
Abstract
Pattern recognition receptors (PRRs) are known for many years for their role in the recognition of microbial products and the subsequent activation of the immune system. The 2011 Nobel Prize for medicine indeed rewarded J. Hoffmann/B. Beutler and R. Steinman for their revolutionary findings concerning the activation of the immune system, thus stressing the significance of understanding the mechanisms of activation of the innate immunity. Such immunostimulatory activities are of major interest in the context of cancer to induce long-term antitumoral responses. Ligands for the toll-like receptors (TLRs), a well-known family of PRR, have been shown to have antitumoral activities in several cancers. Those ligands are now undergoing extensive clinical investigations both as immunostimulant molecules and as adjuvant along with vaccines. However, when considering the use of these ligands in tumor therapy, one shall consider the potential effect on the tumor cells themselves as well as on the entire organism. Recent data indeed demonstrate that TLR activation in tumor cells could trigger both pro- or antitumoral effect depending on the context. This review discusses this balance between the intrinsic activation of PRR in tumor cells and the extrinsic microenvironment activation in term of overall effect of PRR ligands on tumor development. We review recent advances in the field and underline appealing prospects for clinical development of PRR agonists in the light of our current knowledge on their expression and activation.
Similar content being viewed by others


Background
Cancer immunotherapy consists of approaches that mobilize components of the immune system as cancer treatment (reviewed in [1]). The history of cancer immunotherapy dates back to Coley's original observation of the late nineteenth century of antitumor effects from a bacterial concoction [2]. Activation of the immune system was proposed as the mechanism leading to the regression of these measurable tumors. These interpretations are now reinforced by the successful use of purified microbial extracts in cancer treatment as well as the understanding of their mechanism of action.
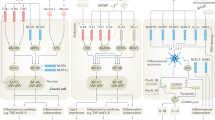
The recognition of pathogens is the key to understand the activation of immune response not only by infectious non-self agents but also by endogenous “danger” signals. Our understanding of innate immunity has led to the discovery of innate sensors or pattern recognition receptor (PRR) being able to recognize pathogen-associated molecular patterns (PAMPs) of all class of microorganisms [3]. Such PRR consists of four families including toll-like receptors (TLRs), nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs), retinoic acid inducible gene-I (RIG-I)-like receptors (RLRs), and DNA sensors (reviewed in [4–6]). Those receptors were recently involved in the sensing of endogenous ligands, referred as danger-associated molecular patterns (DAMP), released by cells in non-infectious conditions such as stress, injury, or cell death [7].
Localization of those receptors at the membranes (TLR) or in the cytosol (RLR, NLR, and DNA sensors) is intimately linked to the nature of the DAMP/PAMP they sense [6]. For instance, TLRs (1/2/4/5/6) involved in bacteria and fungi recognition are expressed at the cell surface, with TLRs specialized in virus recognition through sensing of nucleic acids (TLR3/7/8/9) are located within the endosomes (summarized in Table 1, [6, 8]). In addition to the TLRs, two recently described families of cytosolic innate sensors are involved in virus recognition: (a) the RNA helicases [4] and (b) the cytosolic DNA receptors (DAI, AIM2, and IFI16/p204) [5]. The cytoplasmic RNA helicases, RIG-I, and melanoma differentiation-associated gene 5 (MDA5) and DDX1/DDX21/DHX36 complex [9], sense RNA or DNA viruses that replicate in the cytosol via an RNA intermediate. DNA sensors would be involved in the detection of DNA from viruses and intracellular bacteria [herpes simplex virus (HSV)-1, Listeria monocytogenes, Francisella tularensis, and vesicular stomatitis virus (VSV)] [10]. Finally, NLRs include NOD1/2 involved in bacterial recognition as well as the NLRP family members that sense various PAMP and DAMP [11].
The activation of these pathways relies on various adaptors downstream of the different PRR: myeloid differentiation factor 88 (MYD88) and toll/IL-1 receptor (TIR) domain-containing adaptor inducing interferon β (TRIF) for TLR, mitochondrial antiviral signaling (MAVS) (also known as IPS1/CARDIF/VISA) for RNA helicases, stimulator of interferon genes for most DNA sensors [10], and apoptosis-associated speck-like protein for NLRP.
Triggering of those PRRs results in the induction of proinflammatory cytokines, type I interferon (IFN), and chemokines that altogether participate in the establishment of an adjusted innate and adaptive immune response. Type I IFNs, which have a key role in antiviral immunity, are produced by most cell types through the RNA helicase pathway in response to virus infection [12, 13]. In contrast, plasmacytoid dendritic cells (pDCs), the predominant source of type I IFNα in vivo, are triggered via TLR7 and TLR9 particularly expressed on those cells. As a general theme, membrane-bound PRRs recruit members of the interleukin-1 receptor-associated kinase and tumor necrosis factor receptor-associated factor families to stimulate gene expression via activation of transcription factors, mostly NF-κB, the interferon regulatory factors (IRFs), and MAP kinases. In contrast, the activation of NLRP-1 and -3 [14–19], as well as the DNA sensors AIM2 [14, 20–23], triggers the formation of a multimolecular complex termed inflammasome, which leads to the proteolytic cleavage of pro-interleukin (IL)-1/IL-18 after microbial, DAMPs, or genomic DNA sensing (reviewed in [24]).
Besides cytokine/chemokine production, PRR activation provides essential requirements for the initiation of T cell response: enhancement of antigen uptake, processing, and presentation (through the upregulation of major histocompatibility complex (MHC) and costimulatory molecules expression) by antigen-presenting cells (APCs) including dendritic cells (DCs), followed by migration of those DCs after modulation of chemokine receptor expression. All those events contribute to the activation of antigen-specific T cell [25].
Therefore, the engagement of the innate system via PRR on both immune and tumor cells combined with the triggering of adaptive immune responses provides us with interesting prospects regarding the treatment of cancers. This review discusses the advantages of associating PRR ligands, with a specific focus on TLR and RLR, to conventional therapies in order to promote direct antitumor activity as well as antitumor immune response. Although several mouse models support such therapeutic approach, it is worth remembering that differences in expression and signaling capacity between murine and human PRRs are likely to affect the efficacy for controlling cancer cells. We have therefore essentially focused the discussion on human PRRs.
Toll-like receptors
TLR belongs to the most largely described family of PRRs. Toll was initially discovered in the fruit fly where it was shown to mediate Drosophila dorsoventral embryonic development and innate immune response [26, 27]. The first mammalian homolog of Toll, now known as TLR4, was reported in 1997 by Medzhitov et al. [28]. TLR4 was shown to be involved in the recognition of lipopolysaccharide (LPS), a major cell wall component of gram-negative bacteria. Ten human TLRs have been identified since then. TLRs are type I transmembrane proteins with an extracellular domain consisting of leucine-rich repeats for ligand binding and a cytoplasmic TIR domain for downstream signaling. TLR signaling involves a family of five TIR domain-containing adaptor molecules (MYD88, TRIF, TRAM, MAL, and SARM) [3, 6]. All TLRs require MYD88 for signaling, except TLR3, which uses exclusively TRIF and TLR4 that recruits both adaptors to initiate different signaling pathways. Moreover, besides MYD88 and TRIF, TLR4 uses as well the adaptor molecule MAL for MYD88-dependent NF-κB activation and TRAM to drive TRIF-dependent type I IFN induction via IRF activation. Ensuring that these responses are tightly regulated in order to maintain homeostasis and prevent overwhelming immune activation is thereby crucial and relies on many TLR regulators described over the years [3, 6].
TLR1/2/4/5/6 are mostly involved in bacteria and fungi PAMP recognition (Table 1) [6]. The receptors for nucleic acid-based PAMPs, i.e., TLR3, -7/8, and -9, recognize double stranded (ds), single stranded (ss) viral RNA, and unmethylated CpG DNA, respectively. Ligand for hTLR10 has not yet been described. In contrast with mice [29, 30], the human TLR11 gene is not functional. TLR activation results in transcription of proinflammatory cytokines such as TNFα, IL-1β, IL-12, and IL-6, and in some cases, in type I IFN genes. The cytokine induction pattern is determined by the type of TLR activated, the nature of the ligand, and activated cell.
Expression of TLRs was first documented in immune cells. TLR1 and 6 are ubiquitously expressed in human leukocytes [8, 31, 32]. TLR2/4/5 are restricted to myelomonocytic cells [32–34]. TLR3 is expressed in resting cells by mDC [32] and by some subsets of human B cells [35] and is upregulated in LPS-stimulated monocytes [36]. pDC and B cell specifically express TLR-7, 9, and -10 [8, 37–39]. TLR1/2/6 triggering by bacterial PAMP induce a proinflammatory response in myelomonocytic cells (Table 1). In contrast, triggering of pDC by nucleic acid PAMP induces production of type I IFN, critical for innate antiviral immunity and development of an adequate adaptive immune response [39]. Type I IFN production relies on the activation of transcription factors, especially IRF-3 and -7. TLR3/4 activate IRF3 via TRIF [40, 41], while TLR7/9 induce a response exclusively via MYD88 [42, 43]. In humans, this later response occurs mostly in pDC and depends on IRF7 rather than on IRF3. Unlike other type of cells, pDC constitutively expresses high levels of IRF7 and are therefore preprogrammed to quickly produce high levels of IFNα [42, 43].
While TLR are primarily expressed on immune cells, they have been described in human keratinocytes [44, 45], epithelial cells from the intestinal, urogenital, and respiratory tracts [46], endothelial cells [47], mesenchymal stem cells [48], and various neural cells [49]. In these tissues, they are likely to provide a first line of innate antimicrobial defense. Moreover, several studies have shown that TLRs, as other PRR indeed [50], are capable of controlling intestinal epithelial homeostasis through proliferation and protection from injury [51, 52]. Hasan et al. reported that stimulation of TLRs induces, via MYD88 recruitment, cell cycle entry and progression in fibroblasts [37].
TLR have garnered an extraordinary amount of interest in cancer research due to their role in tumor progression. Besides the extrinsic role of TLR on cells from the tumor microenvironment (immune cells, fibroblasts, and endothelial cells), the intrinsic role of TLR on tumor cells needs to be considered as well. The importance of TLR in host response to tumors became evident as polymorphisms in TLR genes were associated to susceptibility not only to infections but also to cancers [53]. Increasing evidences suggest that TLR might play a dual role in cancer progression (reviewed in [54]). Activation of different TLR might exhibit the exact opposite outcome, antitumor or protumor effects. For example, TLR9 activation was shown to cause breast tumor cell proliferation and increased invasiveness [55] while it inhibited in vitro and in vivo cell proliferation and induced cell death in neuroblastoma cells [56]. Furthermore, TLR expression is modulated on tumor cells, where they might influence tumor growth and host innate immune response. TLR9 expression was shown for instance to be downregulated in head and neck and cervical cancers [57] and upregulated in breast cancer with poorly differentiated phenotype [58]. Increase of TLR3 expression in a percentage of breast cancer patients was for instance associated to a high response rate to synthetic dsRNA, polyadenylic–polyuridylic (Poly[A:U]) treatment [59]. The expression of TLR1 to TLR10 is induced in lymphocytes and macrophages from healthy volunteers following exposure to anticancer agents that induce the p53 network suggesting that these immune receptors can be modulated by DNA metabolic stress [60].
Manipulating TLR signaling in the context of cancer thereby requires a special care particularly with regard to the effect of TLR activation on tumor cells as well as the tumor microenvironment. Moreover, as TLR expression differs between mouse and human, results from preclinical study should be taken very carefully for transfer towards clinical trials.
RNA helicases
RIG-I/DDX58 and MDA5/RH116 (also known as IFIH1 and Helicard) are ubiquitously expressed cytoplasmic sensors involved in RNA detection [4, 13, 61], called RIG-like receptors. These helicases belong to the DExD/H box RNA helicase family including a large number of proteins involved in RNA metabolism [62]. They are able to bind and unwind ssRNA through their helicase domain and signal via their caspase recruitment domains (CARD). Activation of these helicases induces their recruitment to the adaptor protein MAVS located to the mitochondria [63–66]. MAVS relays signals via phosphorylation cascade and nuclear translocation of transcription factors such as NF-κB and IRF3 to coordinate type I IFN production mostly, as well as secretion of proinflammatory cytokines. The third member of the family, laboratory of genetics and physiology 2 (LGP2), lacks CARD domains and acts as a regulator of RIG-I and MDA5 signaling [67–69]. These helicases are involved in virus detection through sensing of their RNA genome. Both helicases sense RNA molecules but with distinct specificity; RIG-I sense 5′-triphosphate (5′tri-P) moiety on RNA backbone [70, 71] while MDA5 is more specific of long dsRNA mimicked in vitro using synthetic polyinosinic–polycytidylic acid (Poly[I:C]) [72] (reviewed in [73]). In that context, RIG-I is essential for the production of type I IFN in response to in vitro-transcribed RNA and RNA viruses including paramyxoviruses (SeV, NDV, and VSV), influenza virus, and Japanese encephalitis virus, whereas MDA5 is critical for Poly[I:C] and picornavirus (EMCV) detection [72]. Phenotypic analyses of a RIG-I deficient mouse model further showed that, besides its involvement in type I IFN induction, RIG-I is an essential negative regulator of physiological myeloid cell differentiation [74]. Besides their role in virus sensing, RLRs are also involved in cancer development. MDA5 was indeed initially found and named for its ability to inhibit colony formation in human melanoma cells [61].
Other DEAD-box proteins were also recently involved in PAMP sensing. Liu and colleagues showed that DHX36 and DHX9 interact respectively with CpG-A and CpG-B oligonucleotides (ODN) and that cells deficient for those helicases display reduced cytokine production [75]. DDX36 and DHX6 thus represent the non-TLR9 MYD88-dependent pathway previously described in pDC [76–78]. More recently, the same group identified DDX1, DDX21, and DHX36 as part of a complex that detects Poly[I:C] in murine splenic DC and signals through TRIF. TRIF(−/−) cells were previously shown to be more deeply impaired in their response to Poly[I:C] than TLR3(−/−) cells suggesting a non-TLR3 TRIF-dependent mechanism of Poly[I:C] sensing [79]. Four systems have thus been reported for Poly[I:C] recognition: (a) the endosomal TLR3-TRIF pathways crucial in epithelial cells [79–81], (b) the RIG-I/MDA5/MAVS mitochondrial pathway that play an important role in fibroblasts and DC [12, 82], (c) the protein kinase R pathway, and (d) this DDX/TRIF pathway in murine DC [9].
The engagement of the innate system via PRR in combination with other anticancer agents therefore provides us with a very exciting prospect of the development of new approaches to treat malignancies. As above mentioned, one shall indeed consider several aspects when thinking about the use of such ligands in cancer immunotherapy: (a) the intrinsic effect on the tumor cells as well as (b) the extrinsic role of TLR activation in cells from the microenvironment and especially immune cells and (c) the adjuvant activity when combined to vaccine. Indeed, human and mice tumors often contain infiltrating immune cells. The potential protective role of these cells has been demonstrated in colorectal cancer patients [83]. Moreover, the development of cancers is favored by immunosuppression in human or immunodeficiency in mice, suggesting a need for reactivation of immune activation in cancer therapy. This review will discuss all these aspects in the prospect of using RLR and TLR ligands, divided in bacterial TLR ligands, TLR3, TLR7/8, and TLR9 ligands, to treat cancer.
Bacterial TLR ligands
As mentioned in the “Background” section, systemic antitumor effects of bacterial infections were reported over a century ago. In retrospect, antitumor effects were probably due to PRR activation, and certainly mostly TLR or NLR, by endotoxins and DNA contained in this heat-killed bacteria preparation. Various bacterial lipopolysaccharides and glycoproteins have been of interest in cancer treatment since Coley's observations. Examples include Bacillus Calmette-Guerin (BCG), whose first published use in cancer dates back to 1935, and Corynebacterium parvum (lung cancer), streptococcal preparation OK432 (carcinomatous pleural effusions), biostim®—glycoprotein extract from Klebsiella pneumoniae (advanced colorectal cancer), and bestatin® from Streptomyces olivoreticuli (leukemia and solid tumors), about 50 years later. However, BCG is the only one to have received regulatory approval. Identification of the recognition of bacteria by TLRs led to the development of purified TLR ligands now in use in clinical trials mostly for their adjuvant activity. As many clinical trials are currently using bacterial TLR ligands as monotherapy or adjuvants in cancer therapy, Table 2 only lists phase III clinical trials.
