
Abstract
Infectious diseases impose serious public health burdens and often have devastating consequences. The cyclic lipopeptides elaborated by bacteria Bacillus, Paenibacillus, Pseudomonas, Streptomyces, Serratia, Propionibacterium and fungus Fusarium are very crucial in restraining the pathogens. Composed of a peptide and a fatty acyl moiety these amphiphilic metabolites exhibit broad spectrum antimicrobial effects. Among the plethora of cyclic lipopeptides, only selective few have emerged as robust antibiotics. For their functional vigor, polymyxin, daptomycin, surfactin, iturin, fengysin, paenibacterin and pseudofactin have been integrated in mainstream healthcare. Daptomycin has been a significant part of antimicrobial arsenal since the past decade. As the magnitude of drug resistance rises in unprecedented manner, the urgency of prospecting novel cyclic lipopeptides is being perceived. Intense research has revealed the implication of these bioactive compounds stretching beyond antibacterial and antifungal. Anticancer, immunomodulatory, prosthetic parts disinfection and vaccine adjuvancy are some of the validated prospects. This review discusses the emerging applications, mechanisms governing the biological actions, role of genomics in refining structure and function, semi-synthetic analog discovery, novel strain isolation, setbacks etc. Though its beyond the scope of the current topic, for holistic purpose, the role of lipopeptides in bioremediation and crop biotechnology has been briefly outlined. This updated critique is expected to galvanize innovations and diversify therapeutic recruitment of microbial lipopeptides.
Similar content being viewed by others


Introduction
Lipopeptides constitute a distinguished class of microbial secondary metabolites. For their multifaceted biological roles, they have garnered immense interest since decades (Cochrane and Vederas 2014). Some lipopeptides have consolidated their position as potent therapeutic compounds. Antibacterial, antiviral, antifungal, antitumor and immunomodulator are some of their proven and emerging properties (Meena and Kanwar 2015). Each lipopeptide is made of a hydrophilic peptide and a hydrophobic fatty acyl chain (Reder-Christ et al. 2012). The number of amino acids generally ranges from 7 to 25 while the length of fatty acid ranges from 13 to 17 carbons. So far, Bacillus and Paenibacillus spp. are the predominant lipopeptide producers (Cochrane and Vederas 2014). However, increasing number of lipopeptides are being isolated from Pseudomonas spp. too (Raaijmakers et al. 2010; Janek et al. 2012; Cochrane and Vederas 2014). Actinomycetes is a class of bacteria to have provided a wealth of functional lipopeptides (Singh et al. 2014b). Serratia is another pharmaceutically pertinent lipopeptide producing genus (Thies et al. 2014). Some strains of Propionibacterium have been discovered to elaborate bioactive lipopeptides (Hajfarajollah et al. 2014). Among fungi, Fusarium is the only well-characterized lipopeptide producer (Shiono et al. 2007). The lipopeptides are highly variable and their structural analogues result due to frequent amino acid substitutions (Baumgart et al. 1991).
Resistance of pathogens to conventional antibiotics has serious public health consequences. In this context, the lipopeptides have proved to be potent in inhibiting even the resilient pathogenic strains (Qian et al. 2012b). Also, they possess the ability to neutralize lipopolysaccharide (LPS) toxicity (Huang and Yousef 2014b). Several sagacious reviews have elucidated the therapeutic relevance of lipopeptides, with focus on their pharmacological spectrum and structure–function aspects (Hashizume and Nishimura 2008; Tang et al. 2008; Mandal et al. 2013a; Schneider et al. 2014). Some lipopeptides have been approved by both Food and Drug Administration (FDA) and European Medicines Agency (EMEA). They include daptomycin and echinocandins (caspofungin, micafungin, and anidulafungin) (Pappas et al. 2009; Kofla and Ruhnke 2011). The product homologues and specific applications of top lipopeptide-elaborating bacteria Bacillus and Paenibacillus spp. been reviewed in many insightful reviews (Cochrane and Vederas 2014; Aleti et al. 2015). The synergistic effect of lipopeptides with colistin (polymyxin E) in inhibition of multidrug-resistant (MDR) Acinetobacter baumannii has been reviewed (Claeys et al. 2014). There are many other seminal literature reviews encompassing specific areas of lipopeptide research.
