
Abstract
Plants produce a wide spectrum of secondary metabolites that play critical roles in plant-environment interactions and against biotic and abiotic stresses. Moreover, many secondary metabolites have pharmaceutical efficacy for a wide range of diseases (cancer, malaria, etc.). Controlled transcription of biosynthetic genes is one of the major mechanisms regulating secondary metabolism in plants. Several transcription factor families such as MYC, MYB, WRKY and AP2/ERF have been found to be involved in the regulation of secondary metabolism in different medicinal plants. In addition, the biosynthesis and proper accumulation of secondary metabolites are also induced by signaling molecule jasmonic acid (JA). This review provides an insight into JA signaling pathway and JA-mediated transcriptional regulation of secondary metabolism (vinblastine, nicotine, artemisinin, taxol and ginsenoside) in a range of medicinal plant species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Plants produce a variety of low molecular weight organic compounds that are usually ramified into two large classes: primary and secondary metabolites. Primary metabolites are essential for plant growth and development, while secondary metabolites act as defense molecules and protect plants in various adverse conditions. Many secondary metabolites have high pharmaceutical value and have been used for the treatment of wide range of diseases [1]. For example, vinblastine and nicotine are used for treatment of tumor [2] and asthma [3] diseases, respectively. Artemisinin, another essential secondary metabolite, is extensively employed as antimalarial drug [4]. Furthermore, two additional pharmaceutically important secondary metabolites, taxol and ginsenosides, are generally used for the treatment of breast, ovarian and non-small cell lung cancers [5, 6]. Because of their beneficial effects to health, many researchers have been focusing on the comprehensive study of regulation and production of secondary metabolites. The biosynthesis and proper accumulation of secondary metabolites are strictly controlled in a spatial and temporal manner and influenced by a number of biotic and abiotic factors. The spatiotemporal transcriptional regulation of metabolic pathways is controlled by a complex network involving many regulatory proteins known as transcription factors (TFs). TFs are sequence-specific DNA-binding proteins that interact with the regulatory regions of the target genes and modulate the rate of transcriptional initiation by RNA polymerase [7]. Many TFs have been characterized for their roles in regulating biosynthetic pathways at the transcriptional level. TFs include a wide number of proteins that initiate and regulate the transcription of several genes. These proteins regulate gene transcription depending on tissue type and in response to external and internal signals [8]. Jasmonic acid (JA) that presents in the scent of jasmine flowers (Jasminum grandiflorum) is known as an essential signaling molecule which can coordinate a lot of cellular activities [9]. JA and its cyclic precursors and derivatives (such as methyl jasmonate, MeJA) are collectively referred to as jasmonates (JAs). JAs act as ubiquitous and conserved elicitors for the production of secondary metabolites across the plant kingdom, from gymnosperms to angiosperms [10, 11]. The promising depiction of transcriptional regulation of secondary metabolism suggests that, in response to phytohormones (JAs), TFs form dynamic regulatory networks that fine-tune the timing, amplitude and tissue-specific expression of pathway genes and the subsequent accumulation of secondary metabolites [8]. Here, we give an updated review on JA-responsive signaling pathway and speculate on JA-mediated transcriptional regulation of secondary metabolism (e.g., vinblastine, nicotine, artemisinin, taxol and ginsenoside) in a range of medicinal plant species.
2 Key players in the JA-mediated signaling pathway
JAs are oxylipin-derived phytohormones that regulate a wide variety of physiological processes in plants including growth, development, reproduction and defense [12]. JAs are synthesized from the fatty acid α-linolenic acid via the octadecanoid pathway [13]. In general, the JAs signaling pathway is accomplished in two steps.
2.1 Formation of SCFCOI1 complex and degradation of JAZ proteins by 26S proteasome
In JA signaling pathway, the F-box protein CORONATINE INSENSITIVE 1 (COI1) connects with Skp–Cullin–F-box-type E3 ubiquitin ligase and forms SCFCOI1 complex to which it provides substrate specificity [10, 12]. The targets of the SCFCOI1 complex are the JA ZIM domain (JAZ) family of repressor proteins. JAZ is one of the key molecules in this pathway that serves as the on/off switch for JA signaling. JAZ and COI1 proteins directly interact in the presence of the bioactive JA–isoleucine (JA-Ile) conjugate to form a co-receptor complex. Although ubiquitination of the JAZ proteins also joins with SCFCOI1, this interaction ultimately triggers the degradation of the JAZ proteins by the 26S proteasome (Fig. 1) [9, 14]. The JAZ proteins contain a conserved TIFY motif within the ZIM domain that mediates homo- and hetero-dimeric interactions between different JAZ proteins. The ZIM domain also functions to recruit transcriptional corepressors, such as TOPLESS (TPL), through the novel interactor of JAZ (NINJA) protein. The JAZ proteins are further characterized by a Jas domain that is required for the interaction of both COI1 and a broad array of TFs [14, 15].