Bacterial TLR: ligands and cellular expression
TLR ligands involved in bacterial recognition include TLR1/2/4/5/6. mTLR11 was shown to sense uropathogenic Escherichia coli bacteria and profiling-like protein from Toxoplasma gondii [29, 30]; however, its human equivalent contains several stop codons and does not code for a full-length protein [84]. TLR2 acts as heterodimer with TLR1 or TLR6 and senses PAMP from bacteria (lipoarabinomannan, lipoteichoic acid, peptidoglycan, diacyl, or triacyl lipopeptide), fungi (zymosan, together with dectin 1), and parasite (GPI from T. gondii) (Table 1) [6]. As mentioned earlier, TLR4 recognizes LPS, a cell wall component of gram-negative bacteria, and its minimal unit the lipid A. TLR4 was shown too to bind viral proteins from RSV [85, 86], VSV [87], retroviruses [88, 89], and HIV [90]. Moreover, TLR4 was shown to sense self-DAMP liberated during tissue injury (hyaluronan, fibronectin, and heat shock proteins 60 and 70 [30]) or upon chemotherapeutic treatment (high mobility group box-1 (HMGB1); [91]).
TLR1 and 6 are ubiquitously expressed in human leukocytes [8, 31, 32]. TLR2, TLR4, and TLR5 are restricted to myelomonocytic cells, such as mDC, monocytes, neutrophils, and basophils [32–34]. Besides immune cells, such TLRs are also expressed on endothelial cells (TLR2/4 [47]), mesenchymal cells (TLR2/4/5 [48]), neural cells (TLR2/4 [49]), keratinocytes (TLR4/5 [45]), and epithelial cells from intestinal, urogenital, and respiratory tracts. TLR2/4 triggering by bacterial PAMP induce a proinflammatory response in myelomonocytic cells (Table 1). Signaling in epithelial cells via TLR4 and TLR5, in concert with TLR signaling in leukocytes, does participate to the development of intestinal inflammation [46]. Upon ligand binding and endocytosis, activated TLR4 might result as well in type I IFN secretion [92].
Dual role of intrinsic TLR triggering on tumor cells
Hundreds of single nucleotide polymorphisms (SNPs) have been identified in TLRs but their functional consequences are largely unknown. Many associations have been reported between TLR polymorphisms and infectious disease. For instance SNPs in either TLR2/4/5, which are expressed in epithelial cells and involved in bacterial recognition, result in increased risk of sepsis and bacterial infection (reviewed in [53]). Recent analysis of data compiled from several studies showed that in contrast to initial data, no overall association exists between SNPs in the TLR4 and TLR6-1-10 loci and risk of prostate cancer [93]. It seems that in most case, the infection and chronic inflammation most likely mediates the increased risk of cancer development [53].
As for other PRR, expression of TLR2/4/5 is upregulated in various tumor epithelial cells. The upregulation of TLR2/4/5 in gastric epithelial cells from patients with gastric dysplasia and carcinoma suggests that these receptors may play a role in adenocarcinoma development [94]. Similarly, TLR4 was shown to be upregulated in human ovarian [95], colon [96], and head and neck tumor cells [97]. TLR triggering in such epithelium does induce inflammation that has indeed been linked to tumor cell resistance to chemotherapies and cell growth. Triggering TLR4–MYD88 pathway in human ovarian [95] and head and neck tumor cells [97] promoted chemoresistance and tumor growth in vitro [95, 97]. In addition, the MYD88 expression in ovarian cancer tissues correlated with a lower patients' progression-free survival [95]. Pro-tumoral activity of TLR4 signaling was shown to be mediated by both intrinsic and extrinsic mechanisms. LPS triggering of TLR4 directly increases tumor cell proliferation [97]. In parallel, tumor growth was favored via synthesis of soluble factors (IL-6, inducible nitric oxide synthase, IL-12) favoring a pro-tumoral microenvironment. These factors have indeed been reported to increase resistance of tumor cells to cytotoxic attack by both T and NK cells [95, 96] and to favor the development of myeloid-derived suppressor cells. Similarly, TLR2 stimulation by L. monocytogenes promoted tumor growth via the production of immunosuppressive molecules such as IL-6 or nitric oxide [98]. In contrast, intrinsic activation of TLR5 by flagellin inhibited proliferation of breast cancer cells and tumor growth in xenograft model [99]. Altogether, the activation of different TLRs might display completely different outcome.
Triggering of bacterial TLR ligands on immune cells for antitumoral activity
Nowadays, live, attenuated culture preparation of the BCG strain of Mycobacterium bovis [TICE® (Organon Teknika Corp) and PACIS® (BioChem Pharma)] is currently in use for cancer therapy. They were granted FDA approval over 30 years ago for intravesical treatment of in situ carcinoma (CIS) of the urinary bladder and primary or recurrent state papillary tumors (stage I, Ta, and T1). Several multicenter clinical trials showed increased benefits when compared to intravesical doxorubicin treatment based on complete response rate for patients with CIS (70% vs 34% [100], 71% vs 54% [101]) and 5-year progression-free survival for stage I patients (37% vs 17% [100]) (summarized in [102]). BCG is still nowadays the treatment of choice for bladder CIS. However, considering some tolerability issues, alternative immunotherapeutic approaches exploiting the immunogenic components of the mycobacterium (purified cell wall content, CpG DNA) have been trialed in preclinical and clinical studies (summarized in [103]). Almost 20 clinical trials (8 completed, 11 active/recruiting or not—www.clinicaltrials.gov) investigate strategies that could either enhance the efficacy of BCG or replace it with lower toxicity (Table 2).
The precise mechanism by which BCG exerts its antitumor effects is still not well understood. BCG effects do not seem to be mediated by direct proapoptotic effect but rather through a nonspecific immune response [104]. BCG indeed induced a massive increase in the proportion of T helper 1 (Th1) CD4 and γδ T cells, while no significant variation of CD8 and NK cells was found [105]. In vitro killing of human T24 bladder cancer cells induced by live BCG-infected DCs was mostly achieved by γδT cells and NKT cells but not conventional CD8+ CTLs [106]. DC and macrophages are stimulated by BCG preparations through sensing of cell wall skeleton, peptidoglycan, and bacterial DNA by TLR2/4 [107] and TLR9 [108], respectively.
In therapeutic conditions (i.e., following chemotherapy), dead tumor cells express DAMPs that mediate the activation of a protective immune response through the triggering of innate sensors. The group of L. Zitvogel showed over the years that anthracyclin-induced cell death of tumor cells induces expression of cell surface (calreticulin [109]) and soluble molecules (HMGB1 [91], ATP [110]) resulting in tumor elimination by tumor-specific T cells (reviewed in detail in [111]). For instance, the release of HMGB1 by dying cells was shown to trigger TLR4 on DC [91]. Functional TLR4 was shown to be critical for anthracyclin-mediated antitumor response both in mouse model and in individuals with breast cancer carrying a loss-of-function allele of TLR4 [91]. Those data were recently confirmed in vitro in human cellular models [112]. Interestingly, the activation of the NLRP3 inflammasome in DC is crucial as well for generation of tumor-specific T cell [110]. Those data suggest the possible need for the activation of multiple PRR pathways for efficient antitumoral activity.
Adjuvant activity of purified TLR ligands
Lipid A, the biologically active portion of LPS, is known to have potent immunostimulatory properties and has been evaluated for decades as an adjuvant for promoting immune responses to minimally immunogenic antigens, including tumor-associated antigens. Although several lipid A species have been tested, only 3-O-desacyl-4′-monophosphoryl lipid A (MPL) has been evaluated as a cancer vaccine adjuvant in published human clinical trials. MPL, which lacks the saccharide group and most phosphates present in LPS, induces many of the immunostimulatory properties of LPS but is at least 100-fold less toxic. Adjuvant System 04 (AS04, GlaxoSmithKline (GSK)), consisting of MPL and aluminum hydroxide, is currently a component in two licensed vaccines, Cervarix™ and Fendrix™, against human papillomavirus (HPV) and hepatitis B virus (HBV), respectively. AS04 is the first TLR ligand approved as a vaccine adjuvant in humans worldwide. The benefit of combining MPL to aluminum salt has been shown by better adaptive immune responses to immunization with HPV VLP vaccines [113]. MPL leads to a rapid activation of innate immune response that could explain the strong adaptive responses achieved with AS04-adjuvanted vaccines [114]. Clinical trials are being conducted to evaluate the safety and/or efficacy of cancer vaccines containing MPL combined with other immunostimulants, such as cell wall skeleton of Mycobacterium phlei, the saponin QS-21 or with QS-21 and CpG ODN [115] (Table 2).
GSK AS02B (MPL/QS21) or AS15 (CpG/MPL/QS21) adjuvant preparations were tested in melanoma-associated antigen 3 (MAGE-A3) protein vaccine in phase II clinical trials in patients with melanoma and non-small cell lung carcinoma (NSCLC). The adjuvant was shown to be essential for the development of strong humoral and cellular responses against MAGE-A3 epitopes [116]. Combination of MAGE-A3 and AS15 yielded higher specific immune responses and long-lasting clinical response [117] than with AS02B, which lack TLR9 agonist CpG. Large, double-blind phase III clinical trials testing MAGE-A3/AS15 combination (GSK1572932A) in patients with advanced melanoma or NSCLC are currently ongoing. Multicenter, open-label phase I/II trials are currently recruiting to test safety and efficacy of recombinant HER2 protein with AS15 adjuvant in patients with metastatic breast cancer.
MPL adjuvant is also a component of Stimuvax®, a lyophilized liposomal preparation containing BLP25 (MUC1 core peptide) lipopeptide. Phase IIB trial in patients with stage IIIB or IV NSCLC showed an increase in median survival time of 4.5 months in patients receiving BLP25 plus best supportive care (BSC) vs BSC alone [118]. Based on those data, phase III trials have been conducted in NSCLC and breast cancer. As a result of suspected unexpected serious adverse reaction, the company has temporarily suspended those trials (press release).
Finally, DETOX™ adjuvant (Biomira, Inc.), combining MPL and M. phlei wall extract, was administered with cellular or peptide vaccines in phase I/II studies to patients with metastatic breast cancer, melanoma, colon, pancreatic, or lung cancer. A vaccine (Theratope™) combining synthetic carbohydrate molecules (sialyl-Tn (STn)), KLH, and DETOX™ was shown to induce strong anti-STn IgG and IgM responses with minimal side effects in metastatic breast cancer patients [119]. However, recent results from a phase III trial in a large cohort of metastatic breast cancer patients showed no overall benefit in time to progression or survival [120].
Besides its direct antitumoral effect, BCG has been evaluated as well with cellular vaccines in colorectal cancer and melanoma [121, 122]. Clinical benefit along with good safety profile was observed in those clinical trials, though insufficient control arms make it hard to conclude on the benefit of BCG on the actual regimen [121, 122]. Ongoing trials are testing BCG benefit in the adjuvant treatment of melanoma, neuroblastoma, and breast, lung, colon, and ovarian cancer (www.clinicaltrials.gov).
TLR3 ligands
TLR3: ligands and cellular expression
Synthetic Poly[I:C] and Poly[A:U] are known for decades for their potent inhibitor effects on the growth of virus-induced and virus-independent mouse tumors [123, 124]. However, it was only in 2001 that TLR3 was identified as a sensor of dsRNA by Alexopoulou et al. [80]. As previously emphasized, the effects of dsRNA on cancer must not necessarily be attributed to TLR3, as several cytosolic receptors exist [9], which may all contribute to tumor control. Nevertheless, several recent reports have established that tumor cells of various origins express TLR3—its expression can be upregulated by type I IFNs—and that its activation by dsRNA inhibits tumor cell survival. Besides exerting intrinsic activities on TLR3-expressing cancer cells, dsRNA activates the cells from both innate and adaptive immunity and modulates the tumor microenvironment. Those extrinsic roles of TLR3 on tumor progression reflect the expression of the receptor, present not only in immune cells but also in non-immune cells, including epithelial cells, endothelial cells, and fibroblasts.
Intrinsic roles of TLR3 in cancer cells growth, metastasis, and death
To date, the direct inhibition of tumor growth by TLR3 agonists has been reported in vitro for human breast, melanoma, prostate, head and neck, multiple myeloma, clear renal carcinoma, colon, lung, and cervical cancer cells [125–137], suggesting that targeting TLR3 could represent an opportunity for antitumor therapy. TLR3 activation inhibits the growth of cancer cells through two main processes: (a) decrease of proliferation and (b) induction of apoptotic cell death.
A decrease of cell proliferation in response to TLR3 activation by Poly[I:C] dsRNA has been demonstrated by BrDu incorporation experiments for breast and prostate cancer cells [130, 131] and likely participates to the dsRNA antitumoral effect in the other types of cancers listed above. The molecular mechanism remains so far uncertain but could be related to autocrine IFN signaling and blockade of cell cycle through combined downregulation of cyclin D1 and upregulation of cyclin-dependent kinase inhibitor p27 [44, 130, 131]. In contrast, Poly[I:C] stimulation has been reported to increase cell proliferation of head and neck and multiple myeloma cell lines in a c-Myc- and NF κB-dependent manner, respectively [137, 138]. Although TLR3 requirement for these effects remains to be clearly established, production of IL-6 vs IFNα by multiple myeloma cells in response to Poly[I:C] could switch cell decision to proliferate or die [137].
The first demonstration of a direct antitumoral effect of TLR3 activation on tumor cells was reported for breast cancer cell lines in which Poly[I:C] treatment induces caspase-8 activation and caspase-dependent apoptotic cell death [130]. Requirement for caspase-8 activation in TLR3-induced apoptosis was further confirmed in other cancers such as melanoma, nasopharyngeal, prostate, and cervical cancers [127, 129, 131, 134–136] indicating that the extrinsic pathway of apoptosis is triggered by TLR3. TLR3 death-signaling pathway is still under intensive investigations. A recent report [136], as well as our submitted data, led to the identification of a proapoptotic molecular complex containing caspase-8 and responsible for TLR3-induced cell death in cancer cells. The molecular basis of this death complex reminds that of classical death receptors of the TNFR superfamily such as TNF or TRAIL receptors [139]. Its formation requires the protein RIP1, a kinase playing a key role in the integration of signaling pathways from genotoxic stress, death receptors, or microbial infections and closely involved in the cell's decision to live or die [140].
Triggering the extrinsic pathway of apoptosis by TLR3 activation in cancer cells leads the way to combine TLR3 ligands with conventional therapies, such as chemotherapies or radiotherapy that preferentially target the intrinsic mitochondrial pathway of apoptosis, to improve clinical outcomes. In vitro combinations of Poly[I:C] with 5-fluorouracil, cisplatin, or etoposide anticancer drugs indeed increase cell death of HCT116 colon cancer cell line[128]. These in vitro preliminary data are encouraging but need to be extended to other drugs and cancers and to be validated with in vivo models. Furthermore, detailed understanding of molecular mechanisms of TLR3-induced apoptosis in cancer cells should help to the design of novel sensitizing combinations. Typically, Smac mimetic compounds (currently in phase I clinical trial), which inhibit the inhibitor of apoptosis proteins (IAPs) XIAP, cIAP1, and cIAP2, strongly synergize with dsRNA for inducing the apoptosis of various tumor cell lines [127, 134–136]. The molecular mechanism may rely on alterations of RIP1 ubiquitination and stability (our unpublished data). Recently, a large signaling platform termed Ripoptosome has been described to transmit apoptotic and necrotic signals emanating from TLR3 [136]. In response to Smac mimetic compounds, this molecular complex assembles spontaneously in some tumor cells, a feature that could be linked to the level of stress signals present in cancer cells compared to normal cells [136, 141]. The absence of Ripoptosome might explain why normal cells are not killed by Poly[I:C] alone or in combination with IAP antagonists, but undergo only a moderate proliferation slowdown ([134] and our unpublished data). Thus, the benefits of combining IAP inhibitors with TLR3 ligand should be further investigated, notably in preclinical animal models.