Major cyclic lipopeptides
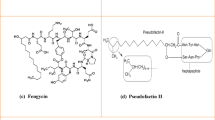
Lipopeptides can be cyclic or linear, based on the topology of the peptide chain (Ortíz-López et al. 2015). Polymyxins, daptomycin, surfactin, iturin, fengycin, paenibacterin, pseudofactin are the most prominent cyclic lipopeptides (Meena and Kanwar 2015; Laverty et al. 2011). TAN 1511, tridecaptin A1, tridecaptin B1, SRCAM and gageostatins A–C are linear lipopeptides (Lohans et al. 2014; Tareq et al. 2014; Cochrane et al. 2014). Here, the cyclic lipopeptides have been discussed as they encompass most biologically relevant lipopeptides. The list of current and emerging therapeutic cyclic lipopeptides has been presented in Table 1. It is evident from the table that most of cyclic lipopeptides have been discovered in last 5 years following the commercialization of daptomycin. The structures of the natural lipopeptides have been illustrated in Fig. 1. The structure of pseudofactin has been illustrarted in Fig. 2. The key charactesristics of the predominant cyclic lipopeptides have been summarized below.

Pharmaceutically and economically important natural cyclic lipopeptides

Structure of pseudofactin
Polymyxins
Polymyxin is a class of cationic cyclic lipopeptides discovered in the 1940’s (Velkov et al. 2013). The variants of polymyxins are elaborated by different species of Paenibacillus genus such as P. amylolyticus, P. polymyxa etc. (DeCrescenzo Henriksen et al. 2007; Yoshino et al. 2013). Polymyxin E or colistin has got wide commercial application (Lim et al. 2010; Dijkmans et al. 2014). Also, polymyxin B is another commercially licensed lipopeptide from this group (Kwa et al. 2007). The basic structure of this lipopeptide is a heptapeptide loop with a tripeptide side chain acylated by a fatty acid at amino terminal (Yu et al. 2015; Velkov et al. 2013; Fig. 1). This class of lipopeptides had been snubbed for their nephrotoxicity and neurotoxicity (Falagas and Kasiakou 2006), yet the dramatic rise of multi-drug resistance has revived interest in them. Their efficacy against resistant Pseudomonas aeruginosa, Acinetobacter baumannii and Klebsiella pneumonia Carbapenemase-producing strains has intensified studies on them (Arnold et al. 2011; Kanj and Kanafani 2011). Some insightful reviews have assessed the scopes and pitfalls in their broader usage (Landman et al. 2008). The chemistry, pharmacokinetics and pharmacodynamics of colistin has been revisited from the perspective of mitigating drug resistance (Gupta et al. 2009).
Daptomycin
Daptomycin is an acidic cyclic depsipeptide produced by Streptomyces roseosporus (Steenbergen et al. 2005; Baltz et al. 2006). It is made of 13 amino acids, a 10-member (decanoyl) macrolactone ring and three exocyclic residues attached to a C10-C13 fatty acyl chain (Shoemaker et al. 2006; Robbel and Marahiel 2010). It is well-established as an antibiotic since 2003, marketed as Cubicin® (Chan Tompkins and Harnicar 2008). Daptomycin has been successfully administered for skin infections, endocarditis, osteomyelitis, soft-tissue infections, even anthrax (Kaya et al. 2013; Gould et al. 2013; Xing et al. 2014).
Surfactin
Surfactin is a major class of antibiotic lipopeptides elaborated by Bacillus sp. The name justifies their outstanding surface tension-lowering ability (Angelini et al. 2009). This group of lipopeptides consists of a heptapeptide and C13–C15 fatty acyl chain (Shao et al. 2015). Though it is effective against both Gram positive and Gram negative bacteria, the non-specificity of cell lysis puts mammalian cells at risk. Further most surfactins are hemolytic, hence, they have limited medical usage (Duarte et al. 2014). Surfactin with a hydrophobic pentadecanoic fatty acyl chain was characterized to possess the hemolytic activity (Deleu et al. 2003). WH1fungin is a type of surfactin elaborated by Bacillus amyloliquefaciens WH1 (Qi et al. 2010). This lipopeptide demonstrated amelioration of type 1 diabetes mellitus in mouse model by immunomodulation (Gao et al. 2014).