Jasmonic acid (JA) signaling pathway. In the presence of JA–isoleucine (JA-Ile), CORONATINE INSENSITIVE 1 (COI1) binds to Skp–Cullin–F-box-type E3 ubiquitin ligase and form SCFCOI1 complex, thereby recruiting the JAZ proteins, targeting them for their degradation by the 26S proteasome and ultimately leading to the release of the TFs that can modulate JA-responsive gene expressions. COI1, CORONATINE INSENSITIVE 1, JA jasmonic acid, JA-Ile JA–isoleucine, JAZ JA ZIM domain, SCF Skp–Cullin–F-box-type E3 ubiquitin ligase, TFs transcription factors
2.2 Transcriptional coordination and enhance physiological activity
In the presence of JA or its bioactive derivatives, JAZ proteins are degraded and freeing TFs for expression of specific sets of JA-responsive genes, thereby promoting physiological activity [9], such as produce specific sets of secondary metabolites (Fig. 1). However, in the absence of JA, JAZ proteins bind to downstream TFs and limit their activity [16]. The best-studied TFs acting in this pathway belong to the MYC family (of which there are three, MYC2, 3 and 4) that characterized by a basic helix-loop-helix (bHLH) DNA-binding motif. For example, in Arabidopsis thaliana, the bHLH TF MYC2 has been shown to involve in regulating secondary metabolite induction both directly and indirectly. It positively regulates TFs and biosynthetic enzymes of flavonoid biosynthesis, but negatively controls tryptophan-derived indole glucosinolate biosynthesis [17]. Related bHLH TFs MYC3 and MYC4 also regulate both overlapping and distinct MYC2-regulated functions in Arabidopsis, while MYC2 orthologs act as “master switches” that regulate JA-mediated biosynthesis of secondary metabolites [18]. In various medicinal plant species, the MYC2 is also well-known target of the JAZ proteins. For example, MYC2 homolog regulates the expression of the ORCA TFs by direct binding to the promoter of the ORCA3 gene, thereby controlling expression of several terpenoid indole alkaloid (TIA) biosynthesis genes in Catharanthus roseus [19]. In Nicotiana tabacum, MYC2 proteins up-regulate the ORCA-related NIC2 locus Apetala2/ethylene response factor (AP2/ERF) that regulates nicotine biosynthesis [20, 21]. In the related species Nicotiana benthamiana, the MYC2 homologs (NbbHLH1 and NbbHLH2) also play as positive regulators in the JA-mediated activation of nicotine biosynthesis [22]. JAZ proteins also interact with other TFs with an embedded role in the biosynthesis of secondary metabolites, such as the R2R3-MYB TFs [23]. A number of TFs with a proven role in JA-mediated elicitation of a specific metabolic pathway are given in Table 1. In some cases, two or more TFs combine together and regulate diverse secondary metabolites such as bHLH and R2R3-MYB TFs which together compose transcriptional activator complexes that control anthocyanin biosynthesis in Arabidopsis and other plant species [24]. Based on these evidence, it is intriguing to recommend that many TFs are highly conserved and play a central role in the JA-mediated signaling pathway.
3 Transcriptional regulation of secondary metabolism in medicinal plants
Transcriptional regulation is the change in gene expression levels by altering transcription rates. Regulation of transcription is depending on the combined effects of structural properties and the interactions of TFs. The expression of genes encoding enzymes involved in one particular secondary metabolic pathway often displayed a marked concerted up-regulation after JA elicitation, leading to the recognition of so-called transcriptional regulons. Several genome-wide transcript profiling studies have demonstrated that JAs treatment triggers an extensive transcriptional reprogramming of metabolism [10]. Several TFs that can be elicited by JAs and regulate specific metabolic pathway are given in Table 1.
3.1 Vinblastine
The plant Madagascar periwinkle (C. roseus), a member of the Apocynaceae family, produces a large array of TIAs. TIAs can be characterized into two principle groups: monoterpene indole alkaloids (MIAs) and the bisindole alkaloids that composed of two joined MIAs. These TIAs, either natural or semisynthetic forms, have pharmaceutical applications. For example, the MIAs serpentine and ajmalicine are used as a tranquillizer and to reduce hypertension, respectively, and the bisindole alkaloids vincristine and vinblastine are potent as antitumor drugs [2, 42]. Moreover, vinblastine also shows a strong antimicrobial activity [43]. The large interest in the anticancer compound vinblastine, which is derived from the coupling of catharanthine and vindoline, contrasts with the low amounts of these compounds found in the plants, making its extraction a very expensive process [44]. The formation of vindoline requires deacetylvindoline 4-O-acetyltransferase (DAT). DAT transfers an acetyl group on to deacetylvindoline to produce vindoline. JA has been demonstrated as regulators of DAT expression [45]. Production of vinblastine through TIA pathway is notably complex, and several key committed steps have been the primary targets for transcriptional regulation. The biosynthesis of TIAs involves a large number of enzymes. Strictosidine is the central intermediate in the biosynthesis of different TIAs (including vinblastine), which is formed by the condensation of secologanin and tryptamine. In fact, the transcript level of STR is induced by JA treatment [46, 47]. Most of genes codifying enzymes from TIA pathway are tightly regulated by specific TFs in a coordinate manner together with developmental and environmental factors. In C. roseus, only a few TFs have been isolated and characterized, highlighting the CrORCA2, CrORCA3, CrBPF1, CrWRKY1, CrMYC1 and CrMYC2. These TFs respond to JAs and/or other elicitors [46, 48–50]. Two bHLH TFs, CrMYC1 and CrMYC2, were isolated and characterized from C. roseus, both of which were involved in the response to JA [19, 25], suggesting that these (CrMYC1 and CrMYC2) TFs may be involved in the regulation of TIA biosynthesis in response to JA signals. Moreover, CrMYC2 activates the expression of two TFs, ORCA2 and ORCA3 (Fig. 2), which is the members of the AP2/ERF-domain class. The AP2/ERF TFs form large families unique to plants and are characterized by the AP2/ERF DNA-binding domain [19]. Both ORCA2 and ORCA3 bind to the JERE (a cis-acting element involved in JA and elicitor responses) element of the STR promoter. The JRE is composed of an A/T-rich quantitative sequence responsible for high-level expression and a qualitative component with a T/G-box element acting as an on/off switch in response to JAs. Both ORCA2 and ORCA3 positively respond to JAs [51–53]. ORCA2 is a TF key in regulating genes encoding the enzymes STR, tryptophan decarboxylase (TDC) and strictosidine β-d-glucosidase (SGD) involved in the TIA pathway [53, 54]. On the other hand, ORCA3 overexpression resulted in increasing levels of some genes, specially TDC, STR and desacetoxyvindoline-4-hydroxylase (D4H) [55, 56], which, in turn, provokes the accumulation of vinblastine and other secondary metabolites in TIA pathway [57, 58].

JA-mediated transcriptional regulation of vinblastine synthesis in Catharanthus roseus and nicotine synthesis in Nicotiana tabacum. Solid and dashed arrows indicate proven and hypothetical (yet to be experimentally established) links, respectively. Arrows indicate positive interactions and T-bars indicate negative interactions. COI1 CORONATINE INSENSITIVE 1, ERF ethylene response factor, JA jasmonic acid, JA-Ile JA–isoleucine; JAZ jasmonic acid ZIM domain, JAM1 JA factor-stimulating MAPKK 1, ORCA 2/3 octadecanoid-responsive Catharanthus AP2-domain proteins 2 and 3, SCF Skp–Cullin–F-box-type E3 ubiquitin ligase, ZCT zinc-finger Catharanthus transcription factor
Other TF involved in the biosynthesis of TIAs is the WRKY family. Recently, a WRKY TF, CrWRKY1, isolated from Catharanthus seedlings has been shown to play a crucial role in TIA biosynthesis and induced by JAs [28]. Overexpression of CrWRKY1 in Catharanthus hairy roots results in up-regulation of several TIA pathway genes, especially TDC. CrWRKY1 binds the W-box motif in the TDC promoter and trans-activates the TDC promoter in Catharanthus cells [48, 59]. Depiction of the CrWRKY1 promoter revealed the importance of several cis-elements, including the TGACG motifs, and suggests the involvement of novel TFs in this regulatory network [60, 61]. These findings demonstrate that the preferential expression of CrWRKY1 and its interaction with other TFs (including CrORCAs and CrMYCs) play a key role in determining the accumulation of vinblastine in C. roseus.