Interestingly, phase II clinical trials on glioblastoma suggest that poly-ICLC (Poly[I:C] stabilized with polylysine and carboxymethylcellulose) may improve patient survival when combined with radiotherapy alone or with radiotherapy plus the alkylating agent temolozomide [142, 143] (Table 3). Current phase II trial is analyzing intramuscular injection of poly-ICLC for the treatment of low-grade gliomas. The relevance of TLR3 expression in cancer cells for dsRNA antitumor effects has been recently demonstrated in several immunodeficient mouse models [59]. Moreover, retrospective analysis of primary breast cancers by the use of a new monoclonal antibody anti-TLR3 revealed that TLR3 expression by the tumor is a biomarker for the therapeutic efficacy of dsRNA on metastatic relapse [59]. Complementary data have been obtained with syngeneic tumor mouse models expressing TLR3, for which Poly[A:U] exerts its antitumoral effects by acting on both host and tumor cells TLR3 [144]. While confirming the potential of TLR3 stimulation in immunotherapy (see below), and stressing a role of this receptor in tumor immune surveillance, another study using syngeneic and transgenic mouse prostate tumor concluded that the antitumoral effect of dsRNA was mainly dependent on TLR3-expressing host cells [145]. Such discrepancies might reflect key differences between human and mouse TLR3, regarding both the expression and the signaling of apoptosis and of cytokine induction [146–148]. Indeed, most mouse tumor cells lines were found to be resistant to dsRNA/TLR3-induced apoptosis compared with human tumors (unpublished data). Such variances between mouse and human cancer cells must be kept in mind when transposing to human the results of dsRNA/TLR3-based immunotherapy assays carried on in mouse.
Extrinsic roles of TLR3-L on tumor environment and adjuvant activities
The capacity of TLR3 ligands to activate mDC appears to be the major determinant of its adjuvant ability. Several phase 0/I/II clinical trials are in progress to determine the potential of dsRNA poly-ICLC as adjuvant for antigen peptide vaccinations for various types of cancer (Table 3). Ampligen® Poly[I:C(12)U] (Hemispherx Biopharma of Philadelphia), a GMP-grade synthetic analog of Poly(I:C) shown to specifically target TLR3 [149] is under investigation as an anticancer drug [150]. Poly[I:C(12)U] was previously tested in the treatment of chronic fatigue and AIDS and is currently under clinical investigation with tumor cell lysate for the treatment of ovarian, fallopian, or peritoneal cancers.
Poly[I:C] triggers the upregulation of co-stimulatory molecules on DCs that is partially TRIF-dependent [151] and increases the cross-presentation of exogenous antigen to CD8+ T cells [152]. Combined with antigen, TLR3 agonists thus drive a potent and type I IFN-dependent antigen-specific CD4+ Th1 response [153] and IFN γ production by CD8+ T cells [154].
Secretion of type I IFN and IL 12 by DC or by other accessory cells in response to Poly[I:C] is also responsible for the indirect activation of NK cells associated with increased cytotoxicity and IFN γ secretion [155, 156]. In addition, the membrane-associated tetraspanin-like named IRF 3-dependent NK-activating molecule was shown to be upregulated on the surface of mDC by Poly[I:C] stimulation and to participate to the activation of NK cells via cell–cell contact. Besides these soluble and membrane-bound signals provided by DC, NK requires in addition a direct stimulation of RIG-I by dsRNA to become activated [156].
Related to those combined effects on T cells and NK cells, TLR3 activation was shown to be critically important for cross-presentation of viral antigen by DCs [157] and for priming antiviral CD8+ T cells during acute viral infection. Moreover, Poly[I:C] has been shown to be among the most powerful adjuvant for tumor vaccine [158, 159] requiring both TLR3 [160] and MDA5 [153, 161] to obtain fully protective and long-term immunization. The adjuvant properties of Poly[I:C] in tumor vaccine can still be increased by preventing the type I interferon-dependent myeloid DC apoptosis [154] and the upregulation of B7-H1 co-inhibitory molecule expression by DC [162] that both limit the magnitude of the CD8 T cell response
TLR3 activation triggers the secretion of various chemokines by cells of the microenvironment. Through these chemokines, macrophages, neutrophils, and lymphocytes can be recruited in situ and participate in wound healing [163] or, in the case of cancer, either to the eradication of the tumor or to the establishment of local immunosuppression. Indeed, uncoupling the effect of the immunosuppressive chemokine CCL5 from the release of the immunostimulatory CXCL10 has been shown to augment the anticancer efficacy of TLR3 ligands [144]. Moreover, injection of Poly[I:C] was found to increase the infiltration of T lymphocytes and NK cells within mouse prostate tumor and to enhance tumor eradication [145]. Lastly, synthetic dsRNAs may also interfere in different ways with tumor vascularization by stimulating tumor cells to secrete VEGF [164], by directly increasing the pro-coagulant functions of the endothelial cells that express TLR3 [165], or indirectly by driving the secretion of IFN-γ by immune cells present at the site of the tumor [166].
In conclusion, TLR3 agonists represent promising components of new drug combinations for cancer therapy due to the synergy of direct inhibitory effects on TLR3-expressing cancer cells and indirect activities on the immune and non-immune cells of the tumor microenvironment. Better understanding of the mechanisms of action of TLR3 and of the involvement of other receptors in the response to TLR3 agonists is now required for optimizing the therapeutic potential.
TLR7/8 ligands
TLR7/8/: ligands and cellular expression
TLR7 and TLR8 are structurally closely related members of the TLR family involved in virus recognition. hTLR7 is predominantly expressed in lung, placenta, and spleen. hTLR8 is more abundant in lung and peripheral blood leukocytes [167]. hTLR7 is expressed in B cells, pDC, and eosinophils and hTLR8 in monocytes and mDC including recently described hBDCA3+ blood DC [8, 168]. In line with those data, hBDCA3+ DC responds to TLR8-L but not TLR7L [169, 170]. NK cells functionally express both TLR7 and TLR8 [171].
Viral single-stranded RNAs are the natural ligands for TLR7 and TLR8 [172, 173]. Even though redundant system of ssRNA recognition exists, mTLR7 was shown to be central for in vivo IFNα induction, mediated by pDC, in response to ssRNA viruses (VSV, influenza) [173]. Stimulation of TLR7 and TLR8 is sequence dependent. AU-rich sequences only stimulate hTLR8 responses [174]. GU-rich sequences can trigger the activation of human and mouse TLR7/8 with the exception of mTLR8 [172] leading to the belief that TLR8 is biologically inactive in mice. Recent analysis of TLR8-deficient mice showed increased TLR7 expression and subsequent response to its ligand and elevated serum levels of autoantibodies [175]. This suggests a central role for mTLR8 in the regulation of mTLR7 expression and prevention of spontaneous autoimmunity. However, this highlights a major difference between human and mice in term of response to TLR8-L. Results from preclinical studies in mouse should thus be taken very carefully for transfer to clinical trials.
Additionally, Jurk et al. recently identified a non-uridine-rich RNA sequence motif on a phosphodiester backbone that induces strong IFNα production by PBMC through TLR7 [176]. Silencing RNAs (siRNAs) also trigger the production of IFNα and IFNγ in vivo and in vitro [177, 178]. Their immunostimulatory potential is mediated by PRRs, including TLR7/8 [177, 178] but also TLR3 [179] and RIG-I [71]. Although RNA-based compounds would appear to be interesting candidates for therapeutic approaches, susceptibility to nuclease degradation limits their use. New class of synthetic TLR7/8 agonists referred to as stabilized immune-modulatory RNA compounds have been developed [180, 181]. They display increased stability, induce cytokine/chemokine production in vitro, and were shown to trigger potent antitumor activity in colon and lung cancer model [180]. This antitumoral activity was associated to a decrease in CD4+CD25+FoxP3+ regulatory T cells (Treg) and increase TA-specific IFNγ response [182]. Finally, synthetic small molecules such as imidazoquinolines and nucleoside analogs also activate TLR7 and/or TLR8. Imiquimod® (R837), Gardiquimod, CL097 (3M-001), and Loxoribine preferentially activate TLR7 in contrast to CL075 (3M-002) that mainly targets TLR8. Resiquimod® (R848) triggers both receptors and was shown to be more soluble and more potent in inducing cytokine expression than its family member imiquimod.
Antitumoral efficacy of TLR7/8 agonists as monotherapy in human trials
Two topical formulations of Imiquimod, Zyclara™ (Graceway Pharmaceuticals; 3.75% imiquimod) and Aldara™ (Graceway Pharmaceuticals; 5% imiquimod), have received legal approval in 1997 for the treatment of HPV-related warts and skin keratosis. Aldara™ is approved as well for the treatment of small superficial basal cell carcinoma (BCC) by the FDA and the European Union. Combined clinical and histological assessments showed a 73–75% clearance rate of BCC in two phase III clinical trials [183]. Imiquimod is currently in phases III (with cryosurgery) and IV (with IFNα) trials in patients with BCC. Besides BCC, objective responses have been observed in some patients with melanoma or lentigo maligna (melanoma in situ) when imiquimod was applied to skin metastasis [184–188]. Finally, two ongoing trials are assessing imiquimod topical treatment in patients with breast cancer with chest wall recurrence or skin metastasis (phase II).
More recently, a specific TLR7 agonist, 852A (3M Pharmaceuticals), is being tested in patients with solid tumors and hematologic malignancies. Unlike imiquimod, 852A has pharmaceutical properties that enable it to be formulated for systemic delivery. Initial data from phase I trial showed that intravenous injection of 852A was well tolerated and induced systemic immune activation [189, 190] that eventually resulted in prolonged disease stabilization in patients with stage IV metastatic melanoma [190]. 852A delays tumor growth in mouse model via inhibition of tumor cell line proliferation through type I IFN induction by pDC [191]. 852A is currently in phase II clinical studies for treatment of cancer, including melanoma. Finally, VTX-2337 (VentiRx Pharmaceuticals), a new specific TLR8 ligand, is in phase I/II clinical trial in combination with current regimen for the treatment of lymphoma and various solid tumors. Results of phase I trial in advanced solid tumors including colorectal (n = 9), pancreatic (n = 6), and melanoma (n = 5) cancers showed good safety and tolerability [192].
Topical application of such TLR7/8 agonists is highly effective, whereas their systemic application has been largely unsuccessful for cancer treatment. Explanations might be variable in tumor type and stages, tumor accessibility, as well as the induction of immune unresponsiveness upon systemic injection of TLR ligands. To circumvent such state, Bourquin et al. designed a novel protocol of treatment with repeated R848 injections and treatment-free intervals and showed improvement of the efficacy of cancer therapy [193]. Treatment failure in solid tumors has been associated in a mouse breast cancer model to increased levels of IL 10 [194]. Blockade of IL 10, but not TGFβ, enhanced the antitumor effect of imiquimod by significantly improving the survival in treated mice [194].
Strong cellular antitumor immune responses have been observed in skin tumors treated topically with dual TLR7/8 ligands. pDCs have been involved in antitumoral immunity, especially upon imiquimod treatment. Topical imiquimod application resulted in pDC recruitment to skin lesions in patients with BCC. Clinical response to imiquimod treatment was correlated to pDC number in a melanoma mouse model [195]. Antitumoral activity of pDC was correlated to tumor cell apoptosis via type I IFN production [168] and TRAIL upregulation [196]. In addition to TLR7-induced IFNα production by pDC, TLR8 triggering on human monocytes and mDC leads to the secretion of proinflammatory cytokines such as TNFα, IL12, and MIP1 [197]. The antitumoral activities of TLR7/8 agonists has thus been associated to their ability to induce bioactive IL12p70 secretion by DC, enhancement of NK cell activation [198], as well as a decrease in CD4+CD25+Foxp3+ Treg [199]. TLR8 ligand may even prove superior NK cell activation than imiquimod [198]. Their ability to induce a potent Th1-type immune response and decrease the number of Tregs could lead to a tumor microenvironment more favorable for the host immune system to respond strongly against the tumor.
While TLR7/8 agonists are potent immunomodulators, direct intrinsic effect on tumor cells shall be considered. A recent study indeed stressed the potential adverse effects of TLR7/8 agonists in the context of lung cancer [200]. TLR7/8 are naturally expressed in human lung epithelial cells and upregulated in lung tumor cells. Activation of those receptors was associated with increased tumor cell survival and resistance to apoptosis induced by several chemotherapies in vitro through NF-κB activation and Bcl2 upregulation [200].
Adjuvant activities of TLR7/8 agonists
TLR7/8 ligands were shown to display as well strong adjuvant activities. TLR7/8 ligands (CL075 or R848) induced maturation of monocyte-differentiated DC. Moreover combination with [poly(I:C)] [201] or LPS [202] yielded DC that showed even better profiles in terms of phenotype, migration, as well as capacity to activate NK cells and IFNγ-producing CD8+ CTL [201, 203]. Imiquimod is thus often associated to DC vaccines. Topical application of imiquimod in phase I trial was shown to augment the immunogenicity of melanoma peptide vaccine when administered with systemic FLT3L, to mobilize DC [204]. Those data suggested a potent effect of imiquimod to activate in situ DC to uptake the antigen and induce specific T cell response. Follow-up trial of topical resiquimod in combination with a NY-ESO1 protein in montanide emulsion is ongoing. Several trials are currently testing the vaccination of patients with tumor lysate-loaded DC in the context of brain tumors (phase II) (Table 4). Imiquimod and gardiquimod, two TLR7 ligands, were indeed shown to improve the antitumor effects of tumor lysate-loaded DCs in murine models [205, 206]. Vaccination in patients with ovarian cancer with dendritic cell/tumor fusions with granulocyte–macrophage colony-stimulating factor (GM-CSF) and imiquimod is also being tested (phase II). Finally, Celldex Therapeutics is testing its antibody-based combination immunotherapy in phase II trials in bladder cancer (CDX-1307 co-administered with GM-CSF, Poly-ICLC, and Resiquimod) and various NY-ESO1 malignancies (CX-1401 along with Resiquimod and/or Poly-ICLC). This technology allows targeting of peptide vaccine to DC using antibodies that specifically target those cells (anti-DEC205, anti MR) inducing antigen uptake, further cross-presentation to specific T cells to induce protective antitumoral immune response as shown in mouse models [207, 208]. Targeting NY-ESO-1 to DC using CDX-1401 with topical resiquimod induces robust and broad humoral and cellular responses in advanced cancer patients (press release). CDX-1307 trial has been terminated for unknown reasons (http://clinicaltrial.gov).
TLR7/8 ligands are also used as adjuvants in protein and peptide vaccines in clinical trial in melanoma [MAGE-3 (two phases I/II), gp100 (two phase II), NY-ESO-1 (phase I completed [209], phase I)], glioma (phase I), and neuroblastoma (phase I) (Table 4). Published results from first phase I trial described the use of imiquimod in a series of vaccinations against the NY-ESO-1 antigen in patients with malignant melanoma [209]. Topical imiquimod is well tolerated and induces local inflammation with a high recruitment of APC and T cell. Overall, the vaccine combination induces measurable NY-ESO-1-specific antibodies and IFNγ + CD4 + T cell responses [209].
TLR9 ligands
TLR9: ligands and cellular expression
TLR9 expression is limited in tissue distribution. In humans, its constitutive expression is confined to B cells, pDCs, and polymorphonuclear leukocytes [32–34]. TLR9 is similarly expressed on mouse B cells, macrophages, and neutrophils; however, major differences were observed in mouse DC as all DC subpopulations do express TLR9 [210]. Moreover, TLR9 has been reported to be expressed by some non-immune cells such as gut, respiratory and cervical epithelia [211, 212], and keratinocytes [44, 213].