Paenibacterin
Paenibacterin produced by Paenibacillus thiaminolyticus OSY-SE consists of 13 amino acids and a C15 fatty acyl chain (Huang et al. 2014a, b). It affects viability of both Gram negative and Gram positive human pathogens (Huang et al. 2012). Paenibacterin, owing to its positive charge binds to the negatively charged Gram negative endotoxins in vitro and, inhibits drug-resistant P. aeruginosa in vivo (Huang and Yousef 2014a).
Iturin
Iturin is an antifungal cyclic lipopeptide produced by Bacillus spp. It has a heptapeptide moiety and a β-hydroxy fatty acid chain of length C13–C17. Amino acid variations have led to high polymorphism in iturins. The characterized variants are iturin A, iturin C, iturin D, iturin E, bacillomycin D, bacillomycin F, bacillomycin L, bacillomycin Lc and mycosubtilin (Roongsawang et al. 2010; Pathak and Keharia 2013; Ali et al. 2014). Iturins are well-known biocontrol agents towards plant pathogens such as Xanthomonas campestris Cucurbitae, Pectobacterium carotovorum subsp. Carotovorum, Rhizoctonia solani, Fusarium graminearum etc. (Zeriouh et al. 2011; Gong et al. 2015). There is no information if iturins are medically-relevant.
Fengycin
Fengycin is an antifungal lipopeptide complex produced by some Bacillus and Paenibacillus strains (Cochrane and Vederas 2014). It consists of a decapeptide and C14-C18 fatty acyl chain (Nasir et al. 2013). It acts on plasma membrane of cells by inducing lipid bilayer perturbations and local electrostatic-driven remodeling (Horn et al. 2013). It acts widely against filamentous fungi. It has been observed to inhibit Rhizoctonia solani in the cotton rhizosphere (Guo et al. 2014). Though currently, it has limited implication in pharmaceutical field, its milder hemolytic activity compared to other lipopeptides suggests its possible medical applications. Its advocated to be a potent candidate to treat dermatomycoses (candidiasis and ringworm) (Eeman et al. 2014).
Pseudofactin
Pseudofactin II is a cyclic lipopeptide secreted by Pseudomonas fluorescens BD5. This biosurfactant consists of an octapeptide linked to palmitic acid. It exerts outstanding anti-adhesive and anti-biofilm properties, that might be exploited for therapeutic purposes (Janek et al. 2012). Further, it exerted cytotoxicity towards human melanoma A375 cells mediated by plasma membrane permeabilization (Janek et al. 2013).
Apart from the above mentioned cyclic lipopeptides, there are many other cyclic lipopeptides with potential roles in medical fields which include Bacillus licheniformis-elaborated lichenysin (Nerurkar 2010), Bacillus subtilis SSE4-elaborated subtulene A (Thasana et al. 2010), Paenibacillus kobensis M-elaborated mattacin (Martin et al. 2003), Paenibacillus tianmuensis-elaborated battacin (Qian et al. 2012b), Paenibacillus elgii-elaborated pelgipeptin (Qian et al. 2012a), Micromonospora-elaborated rakicidin (Takeuchi et al. 2011), Actinoplanes friuliensis HAG 010964-elaborated friulimicin (Schneider et al. 2009; Rückert et al. 2014), Serratia marcescens-elaborated serrawettin W1 (Thies et al. 2014), Pseudomonas corrugata CFBP 5454-elaborated corpeptins (Strano et al. 2015), Pseudomonas fluorescens SS101-elaborated massetolide (Song et al. 2014) etc. In fact, the list is exhaustive and it reflects the rationale of searching for novel lipopeptides.