3.2 Nicotine
Nicotiana, a member of the Solanaceae family, is one of the most important research model plants and of high economic value worldwide. The main secondary metabolites found in Nicotiana species are tobacco alkaloids [62]. The biosynthesis of nicotine involves pyrrolidine ring formation, pyridine ring formation and the coupling of both rings. Several enzymes and corresponding genes are involved in the major steps of the nicotine synthetic pathway [63]. In N. tabacum, arginine decarboxylase (ADC), ornithine decarboxylase (ODC), putrescine N-methyltransferase (PMT) and N-methyl-putrescine oxidase (MPO) are involved in the formation of the pyrrolidine ring; on the other hand, aspartate oxidase (AO), quinolinate synthase (QS) and quinolinic acid phosphoribosyltransferase (QPT) are responsible for the biosynthesis of the pyridine ring [64]. Furthermore, a PIP (pinoresinol–lariciresinol reductase, isoflavone reductase and phenylcoumaran benzylic ether reductase) family isoflavone reductase-like protein A622 and berberine bridge enzyme-like protein (BBL) are required for nicotine ring coupling [62]. However, it remains unclear how the two rings are joined to form nicotine. Data from a range of studies revealed that the complex regulation of nicotine biosynthesis involves a number of TFs and JAs is a positive regulator for nicotine biosynthesis [62, 64, 65]. In response to JA, the JA receptor NtCOI1 mediates degradation of the NtJAZs protein, a transcriptional repressor, resulting in inducible expression of PMT (Fig. 2) [36]. In N. benthamiana, two bHLH TFs bHLH1 and bHLH2, which are homologs of MYC2, bind to the PMT promoter to provide positive regulation of nicotine synthesis. A series of ERF TFs that regulate nicotine levels play a role in regulating nicotine content [22]. In N. tabacum, 7 AP2/ERF TFs were found in the NIC2 locus [36].The JA-inducible ORC1 (also known as ERF221) and ERF189 TFs cluster together in the NIC2 locus and are up-regulated by JAs elicitation. ORC1 and ERF189 have overlapping but non-redundant roles in regulating nicotine biosynthetic genes [36, 66]. ORC1 recognition of the GCC-box of the PMT promoter was necessary but not sufficient for activation. ORC1 is a close homolog of C. roseus ORCA3, and its overexpression stimulated alkaloid biosynthesis [66]. NtMYC2 directly regulates the expression of selected nicotine biosynthetic genes and also regulates nicotine biosynthesis through regulation of the NIC2 AP2/ERF TFs [20, 67]. In N. benthamiana, MYC2 homolog NbbHLH1 functions in combination with ORC1, in binding to the G and GCC-boxes of PMT1 and quinolinate phosphoribosyl transferase (QPRT2) promoters, respectively; both TFs are necessary for optimal PMT1 and QPRT2 activation [66]. Future work is needed to determine the roles of the other five NIC2 locus ERFs specifically play in regulating nicotine biosynthesis. In addition, JA-induced phosphorylation cascade has also been shown to play a crucial role in nicotine biosynthesis; for example, the activity of both ORC1 and the MYC-type bHLH proteins can be posttranslationally up-regulated by a JA-modulated phosphorylation cascade. By contrast, in tobacco, the JA-inducible MYBJS1 (a R2R3-MYB TF) was shown to induce phenylpropanoid biosynthetic genes, and the accumulation of phenylpropanoid polyamine conjugates during stress [35]. A related MYB, MYB8, was recently found to control inducible phenolamide levels in Nicotiana attenuata [37]. Collectively, these findings indicate the intricate interaction between different TFs in the regulation of nicotine and other secondary metabolites in Nicotiana species.
3.3 Artemisinin
Artemisinin, a sesquiterpene lactone endoperoxide that isolated from Artemisia annua (family Asteraceae), has been widely employed as antimalarial drugs [4, 30]. Artemisinin is effective against chloroquine- and quinine-resistant Plasmodium falciparum as well as other malaria-causing parasites [68]. Besides their antimalarial activity, recently artemisinins have also been reported to possess antiviral [31], anticancer [32] and antischistosomal activities [69]. To date, A. annua is the only natural source of artemisinin. The relatively low yield (0.1 %–0.8 % by dry weight) of artemisinin in A. annua is a major limitation to the commercialization of the drug, so the enhanced production of artemisinin is highly desirable [70]. Caretto et al. [71] have recognized that the plant A. annua are able to produce high content of artemisinin in respond to the elicitor effect of JAs. So, controlled transcription of biosynthetic genes by JA-responsive TFs is an alternative way to produce high artemisinin from A. annua. Recently, the artemisinin biosynthetic pathway has been almost completely revealed by several groups [72]. In artemisinin biosynthesis pathway, amorpha-4,11-diene synthase (ADS) catalyzes the conversion of farnesyl diphosphate into amorpha-4,11-diene, which is the first committed step in A. annua [73]. In the second step, amorpha-4,11-diene is hydroxylated to yield artemisinic alcohol, which is subsequently oxidized to form artemisinic aldehyde. The two steps are catalyzed by a bifunctional cytochrome P450-dependent hydroxylase (CYP71AV1) together with the NADPH: cytochrome P450 oxidoreductase (CPR) [74]. Most of the genes involved in the artemisinin biosynthetic pathway are likely to be trichome specific and to be controlled by one or several TFs [75]. Two jasmonate-responsive AP2/ERF TFs AaERF1 and AaERF2 can regulate the transcription of ADS and CYP sesquiterpene oxidase (CYP71AV1) and to activate expression of both genes. Overexpression of AaERF1 or AaERF2 led to increased accumulation of artemisinin and artemisinic acids [30]. The promoters of ADS contain two reverse-oriented TTGACC W-box cis-acting elements, which are the proposed binding sites of WRKY TFs. Ma et al. [4] first reported about WRKY1 TF in A. annua by isolating full-length cDNA of AaWRKY1. In A. annua, JA-responsive WRKY1 can transactivate the promoter of the ADS gene and might participate in the regulation of artemisinin biosynthesis. Majority of the artemisinin biosynthetic gene expressions were activated by transient expression of AaWRKY1 cDNA [4, 76]. Recently, the AP2/ERF TF AaORA1 has been shown to regulate accumulation of artemisinin [77]. With some exception, AaORA1 regulates the same enzymes as AaWRKY1 that directing the pathway toward artemisinin formation. Moreover, Ji et al. [70] report about the isolation and characterization of novel bHLH TF (AabHLH1) from A. annua. AabHLH1 can positively regulate the transcription of ADS and CYP71AV1 by binding to the E boxes in the promoters, and transient expression of the AabHLH1 TF in leaves of A. annua resulted in increased levels of ADS and CYP71AV1. The previous study indicates that JA-responsive expression of alkaloid biosynthesis genes in C. roseus is controlled by a TF cascade consisting of the bHLH protein, CrMYC2, which regulates the gene expression AP2/ERF-domain transcription factor ORCA (ORCA2 and ORCA3) [54]. It should be interesting to investigate in the future whether there is a TF cascade consisting of AabHLHs regulating AP2/ERF-domain TFs (AaERF1 and AaERF2) in artemisinin biosynthesis in A. annua.