TLR9 activity has been characterized mainly in response to the synthetic CpG motif-containing ODN [214]. These motifs are expressed as unmethylated CpG motifs in bacterial or viral DNA. It has been shown that TLR9 is critical in the control of bacterial infections with Brucella abortus [215], Streptococcus pneumoniae [216], and Mycobacterium tuberculosis [217]. TLR9 polymorphisms have been associated with an increased risk for Helicobacter infection, which has high incidence in gastric cancers, suggesting that TLR9 is involved in recognition and clearance of Helicobacter [218]. TLR9 also detects many DNA viruses including HSV type 1 and type 2 [219, 220], murine cytomegalovirus [220, 221], adenovirus [220, 222], HPV CpG motifs [57], epstein barr virus (EBV) [223], and recently, adenovirus-associated virus [224]. In addition to the CpG motifs, recent studies have indicated an important role of nucleotide derivatives and the DNA sugar backbone in determining TLR9-dependent immune stimulatory activities [225]. There is increasing evidences suggesting that TLR9 can also sense DAMPs. The release of DAMPs can be linked to the ability of TLRs to induce an immune response and/or to control mechanisms that control tissue integrity [226–229]. For instance, pDCs were shown to sense genomic DNA associated to the cationic antimicrobial peptide LL37. Such complex triggers robust IFN responses by activating endosomal TLR9 thus leading to robust Th1-mediated adaptive response [230]. TLR9 activation by self-DNA–LL-37 conjugate was shown to be involved in autoimmune diseases, as psoriasis and systemic lupus erythematosus [230].
TLR9 recognizes CpG motifs as well as self-DNA itself with certain structures. There are three structurally distinct classes of CpG ODN: CpG-A, CpG-B, and CpG-C. CpG-A preferentially stimulates pDCs to produce type I IFNs, but very little maturation of APC and B cell activation [231]. CpG-B induces very strong B cell proliferation and differentiation and APC maturation and low levels of type I IFN production. CpG-C has combined activities of CpG-A and CpG-B [231]. The optimal CpG motif in mice consists of an unmethylated CpG dinucleotide flanked by two 5′ purines and two 3′ pyrimidines, whereas the optimal motif in humans is TCGTT and/or TCGTA [232].
Two important events for TLR9 signaling were unraveled in the recent years using mouse models. First, localization of TLR9, as TLR3 and TLR7, to the endosome is crucial for its function and requires the multi-transmembrane protein domain-containing protein named UNC93B [233–235]. Second, proteolytic cleavage in an endolysosomal compartment is required for TLR9 activation by CpG [236–238]. In addition, TLR7 and TLR3 were recently shown to be processed in an analogous manner [239].
pDC and B cell activation by TLR9 in response to CpG motifs lead to a robust inflammatory and type I IFN immune responses. IFNα secretion will in turn modulate DC biology by increasing their APC function and migratory capacities, as well as B and T cell trafficking to the lymph node [240]. Ligand activity on TLR9 expressed in human B cells induces proliferation, class-switch recombination, and enhanced antigen-specific antibody production [241]. TLR9 agonists on human B cells induce the expression of the cytokine and chemokine genes (IL-6, MIP1-α, MIP1-β, TNF-α…) as well as the co-stimulatory molecules, Fc receptors, but also anti-apoptotic gene (bcl2), the transcription factors MYC and TCFL5, and genes critical for B cell proliferation and differentiation [242]. TLR9 activation, along with B cell antigen receptor, induces naive B cell differentiation into plasma cells. Direct conjugation of antigen and CpG reveals a mechanism that may operate during the initiation of primary immune responses, leading to enhanced antigen-specific B cell proliferation and differentiation to form extrafollicular plasma cells [243, 244].
TLR9L as immunotherapy in cancer treatment
TLR9 CpG ODNs have been added to the large list of anticancer drugs as monotherapy or in combination with chemotherapy, radiotherapy, and other immunotherapeutic approaches as they increase antigen presentation and boost antitumor T and B cell responses. Several synthetic TLR9 agonists have been developed for clinical grade use and displayed substantial efficacy in the preclinical and clinical models (discussed in detail in [245]). Sequences dedicated to stimulating TLR9 are termed immune-modulatory ODN (IMOs). IMO-2055 (Idera Pharmaceuticals/Merck KGaA) was described in a mouse model when used as a monotherapy and its antitumoral activity was amplified when it was used in combination with chemotherapeutic agents [246, 247] (Table 5). This compound is currently being analyzed in two phase Ib trials in NSCLC (with bevacizumab and erlotinib) and colorectal cancer (with cetuximab and irinotecan) [248]. Based on increased incidence of neutropenia and electrolyte imbalances reported in its phase 1 trial in patients with squamous cell carcinoma of head and neck (SCCHN), Merck will not conduct further clinical development of IMO-2055 at this stage (press release July 2011).
A second panel of ODN has been created termed immunostimulatory sequences (ISS). These short DNA sequences increase the activity of TLR9 and enhance the production of Th1 cells. ISS can be linked to antigens or used alone to suppress a Th2 response. The lead compound ISS-1018 (Dynavax) induces the production of immunoglobulin and type I IFN in vitro by B cells and of IFNβ, IL-12, and TNF-α by pDC [249]. Potential enhancement of antibody-dependent cell-mediated cytotoxicity is also regarded with interest, and studies are ongoing with ISS-1018 in combination with the anti-CD20 or anti-HER2 in lymphomas and breast cancer patients, respectively [250]. The ISS-1018 ODN sequence has been also used in clinical trials alone and combined with current regimen for non-Hodgkin's lymphoma and metastatic colorectal cancer. CpG-7909 (also known as Agatolimod or PF3512676, Pfizer) is another CpG-B-type ODN that is being developed in therapeutic combinations to treat cancers as well as allergy, asthma, and HBV infection (Table 5). The use of CpG-7909 is discussed in depth in the review by Holtick et al. [245]. It is currently in phase II clinical trials for treatment of melanoma, cutaneous T cell, non-Hodgkin, and follicle center and marginal zone B cell lymphomas. However, evaluation in phase III trials in NSCLC was discontinued when it did not show increased efficacy over standard chemotherapy [251]. Interesting phase I/II results have recently been presented by Brody and colleagues. In this study, the combination of local radiotherapy and CpG-7909 to lymphoma sites led to tumor regression even at distant tumor localizations [252].
CpG ODN as adjuvant in peptide vaccine trials
MGN-1703 and MGN-1706 (Mologen AG) are double stem-loop immunomodulating adjuvants that consist of noncoding DNA and are being developed as anticancer TLR9 agonists (Table 5). Individually, these compounds have anticancer effects in preclinical models [253] and positive results were obtained in a phase I clinical trial of MGN-1703 in prostate and colorectal cancer. CpG7909 is being tested too as adjuvant in phase I/II clinical trials along with peptide vaccine, in esophageal cancer (URCL10-177 and TTK-567 peptides), NY-ESO-1 expressing tumors such as prostate cancer (with NY-ESO-1 antigen), and melanoma (MART1, gp100, MAGE-A3, Melan-A, MAGE10). CpG 7909 was tested with KLH and TT protein vaccine in treating patients who have undergone autologous stem cell transplant in phase I trial that was terminated, as risk to benefit profile did not justify continuation of the trials. CpG7909 is being tested along with MPL-based AS15 adjuvant and IL-2 in recombinant MAGE-A3 peptide vaccine. Finally, CpGs are, as previously mentioned, a component of the AS15 adjuvant currently in phase III trials testing MAGE-A3 vaccine in patients with melanoma or NSCLC, and a phase II trial with rHER2 in patients with metastatic breast cancer.
Intrinsic activities of TLR9 in cancers
The role for TLRs in response to infection is clear, in that TLR9 recognizes PAMPs and responds by eliciting a robust antiviral response. However, viruses associated to cancers have developed clever strategies to transform the host cell of infection and also escape immune attack by deregulating somehow the immune response.
TLR9 is differentially expressed on tumor and healthy tissues. In some tumors, TLR9 has been found upregulated when compared to healthy tissue. In prostate or lung cancers, TLR9 and ERα expression is simultaneously increased especially in poorly differentiated tumors [254]. Conversely, a study on the clinical relevance of TLRs in breast cancer showed that tumors with high TLR9 expression by fibroblast-like cells were associated with low probability of metastasis [255]. On the opposite, TLR9 levels are abolished for instance during the transformation of bone marrow cells from myelodysplastic syndromes to overt leukemia [256] and in cervical cancers [57]. We found that TLR9 expression and function were decreased in cervical cancer cell lines and biopsies associated with high-risk type HPV but not with low-risk type HPV [57]. This downmodulation of tlr9 was shown to be mediated by HPV16 oncoproteins via the Nκ-FB pathway [57]. A study by Daud et al. analyzed the changes in TLR expression in women that either cleared or not HPV16 infection and showed that virus clearance was associated with an increase in TLRs including TLR9, while the persistence correlated with a decrease in TLR3, TLR7/8, and TLR9 expression [257]. In all, we assume that viruses have developed an escape mechanism based on the interaction of virus particles with TLR9 (reviewed in [188]). This phenomenon is not only restricted to chronic infections associated to cancer but also to non-viral-induced cancers. TLR9 has also been demonstrated to be expressed in solid tumors including hepatocellular carcinoma, breast, prostate, and lung cancers as well as hematologic malignancies, especially B cell lymphoma and multiple myeloma [55, 258, 259]. Although TLR9 expression has been shown to be associated to several solid tumors, the exact cell type in a given immunohistochemical block needs to be microdissected to fully understand the implication of the receptor in a given cell type.
TLRs and especially TLR9 are differentially regulated depending on the type of cancer. Their dysregulation might impact on the ability of TLR ligands to drive cell proliferation or apoptosis. TLR agonists have been used in a plethora of xenograft models or in vitro studies to determine their pro- or antitumor role on tumor growth. On one hand, CpG-mediated targeting of TLR9 in xenograft models of human ovarian carcinoma, small cell lung cancer, colon cancer, lung adenocarcinoma, and neuroblastoma, led to a decrease of cell proliferation and in the end tumor size when compared to controls [56, 260, 261]. TLR9 agonists were shown as well in vitro to decrease human colon cancer cell proliferation and survival.
In contrast, TLR9 engagement has been shown in other studies to induce proliferation and invasiveness of cancer cells. Triggering of TLR9 using CpG DNA promoted tumor invasion [55] and migration [58] in breast and prostate cancers [262] in in vitro tumor models. This was associated to increased NF κB [58] and matrix metalloproteinase activities, especially MMP13 [55]. Finally, stimulation of TLRs has been shown to be either proapoptotic [263, 264] or anti-apoptotic [265] depending on the TLR, the cell type, and the metabolic condition of the cell. It was for instance shown that treatment with TLR9 ligand increases the survival of nude mice with experimentally induced brain tumors by inducing apoptosis [263]. However, the exact mechanisms remain unclear.
Triggering of extrinsic TLR9 activity in tumor microenvironment
Besides their intrinsic effect on the tumor cells, TLR9 ligands display also extrinsic activities on the microenvironment that could favor or fight against tumor development. The role of TLRs in tumor angiogenesis is quite diverse just as cancer is itself. Cyclooxygenase-2 (COX-2) is known to play a crucial role in Helicobacter pylori-associated gastric cancer. H. pylori was shown to act through TLR2 and 9 to activate the MAPK cascade, leading to COX-2-dependent prostaglandin E2 (PGE2) release and thereby contributing to cancer cell invasion and angiogenesis [266]. On the other hand, TLR9 ligands can also cause antitumor activity, interfering with angiogenesis. For instance, IMO TLR9 agonist inhibited vessel formation and tumor growth [267] by a mechanism still not well understood.
Based of their ability to stimulate human pDC and B cell, TLR9 agonists could provide strong humoral and Th1 cellular responses. Interestingly, the efficacy of CpG in mouse melanoma B16F10 model was shown to be dependent on the cross-talk between pDCs and specific subsets of cDCs [268]. However, lack of TLR9 responsiveness of pDC in breast, ovarian [269], SCCHN [270], and NSCLC [271] cancers and several virus infections is a well-documented observation (discussed in [188]). This shall be taken into consideration in the prospect of using TLR9 ligands in immunotherapy. pDCs from HCV/HIV-infected patients or isolated from ovarian or breast tumors display weak production of IFNα after ex vivo exposure to TLR9 agonists [269, 272–274]. The mechanism involved in lack of TLR9 responsiveness in cancers was sometimes associated to a decreased expression of TLR9 [270] but is still not well understood. Similarly, in vitro infection of B cell by EBV (lymphoblastoid cell line) led to TLR9 downregulation and decreased cytokine response (TNFα, IL8) to CpG DNA [223]. Due to in vitro limitation, the effect of TLR9 downregulation on plasma cell differentiation and humoral responses was not assessed.
RLR
RIG-I and MDA5 are two RNA helicases now considered as a first line of defense against viral dsRNA [72]. MDA5 was first described as a melanoma differentiation-associated gene [61]. It is expressed at low abundance in many normal tissues, whereas expression is induced in normal and cancer cells by type I IFN [275]. MDA5 was shown to display proapoptotic activities when expressed ectopically at high levels [275, 276]. Mutational analysis demonstrated that both CARD and helicases domains were required for full growth inhibition [61]. However, it was unclear how MDA5 was driving apoptosis on its own. Its triggering by complexed Poly[I:C] was indeed recently shown to induce the formation of autophagy vesicles (autophagosomes) and NOXA-dependent cell death [277]. Furthermore, upon apoptotic stimuli, MDA5 is cleaved by caspases, thereby separating the CARD domains from the helicase domain. While MDA5 localizes in the cytoplasm, the helicase-containing fragment is found in the nucleus where it accelerates FasL-induced DNA degradation [276].
While RLR ligands are not yet under clinical trial, in vitro data strongly suggest that triggering MDA5 or RIG-I would represent a promising approach in cancer treatment. Because recognition of 5′-triphosphate RNA (5′triP-RNA) by RIG-I is mostly independent of the RNA sequence and gene silencing is not inhibited by the presence of a 5′triP moiety, both biological activities can be induced by one short dsRNA molecule. Poeck et al. demonstrated that systemic administration of 5′triP-siRNA designed to specifically silence bcl2 and activate RIG-I, triggers melanoma tumor growth inhibition reflected by massive melanoma tumor apoptosis [278]. Downregulation of bcl2 and immune activation via RIG-I (type I IFN, CXCL10, and MHC class I production) lead to apoptosis of melanoma cells [278]. 5′triP-RNA triggering preferentially induced apoptosis in tumor cell, whereas primary cells are less susceptible due to an intact Bcl-xL counter-regulatory pathway [279]. This response requires recruitment of NK cell at tumor site and is associated with the induction of systemic Th1 cytokines (IFN α, IL-12p40, and IFN γ) [278]. Similarly, ovarian cancer cells treated with complexed Poly[I:C] reacted with enhanced expression of HLA-class I, release of cytokines (CXCL10, IL-6, and type I interferon), as well as tumor cell apoptosis [280]. DC engulfed MDA5-activated cancer cells, became activated, and subsequently provided a proinflammatory milieu promoting cytolytic activity and IFN γ secretion by NK cells [280].
RIG-I has been involved as well in antitumor activity in acute myeloid leukemia (AML). RIG-I was shown to be highly upregulated in all-trans retinoic acid (ATRA)-induced terminal granulocytic differentiation of APC cell line [281]. RIG-I-deficient mice displayed an abnormal Gr-1hi/Mac-1hi granulocytic compartment in peripheral blood leukocytes [74], which would evolve to a CML-like phenotype. Furthermore, in vitro colony assays revealed an intrinsic defect of RIG-I-deficient myeloid progenitors to proliferate and differentiate suggesting that RIG-I exerts a critical role in basic regulatory mechanism controlling cell growth and survival [74]. The same group recently published that ATRA-induced RIG-I induction exerts a critical feedback effect, similar to what is seen using type I IFN or ATRA, to restrain cellular proliferation of AML in vivo and in vitro [282]. RIG-I effect is mediated by a strong activation of STAT1 and interferon-stimulated gene induction (such as ICSBP) [282].
Altogether, those data provide strong proof of concept that the engagement of MDA5 (ovarian cancer) or RIG-I (melanoma, AML) exhibits anticancer effect through two essential and interconnected effects: (a) inhibition of proliferation and induction of apoptosis of tumor cells and (b) mobilization of antitumor immune responses. Those RLR further represents promising ligands to use in cancer therapy.