Biosynthetic mechanisms
Identification of genes involved in lipopeptides synthesis and understanding their genomic organization holds immense scope for functional manipulation (Walsh 2002). Transposon mutagenesis experiments have revealed many genes governing lipopeptide generation (Martin et al. 2003). The key machinery for the synthesis is multi-modular and consists of non-ribosomal peptide synthetases (NRPSs) (Roongsawang et al. 2010). The integrated system introduces significant heterogeneity among the lipopeptides with respect to the arrangement of amino acids, cyclization of peptide and length of fatty acyl chain (Ongena and Jacques 2008). An in-depth review depicting the regulatory mechanisms of lipopeptide biosynthesis has been published (Raaijmakers et al. 2010). Another insightful review discusses the region- and stereo-specific assembly of the peptides. (Strieker et al. 2010). Catalytic role of NRPS components in selection of the amino acids, subsequent condensations,, termination and cyclization of the peptide chain has been well-documented (Roongsawang et al. 2010). The stereo-controlled pathways incorporate nonproteinogenic amino acids into the peptide moiety e.g. l-kynurenine in daptomycin (Robbel and Marahiel 2010), L-2,4-diaminobutyrate in pelgipeptin (Qian et al. 2012a), pipecolinic acid, methylaspartic acid, and 2,3-diaminobutyric acid in friulimicin etc.
The peptide moiety is inactive until its coupled with a fatty acyl chain. As the aliphatic chain of variable length fuses with the N-terminal residue of the peptide chain, the bioactive lipopeptide is generated (Malina and Shai 2005). The conjugates with a macrocylic ring structure, an exocyclic tail and fatty acyl chain serve as an amphiphilic molecules (Schneider et al. 2014).
To understand mechanistic variation, lipopeptide synthesis in different strains have been studied. Here, some important findings have been summarized. The investigation of P. thiaminolyticus OSY-SE, revealed the location of paenibacterin-encoding gene cluster within a 52-kb DNA region. These genes encoded NRPS members PbtA, PbtB and PbtC, and two ABC-transporters, PbtD and PbtE (Qian et al. 2012a, b). Role of these peptide synthetases in amino acids assembly was recognized (Huang and Yousef 2014b). Daptomycin production by S. roseosporus SW0702 at transcriptional level was investigated. A gene dptR2, encoding a DeoR-type regulator located close to the daptomycin biosynthesis gene cluster was found essential for daptomycin production (Wang et al. 2014). Also, in Pseudomonas corrugata, the genes encoding NRPS and an ABC-type transport system for lipopeptide corpeptin, belonged to the same transcriptional unit (Strano et al. 2015). The genome of B. subtilis 916 has four NRPS gene clusters such as srf, bmy, fen, and loc, for biosynthesis of surfactins, bacillomycin L, fengycins, and locillomycins, respectively (Luo et al. 2015).
Biomedical roles
The cyclic lipopeptides have been validated to exert a diverse array of biological effects such as antibacterial, antifungal, immunomodulation, antitumor and prosthetic disinfection. Further, adequate understanding of the action of the lipopeptides is paramount for their therapeutic optimization and valorization. So, the therapeutic roles and the underlying mechanisms have been discussed below.
Antibacterial
Among antibacterial cyclic lipopeptides, daptomycin is most prominent. It upsets the membrane potential of Gram-positive bacteria (Kanafani and Corey 2007). When administered to left heart endocarditis patients, it inhibited methicillin-resistant Staphylococcus aureus (MRSA) and Streptococcus mutans without any side effects (Kaya et al. 2013). At an MIC value of 0.78 μg/ml, it eradicated the anthrax pathogen Bacillus anthracis AP422, by perturbing the latter’s membrane potential. The efficacy was comparable to ciprofloxacin and penicillin G, though the mechanism of inhibition varied (Xing et al. 2014). The calcium dependent attachment as well as insertion of the lipophilic part to pathogen’s cytoplasm, followed by rapid potassium efflux leading to depolarization of the cell membrane has been understood (Silverman et al. 2003; Beiras-Fernandez et al. 2010). Daptomycin-caused pores were selective for cations, with permeability being highest for sodium, potassium and other alkali metal ions (Zhang et al. 2014b). Streptomyces amritsarensis sp. nov. produced a lipopeptide which abolished Bacillus subtilis (MTCC 619), Staphylococcus epidermidis (MTCC 435), Mycobacterium smegmatis (MTCC 6) and MRSA at MIC of 10, 15, 25 and 45 μg/ml, respectively. The lipopeptide was pH and protease stable. The antimicrobial action could be attributed to the surfactant nature of the lipopeptide (Sharma et al. 2014). Paenibacterin acted on both Gram negative and Gram positive bacteria. It disrupted membrane structure and caused efflux of potassium ions from cells of E. coli and S. aureus. The positive charge on the antibiotic neutralized the negatively charged-LPS on the outer membrane of E. coli (Huang and Yousef 2014a). Propionibacterium freudenreichii subsp. freudenreichii elaborated a biosurfactant that inhibited Rhodococcus erythropolis and exhibited anti-adhesion towards Pseudomonas aeruginosa (Hajfarajollah et al. 2014). The action of daptomycin against Gram positive pathogens and paenibacterin against Gram negative pathogens has been presented in Fig. 3.