3.4 Taxol
Taxol (generic name paclitaxel), a complex diterpene found in the gymnosperm genus Taxus, is one of the most promising anticancer agents and widely used for the treatment of breast, ovarian and non-small cell lung cancers as well as AIDS-related Kaposi’s sarcoma [5]. Paclitaxel is also being investigated to use in the treatment of neurological disorders and in postsurgery heart patients. Production of taxol from yew trees remains a challenging problem due to the limited resources of Taxus sp. Moreover, paclitaxel found small quantities in plants (0.01 %–0.03 % of the dry weight of the bark of Taxus), and total biosynthesis comprises several steps and is therefore low yielding [39]. The biosynthetic pathway is believed to contain 19 steps; however, there are certain transformations which have yet to be fully characterized [78]. Like other secondary metabolites, JAs can significantly induce the production of taxol and related taxanes in Taxus sp. [39, 79]. The time-course analysis revealed that there are two regulatory steps existing in the paclitaxel biosynthesis: (i) the taxane ring formation step that is up-regulated by JAs and (ii) the acylation step at the C-13 position. The enzyme 10-deacetylbaccatin III-10 β-o-acetyl transferase (DBAT) is a key rate-limiting step in the biosynthesis of taxol [80]. Li et al. [81] identified a WRKY TF, TcWRKY1, with the yeast one-hybrid system from Taxus chinensis cells, as a regulator of DBAT. The gene expression of TcWRKY1 in T. chinensis suspension cells was specifically induced by JAs. Biochemical analysis indicated that TcWRKY1 protein specifically interacts with the two W-box (TGAC) cis-elements among the important regulatory elements. Overexpression of TcWRKY1 enhanced DBAT expression in T. chinensis suspension cells, and RNA interference (RNAi) reduced the transcript levels of DBAT. These results suggest that TcWRKY1 participates in regulation of taxol biosynthesis in T. chinensis cells and DBAT is a target gene of this WRKY TF. Recently, Lenka et al. [39] identified three JA-inducible MYC TFs TcJAMYC1, TcJAMYC2 and TcJAMYC4 from Taxus cuspidata. The MYC TFs from Taxus sp. exhibit high similarity to Arabidopsis MYC2. The similarity in sequence and expression between TcJAMYC(s) and the well-characterized AtMYC2 suggests a conserved response to JAs despite significant divergence between the gymnosperm and angiosperm lineages. The results suggest that TcJAMYC(s) are regulators of JA-mediated expression of taxane biosynthetic genes in Taxus, but that they function as negative rather than positive regulators of these genes. Further analysis of other TFs will be helpful to understand the regulation patterns upon JAs elicitation and the molecular mechanisms of JA-mediated taxane biosynthesis in Taxus sp.
3.5 Ginsenoside
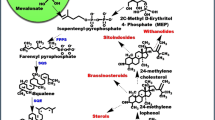
Ginseng (Panax ginseng C.A. Meyer) belonging to the family Araliaceae has been used as a traditional medicine since the ancient times. The major active component of ginseng roots is ginsenoside, a triterpenoid saponin. Saponins have been shown to have many beneficial bioactive effects on human health, such as antitumor, antihyperglycemic, antiinflammatory, antistress, antiaging and improving immune function [82, 83]. To date, more than 40 ginsenosides have been characterized, which can be divided into three categories according to the position of side chains on the shared a glycine skeleton. These three types are the protopanaxadiol (PPD), protopanaxatriol (PPT) and oleanolic acid [83]. Due to their broad pharmaceutical properties, special attention has been paid to genes involved in ginsenoside biosynthesis for large-scale production. Ginsenosides are synthesized by the cyclization of 2,3-oxidosqualene via the mevalonate pathway. This cyclization occurs at the branching point leading to oleanane- and dammarene-type ginsenosides. Dammarene-type triterpenes undergo further modifications such as oxidation/hydroxylation, substitution and glycosylation leading to PPD- and PPT-type ginsenosides. Despite commercial interest in ginsenosides, very little is known about the genes involved in the biosynthetic pathway of triterpenoid saponins [83, 84].
Various elicitation strategies have been developed to stimulate ginsenoside biosynthesis of ginseng plants. For example, JAs treatment can increase the production of ginsenosides. The previous study indicates that JAs (especially MeJA) have a great impact on ginsenoside production in ginseng hairy root and adventitious root cultures. The stimulation of ginsenosides production with JAs may be mediated by the up-regulation of the genes involved in the biosynthesis and accumulation of saponins [85–87]. Recently, Sun et al. [41] isolated one JA-responsive WRKY family TF (PqWRKY1) that positively regulate triterpene ginsenoside biosynthesis in P. quinquefolius. In our work, one MYB TF (PgMYB1) has been isolated from P. ginseng that exhibit negative transcript effect by JAs [88]. However, transcriptional regulation of ginsenoside is little known, and the research in this field will be emerged in near future.