Concluding remarks
Agonists of TLR and other innate sensors are actively pursued for therapeutic applications in oncology based on three major rationales. First, TLR agonists are potent immunomodulators that can strongly stimulate innate immunity and initiate long-lasting adaptive immunity through systemic activity. The more advanced molecules developed in clinic along with this rational mainly included (a) whole bacteria and defined TLR2/4 ligands and (b) TLR9 ligands. Second, certain ligands have demonstrated antitumor activity based on direct tumoricidal activity on tumor cells, these mainly included TLR3 and TLR7 ligands. Third, vaccine companies are developing TLR ligands as vaccine adjuvant based on local delivery together with formulated tumor antigen leading to local activation and DC maturation resulting in initiation of long-lasting adaptive immunity. Many TLR ligands are developed along this line; the most advanced in clinic include TLR9 and TLR4 ligands. Finally, recent advances in the field of RLR lead to very exciting preclinical data promoting RLR ligands to the level of TLR agonists as potential immunotherapeutic agents in cancer treatment.
For all these lines of investigations, the development has been held back by the limited transposition of mouse preclinical models to human clinic. For example, the wide distribution of TLR9 in mouse APC compared to its limited expression to pDC and B cells in human makes the mouse preclinical models for both vaccine development and systemic delivery not predictable.
For clinical development, we also have to take into consideration detrimental activities of TLR ligands such as pro-tumoral activity through indirect inflammatory process or protective activity against chemotherapy through direct pro-survival effect. The clinical development of these ligands aimed at treating patients with advanced tumors and potential immune defect. Importantly, it will be critical for the future clinical trials to (a) define the expected mechanisms of action and (b) determine the functional competence of the identified TLR target cells in patients (DC in periphery or at tumor sites). Based on these understandings, it is expected that TLR ligands will undoubtedly be successfully developed in clinical trials and will allow to reach therapeutic responses in cancer patients through elicitation of long-lasting antitumor immunity.
References
Dillman RO (2011) Cancer immunotherapy. Cancer Biother Radiopharm 26:1–64
Coley WB (1991) The treatment of malignant tumors by repeated inoculations of erysipelas. With a report of ten original cases. 1893. Clin Orthop Relat Res 262:3–11
Goutagny N, Fitzgerald KA (2006) Pattern recognition receptors: an update. Expert Rev Clin Immunol 2:569–583
Akira S, Uematsu S, Takeuchi O (2006) Pathogen recognition and innate immunity. Cell 124:783–801
Barber GN (2011) Innate immune DNA sensing pathways: STING, AIMII and the regulation of interferon production and inflammatory responses. Curr Opin Immunol 23:10–20
Barton GM, Kagan JC (2009) A cell biological view of Toll-like receptor function: regulation through compartmentalization. Nat Rev Immunol 9:535–542
Kono H, Rock KL (2008) How dying cells alert the immune system to danger. Nat Rev Immunol 8:279–289
Hornung V, Rothenfusser S, Britsch S, Krug A, Jahrsdorfer B, Giese T et al (2002) Quantitative expression of toll-like receptor 1–10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J Immunol 168:4531–4537
Zhang Z, Kim T, Bao M, Facchinetti V, Jung SY, Ghaffari AA et al (2011) DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 34:866–878
Ishikawa H, Ma Z, Barber GN (2009) STING regulates intracellular DNA-mediated, type I interferon-dependent innate immunity. Nature 461:788–792
Bryant C, Fitzgerald KA (2009) Molecular mechanisms involved in inflammasome activation. Trends Cell Biol 19:455–464
Kato H, Sato S, Yoneyama M, Yamamoto M, Uematsu S, Matsui K et al (2005) Cell type-specific involvement of RIG-I in antiviral response. Immunity 23:19–28
Yoneyama M, Kikuchi M, Natsukawa T, Shinobu N, Imaizumi T, Miyagishi M et al (2004) The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol 5:730–737
Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL et al (2008) Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat Immunol 9:847–856
Kanneganti TD, Ozoren N, Body-Malapel M, Amer A, Park JH, Franchi L et al (2006) Bacterial RNA and small antiviral compounds activate caspase-1 through cryopyrin/Nalp3. Nature 440:233–236
Kanneganti TD, Body-Malapel M, Amer A, Park JH, Whitfield J, Franchi L et al (2006) Critical role for Cryopyrin/Nalp3 in activation of caspase-1 in response to viral infection and double-stranded RNA. J Biol Chem 281:36560–36568
Mariathasan S, Newton K, Monack DM, Vucic D, French DM, Lee WP et al (2004) Differential activation of the inflammasome by caspase-1 adaptors ASC and Ipaf. Nature 430:213–218
Mariathasan S, Weiss DS, Newton K, McBride J, O’Rourke K, Roose-Girma M et al (2006) Cryopyrin activates the inflammasome in response to toxins and ATP. Nature 440:228–232
Martinon F, Petrilli V, Mayor A, Tardivel A, Tschopp J (2006) Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 440:237–241
Fernandes-Alnemri T, Yu JW, Datta P, Wu J, Alnemri ES (2009) AIM2 activates the inflammasome and cell death in response to cytoplasmic DNA. Nature 458:509–513
Kim S, Bauernfeind F, Ablasser A, Hartmann G, Fitzgerald KA, Latz E et al (2010) Listeria monocytogenes is sensed by the NLRP3 and AIM2 inflammasome. Eur J Immunol 40:1545–1551
Rathinam VA, Jiang Z, Waggoner SN, Sharma S, Cole LE, Waggoner L et al (2010) The AIM2 inflammasome is essential for host defense against cytosolic bacteria and DNA viruses. Nat Immunol 11:395–402
Roberts TL, Idris A, Dunn JA, Kelly GM, Burnton CM, Hodgson S et al (2009) HIN-200 proteins regulate caspase activation in response to foreign cytoplasmic DNA. Science 323:1057–1060
Petrilli V, Dostert C, Muruve DA, Tschopp J (2007) The inflammasome: a danger sensing complex triggering innate immunity. Curr Opin Immunol 19:615–622
Iwasaki A, Medzhitov R (2004) Toll-like receptor control of the adaptive immune responses. Nat Immunol 5:987–995
Anderson KV, Bokla L, Nusslein-Volhard C (1985) Establishment of dorsal-ventral polarity in the Drosophila embryo: the induction of polarity by the Toll gene product. Cell 42:791–798
Hoffmann JA (2003) The immune response of Drosophila. Nature 426:33–38
Medzhitov R, Preston-Hurlburt P, Janeway CA Jr (1997) A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 388:394–397
Yarovinsky F, Zhang D, Andersen JF, Bannenberg GL, Serhan CN, Hayden MS et al (2005) TLR11 activation of dendritic cells by a protozoan profilin-like protein. Science 308:1626–1629
Zhang Z, Schluesener HJ (2006) Mammalian toll-like receptors: from endogenous ligands to tissue regeneration. Cell Mol Life Sci 63:2901–2907
Hayashi F, Means TK, Luster AD (2003) Toll-like receptors stimulate human neutrophil function. Blood 102:2660–2669
Muzio M, Bosisio D, Polentarutti N, D’Amico G, Stoppacciaro A, Mancinelli R et al (2000) Differential expression and regulation of toll-like receptors (TLR) in human leukocytes: selective expression of TLR3 in dendritic cells. J Immunol 164:5998–6004
Komiya A, Nagase H, Okugawa S, Ota Y, Suzukawa M, Kawakami A et al (2006) Expression and function of toll-like receptors in human basophils. Int Arch Allergy Immunol 140(Suppl 1):23–27
Mansson A, Cardell LO (2009) Role of atopic status in Toll-like receptor (TLR)7- and TLR9-mediated activation of human eosinophils. J Leukoc Biol 85:719–727
Genestier L, Taillardet M, Mondiere P, Gheit H, Bella C, Defrance T (2007) TLR agonists selectively promote terminal plasma cell differentiation of B cell subsets specialized in thymus-independent responses. J Immunol 178:7779–7786
Pan ZK, Fisher C, Li JD, Jiang Y, Huang S, Chen LY (2011) Bacterial LPS up-regulated TLR3 expression is critical for antiviral response in human monocytes: evidence for negative regulation by CYLD. Int Immunol 23:357–364
Hasan UA, Trinchieri G, Vlach J (2005) Toll-like receptor signaling stimulates cell cycle entry and progression in fibroblasts. J Biol Chem 280:20620–20627
Jarrossay D, Napolitani G, Colonna M, Sallusto F, Lanzavecchia A (2001) Specialization and complementarity in microbial molecule recognition by human myeloid and plasmacytoid dendritic cells. Eur J Immunol 31:3388–3393
Kadowaki N, Ho S, Antonenko S, Malefyt RW, Kastelein RA, Bazan F et al (2001) Subsets of human dendritic cell precursors express different toll-like receptors and respond to different microbial antigens. J Exp Med 194:863–869
Fitzgerald KA, Rowe DC, Barnes BJ, Caffrey DR, Visintin A, Latz E et al (2003) LPS-TLR4 signaling to IRF-3/7 and NF-kappaB involves the toll adapters TRAM and TRIF. J Exp Med 198:1043–1055
Oshiumi H, Matsumoto M, Funami K, Akazawa T, Seya T (2003) TICAM-1, an adaptor molecule that participates in Toll-like receptor 3-mediated interferon-beta induction. Nat Immunol 4:161–167
Honda K, Yanai H, Mizutani T, Negishi H, Shimada N, Suzuki N et al (2004) Role of a transductional-transcriptional processor complex involving MyD88 and IRF-7 in Toll-like receptor signaling. Proc Natl Acad Sci USA 101:15416–15421
Kawai T, Sato S, Ishii KJ, Coban C, Hemmi H, Yamamoto M et al (2004) Interferon-alpha induction through Toll-like receptors involves a direct interaction of IRF7 with MyD88 and TRAF6. Nat Immunol 5:1061–1068
Hasan UA, Caux C, Perrot I, Doffin AC, Menetrier-Caux C, Trinchieri G et al (2007) Cell proliferation and survival induced by Toll-like receptors is antagonized by type I IFNs. Proc Natl Acad Sci USA 104:8047–8052
Kollisch G, Kalali BN, Voelcker V, Wallich R, Behrendt H, Ring J et al (2005) Various members of the Toll-like receptor family contribute to the innate immune response of human epidermal keratinocytes. Immunology 114:531–541
Gribar SC, Anand RJ, Sodhi CP, Hackam DJ (2008) The role of epithelial Toll-like receptor signaling in the pathogenesis of intestinal inflammation. J Leukoc Biol 83:493–498
Grote K, Schuett H, Schieffer B (2011) Toll-like receptors in angiogenesis. Sci World J 11:981–991
Hwa CH, Bae YC, Jung JS (2006) Role of toll-like receptors on human adipose-derived stromal cells. Stem Cells 24:2744–2752
Bsibsi M, Ravid R, Gveric D, van Noort JM (2002) Broad expression of Toll-like receptors in the human central nervous system. J Neuropathol Exp Neurol 61:1013–1021
Dupaul-Chicoine J, Yeretssian G, Doiron K, Bergstrom KS, McIntire CR, LeBlanc PM et al (2010) Control of intestinal homeostasis, colitis, and colitis-associated colorectal cancer by the inflammatory caspases. Immunity 32:367–378
Salcedo R, Worschech A, Cardone M, Jones Y, Gyulai Z, Dai RM et al (2010) MyD88-mediated signaling prevents development of adenocarcinomas of the colon: role of interleukin 18. J Exp Med 207:1625–1636
Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R (2004) Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 118:229–241
El-Omar EM, Ng MT, Hold GL (2008) Polymorphisms in Toll-like receptor genes and risk of cancer. Oncogene 27:244–252
Huang B, Zhao J, Unkeless JC, Feng ZH, Xiong H (2008) TLR signaling by tumor and immune cells: a double-edged sword. Oncogene 27:218–224
Merrell MA, Ilvesaro JM, Lehtonen N, Sorsa T, Gehrs B, Rosenthal E et al (2006) Toll-like receptor 9 agonists promote cellular invasion by increasing matrix metalloproteinase activity. Mol Cancer Res 4:437–447
Brignole C, Marimpietri D, Di PD, Perri P, Morandi F, Pastorino F et al (2010) Therapeutic targeting of TLR9 inhibits cell growth and induces apoptosis in neuroblastoma. Cancer Res 70:9816–9826
Hasan UA, Bates E, Takeshita F, Biliato A, Accardi R, Bouvard V et al (2007) TLR9 expression and function is abolished by the cervical cancer-associated human papillomavirus type 16. J Immunol 178:3186–3197
Berger R, Fiegl H, Goebel G, Obexer P, Ausserlechner M, Doppler W et al (2010) Toll-like receptor 9 expression in breast and ovarian cancer is associated with poorly differentiated tumors. Cancer Sci 101:1059–1066
Salaun B, Zitvogel L, Asselin-Paturel C, Morel Y, Chemin K, Dubois C et al (2011) TLR3 as a biomarker for the therapeutic efficacy of double-stranded RNA in breast cancer. Cancer Res 71:1607–1614
Menendez D, Shatz M, Azzam K, Garantziotis S, Fessler MB, Resnick MA (2011) The Toll-like receptor gene family is integrated into human DNA damage and p53 networks. PLoS Genet 7:e1001360
Kang DC, Gopalkrishnan RV, Wu Q, Jankowsky E, Pyle AM, Fisher PB (2002) mda-5: an interferon-inducible putative RNA helicase with double-stranded RNA-dependent ATPase activity and melanoma growth-suppressive properties. Proc Natl Acad Sci USA 99:637–642
Linder P (2006) Dead-box proteins: a family affair—active and passive players in RNP-remodeling. Nucleic Acids Res 34:4168–4180
Kawai T, Takahashi K, Sato S, Coban C, Kumar H, Kato H et al (2005) IPS-1, an adaptor triggering RIG-I- and Mda5-mediated type I interferon induction. Nat Immunol 6:981–988
Meylan E, Curran J, Hofmann K, Moradpour D, Binder M, Bartenschlager R et al (2005) Cardif is an adaptor protein in the RIG-I antiviral pathway and is targeted by hepatitis C virus. Nature 437:1167–1172
Seth RB, Sun L, Ea CK, Chen ZJ (2005) Identification and characterization of MAVS, a mitochondrial antiviral signaling protein that activates NF-kappaB and IRF 3. Cell 122:669–682
Xu LG, Wang YY, Han KJ, Li LY, Zhai Z, Shu HB (2005) VISA is an adapter protein required for virus-triggered IFN-beta signaling. Mol Cell 19:727–740
Rothenfusser S, Goutagny N, DiPerna G, Gong M, Monks BG, Schoenemeyer A et al (2005) The RNA helicase Lgp2 inhibits TLR-independent sensing of viral replication by retinoic acid-inducible gene-I. J Immunol 175:5260–5268
Satoh T, Kato H, Kumagai Y, Yoneyama M, Sato S, Matsushita K et al (2010) LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses. Proc Natl Acad Sci USA 107:1512–1517
Venkataraman T, Valdes M, Elsby R, Kakuta S, Caceres G, Saijo S et al (2007) Loss of DExD/H box RNA helicase LGP2 manifests disparate antiviral responses. J Immunol 178:6444–6455
Pichlmair A, Schulz O, Tan CP, Naslund TI, Liljestrom P, Weber F et al (2006) RIG-I-mediated antiviral responses to single-stranded RNA bearing 5′-phosphates. Science 314:997–1001