Action of daptomycin against Gram positive pathogens and paenibacterin against Gram negative pathogens
Antifungal
Bacillus amyloliquefaciens WH1-elaborated WH1fungin inhibited the growth of Rhizoctonia solani, Helminthosprium maydis, Fusarium oxysporium, Botrytis cinereapers, Gibberella saubinetii, Colletotrichum gossypii, C. capsici, Physalospora piricola, Sclerotinia sclerotiorum and Candida albicans (Qi et al. 2010; De Brucker et al. 2011). At low dose, WH1 fungin impeded glucan synthesis and ATPase activity. Eventually apoptosis of fungal cells occurred (De Brucker et al. 2011). However, at higher concentration, it formed pores in the fungal pathogens (Qi et al. 2010). Candida albicans is a common human pathogen and its resilience is attributed to its variable morphology and biofilm synthesis. Bacillus amyloliquefaciens AR2 lipopeptide hindered the biofilm formation and demonstrated strong fungicidal effect (Rautela et al. 2014). Antifungal action of Bacillus amyloliquefaciens-derived bacillomycin L was unravelled to be mediated by the modulation of fungal membrane permeability as well as by interaction with other intracellular targets (Zhang et al. 2013a).
Immunomodulation
Lipopeptides have been recognized as immunomodulators that interact with pattern recognition receptors such as Toll-like receptors (TLRs) expressed on antigen presenting cells (macrophages and dendritic cells) (Kelesidis 2014). The immunomodulatory role of Bacillus-produced surfactin WH1fungin against Type 1 diabetes mellitus was assessed. When orally given to NOD mice at 5–25 mg/kg for 4 weeks, it tilted the immune response from Th1- to Th2-type. The observed changes in immune landscape were suppressed T cells proliferation, down-regulated activated CD8+T cells and effector molecules (tumor necrosis factor (TNF)-α and interferon (IFN)-γ), and increased regulator T cells (Tregs). These immune alteration contributed to lowered incidence of the autoimmunity-driven diabetes (Gao et al. 2014). Daptomycin possesses immunomodulatory properties, manifested in suppression of cytokine expression by host immunity on MRSA infection (Tirilomis 2014). It has been observed that in presence of divalent ions, daptomycin interacts with phospholipids of immune cell membranes and permeates through them (Kelesidis 2014). Exploitation of the lipopeptides for immunity correction has barely been explored and it holds immense prospect.
Antitumor
Rising instances of drug resistance and normal tissue toxicity has necessitated the prospecting of benign yet effective therapeutics (Sagar et al. 2006). In this context, the lipopeptides appear promising for their ability to induce apoptosis and prevent proliferation by manipulation of signaling pathways (Dey et al. 2015). Surfactin from Bacillus subtilis induce anti-proliferation on colon carcinoma LoVo cells by apoptosis induction, cell cycle arrest and survival signaling suppression (Kim et al. 2007). The effect of Bacillus natto T2-elaborated cyclic lipopeptide on human leukemic K562 cells was investigated. The lipopeptide induced a sustained increment in the concentration of intracellular Ca2+, which induced cell apoptosis and ERK phosphorylation. The kinase subsequently activated Bax, cytochrome c and caspase-3, leading to apoptosis (Wang et al. 2009). A marine Bacillus circulans DMS-2 elaborated surfactin and fengycin with pronounced antiproliferative activity against the human colon cancer HCT-15 and HT-29 cell lines (Sivapathasekaran et al. 2010). Rakicidin A produced by the actinomycete Micromonospora strain, induced hypoxia-selective cytotoxicity in solid tumors. It induced death of chronic myelogenous leukemia cells by caspase-dependent as well as independent pathways. The result predicted that rakicidin A can be used to target the drug-resistant dormant cancer cells in hypoxic milieu (Takeuchi et al. 2011). Bacillus amyloliquefaciens strain fiply 3A produced a bacillomycin D, which dose-dependently killed human cancer cell lines such as alveolar adenocarcinoma A549, renal carcinoma A498, and colon adenocarcinoma HCT-15. Apoptosis mediated cell death was confirmed through the observed cell shrinkage, nuclear condensation and fragmentation of nuclei (Hajare et al. 2013). A review discussing the anticancer efficacy of lipopeptides has been published. Their cancer inhibition property has been correlated to cell cycle arrest, intervention with crucial signaling pathways such as Akt, ERK/c-Jun N-terminal kinase (JNK), Janus kinase/signal transducer and activator of transcription (JAK/STAT), activation of natural killer T (NKT) cells, suppression of cell surface receptors (EGFR, VEGFR, PDGFR and IGFR), prevention of angiogenesis and induction of apoptosis (Dey et al. 2014). Though these amphiphiles have only recently been investigated to tackle cancer, their low toxicity, safety, efficacy against resistant pathways are emerging and they seem poised to contribute more to oncology in near future. Figure 4 illustrates the anticancer mechanism of lipopeptides.