4 JA-responsive transcriptional networks and elicitation of secondary metabolites
Secondary metabolites play diverse roles in plants. Biosynthesis of defensive secondary metabolites is often induced by the following attack of plant. Primary signals specifying attack which are recognized by the plant are called elicitors. These elicitors activate signal transduction pathways that generate secondary signals produced by plants [11]. Three major secondary signaling molecules are JAs, ethylene and salicylic acid [89, 90]. Production of these hormones generates a signal transduction network that leads to a cascade of events responsible for the physiological adaptation of the plant cell to the external stress [91]. Among these three key regulatory signals, by far the most important molecules for induction of secondary metabolism are the JAs. JAs play key roles in defense against insects and certain pathogens, among others by controlling the biosynthesis of protective secondary metabolites [92]. Coordinate transcriptional control of biosynthetic genes emerges as a major mechanism dictating the final levels of secondary metabolites in plant cells. This regulation of biosynthetic pathways is achieved by specific TFs. TFs form useful tools for engineering the production of valuable secondary metabolites [7]. Studies related to the regulation of secondary metabolite pathways have been focused mainly on characterization of TFs regulating the structural genes. However, only a few have provided mechanistic insight about regulation. The bHLH TF MYC2 forms a very valuable tool, since it occupies the highest position in the regulatory hierarchy and it controls the expression of both the ORCA2 and ORCA3 genes and possibly of other genes encoding TFs involved in alkaloid biosynthesis in C. roseus. Activation of CrMYC2 involves JA-induced degradation of JAZ proteins that forms a repressor complex with CrMYC2 in the absence of JA [19]. Similarly, biosynthesis of nicotine is also induced by JA in a COI1-JAZ-dependent manner. Zhang et al. [21] have shown that tobacco contains multiple MYC2 genes (NtMYC2a, NtMYC2b and NtMYC2c) involved in nicotine biosynthesis. The similarities between the components involved in regulation of JA-responsive alkaloid metabolism in tobacco and in C. roseus suggest that the overall regulatory circuits are highly similar (Fig. 2). The above finding indicates that JAZ proteins are conserved components of JAs signaling in plants. However, JA-mediated WRKY TF also acts as a powerful tool for medicinal plants secondary metabolism. In C. roseus, CrWRKY1 interacts with other TFs and helps to accumulate important secondary metabolites [93]. AaWRKY1 and TcWRKY1 also participate in regulation of artemisinin and taxol biosynthesis in A. annua and T. chinensis, respectively [4, 82]. Recently, Sun et al. [41] reported about PqWRKY1, which positively regulates triterpene ginsenoside biosynthesis in P. quinquefolius. These evidences suggest that the concerted transcriptional activation of whole pathways is not necessarily activated by the action of a single TF; quite the opposite, a combinatorial role for several TFs in the regulation of different enzymes or suites of enzymes seems currently more reasonable to clarification for the control of biosynthetic pathways by JAs. Combinatorial exploit of TFs has already been confirmed in non-medicinal secondary metabolite flavonoid biosynthesis [94]. In case of medicinal plants, the JA-mediated elicitation of tobacco nicotine biosynthesis also involves the concerted action of AP2/ERF and bHLH TFs [67]. It implies that combinatorial interactions of TF families are proficient to biosynthesis various important secondary metabolites.
JAs have an evolutionarily conserved role in the reprogramming of plant secondary metabolism in response to various environmental or developmental stimuli, and JA-mediated induction of secondary metabolism is strongly and rapidly influenced by defense responses [95]. When the plant is under attack, the JA signal needs to persist or even to intensify for the induction of secondary metabolite accumulation. As a result, plants have developed a positive response system with loops at various control points, termed as autoregulatory JA loop [96]. The mechanisms involved in the autoregulatory JA loop have mainly been studied in Arabidopsis, but the phenomenon is conserved among plants. Positive and negative feedback loops might be increased the system of secondary metabolism but not at all costs. In case of positive loops, a control point is situated at the genes encoding JAs biosynthesis enzymes that are all JAs inducible and controlled by JA-responsive TFs (such as MYC2), that increase bioactive JAs synthesis [10, 96]. After that, many genes encoding TFs involved in the primary JAs response are rapidly induced by the same signal and can modulate their own expression [10, 17]. Finally, some other sets of regulators and a number of auxin response factors (ARF6 and ARF8) can positively influence JA biosynthesis in a developmental context [97]. In addition to positive loops, negative feedback is also needed to shut down plant defense and stress responses to avoid energy being wasted. Multiple regulatory mechanisms have been developed to maintain energy consumption. As a result, negative regulators are key components in the control of stress-related gene expression. ERF-associated amphiphilic repression (EAR)-domain-containing proteins have been identified as characteristic elements of transcriptional repression of gene expression in plants [18]. Some EAR proteins have been shown to be active in metabolic pathways. In C. roseus, three members of the EAR-domain-containing TF IIIA-type zinc-finger protein family, ZCT1, ZCT2 and ZCT3, were reported to bind the promoters of the STR and TDC genes and therefore repress their expression [29]. Such information can potentially be used to engineer metabolic pathways to enhance the accumulation of important secondary compounds in medicinal plants.
5 Conclusions and perspectives
The plant secondary metabolites play important roles in plant-environment interactions. Also they have been proven to be beneficial for human health, and many secondary metabolites are highly valuable in the pharmacological application. However, direct extraction of these metabolites is usually complicated and inefficient due to their low concentrations in plant tissues. Biosynthesis and regulation of secondary metabolites is multilayer network that requires common signaling pathways and key molecules such as JAs. JAs are instrumental in controlling plant insect defense, wounding responses, development and secondary metabolite biosynthesis. TFs play a paramount role in regulating genes involved in likely all aspects of plant growth and development, including secondary metabolism. In the last decades, an increasing number of TFs and the underlying mechanisms of secondary metabolism have been elucidated. Although the basic JA signaling pathway that releases many TFs is much clearer, the binding sites of TFs and other details need to be further clarified. Meanwhile, JA-responsive transcriptional regulations have been identified mostly in model plants, but the cases in medicinal plants remain elusive. JAs have an evolutionarily conserved role in reprogramming plant secondary metabolism. An important aspect herein is the concerted transcriptional activation of the genes encoding the enzymes that catalyze the secondary metabolic reactions. Often, JAs simultaneously induce all known biosynthetic genes from a particular pathway, as illustrated by the vinblastine, nicotine, artemisinin, taxol and ginsenoside pathways in C. roseus, N. tabacum, A. annua, Taxus sp. and P. ginseng, respectively. Increasing secondary metabolites by engineering TFs is a promising method in near future.
