://books.google.fr/books?id=85AbeqOJkkcC
Scordia D, Scalici G, Clifton-Brown J et al (2020) Wild Miscanthus germplasm in a drought-affected area: physiology and agronomy appraisals. Agronomy 10:679. https://doi.org/10.3390/agronomy10050679
Blum A (2005) Drought resistance, water-use efficiency, and yield potential—are they compatible, dissonant, or mutually exclusive? Aust J Agric Res 56:1159. https://doi.org/10.1071/AR05069
Nunn C, Hastings AFStJ, Kalinina O, et al (2017) Environmental influences on the growing season duration and ripening of diverse miscanthus germplasm grown in six countries. Front Plant Sci 8:907. https://doi.org/10.3389/fpls.2017.00907
Zub HW, Arnoult S, Younous J et al (2012) The frost tolerance of Miscanthus at the juvenile stage: differences between clones are influenced by leaf-stage and acclimation. Eur J Agron 36:32–40. https://doi.org/10.1016/j.eja.2011.08.001
Domon J-M, Baldwin L, Acket S et al (2013) Cell wall compositional modifications of Miscanthus ecotypes in response to cold acclimation. Phytochemistry 85:51–61. https://doi.org/10.1016/j.phytochem.2012.09.001
Fonteyne S, Muylle H, De Swaef T et al (2016) How low can you go?—Rhizome and shoot frost tolerance in miscanthus germplasm. Ind Crops Prod 89:323–331. https://doi.org/10.1016/j.indcrop.2016.05.031
Davey CL, Jones LE, Squance M et al (2017) Radiation capture and conversion efficiencies of Miscanthus sacchariflorus, M. sinensis and their naturally occurring hybrid M. × giganteus. GCB Bioenergy 9:385–399. https://doi.org/10.1111/gcbb.12331
Heaton EA, Long SP, Voigt TB et al (2004) Miscanthus for renewable energy generation: European Union Experience and Projections for Illinois. Mitig Adapt Strat Glob Change 9:433–451. https://doi.org/10.1023/B:MITI.0000038848.94134.be
Cadoux S, Riche AB, Yates NE, Machet J-M (2012) Nutrient requirements of Miscanthus x giganteus: conclusions from a review of published studies. Biomass Bioenerg 38:14–22. https://doi.org/10.1016/j.biombioe.2011.01.015
Zapater M, Catterou M, Mary B et al (2017) A single and robust critical nitrogen dilution curve for Miscanthus × giganteus and Miscanthus sinensis. Bioenerg Res 10:115–128. https://doi.org/10.1007/s12155-016-9781-8
Roncucci N, Di Nasso NO, N, Tozzini C, et al (2015) Miscanthus × giganteus nutrient concentrations and uptakes in autumn and winter harvests as influenced by soil texture, irrigation and nitrogen fertilization in the Mediterranean. GCB Bioenergy 7:1009–1018. https://doi.org/10.1111/gcbb.12209
Quinn LD, Allen DJ, Stewart JR (2010) Invasiveness potential of Miscanthus sinensis: implications for bioenergy production in the United States: INVASIVENESS POTENTIAL OF MISCANTHUS SINENSIS. GCB Bioenergy 2:310–320. https://doi.org/10.1111/j.1757-1707.2010.01062.x
Stewart JR, Toma Y, Fernández FG et al (2009) The ecology and agronomy of Miscanthus sinensis, a species important to bioenergy crop development, in its native range in Japan: a review. GCB Bioenergy 1:126–153. https://doi.org/10.1111/j.1757-1707.2009.01010.x
Vermerris W (2008) Miscanthus: genetic resources and breeding potential to enhance bioenergy production. In: Vermerris W (ed) Genetic Improvement of Bioenergy Crops. Springer, New York, New York, NY, pp 295–308
Lewandowski I, Clifton-Brown J, Trindade LM et al (2016) Progress on optimizing Miscanthus biomass production for the European bioeconomy: results of the EU FP7 Project OPTIMISC. Front Plant Sci 7: https://doi.org/10.3389/fpls.2016.01620
Clifton-Brown J, Hastings A, Mos M et al (2017) Progress in upscaling Miscanthus biomass production for the European bio-economy with seed-based hybrids. GCB Bioenergy 9:6–17. https://doi.org/10.1111/gcbb.12357
Arnoult S, Brancourt-Hulmel M (2015) A review on Miscanthus biomass production and composition for bioenergy use: genotypic and environmental variability and implications for breeding. Bioenerg Res 8:502–526. https://doi.org/10.1007/s12155-014-9524-7
Acknowledgements
We would like to thank all the partners who contributed to the implementation of the study for their participation and for all the additional information provided for the analysis of the results, especially in the framework of Work Package 2 of the Biomass For the Future project. The authors also thank the technical staff of UMR Agronomie for their support in the management and data collection since 2013, and Wageningen University & Research and Genech for providing plant material. Finally, we thank Liz Carey Libbrecht for language editing this paper. This paper is dedicated to the memory of Mathieu Bazot (deceased in 2017).
Funding
The research from which we obtained these results received funding from the French Government, managed by the Agence Nationale de la Recherche (ANR) under the Investment for the Future program (BIOMASS FOR THE FUTURE ANR11‐BTBR‐0006 project).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ouattara, M.S., Laurent, A., Berthou, M. et al. Identifying Factors Explaining Yield Variability of Miscanthus x giganteus and Miscanthus sinensis Across Contrasting Environments: Use of an Agronomic Diagnosis Approach. Bioenerg. Res. 15, 672–685 (2022). https://doi.org/10.1007/s12155-021-10332-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12155-021-10332-x