The anticancer mechanism of lipopeptides
Disinfection of prosthetic implants
Prosthetic joint infection is a noxious issue following joint replacement (Peel et al. 2012). Biofilm formation in the arthroplasty milieu leads to high patient morbidity (McConoughey et al. 2014). Despite, technical innovations, the rising cases of MRSA is a serious concern (Bassetti et al. 2014). The efficacy and safety of high doses of daptomycin (10 mg/kg daily) along with rifampin for tackling the implant pathogens was investigated. Preliminary results showed moderate efficacy towards fluoroquinolone-resistant staphylococci (Lora-Tamayo et al. 2014). A patient with Enetercoccus faecalis prosthetic joint infection, when administered with daptomycin at the above dose for 12 weeks, significant biofilm inhibition was observed (Yuste et al. 2014).
New frontiers in lipopeptide research
Semisynthetic drug designing from natural lipopeptides
As the specter of microbial and cancer drug resitance escalates, novel pharamaceuticals are urgently needed. Despite a vast arsenal of lipopeptides and their diversified actions, the above challenges are far from contained. Modification of the lipopeptides for improved biological specificity might resolve the issues. The unprecedented devlopement in structural elucidation and knowledge on their functional mechanistic has enabled analog fabrication. Some key progress in this field have been mentoned below. Altering the peptide and acyl group led to changed geometry and lipophilicity, thus antifungal potency properties of the analog. Antifungal effect of fungal lipopeptide echinocandin was enhanced by enzymatic deacylation followed by chemical reacylation (Debono et al. 1995). A member of this class, mulundocandin was subjected to hemiaminal function modification and resultant improvement in solution stability and activity was observed (Lal et al. 2003). Also, modification of the N-linked acyl chain of echinocandins reduced the hemolytic effect of the analogs, vital for therapeutic applications (Eschenauer et al. 2007). Laspartomycin C, another cyclic lipopeptide when modified by deacylation, exerted higher efficacy against MRSA, vancomcin-resistant Staphylococcus and vancomycin-resistant Enterococcus (Curran et al. 2007). A54145, a cyclic lipopeptide with high similarity to daptomycin was modified to generate CB-182,462. The derivative was capable of forming oligomers like daptomycin (Alexander et al. 2011; Zhang et al. 2013b). The mentioned examples are only a few of the large array of semi-synthetic lipopeptide analogs.
The pace of superior lipopeptide generation has been punctuated. The integration of combinatorial biosynthesis (Baltz et al. 2006) and genetic engineering (Qian et al. 2012a, b) might accelerate it. The scope of these emerging techniques have been discussed later. Finally, the semisynthetic modification of the natural lipoprptides is intricate task and success is not gurantted. Yet, the works done have furnished information on the behavior of the constituents. The structure–activity relationship in ultrashort cationic lipopeptides was explored. Subjecting the side chain of the amino acids to constraint did not provide any favorable antimicrobial activity (Domalaon et al. 2014). These garnered insights could be applied in diversifying synthetic lipopeptide repertoire.