Hornung V, Ellegast J, Kim S, Brzozka K, Jung A, Kato H et al (2006) 5′-Triphosphate RNA is the ligand for RIG-I. Science 314:994–997
Kato H, Takeuchi O, Sato S, Yoneyama M, Yamamoto M, Matsui K et al (2006) Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 441:101–105
Kato H, Takeuchi O, Mikamo-Satoh E, Hirai R, Kawai T, Matsushita K et al (2008) Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J Exp Med 205:1601–1610
Zhang NN, Shen SH, Jiang LJ, Zhang W, Zhang HX, Sun YP et al (2008) RIG-I plays a critical role in negatively regulating granulocytic proliferation. Proc Natl Acad Sci USA 105:10553–10558
Kim T, Pazhoor S, Bao M, Zhang Z, Hanabuchi S, Facchinetti V et al (2010) Aspartate-glutamate-alanine-histidine box motif (DEAH)/RNA helicase A helicases sense microbial DNA in human plasmacytoid dendritic cells. Proc Natl Acad Sci USA 107:15181–15186
Jung A, Kato H, Kumagai Y, Kumar H, Kawai T, Takeuchi O et al (2008) Lymphocytoid choriomeningitis virus activates plasmacytoid dendritic cells and induces a cytotoxic T-cell response via MyD88. J Virol 82:196–206
Hochrein H, Schlatter B, O’Keeffe M, Wagner C, Schmitz F, Schiemann M et al (2004) Herpes simplex virus type-1 induces IFN-alpha production via Toll-like receptor 9-dependent and -independent pathways. Proc Natl Acad Sci USA 101:11416–11421
Hokeness-Antonelli KL, Crane MJ, Dragoi AM, Chu WM, Salazar-Mather TP (2007) IFN-alphabeta-mediated inflammatory responses and antiviral defense in liver is TLR9-independent but MyD88-dependent during murine cytomegalovirus infection. J Immunol 179:6176–6183
Yamamoto M, Sato S, Hemmi H, Hoshino K, Kaisho T, Sanjo H et al (2003) Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 301:640–643
Alexopoulou L, Holt AC, Medzhitov R, Flavell RA (2001) Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 413:732–738
Yamamoto M, Sato S, Mori K, Hoshino K, Takeuchi O, Takeda K et al (2002) Cutting edge: a novel Toll/IL-1 receptor domain-containing adapter that preferentially activates the IFN-beta promoter in the Toll-like receptor signaling. J Immunol 169:6668–6672
Yoneyama M, Kikuchi M, Matsumoto K, Imaizumi T, Miyagishi M, Taira K et al (2005) Shared and unique functions of the DExD/H-box helicases RIG-I, MDA5, and LGP2 in antiviral innate immunity. J Immunol 175:2851–2858
Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C et al (2006) Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science 313:1960–1964
Balenga NA, Balenga NA (2007) Human TLR11 gene is repressed due to its probable interaction with profilin expressed in human. Med Hypotheses 68:456
Haynes LM, Moore DD, Kurt-Jones EA, Finberg RW, Anderson LJ, Tripp RA (2001) Involvement of toll-like receptor 4 in innate immunity to respiratory syncytial virus. J Virol 75:10730–10737
Kurt-Jones EA, Popova L, Kwinn L, Haynes LM, Jones LP, Tripp RA et al (2000) Pattern recognition receptors TLR4 and CD14 mediate response to respiratory syncytial virus. Nat Immunol 1:398–401
Georgel P, Jiang Z, Kunz S, Janssen E, Mols J, Hoebe K et al (2007) Vesicular stomatitis virus glycoprotein G activates a specific antiviral Toll-like receptor 4-dependent pathway. Virology 362:304–313
Rassa JC, Meyers JL, Zhang Y, Kudaravalli R, Ross SR (2002) Murine retroviruses activate B cells via interaction with toll-like receptor 4. Proc Natl Acad Sci USA 99:2281–2286
Rolland A, Jouvin-Marche E, Viret C, Faure M, Perron H, Marche PN (2006) The envelope protein of a human endogenous retrovirus-W family activates innate immunity through CD14/TLR4 and promotes Th1-like responses. J Immunol 176:7636–7644
Cavaleiro R, Brunn GJ, Albuquerque AS, Victorino RM, Platt JL, Sousa AE (2007) Monocyte-mediated T cell suppression by HIV-2 envelope proteins. Eur J Immunol 37:3435–3444
Apetoh L, Ghiringhelli F, Tesniere A, Obeid M, Ortiz C, Criollo A et al (2007) Toll-like receptor 4-dependent contribution of the immune system to anticancer chemotherapy and radiotherapy. Nat Med 13:1050–1059
Kagan JC, Su T, Horng T, Chow A, Akira S, Medzhitov R (2008) TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-beta. Nat Immunol 9:361–368
Lindstrom S, Hunter DJ, Gronberg H, Stattin P, Wiklund F, Xu J et al (2010) Sequence variants in the TLR4 and TLR6-1-10 genes and prostate cancer risk. Results based on pooled analysis from three independent studies. Cancer Epidemiol Biomarkers Prev 19:873–876
Pimentel-Nunes P, Soares JB, Roncon-Albuquerque R Jr, Dinis-Ribeiro M, Leite-Moreira AF (2010) Toll-like receptors as therapeutic targets in gastrointestinal diseases. Expert Opin Ther Targets 14:347–368
Kelly MG, Alvero AB, Chen R, Silasi DA, Abrahams VM, Chan S et al (2006) TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res 66:3859–3868
Huang B, Zhao J, Li H, He KL, Chen Y, Chen SH et al (2005) Toll-like receptors on tumor cells facilitate evasion of immune surveillance. Cancer Res 65:5009–5014
Szczepanski MJ, Czystowska M, Szajnik M, Harasymczuk M, Boyiadzis M, Kruk-Zagajewska A et al (2009) Triggering of Toll-like receptor 4 expressed on human head and neck squamous cell carcinoma promotes tumor development and protects the tumor from immune attack. Cancer Res 69:3105–3113
Huang B, Zhao J, Shen S, Li H, He KL, Shen GX et al (2007) Listeria monocytogenes promotes tumor growth via tumor cell toll-like receptor 2 signaling. Cancer Res 67:4346–4352
Cai Z, Sanchez A, Shi Z, Zhang T, Liu M, Zhang D (2011) Activation of Toll-like receptor 5 on breast cancer cells by flagellin suppresses cell proliferation and tumor growth. Cancer Res 71:2466–2475
Lamm DL, Blumenstein BA, Crawford ED, Montie JE, Scardino P, Grossman HB et al (1991) A randomized trial of intravesical doxorubicin and immunotherapy with bacille Calmette-Guerin for transitional-cell carcinoma of the bladder. N Engl J Med 325:1205–1209
Khanna OP, Son DL, Mazer H, Read J, Nugent D, Cottone R et al (1990) Multicenter study of superficial bladder cancer treated with intravesical bacillus Calmette-Guerin or adriamycin. Urology 35:101–108
Shelley MD, Mason MD, Kynaston H (2010) Intravesical therapy for superficial bladder cancer: a systematic review of randomised trials and meta-analyses. Cancer Treat Rev 36:195–205
Dovedi SJ, Davies BR (2009) Emerging targeted therapies for bladder cancer: a disease waiting for a drug. Cancer Metastasis Rev 28:355–367
Alexandroff AB, Jackson AM, O’Donnell MA, James K (1999) BCG immunotherapy of bladder cancer: 20 years on. Lancet 353:1689–1694
Ponticiello A, Perna F, Maione S, Stradolini M, Testa G, Terrazzano G et al (2004) Analysis of local T lymphocyte subsets upon stimulation with intravesical BCG: a model to study tuberculosis immunity. Respir Med 98:509–514
Higuchi T, Shimizu M, Owaki A, Takahashi M, Shinya E, Nishimura T et al (2009) A possible mechanism of intravesical BCG therapy for human bladder carcinoma: involvement of innate effector cells for the inhibition of tumor growth. Cancer Immunol Immunother 58:1245–1255
Tsuji S, Matsumoto M, Takeuchi O, Akira S, Azuma I, Hayashi A et al (2000) Maturation of human dendritic cells by cell wall skeleton of Mycobacterium bovis bacillus Calmette-Guerin: involvement of toll-like receptors. Infect Immun 68:6883–6890
von Meyenn F, Schaefer M, Weighardt H, Bauer S, Kirschning CJ, Wagner H et al (2006) Toll-like receptor 9 contributes to recognition of Mycobacterium bovis bacillus Calmette-Guerin by Flt3-ligand generated dendritic cells. Immunobiology 211:557–565
Obeid M, Tesniere A, Ghiringhelli F, Fimia GM, Apetoh L, Perfettini JL et al (2007) Calreticulin exposure dictates the immunogenicity of cancer cell death. Nat Med 13:54–61
Ghiringhelli F, Apetoh L, Tesniere A, Aymeric L, Ma Y, Ortiz C et al (2009) Activation of the NLRP3 inflammasome in dendritic cells induces IL-1beta-dependent adaptive immunity against tumors. Nat Med 15:1170–1178
Zitvogel L, Kepp O, Kroemer G (2011) Immune parameters affecting the efficacy of chemotherapeutic regimens. Nat Rev Clin Oncol 8:151–160
Fucikova J, Kralikova P, Fialova A, Brtnicky T, Rob L, Bartunkova J et al (2011) Human tumor cells killed by anthracyclines induce a tumor-specific immune response. Cancer Res 71:4821–4833
Giannini SL, Hanon E, Moris P, Van MM, Morel S, Dessy F et al (2006) Enhanced humoral and memory B cellular immunity using HPV16/18 L1 VLP vaccine formulated with the MPL/aluminium salt combination (AS04) compared to aluminium salt only. Vaccine 24:5937–5949
Didierlaurent AM, Morel S, Lockman L, Giannini SL, Bisteau M, Carlsen H et al (2009) AS04, an aluminum salt- and TLR4 agonist-based adjuvant system, induces a transient localized innate immune response leading to enhanced adaptive immunity. J Immunol 183:6186–6197
Cluff CW (2009) Monophosphoryl lipid A (PML) as an adjuvant for anti-cancer vaccines: clinical results. In: Jeannin JF (ed) Lipid A in cancer therapy. Landes Bioscience, Austin
Atanackovic D, Altorki NK, Stockert E, Williamson B, Jungbluth AA, Ritter E et al (2004) Vaccine-induced CD4+ T cell responses to MAGE-3 protein in lung cancer patients. J Immunol 172:3289–3296
Kruit WH (2008) Immunization with recombinant MAGE-A3 protein combined with adjuvant systems AS15 or AS02B in patients with unresectabke and progressive metastatic cutaneous melanoma: a randomized open-label phase II study of the EORTC Melanoma Group (16032-18031). J Clin Oncol 26:9065
Butts C, Maksymiuk A, Goss G, Soulieres D, Marshall E, Cormier Y et al (2011) Updated survival analysis in patients with stage IIIB or IV non-small-cell lung cancer receiving BLP25 liposome vaccine (L-BLP25): phase IIB randomized, multicenter, open-label trial. J Cancer Res Clin Oncol 137(9):1337–1342
Miles DW, Towlson KE, Graham R, Reddish M, Longenecker BM, Taylor-Papadimitriou J et al (1996) A randomised phase II study of sialyl-Tn and DETOX-B adjuvant with or without cyclophosphamide pretreatment for the active specific immunotherapy of breast cancer. Br J Cancer 74:1292–1296
Miles D, Roche H, Martin M, Perren TJ, Cameron DA, Glaspy J et al (2011) Phase III multicenter clinical trial of the sialyl-TN (STn)-keyhole limpet hemocyanin (KLH) vaccine for metastatic breast cancer. Oncologist 16(8):1092–1100
Sharma P, Bajorin DF, Jungbluth AA, Herr H, Old LJ, Gnjatic S (2008) Immune responses detected in urothelial carcinoma patients after vaccination with NY-ESO-1 protein plus BCG and GM-CSF. J Immunother 31:849–857
Vermorken JB, Claessen AM, van Tinteren H, Gall HE, Ezinga R, Meijer S et al (1999) Active specific immunotherapy for stage II and stage III human colon cancer: a randomised trial. Lancet 353:345–350
Sarma PS, Shiu G, Neubauer RH, Baron S, Huebner RJ (1969) Virus-induced sarcoma of mice: inhibition by a synthetic polyribonucleotide complex. Proc Natl Acad Sci USA 62:1046–1051
Levy HB, Law LW, Rabson AS (1969) Inhibition of tumor growth by polyinosinic-polycytidylic acid. Proc Natl Acad Sci USA 62:357–361
Yu M, Lam J, Rada B, Leto TL, Levine SJ (2011) Double-stranded RNA induces shedding of the 34-kDa soluble TNFR1 from human airway epithelial cells via TLR3-TRIF-RIP1-dependent signaling: roles for dual oxidase 2- and caspase-dependent pathways. J Immunol 186:1180–1188
Whitson JM, Noonan EJ, Pookot D, Place RF, Dahiya R (2009) Double stranded-RNA-mediated activation of P21 gene induced apoptosis and cell cycle arrest in renal cell carcinoma. Int J Cancer 125:446–452
Weber A, Kirejczyk Z, Besch R, Potthoff S, Leverkus M, Hacker G (2010) Proapoptotic signalling through Toll-like receptor-3 involves TRIF-dependent activation of caspase-8 and is under the control of inhibitor of apoptosis proteins in melanoma cells. Cell Death Differ 17:942–951
Taura M, Fukuda R, Suico MA, Eguma A, Koga T, Shuto T et al (2010) TLR3 induction by anticancer drugs potentiates poly I:C-induced tumor cell apoptosis. Cancer Sci 101:1610–1617
Salaun B, Lebecque S, Matikainen S, Rimoldi D, Romero P (2007) Toll-like receptor 3 expressed by melanoma cells as a target for therapy? Clin Cancer Res 13:4565–4574
Salaun B, Coste I, Rissoan MC, Lebecque SJ, Renno T (2006) TLR3 can directly trigger apoptosis in human cancer cells. J Immunol 176:4894–4901
Paone A, Starace D, Galli R, Padula F, De CP, Filippini A et al (2008) Toll-like receptor 3 triggers apoptosis of human prostate cancer cells through a PKC-alpha-dependent mechanism. Carcinogenesis 29:1334–1342
Morikawa T, Sugiyama A, Kume H, Ota S, Kashima T, Tomita K et al (2007) Identification of Toll-like receptor 3 as a potential therapeutic target in clear cell renal cell carcinoma. Clin Cancer Res 13:5703–5709
Jiang Q, Wei H, Tian Z (2008) Poly I:C enhances cycloheximide-induced apoptosis of tumor cells through TLR3 pathway. BMC Cancer 8:12
Friboulet L, Pioche-Durieu C, Rodriguez S, Valent A, Souquere S, Ripoche H et al (2008) Recurrent overexpression of c-IAP2 in EBV-associated nasopharyngeal carcinomas: critical role in resistance to Toll-like receptor 3-mediated apoptosis. Neoplasia 10:1183–1194
Friboulet L, Gourzones C, Tsao SW, Morel Y, Paturel C, Temam S et al (2010) Poly(I:C) induces intense expression of c-IAP2 and cooperates with an IAP inhibitor in induction of apoptosis in cancer cells. BMC Cancer 10:327
Feoktistova M, Geserick P, Kellert B, Dimitrova DP, Langlais C, Hupe M et al (2011) cIAPs block ripoptosome formation, a RIP1/caspase-8 containing intracellular cell death complex differentially regulated by cFLIP isoforms. Mol Cell 43:449–463
Chiron D, Pellat-Deceunynck C, Amiot M, Bataille R, Jego G (2009) TLR3 ligand induces NF-κB activation and various fates of multiple myeloma cells depending on IFN-κ production. J Immunol 182:4471–4478
Pries R, Hogrefe L, Xie L, Frenzel H, Brocks C, Ditz C et al (2008) Induction of c-Myc-dependent cell proliferation through toll-like receptor 3 in head and neck cancer. Int J Mol Med 21:209–215
Wilson NS, Dixit V, Ashkenazi A (2009) Death receptor signal transducers: nodes of coordination in immune signaling networks. Nat Immunol 10:348–355
Festjens N, Vanden Berghe T, Cornelis S, Vandenabeele P (2007) RIP1, a kinase on the crossroads of a cell’s decision to live or die. Cell Death Differ 14:400–410