References
Verpoorte R, van der Heijden R, Memelink J et al (2000) Engineering the plant cell factory for secondary metabolite production. Trans Res 9:323–343
Roepke J, Salim V, Wu M et al (2010) Vinca drug components accumulate exclusively in leaf exudate of madagascar periwinkle. Proc Natl Acad Sci USA 107:15287–15292
Singh YN (1986) Traditional medicine in Fiji: some herbal folk cures used by Fiji Indians. J Ethnopharmacol 15:57–88
Ma D, Pu G, Lei C et al (2009) Isolation and characterization of AaWRKY1, an Artemisia annua transcription factor that regulates the amorpha-4, 11-diene synthase gene, a key gene of artemisinin biosynthesis. Plant Cell Phys 50:2146–2161
Wilson SA, Roberts SC (2014) Metabolic engineering approaches for production of biochemicals in food and medicinal plants. Curr Opi Biotech 26:174–182
Nag SA, Qin JJ, Wang W et al (2012) Ginsenosides as anticancer agents: In vitro and in vivo activities, structure activity relationships, and molecular mechanisms of action. Front in Pharmacol 3:1–18
Yang CQ, Fang X, Wu XM et al (2012) Transcriptional regulation of plant secondary metabolism. J Integr Plant Biol 54:703–712
Vom Endt D, Kijne JW, Memelink J et al (2002) Transcription factors controlling plant secondary metabolism: what regulates the regulators? Phytochem 61:107–114
Memelink J (2009) Regulation of gene expression by jasmonate hormones. Phytochem 70:1560–1570
Pauwels L, Inzé D, Goossens A et al (2009) Jasmonate-inducible gene: What does it mean? Trend Plant Sci 14:87–91
Zhao J, Davis LC, Verpoorte R et al (2005) Elicitor signal transduction leading to production of plant secondary metabolites. Biotec Adv 23:283–333
Browse J (2009) Jasmonate passes muster: a receptor and targets for the defense hormone. Annu Rev Plant Biol 60:183–205
Schaller F (2001) Enzymes of the biosynthesis of octadecanoid-derived signalling molecules. J Exp Bot 52:11–23
Pauwels L, Goossens A (2011) The JAZ proteins: A crucial interface in the jasmonate signaling cascade. Plant Cell 23:3089–3100
Chung HS, Howe GA (2009) A critical role for the TIFY motif in repression of jasmonate signaling by a stabilized splice variant of the jasmonate zim-domain protein JAZ10 in Arabidopsis. Plant Cell 21:131–145
Chini A, Fonseca S, Fernandez G et al (2007) The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448:666–671
Dombrecht B, Xue GP, Sprague SJ et al (2007) MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 19:2225–2245
Kazan K, Manners JM (2013) MYC2: the master in action. Mol Plant 6:686–703
Zhang H, Hedhili S, Montiel G et al (2011) The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J 67:61–71
Shoji T, Hashimoto T (2011) Tobacco MYC2 regulates jasmonate-inducible nicotine biosynthesis genes directly and by way of the NIC2-locus ERF genes. Plant Cell Physiol 52:1117–1130
Zhang HB, Bokowiec MT, Rushton PJ et al (2012) Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple jasmonate-inducible steps in nicotine biosynthesis. Mol Plant 5:73–84
Todd AT, Liu E, Polvi SL et al (2010) A functional genomics screen identifies diverse transcription factors that regulate alkaloid biosynthesis in Nicotiana benthamiana. Plant J 62:589–600
Qi T, Song S, Ren Q et al (2011) The jasmonate-zim-domain proteins interact with the WD-repeat/ bHLH/ MYB complexes to regulate jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 23:1795–1814
Petroni K, Tonelli C (2011) Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci 181:219–229
Chatel G, Montiel G, Pre M et al (2003) CrMYC1, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J Exp Bot 54:2587–2588
Menke FL, Champion A, Kijne JW et al (1999) A novel jasmonate-and elicitor-responsive element in the periwinkle secondary metabolite biosynthetic gene STR interacts with a jasmonate-and elicitor-inducible AP2-domain transcription factor, ORCA2. EMBO J 18:4455–4463
van der Fits L, Memelink J (2000) ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 289:295–297
Suttipanta N, Pattanaik S, Kulshrestha M et al (2011) The transcription factor CrWRKY1 positively regulates the terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol 157:2081–2093
Pauw B, Hilliou FA, Martin VS et al (2004) Zinc finger proteins act as transcriptional repressors of alkaloid biosynthesis genes in Catharanthus roseus. J Biol Chem 279:52940–52948
Yu ZX, Li JX, Yang CQ et al (2012) The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua. Mol Plant 5:353–365
Romero MR, Serrano MA, Vallejo M et al (2006) Antiviral effect of artemisinin from Artemisia annua against a model member of the Flaviviridae family, the bovine viral diarrhoea virus (BVDV). Planta Med 72:1169–1174
Efferth T (2006) Molecular pharmacology and pharmacogenomics of artemisinin and its derivatives in cancer cells. Curr Drug Targ 7:407–421
Cáceres A, Girón LM, Martínez AM et al (1987) Diuretic activity of plants used for the treatment of urinary ailments in guatemala. J Ethnopharmacol 19:233–245
Binorkar SV, Jani DK (2012) Traditional medicinal usage of tobacco-a review. Spa DD Peer Rev J Complement Med Drug Discov 2:127–134
Gális I, Šimek P, Narisawa T et al (2006) A novel R2R3-MYB transcription factor NtMYBJS1 is a methyl jasmonate-dependent regulator of phenylpropanoid-conjugate biosynthesis in tobacco. Plant J 46:573–592
Shoji T, Kajikawa M, Hashimoto T et al (2010) Clustered transcription factor genes regulate nicotine biosynthesis in tobacco. Plant Cell 22:3390–3409
Onkokesung N, Gaquerel E, Kotkar H et al (2012) MYB8 controls inducible phenolamide levels by activating three novel hydroxycinnamoyl-coenzyme a: polyamine transferases in Nicotiana attenuata. Plant Physiol 158:389–407
Li ST, Zhang P, Zhang M et al (2012) Transcriptional profile of Taxus chinensis cells in response to methyl jasmonate. BMC Genomics 13:295–306