Vaccine formulation prospects
As the onslaught of pathogens mounts, the dire need for vaccines is being perceived. Lipopeptides are promising in this goal as they can serve as non-toxic, non-pyrogenic immunoadjuvants when combined with conventional antigens (Mittenbühler et al. 1997; BenMohamed et al. 2002; Moyle and Toth 2008; Toth et al. 2008). The recognition of lipopeptides by TLRs, result in induction of immune response (Steinhagen et al. 2011). A stable, human TLR2-specific lipopeptide as candidate vaccine was designed and efficacy proved in rabbit model (Salunke et al. 2013). Recently, the interaction of daptomycin with lipid bilayer was explored and its lipid extracting effect was described, which might help in designing vaccines with lipopeptides as adjuvant, since it affects the permeability of a membrane (Chen et al. 2014). Intranasal immunization with polymyxin B and colistin, along with ovalbumin amplified the humoral immune responses dose-dependently. The adjuvanticity was independent of pathogenic toxin and side effect-free (Yoshino et al. 2013). As an adjuvant, WH1 fungin protected the co-administered protein antigens from the hostile gut milieu, promoted their mucosal uptake and increased the expression of cell surface markers and cytokines in dendritic cells (Gao et al. 2014). Improved surveillance of these innate immune cells has direct consequence in pathogen elimination. This finding heralds the inclusion possibility of the surfactin in oral vaccine formulations WH1 fungin-laced hepatitis B surface antigen when administered to mice elicited strong immunity. It indicated the possibility of potent vaccine development using the surfactin (Pan et al. 2014). The development of lipopeptide vaccines, their TLR2 recognition and immune activation mechanism have been reviewed (Zaman and Toth 2013). Design of therapeutic human papilloma virus (HPV) vaccines using lipoimmunogens was met with considerable success (Shen et al. 2015). The next generation of vaccine development against a plethora of pathogens can certainly benefit from integration of these lipopeptides as adjuvants.
Genetic engineering
The remarkable advent in genomics has generated many opportunities for lipopeptide yield enhancement and tailored lipopeptide synthesis. Some of the revolutionary technologies have been discussed below. Genetic engineering is a robust tool to induce biologically important lipopeptide production from non-producing microbes. By introducing the expression plasmids carrying NRPS genes, peptide cores of daptomycin-like lipopeptides were synthesized. On subsequent coupling with desired acyl chains, an array of bioactive lipopeptides could be generated (Miao et al. 2006). The extraordinary structural and functional diversity of natural products has enabled their exploitation for assembly and modification. The usage of engineered enzymes and expression of biosynthetic pathways have broadened the scope of developing effective drug analogs (Sun et al. 2015). NRPS being a multi-enzyme family, is amenable for combinatorial biosynthesis. By selective domain exchange, clustering and pathway-level recombination, novel lipopeptides with efficacy superior than daptomycin were generated (Baltz 2014). The potential of ribosome engineering in activation of these cryptic pathways, leading to higher expression of secondary metabolites like lipopeptides has been reviewed (Ochi and Hosaka 2013). This novel approach based on awakening of silent genes might contribute to lipopeptide production. A library of mutants and plasmids was generated to facilitate combinatorial biosynthesis of broad spectrum lipopeptides (Alexander et al. 2010). Heterologous expression of the peptide synthetase gene cluster-carrying plasmids in mutants can be used to produce lipopeptide with desired peptide architecture. This diversity-based multiplex combinatorial biosynthesis holds immense relevance in target-specific lipopeptide drug production. Promoter replacement for enhanced production of lipopeptide was explored. Substitution of the native promoter of B. subtilis fmbR with the Pspac promoter, augmented surfactin production. The recombinant B. subtilis strain elaborated ten-fold more surfactin than the wild type when induced by isopropyl β-1-thiogalactopyranoside (IPTG) (Sun et al. 2009). Genome shuffling is increasingly being adopted for tailored metabolite production, improving substrate uptake and strain tolerance (Gong et al. 2009). Surfactin yield from a genome-shuffled recombinant strain of B. amyloliquefaciens was enhanced, verified from the higher expression of the sufactin synthetase gene srfA (Zhao et al. 2012). The emergent approaches of genetic engineering is expected to develop microbial phenotypes with high competence for pharmaceutically useful lipopeptides.