Tenev T, Bianchi K, Darding M, Broemer M, Langlais C, Wallberg F et al (2011) The Ripoptosome, a signaling platform that assembles in response to genotoxic stress and loss of IAPs. Mol Cell 43:432–448
Grossman SA, Ye X, Piantadosi S, Desideri S, Nabors LB, Rosenfeld M et al (2010) Survival of patients with newly diagnosed glioblastoma treated with radiation and temozolomide in research studies in the United States. Clin Cancer Res 16:2443–2449
Butowski N, Chang SM, Lamborn KR, Polley MY, Parvataneni R, Hristova-Kazmierski M et al (2010) Enzastaurin plus temozolomide with radiation therapy in glioblastoma multiforme: a phase I study. Neuro Oncol 12:608–613
Conforti R, Ma Y, Morel Y, Paturel C, Terme M, Viaud S et al (2010) Opposing effects of toll-like receptor (TLR3) signaling in tumors can be therapeutically uncoupled to optimize the anticancer efficacy of TLR3 ligands. Cancer Res 70:490–500
Chin AI, Miyahira AK, Covarrubias A, Teague J, Guo B, Dempsey PW et al (2010) Toll-like receptor 3-mediated suppression of TRAMP prostate cancer shows the critical role of type I interferons in tumor immune surveillance. Cancer Res 70:2595–2603
Rehli M (2002) Of mice and men: species variations of Toll-like receptor expression. Trends Immunol 23:375–378
Lundberg AM, Drexler SK, Monaco C, Williams LM, Sacre SM, Feldmann M et al (2007) Key differences in TLR3/poly I:C signaling and cytokine induction by human primary cells: a phenomenon absent from murine cell systems. Blood 110:3245–3252
Heinz S, Haehnel V, Karaghiosoff M, Schwarzfischer L, Muller M, Krause SW et al (2003) Species-specific regulation of Toll-like receptor 3 genes in men and mice. J Biol Chem 278:21502–21509
Gowen BB, Wong MH, Jung KH, Sanders AB, Mitchell WM, Alexopoulou L et al (2007) TLR3 is essential for the induction of protective immunity against Punta Toro Virus infection by the double-stranded RNA (dsRNA), poly(I:C12U), but not Poly(I:C): differential recognition of synthetic dsRNA molecules. J Immunol 178:5200–5208
Jasani B, Navabi H, Adams M (2009) Ampligen: a potential toll-like 3 receptor adjuvant for immunotherapy of cancer. Vaccine 27:3401–3404
Hoebe K, Janssen EM, Kim SO, Alexopoulou L, Flavell RA, Han J et al (2003) Upregulation of costimulatory molecules induced by lipopolysaccharide and double-stranded RNA occurs by Trif-dependent and Trif-independent pathways. Nat Immunol 4:1223–1229
Datta SK, Redecke V, Prilliman KR, Takabayashi K, Corr M, Tallant T et al (2003) A subset of Toll-like receptor ligands induces cross-presentation by bone marrow-derived dendritic cells. J Immunol 170:4102–4110
Longhi MP, Trumpfheller C, Idoyaga J, Caskey M, Matos I, Kluger C et al (2009) Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J Exp Med 206:1589–1602
Fuertes Marraco SA, Scott CL, Bouillet P, Ives A, Masina S, Vremec D et al (2011) Type I interferon drives dendritic cell apoptosis via multiple BH3-only proteins following activation by PolyIC in vivo. PLoS One 6:e20189
McCartney S, Vermi W, Gilfillan S, Cella M, Murphy TL, Schreiber RD et al (2009) Distinct and complementary functions of MDA5 and TLR3 in poly(I:C)-mediated activation of mouse NK cells. J Exp Med 206:2967–2976
Perrot I, Deauvieau F, Massacrier C, Hughes N, Garrone P, Durand I et al (2010) TLR3 and Rig-like receptor on myeloid dendritic cells and Rig-like receptor on human NK cells are both mandatory for production of IFN-gamma in response to double-stranded RNA. J Immunol 185:2080–2088
Schulz O, Diebold SS, Chen M, Naslund TI, Nolte MA, Alexopoulou L et al (2005) Toll-like receptor 3 promotes cross-priming to virus-infected cells. Nature 433:887–892
Wick DA, Martin SD, Nelson BH, Webb JR (2011) Profound CD8+ T cell immunity elicited by sequential daily immunization with exogenous antigen plus the TLR3 agonist poly(I:C). Vaccine 29:984–993
Aranda F, Llopiz D, Diaz-Valdes N, Riezu-Boj JI, Bezunartea J, Ruiz M et al (2011) Adjuvant combination and antigen targeting as a strategy to induce polyfunctional and high-avidity T-cell responses against poorly immunogenic tumors. Cancer Res 71:3214–3224
Salaun B, Greutert M, Romero P (2009) Toll-like receptor 3 is necessary for dsRNA adjuvant effects. Vaccine 27:1841–1847
Wang Y, Cella M, Gilfillan S, Colonna M (2010) Cutting edge: polyinosinic:polycytidylic acid boosts the generation of memory CD8 T cells through melanoma differentiation-associated protein 5 expressed in stromal cells. J Immunol 184:2751–2755
Pulko V, Liu X, Krco CJ, Harris KJ, Frigola X, Kwon ED et al (2009) TLR3-stimulated dendritic cells up-regulate B7-H1 expression and influence the magnitude of CD8 T cell responses to tumor vaccination. J Immunol 183:3634–3641
Lin Q, Fang D, Fang J, Ren X, Yang X, Wen F et al (2011) Impaired wound healing with defective expression of chemokines and recruitment of myeloid cells in TLR3-deficient mice. J Immunol 186:3710–3717
Paone A, Galli R, Gabellini C, Lukashev D, Starace D, Gorlach A et al (2010) Toll-like receptor 3 regulates angiogenesis and apoptosis in prostate cancer cell lines through hypoxia-inducible factor 1 alpha. Neoplasia 12:539–549
Zimmer S, Steinmetz M, Asdonk T, Motz I, Coch C, Hartmann E et al (2011) Activation of endothelial toll-like receptor 3 impairs endothelial function. Circ Res 108:1358–1366
Berge M, Bonnin P, Sulpice E, Vilar J, Allanic D, Silvestre JS et al (2010) Small interfering RNAs induce target-independent inhibition of tumor growth and vasculature remodeling in a mouse model of hepatocellular carcinoma. Am J Pathol 177:3192–3201
Chuang TH, Ulevitch RJ (2000) Cloning and characterization of a sub-family of human toll-like receptors: hTLR7, hTLR8 and hTLR9. Eur Cytokine Netw 11:372–378
Robbins SH, Walzer T, Dembele D, Thibault C, Defays A, Bessou G et al (2008) Novel insights into the relationships between dendritic cell subsets in human and mouse revealed by genome-wide expression profiling. Genome Biol 9:R17
Poulin LF, Salio M, Griessinger E, Anjos-Afonso F, Craciun L, Chen JL et al (2010) Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8alpha + dendritic cells. J Exp Med 207:1261–1271
Jongbloed SL, Kassianos AJ, McDonald KJ, Clark GJ, Ju X, Angel CE et al (2010) Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J Exp Med 207:1247–1260
Hart OM, Athie-Morales V, O’Connor GM, Gardiner CM (2005) TLR7/8-mediated activation of human NK cells results in accessory cell-dependent IFN-gamma production. J Immunol 175:1636–1642
Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S et al (2004) Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science 303:1526–1529
Lund JM, Alexopoulou L, Sato A, Karow M, Adams NC, Gale NW et al (2004) Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc Natl Acad Sci USA 101:5598–5603
Forsbach A, Nemorin JG, Montino C, Muller C, Samulowitz U, Vicari AP et al (2008) Identification of RNA sequence motifs stimulating sequence-specific TLR8-dependent immune responses. J Immunol 180:3729–3738
Demaria O, Pagni PP, Traub S, de Gassart A, Branzk N, Murphy AJ et al (2010) TLR8 deficiency leads to autoimmunity in mice. J Clin Invest 120:3651–3662
Jurk M, Chikh G, Schulte B, Kritzler A, Richardt-Pargmann D, Lampron C et al (2011) Immunostimulatory potential of silencing RNAs can be mediated by a non-uridine-rich Toll-like receptor 7 motif. Nucleic Acid Ther 21(3):201–214
Hornung V, Guenthner-Biller M, Bourquin C, Ablasser A, Schlee M, Uematsu S et al (2005) Sequence-specific potent induction of IFN-alpha by short interfering RNA in plasmacytoid dendritic cells through TLR7. Nat Med 11:263–270
Sledz CA, Holko M, de Veer MJ, Silverman RH, Williams BR (2003) Activation of the interferon system by short-interfering RNAs. Nat Cell Biol 5:834–839
Kleinman ME, Yamada K, Takeda A, Chandrasekaran V, Nozaki M, Baffi JZ et al (2008) Sequence- and target-independent angiogenesis suppression by siRNA via TLR3. Nature 452:591–597
Lan T, Kandimalla ER, Yu D, Bhagat L, Li Y, Wang D et al (2007) Stabilized immune modulatory RNA compounds as agonists of Toll-like receptors 7 and 8. Proc Natl Acad Sci USA 104:13750–13755
Kandimalla ER, Struthers M, Bett AJ, Wisniewski T, Dubey SA, Jiang W et al (2011) Synthesis and immunological activities of novel Toll-like receptor 7 and 8 agonists. Cell Immunol 270(2):126–134
Wang D, Precopio M, Lan T, Yu D, Tang JX, Kandimalla ER et al (2010) Antitumor activity and immune response induction of a dual agonist of Toll-like receptors 7 and 8. Mol Cancer Ther 9:1788–1797
Geisse J, Caro I, Lindholm J, Golitz L, Stampone P, Owens M (2004) Imiquimod 5% cream for the treatment of superficial basal cell carcinoma: results from two phase III, randomized, vehicle-controlled studies. J Am Acad Dermatol 50:722–733
Bong AB, Bonnekoh B, Franke I, Schon MP, Ulrich J, Gollnick H (2002) Imiquimod, a topical immune response modifier, in the treatment of cutaneous metastases of malignant melanoma. Dermatology 205:135–138
Turza K, Dengel LT, Harris RC, Patterson JW, White K, Grosh WW et al (2010) Effectiveness of imiquimod limited to dermal melanoma metastases, with simultaneous resistance of subcutaneous metastasis. J Cutan Pathol 37:94–98
Wolf IH, Cerroni L, Kodama K, Kerl H (2005) Treatment of lentigo maligna (melanoma in situ) with the immune response modifier imiquimod. Arch Dermatol 141:510–514
Wolf IH, Smolle J, Binder B, Cerroni L, Richtig E, Kerl H (2003) Topical imiquimod in the treatment of metastatic melanoma to skin. Arch Dermatol 139:273–276
Hirsch I, Caux C, Hasan U, Bendriss-Vermare N, Olive D (2010) Impaired Toll-like receptor 7 and 9 signaling: from chronic viral infections to cancer. Trends Immunol 31:391–397
Dudek AZ, Yunis C, Harrison LI, Kumar S, Hawkinson R, Cooley S et al (2007) First in human phase I trial of 852A, a novel systemic toll-like receptor 7 agonist, to activate innate immune responses in patients with advanced cancer. Clin Cancer Res 13:7119–7125
Dummer R, Hauschild A, Becker JC, Grob JJ, Schadendorf D, Tebbs V et al (2008) An exploratory study of systemic administration of the toll-like receptor-7 agonist 852A in patients with refractory metastatic melanoma. Clin Cancer Res 14:856–864
Inglefield JR, Dumitru CD, Alkan SS, Gibson SJ, Lipson KE, Tomai MA et al (2008) TLR7 agonist 852A inhibition of tumor cell proliferation is dependent on plasmacytoid dendritic cells and type I IFN. J Interferon Cytokine Res 28:253–263
Cohen P, Northfelt D, Weiss GJ, Von Hoff DD, Manjarrez K, Ditesch G et al (2011) Phase I clinical trial of VTX-2337, a selective toll-like receptor 8 (TLR8) agonist, in patients with advanced solid tumors. J Clin Oncol 29:2537
Bourquin C, Hotz C, Noerenberg D, Voelkl A, Heidegger S, Roetzer LC et al (2011) Systemic cancer therapy with a small molecule agonist of Toll-like receptor 7 can be improved by circumventing TLR tolerance. Cancer Res 71:5123–5133
Lu H, Wagner WM, Gad E, Yang Y, Duan H, Amon LM et al (2010) Treatment failure of a TLR-7 agonist occurs due to self-regulation of acute inflammation and can be overcome by IL-10 blockade. J Immunol 184:5360–5367
Palamara F, Meindl S, Holcmann M, Luhrs P, Stingl G, Sibilia M (2004) Identification and characterization of pDC-like cells in normal mouse skin and melanomas treated with imiquimod. J Immunol 173:3051–3061
Stary G, Bangert C, Tauber M, Strohal R, Kopp T, Stingl G (2007) Tumoricidal activity of TLR7/8-activated inflammatory dendritic cells. J Exp Med 204:1441–1451
Gorden KB, Gorski KS, Gibson SJ, Kedl RM, Kieper WC, Qiu X et al (2005) Synthetic TLR agonists reveal functional differences between human TLR7 and TLR8. J Immunol 174:1259–1268
Gorski KS, Waller EL, Bjornton-Severson J, Hanten JA, Riter CL, Kieper WC et al (2006) Distinct indirect pathways govern human NK-cell activation by TLR-7 and TLR-8 agonists. Int Immunol 18:1115–1126
Peng G, Guo Z, Kiniwa Y, Voo KS, Peng W, Fu T et al (2005) Toll-like receptor 8-mediated reversal of CD4+ regulatory T cell function. Science 309:1380–1384
Cherfils-Vicini J, Platonova S, Gillard M, Laurans L, Validire P, Caliandro R et al (2010) Triggering of TLR7 and TLR8 expressed by human lung cancer cells induces cell survival and chemoresistance. J Clin Invest 120:1285–1297
Spranger S, Javorovic M, Burdek M, Wilde S, Mosetter B, Tippmer S et al (2010) Generation of Th1-polarizing dendritic cells using the TLR7/8 agonist CL075. J Immunol 185:738–747
Pufnock JS, Cigal M, Rolczynski LS, Andersen-Nissen E, Wolfl M, McElrath MJ et al (2011) Priming CD8+ T cells with dendritic cells matured using TLR4 and TLR7/8 ligands together enhances generation of CD8+ T cells retaining CD28. Blood 117:6542–6551
Zobywalski A, Javorovic M, Frankenberger B, Pohla H, Kremmer E, Bigalke I et al (2007) Generation of clinical grade dendritic cells with capacity to produce biologically active IL-12p70. J Transl Med 5:18
Shackleton M, Davis ID, Hopkins W, Jackson H, Dimopoulos N, Tai T et al (2004) The impact of imiquimod, a Toll-like receptor-7 ligand (TLR7L), on the immunogenicity of melanoma peptide vaccination with adjuvant Flt3 ligand. Cancer Immun 4:9
Prins RM, Craft N, Bruhn KW, Khan-Farooqi H, Koya RC, Stripecke R et al (2006) The TLR-7 agonist, imiquimod, enhances dendritic cell survival and promotes tumor antigen-specific T cell priming: relation to central nervous system antitumor immunity. J Immunol 176:157–164
Ma F, Zhang J, Zhang J, Zhang C (2010) The TLR7 agonists imiquimod and gardiquimod improve DC-based immunotherapy for melanoma in mice. Cell Mol Immunol 7:381–388
Mahnke K, Qian Y, Fondel S, Brueck J, Becker C, Enk AH (2005) Targeting of antigens to activated dendritic cells in vivo cures metastatic melanoma in mice. Cancer Res 65:7007–7012
Bonifaz L, Bonnyay D, Mahnke K, Rivera M, Nussenzweig MC, Steinman RM (2002) Efficient targeting of protein antigen to the dendritic cell receptor DEC-205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8+ T cell tolerance. J Exp Med 196:1627–1638
Adams S, O’Neill DW, Nonaka D, Hardin E, Chiriboga L, Siu K et al (2008) Immunization of malignant melanoma patients with full-length NY-ESO-1 protein using TLR7 agonist imiquimod as vaccine adjuvant. J Immunol 181:776–784
Edwards AD, Diebold SS, Slack EM, Tomizawa H, Hemmi H, Kaisho T et al (2003) Toll-like receptor expression in murine DC subsets: lack of TLR7 expression by CD8 alpha+ DC correlates with unresponsiveness to imidazoquinolines. Eur J Immunol 33:827–833