Lenka SK, Nims NE, Vongpaseuth K et al (2015) Jasmonate-responsive expression of paclitaxel biosynthesis genes in Taxus cuspidata cultured cells is negatively regulated by the bHLH transcription factors TcJAMYC1, TcJAMYC2 and TcJAMYC4. Front Plant Sci. doi:10.3389/fpls.2015.00115
Sun C, Li Y, Wu Q et al (2010) De novo sequencing and analysis of the American Ginseng root transcriptome using a GS FLX Titanium platform to discover putative genes involved in ginsenoside biosynthesis. BMC Genomics 11:262–274
Sun Y, Niu Y, Xu J et al (2013) Discovery of WRKY transcription factors through transcriptome analysis and characterization of a novel methyl jasmonate-inducible PqWRKY1 gene from Panax quinquefolius. Plant Cell Tiss Org Cult 114:269–277
Burlat V, Oudin A, Courtois M et al (2004) Co-expression of three mep pathway genes and geraniol 10-hydroxylase in internal phloem parenchyma of Catharanthus roseus implicates multicellular translocation of intermediates during the biosynthesis of monoterpene indole alkaloids and isoprenoid-derived primary metabolites. Plant J 38:131–141
Koul M, Lakra NS, Chandra R et al (2013) Catharanthus roseus and prospects of its endophytes: A new avenue for production of bioactive metabolites. Int J Pharm Sci Res 4:2705–2716
Rai V, Tandon PK, Khatoon S (2014) Effect of chromium on antioxidant potential of Catharanthus roseus varieties and production of their anticancer alkaloids: vincristine and vinblastine. Bio Med Res Int. doi:10.1155/2014/934182
Goldhaber-Pasillas GD, Mustafa NR, Verpoorte R et al (2014) Jasmonic acid effect on the fatty acid and terpenoid indole alkaloid accumulation in cell suspension cultures of Catharanthus roseus. Molecules 19:10242–10260
Almagro L, Gutierrez J, Pedreño MA et al (2014) Synergistic and additive influence of cyclodextrins and methyl jasmonate on the expression of the terpenoid indole alkaloid pathway genes and metabolites in Catharanthus roseus cell cultures. Plant Cell Tissue Organ Cult 119:543–551
Zhou ML, Zhu XM, Shao JR et al (2010) Transcriptional response of the catharanthine biosynthesis pathway to methyl jasmonate/nitric oxide elicitation in Catharanthus roseus hairy root culture. Appl Microbiol Biotechnol 88:737–750
Chen Q, Chen Z, Lu L et al (2013) Interaction between abscisic acid and nitric oxide in PB90 induced catharanthine biosynthesis of Catharanthus roseus cell suspension cultures. Biotechnol Prog 29:994–1001
Ganteta P, Memelink J (2002) Transcription factors: tools to engineer the production of pharmacologically active plant metabolites. Trend Pharmacol Sci 12:563–569
Almagro L, Pérez FF, Pedreño MA (2015) Indole alkaloids from Catharanthus roseus: bioproduction and their effect on human health. Molecules 20:2973–3000
Memelink J, Gantet P (2007) Transcription factors involved in terpenoid indole alkaloid biosynthesis in Catharanthus roseus. Phytochem Rev 6:353–362
Tang KX, Liu DH, Wang YL et al (2011) Overexpression of transcriptional factor ORCA3 increases the accumulation of catharanthine and vindoline in Catharanthus roseus hairy roots. Russ J Plant Physiol 58:415–422
Liu DH, Ren WW, Cui LJ et al (2011) Enhanced accumulation of catharanthine and vindoline in Catharanthus roseus hairy roots by overexpression of transcriptional factor ORCA2. Afr J Biotech 10:3260–3268
Li CY, Leopold AL, Sander GW et al (2013) The ORCA2 transcription factor plays a key role in regulation of the terpenoid indole alkaloid pathway. BMC Plant Biol 13:155–172
Van Der Fits L, Memelink J (2001) The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via interaction with a jasmonate-responsive promoter element. Plant J 25:43–53
Peebles CAM, Hughes EH, Shanks JV et al (2009) Transcriptional response of the terpenoid indole alkaloid pathway to the over expression of ORCA3 along with jasmonic acid elicitation of Catharanthus roseu hairy roots over time. Metab Eng 11:76–86
Pan Q, Wang Q, Yuan F et al (2012) Overexpression of ORCA3 and G10H in Catharanthus roseus plants regulated alkaloid biosynthesis and metabolism revealed by NMR-metabolomics. PLoS One 7:e43038
Wang CT, Liu H, Gao XS et al (2010) Overexpression of G10H and ORCA3 in the hairy roots of Catharanthus roseus improves catharanthine production. Plant Cell Rep 29:887–894
Schluttenhofer C, Pattanaik S, Patra B et al (2014) Analyses of Catharanthus roseus and Arabidopsis thaliana WRKY transcription factors reveal involvement in jasmonate signaling. BMC Genomics 15:502–522
Saiman MZ, Mustafa RN, Pomahacova B et al (2014) Analysis of metabolites in the terpenoid pathway of Catharanthus roseus cell suspensions. Plant Cell Tissue Organ Cult 117:225–239
Patra B, Schluttenhofer C, Wu Y et al (2013) Transcriptional regulation of secondary metabolite biosynthesis in plants. Biochim Biophys Acta 1829:1236–1247
Wang X, Bennetzen JL (2015) Current status and prospects for the study of Nicotiana genomics, genetics, and nicotine biosynthesis genes. Mol Genet Genomics 290:1–11
Sierro N, Battey JN, Ouadi S et al (2014) The tobacco genome sequence and its comparison with those of tomato and potato. Nat Commun 5:1–9
Dewey RE, Xie J (2013) Molecular genetics of alkaloid biosynthesis in Nicotiana tabacum. Phytochemistry 94:10–27
Kato K, Shoji T, Hashimoto T (2014) Tobacco nicotine uptake permease regulates the expression of a key transcription factor gene in the nicotine biosynthesis pathway. Plant Physiol 166:2195–2204
De Boer K, Tilleman S, Pauwels L et al (2011) Apetala2/ethylene response factor and basic helix-loop-helix tobacco transcription factors cooperatively mediate jasmonate-elicited nicotine biosynthesis. Plant J 66:1053–1065
Shoji T, Hashimoto T (2014) Stress-induced expression of NICOTINE2-locus genes and their homologs encoding ethylene response factor transcription factors in tobacco. Phytochemistry. doi:10.1016/j.phytochem.2014.05.017