Pedersen G, Andresen L, Matthiessen MW, Rask-Madsen J, Brynskov J (2005) Expression of Toll-like receptor 9 and response to bacterial CpG oligodeoxynucleotides in human intestinal epithelium. Clin Exp Immunol 141:298–306
Platz J, Beisswenger C, Dalpke A, Koczulla R, Pinkenburg O, Vogelmeier C et al (2004) Microbial DNA induces a host defense reaction of human respiratory epithelial cells. J Immunol 173:1219–1223
Lebre MC, van der Aar AM, van Baarsen L, van Capel TM, Schuitemaker JH, Kapsenberg ML et al (2007) Human keratinocytes express functional Toll-like receptor 3, 4, 5, and 9. J Invest Dermatol 127:331–341
Hemmi H, Takeuchi O, Kawai T, Kaisho T, Sato S, Sanjo H et al (2000) A Toll-like receptor recognizes bacterial DNA. Nature 408:740–745
Copin R, De BP, Carlier Y, Letesson JJ, Muraille E (2007) MyD88-dependent activation of B220-CD11b+LY-6C+ dendritic cells during Brucella melitensis infection. J Immunol 178:5182–5191
Lee KS, Scanga CA, Bachelder EM, Chen Q, Snapper CM (2007) TLR2 synergizes with both TLR4 and TLR9 for induction of the MyD88-dependent splenic cytokine and chemokine response to Streptococcus pneumoniae. Cell Immunol 245:103–110
Bafica A, Scanga CA, Feng CG, Leifer C, Cheever A, Sher A (2005) TLR9 regulates Th1 responses and cooperates with TLR2 in mediating optimal resistance to Mycobacterium tuberculosis. J Exp Med 202:1715–1724
Anderson AE, Worku ML, Khamri W, Bamford KB, Walker MM, Thursz MR (2007) TLR9 polymorphisms determine murine lymphocyte responses to Helicobacter: results from a genome-wide scan. Eur J Immunol 37:1548–1561
Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A (2003) Toll-like receptor 9-mediated recognition of herpes simplex virus-2 by plasmacytoid dendritic cells. J Exp Med 198:513–520
Sasai M, Linehan MM, Iwasaki A (2010) Bifurcation of Toll-like receptor 9 signaling by adaptor protein 3. Science 329:1530–1534
Krug A, French AR, Barchet W, Fischer JA, Dzionek A, Pingel JT et al (2004) TLR9-dependent recognition of MCMV by IPC and DC generates coordinated cytokine responses that activate antiviral NK cell function. Immunity 21:107–119
Zhu J, Huang X, Yang Y (2007) Innate immune response to adenoviral vectors is mediated by both Toll-like receptor-dependent and -independent pathways. J Virol 81:3170–3180
Fathallah I, Parroche P, Gruffat H, Zannetti C, Johansson H, Yue J et al (2010) EBV latent membrane protein 1 is a negative regulator of TLR9. J Immunol 185:6439–6447
Andersen JM, Al-Khairy D, Ingalls RR (2006) Innate immunity at the mucosal surface: role of toll-like receptor 3 and toll-like receptor 9 in cervical epithelial cell responses to microbial pathogens. Biol Reprod 74:824–831
Kandimalla ER, Bhagat L, Li Y, Yu D, Wang D, Cong YP et al (2005) Immunomodulatory oligonucleotides containing a cytosine-phosphate-2′-deoxy-7-deazaguanosine motif as potent toll-like receptor 9 agonists. Proc Natl Acad Sci USA 102:6925–6930
Wakefield D, Gray P, Chang J, Di GN, McCluskey P (2010) The role of PAMPs and DAMPs in the pathogenesis of acute and recurrent anterior uveitis. Br J Ophthalmol 94:271–274
Rubartelli A, Lotze MT (2007) Inside, outside, upside down: damage-associated molecular-pattern molecules (DAMPs) and redox. Trends Immunol 28:429–436
Garg AD, Nowis D, Golab J, Vandenabeele P, Krysko DV, Agostinis P (2010) Immunogenic cell death, DAMPs and anticancer therapeutics: an emerging amalgamation. Biochim Biophys Acta 1805:53–71
Carta S, Castellani P, Delfino L, Tassi S, Vene R, Rubartelli A (2009) DAMPs and inflammatory processes: the role of redox in the different outcomes. J Leukoc Biol 86:549–555
Gilliet M, Lande R (2008) Antimicrobial peptides and self-DNA in autoimmune skin inflammation. Curr Opin Immunol 20:401–407
Vollmer J, Weeratna R, Payette P, Jurk M, Schetter C, Laucht M et al (2004) Characterization of three CpG oligodeoxynucleotide classes with distinct immunostimulatory activities. Eur J Immunol 34:251–262
Krieg AM, Yi AK, Matson S, Waldschmidt TJ, Bishop GA, Teasdale R et al (1995) CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 374:546–549
Brinkmann MM, Spooner E, Hoebe K, Beutler B, Ploegh HL, Kim YM (2007) The interaction between the ER membrane protein UNC93B and TLR3, 7, and 9 is crucial for TLR signaling. J Cell Biol 177:265–275
Tabeta K, Hoebe K, Janssen EM, Du X, Georgel P, Crozat K et al (2006) The Unc93b1 mutation 3d disrupts exogenous antigen presentation and signaling via Toll-like receptors 3, 7 and 9. Nat Immunol 7:156–164
Kim YM, Brinkmann MM, Paquet ME, Ploegh HL (2008) UNC93B1 delivers nucleotide-sensing toll-like receptors to endolysosomes. Nature 452:234–238
Sepulveda FE, Maschalidi S, Colisson R, Heslop L, Ghirelli C, Sakka E et al (2009) Critical role for asparagine endopeptidase in endocytic Toll-like receptor signaling in dendritic cells. Immunity 31:737–748
Ewald SE, Lee BL, Lau L, Wickliffe KE, Shi GP, Chapman HA et al (2008) The ectodomain of Toll-like receptor 9 is cleaved to generate a functional receptor. Nature 456:658–662
Park B, Brinkmann MM, Spooner E, Lee CC, Kim YM, Ploegh HL (2008) Proteolytic cleavage in an endolysosomal compartment is required for activation of Toll-like receptor 9. Nat Immunol 9:1407–1414
Ewald SE, Engel A, Lee J, Wang M, Bogyo M, Barton GM (2011) Nucleic acid recognition by Toll-like receptors is coupled to stepwise processing by cathepsins and asparagine endopeptidase. J Exp Med 208:643–651
Kemp TJ, Elzey BD, Griffith TS (2003) Plasmacytoid dendritic cell-derived IFN-alpha induces TNF-related apoptosis-inducing ligand/Apo-2L-mediated antitumor activity by human monocytes following CpG oligodeoxynucleotide stimulation. J Immunol 171:212–218
Bekeredjian-Ding I, Jego G (2009) Toll-like receptors—sentries in the B-cell response. Immunology 128:311–323
Hanten JA, Vasilakos JP, Riter CL, Neys L, Lipson KE, Alkan SS et al (2008) Comparison of human B cell activation by TLR7 and TLR9 agonists. BMC Immunol 9:39
Bernasconi NL, Traggiai E, Lanzavecchia A (2002) Maintenance of serological memory by polyclonal activation of human memory B cells. Science 298:2199–2202
Eckl-Dorna J, Batista FD (2009) BCR-mediated uptake of antigen linked to TLR9 ligand stimulates B-cell proliferation and antigen-specific plasma cell formation. Blood 113:3969–3977
Holtick U, Scheulen ME, von Bergwelt-Baildon MS, Weihrauch MR (2011) Toll-like receptor 9 agonists as cancer therapeutics. Expert Opin Investig Drugs 20:361–372
Goodchild A, Nopper N, Craddock A, Law T, King A, Fanning G et al (2009) Primary leukocyte screens for innate immune agonists. J Biomol Screen 14:723–730
Agrawal S, Kandimalla ER (2007) Synthetic agonists of Toll-like receptors 7, 8 and 9. Biochem Soc Trans 35:1461–1467
Krieg AM (2008) Toll-like receptor 9 (TLR9) agonists in the treatment of cancer. Oncogene 27:161–167
Rakoff-Nahoum S, Medzhitov R (2009) Toll-like receptors and cancer. Nat Rev Cancer 9:57–63
van Ojik HH, Bevaart L, Dahle CE, Bakker A, Jansen MJ, van Vugt MJ et al (2003) CpG-A and B oligodeoxynucleotides enhance the efficacy of antibody therapy by activating different effector cell populations. Cancer Res 63:5595–5600
Schmidt C (2007) Clinical setbacks for toll-like receptor 9 agonists in cancer. Nat Biotechnol 25:825–826
Brody JD, Ai WZ, Czerwinski DK, Torchia JA, Levy M, Advani RH et al (2010) In situ vaccination with a TLR9 agonist induces systemic lymphoma regression: a phase I/II study. J Clin Oncol 28:4324–4332
Kochling J, Prada J, Bahrami M, Stripecke R, Seeger K, Henze G et al (2008) Anti-tumor effect of DNA-based vaccination and dSLIM immunomodulatory molecules in mice with Ph+ acute lymphoblastic leukaemia. Vaccine 26:4669–4675
Vaisanen MR, Vaisanen T, Jukkola-Vuorinen A, Vuopala KS, Desmond R, Selander KS et al (2010) Expression of toll-like receptor-9 is increased in poorly differentiated prostate tumors. Prostate 70:817–824
Gonzalez-Reyes S, Marin L, Gonzalez L, Gonzalez LO, del Casar JM, Lamelas ML et al (2010) Study of TLR3, TLR4 and TLR9 in breast carcinomas and their association with metastasis. BMC Cancer 10:665
Kuninaka N, Kurata M, Yamamoto K, Suzuki S, Umeda S, Kirimura S et al (2010) Expression of Toll-like receptor 9 in bone marrow cells of myelodysplastic syndromes is down-regulated during transformation to overt leukemia. Exp Mol Pathol 88:293–298
Daud II, Scott ME, Ma Y, Shiboski S, Farhat S, Moscicki AB (2011) Association between toll-like receptor expression and human papillomavirus type 16 persistence. Int J Cancer 128:879–886
Chiron D, Bekeredjian-Ding I, Pellat-Deceunynck C, Bataille R, Jego G (2008) Toll-like receptors: lessons to learn from normal and malignant human B cells. Blood 112:2205–2213
Yu L, Chen S (2008) Toll-like receptors expressed in tumor cells: targets for therapy. Cancer Immunol Immunother 57:1271–1278
Gekeler V, Gimmnich P, Hofmann HP, Grebe C, Rommele M, Leja A et al (2006) G3139 and other CpG-containing immunostimulatory phosphorothioate oligodeoxynucleotides are potent suppressors of the growth of human tumor xenografts in nude mice. Oligonucleotides 16:83–93
Wang H, Rayburn ER, Wang W, Kandimalla ER, Agrawal S, Zhang R (2006) Chemotherapy and chemosensitization of non-small cell lung cancer with a novel immunomodulatory oligonucleotide targeting Toll-like receptor 9. Mol Cancer Ther 5:1585–1592
Ilvesaro JM, Merrell MA, Swain TM, Davidson J, Zayzafoon M, Harris KW et al (2007) Toll like receptor-9 agonists stimulate prostate cancer invasion in vitro. Prostate 67:774–781
El AA, Sonabend AM, Han Y, Lesniak MS (2006) Stimulation of TLR9 with CpG ODN enhances apoptosis of glioma and prolongs the survival of mice with experimental brain tumors. Glia 54:526–535
Fischer SF, Rehm M, Bauer A, Hofling F, Kirschnek S, Rutz M et al (2005) Toll-like receptor 9 signaling can sensitize fibroblasts for apoptosis. Immunol Lett 97:115–122
Jozsef L, Khreiss T, Filep JG (2004) CpG motifs in bacterial DNA delay apoptosis of neutrophil granulocytes. FASEB J 18:1776–1778
Chang YJ, Wu MS, Lin JT, Chen CC (2005) Helicobacter pylori-induced invasion and angiogenesis of gastric cells is mediated by cyclooxygenase-2 induction through TLR2/TLR9 and promoter regulation. J Immunol 175:8242–8252
Damiano V, Caputo R, Bianco R, D’Armiento FP, Leonardi A, De PS et al (2006) Novel toll-like receptor 9 agonist induces epidermal growth factor receptor (EGFR) inhibition and synergistic antitumor activity with EGFR inhibitors. Clin Cancer Res 12:577–583
Nierkens S, den Brok MH, Garcia Z, Togher S, Wagenaars J, Wassink M et al (2011) Immune adjuvant efficacy of CpG oligonucleotide in cancer treatment is founded specifically upon TLR9 function in plasmacytoid dendritic cells. Cancer Res 71(20):6428–6437
Labidi-Galy SI, Sisirak V, Meeus P, Gobert M, Treilleux I, Bajard A et al (2011) Quantitative and functional alterations of plasmacytoid dendritic cells contribute to immune tolerance in ovarian cancer. Cancer Res 71:5423
Hartmann E, Wollenberg B, Rothenfusser S, Wagner M, Wellisch D, Mack B et al (2003) Identification and functional analysis of tumor-infiltrating plasmacytoid dendritic cells in head and neck cancer. Cancer Res 63:6478–6487
Perrot I, Blanchard D, Freymond N, Isaac S, Guibert B, Pacheco Y et al (2007) Dendritic cells infiltrating human non-small cell lung cancer are blocked at immature stage. J Immunol 178:2763–2769
Dolganiuc A, Chang S, Kodys K, Mandrekar P, Bakis G, Cormier M et al (2006) Hepatitis C virus (HCV) core protein-induced, monocyte-mediated mechanisms of reduced IFN-alpha and plasmacytoid dendritic cell loss in chronic HCV infection. J Immunol 177:6758–6768
Conry SJ, Milkovich KA, Yonkers NL, Rodriguez B, Bernstein HB, Asaad R et al (2009) Impaired plasmacytoid dendritic cell (PDC)-NK cell activity in viremic human immunodeficiency virus infection attributable to impairments in both PDC and NK cell function. J Virol 83:11175–11187
Ulsenheimer A, Gerlach JT, Jung MC, Gruener N, Wachtler M, Backmund M et al (2005) Plasmacytoid dendritic cells in acute and chronic hepatitis C virus infection. Hepatology 41:643–651
Kang DC, Gopalkrishnan RV, Lin L, Randolph A, Valerie K, Pestka S et al (2004) Expression analysis and genomic characterization of human melanoma differentiation associated gene-5, mda-5: a novel type I interferon-responsive apoptosis-inducing gene. Oncogene 23:1789–1800
Kovacsovics M, Martinon F, Micheau O, Bodmer JL, Hofmann K, Tschopp J (2002) Overexpression of Helicard, a CARD-containing helicase cleaved during apoptosis, accelerates DNA degradation. Curr Biol 12:838–843
Tormo D, Checinska A, Alonso-Curbelo D, Perez-Guijarro E, Canon E, Riveiro-Falkenbach E et al (2009) Targeted activation of innate immunity for therapeutic induction of autophagy and apoptosis in melanoma cells. Cancer Cell 16:103–114
Poeck H, Besch R, Maihoefer C, Renn M, Tormo D, Morskaya SS et al (2008) 5′-Triphosphate-siRNA: turning gene silencing and Rig-I activation against melanoma. Nat Med 14:1256–1263
Besch R, Poeck H, Hohenauer T, Senft D, Hacker G, Berking C et al (2009) Proapoptotic signaling induced by RIG-I and MDA-5 results in type I interferon-independent apoptosis in human melanoma cells. J Clin Invest 119:2399–2411
Kubler K, Pesch CT, Gehrke N, Riemann S, Dassler J, Coch C et al (2011) Immunogenic cell death of human ovarian cancer cells induced by cytosolic poly(I:C) leads to myeloid cell maturation and activates NK cells. Eur J Immunol 41(10):3028–3039
Liu TX, Zhang JW, Tao J, Zhang RB, Zhang QH, Zhao CJ et al (2000) Gene expression networks underlying retinoic acid-induced differentiation of acute promyelocytic leukemia cells. Blood 96:1496–1504
Jiang LJ, Zhang NN, Ding F, Li XY, Chen L, Zhang HX et al (2011) RA-inducible gene-I induction augments STAT1 activation to inhibit leukemia cell proliferation. Proc Natl Acad Sci USA 108:1897–1902
Conflict of interest
No funds were received in support of this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Goutagny, N., Estornes, Y., Hasan, U. et al. Targeting pattern recognition receptors in cancer immunotherapy. Targ Oncol 7, 29–54 (2012). https://doi.org/10.1007/s11523-012-0213-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11523-012-0213-1