Ariey F, Witkowski B, Amaratunga C et al (2014) A molecular marker of artemisinin-resistant Plasmodium falciparum malaria. Nature 505:50–55
Utzinger J, Xiao SH, Tanner M et al (2007) Artemisinins for schistosomiasis and beyond. Curr Opin Investig Drugs 8:105–116
Ji Y, Jingwei X, Yalin S et al (2014) Cloning and characterization of AabHLH1, a bHLH transcription factor that positively regulates artemisinin biosynthesis in Artemisia annua. Plant Cell Physiol 55:1592–1604
Caretto S, Quarta A, Durante M et al (2011) Methyl jasmonate and miconazole differently affect arteminisin production and gene expression in Artemisia annua suspension cultures. Plant Biol 13:51–58
Abdin MZ, Alam P (2015) Genetic engineering of artemisinin biosynthesis: prospects to improve its production. Acta Physiol Plant 37:1–12
Paddon CJ, Westfall PJ, Pitera DJ et al (2013) High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 496:528–532
Jiang W, Lu X, Qiu B et al (2014) Molecular cloning and characterization of a trichome-specific promoter of artemisinic aldehyde Δ11 (13) reductase (DBR2) in Artemisia annua. Plant Mol Biol Rep 32:82–91
Wang H, Han J, Kanagarajan S et al (2013) Trichome-specific expression of the amorpha-4,11-diene 12-hydroxylase (cyp71av1) gene, encoding a key enzyme of artemisinin biosynthesis in Artemisia annua, as reported by a promoter-GUS fusion. Plant Mol Biol 81:119–138
Han J, Wang H, Lundgren A et al (2014) Effects of over expression of AaWRKY1 on artemisinin biosynthesis in transgenic Artemisia annua plants. Phytochemistry 102:89–96
Lu X, Zhang L, Zhang F et al (2013) AaORA, a trichome-specific AP2/ERF transcription factor of Artemisia annua, is a positive regulator in the artemisinin biosynthetic pathway and in disease resistance to botrytis cinerea. New Phytol 198:1191–1202
Howat S, Park B, Oh IS et al (2014) Paclitaxel: biosynthesis, production and future prospects. New Biotech 31:242–245
Cusido RM, Onrubia M, Sabater AB et al (2014) A rational approach to improving the biotechnological production of taxanes in plant cell cultures of Taxus spp. Biotech Adv 32:1157–1167
Walker K, Croteau R (2000) Molecular cloning of a 10-deacetylbaccatin iii-10-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli. Proc Natl Acad Sci USA 97:583–587
Li S, Zhang P, Zhang M et al (2013) Functional analysis of a WRKY transcription factor involved in transcriptional activation of the DBAT gene in Taxus chinensis. Plant Biol 15:19–26
Subramaniyam S, Mathiyalagan R, Natarajan S et al (2014) Transcript expression profiling for adventitious roots of Panax ginseng Meyer. Gene 546:89–96
Yang JW, Kim SS (2015) Ginsenoside Rc Promotes Anti-Adipogenic Activity on 3T3-L1 Adipocytes by Down-Regulating C/EBPα and PPARγ. Molecules 20:1293–1303
Jayakodi M, Lee SC, Park HS (2014) Transcriptome profiling and comparative analysis of Panax ginseng adventitious roots. J Ginseng Res 4:278–288
Zhao S, Wang L, Liu L et al (2014) Both the mevalonate and the non-mevalonate pathways are involved in ginsenoside biosynthesis. Plant Cell Rep 33:393–400
Khorolragchaa A, Kim YJ, Rahimi S et al (2014) Grouping and characterization of putative glycosyltransferase genes from Panax ginseng Meyer. Gene 536:186–192
Kim YS, Yeung EC, Hahn EJ et al (2007) Combined effects of phytohormone, indole-3-butyric acid, and methyl jasmonate on root growth and ginsenoside production in adventitious root cultures of Panax ginseng C.A Meyer. Biotechnol Lett 29:1789–1792
Afrin S, Zhu J, Cao H et al (2015) Molecular cloning and expression profile of an abiotic stress and hormone responsive MYB transcription factor gene from Panax ginseng. Acta Biochim Biophys Sin 47:267–277
Zhu F, Xi DH, Yuan S et al (2014) Salicylic Acid and Jasmonic Acid Are Essential for Systemic Resistance Against Tobacco mosaic virus in Nicotiana benthamiana. Am Phyto Soc 27:567–577
Groen SC, Whiteman NK (2014) The evolution of ethylene signaling in plant chemical ecology. J Chem Ecol 40:700–716
Miransari M, Smith DL (2014) Plant hormones and seed germination. Environ Exp Bot 99:110–121
Okada K, Abe H, Arimura GI (2015) Jasmonates induce both defense responses and communication in monocotyledonous and dicotyledonous plants. Plant Cell Physiol 56:16–27
Yang Z, Patra B, Li R et al (2013) Promoter analysis reveals cis-regulatory motifs associated with the expression of the WRKY transcription factor CrWRKY1 in Catharanthus roseus. Planta 238:1039–1049
Afrin S, Nuruzzaman M, Zhu J et al (2014) Combinatorial interactions of MYB and bHLH in flavonoid biosynthesis and their function in plants. J Plant Bio Res 3:65–77
Kim J, Chang C, Tucker ML (2015) To grow old: regulatory role of ethylene and jasmonic acid in senescence. Front Plant Sci 6:1–7
Wasternack C (2007) Jasmonates: an update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann Bot 100:681–697
Nagpal P, Ellis CM, Weber H et al (2005) Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 132:4107–4118
Acknowledgements
This work was supported by the National Natural Science Foundation of China (30470189, 81071821, 81250110086 and 81250110551), the Fundamental Research for the Central University of China (721500011), the Key Hunan Provincial Natural Science Foundation of China (13JJ2016 and 07JJ5096), the Science and Technology Program of Hunan Province of China (2014SK4050) and Changsha National High Tech Zone Innovation Leading Talent Program (2014-37).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Afrin, S., Huang, JJ. & Luo, ZY. JA-mediated transcriptional regulation of secondary metabolism in medicinal plants. Sci. Bull. 60, 1062–1072 (2015). https://doi.org/10.1007/s11434-015-0813-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11434-015-0813-0