Role of proteomics and metabolomics
Comparative quantitative proteomics has furnished invaluable insights on the mechanism of lipopetide action and response of the pathogens to them. Pathway analysis at proteome-level unveils the putative modes of the lipopeptides. The modulation of bioenergetic pathways in the pathogens can be harnessed to refine the structure and function of the drugs. Comparative proteomics of different pathogenic strains, provides information on differential abundance of proteins, in response to the lipopeptides. This knowledge can be applied to discriminate between drug-sensitive and drug-resistant strains. Proteomics approach contributed to the daptomycin resistance mechanistic of Staphylococcus aureus (Fischer et al. 2011). Comparative proteomic analysis can furnish information on the functional improvement by metabolic engineering. To find the difference in protein profiles of the parental and genome-shuffled B. amyloliquefaciens strains, their proteomes were analyzed. Protein electrophoresis followed by mass spectrometry, implied higher expression of proteins associated with sufactin biosynthesis (Zhao et al. 2014).
Lipopeptide sequence verification is paramount for deciphering structure–function relationship (Yang et al. 2006). Metabolomics, the omic science of analyte detection has significantly facilitated the peptide analysis task (Zhang et al. 2012). The high-end tools mass spectrometer (MS) and nuclear magnetic resonance (NMR) have immensely contributed to the field. The fast atom bombardment tandem MS, time of flight (TOF) MS and electrospray ionization (ESI) MS has enabled sequencing without hydrolysis of the peptide. NMR (1D and 2D) has identified the substitutions in the lipopeptide variants. An antifungal cyclic lipopeptide kannurin from Bacillus cereus AK1 was elucidated by ESI–MS and Fourier transform infrared (FT-IR). The lipopeptide was made of amino acids Leu-Asp-Val-Leu-Leu-Leu-Leu (Ajesh et al. 2013). Thin layer chromatography (TLC) and FT-IR analysis characterized a surfactant lipopeptide elaborated by Fusarium sp. BS-8 isolated from an oil contaminated site (Qazi et al. 2014). Halobacteriaceae archaeon AS65 produced a strong biosurfactant when grown in minimal salt medium-amended with banana peel and monosodium glutamate. FT-IR, NMR, and MS analysis verified the compound to be a lipopeptide. It exhibited antimicrobial, oil emulsification and bioremediation ability (Chooklin et al. 2014). Streptomyces canus strain FIM0916 elaborated novel lipopeptides, amphomycin, aspartocin D and aspartocin E. Their compositions were elucidated by NMR analysis which revealed the latter two lipopeptides to be the variants of the former. These lipopeptides possessed antimicrobial capacity and they had exact same decapeptide varying only in acyl moiety (Yang et al. 2014a, b). MS, NMR and high resolution mass spectrometry (HR-MS/MS) was recruited to identify the lipopeptide polypeptin C from Paenibacillus ehimensis MA2012 (Naing et al. 2015). Metabolomic analyses revealed that Paenibacillus ehimensis IB-X-bcan elaborate multiple families of cyclic lipopeptides such as bacillomycin L-C15, fengycin/plipastatin A-C16 and their homologues (Aktuganov et al. 2014). Apart from expediting structural elucidation of lipopeptides, metabolomics has proved its role in selecting the novel lipopeptide-producing bacteria strains. Cystargamide was isolated from the fermentation broth of the actinomycete Kitasatospora cystarginea. LC–MS and NMR complemented each other in selecting the most potent strains among 12 isolates (Gill et al. 2014). The role of metabolomics in enriching the lipopeptide domain has just started to be appreciated. Only a negligible fraction of microbial secondary metabolomes have been analyzed to date (Yamanaka et al. 2014). So, many critical findings are expected in coming times.
Synergy with other drugs
Novel combinations of therapeutic agents are indispensable to overcome drug resistance. In this context, exploring the concerted effect of lipopeptides with other antibiotics, other lipopeptides and co-surfactants appear much promising. A study investigated the concomitant application of daptomycin with a β-lactam drug ceftaroline against MRSA-caused bacteremia. The combination substantially lowered the time of bacteremia clearance, by strong sensitization of host innate host defense against the pathogen (Sakoulas et al. 2014). The synergy of daptomycin and colistin was highly active against multidrug-resistant Acinetobacter baumannii in both in vitro assays and honeycomb moth larvae model of infection (Yang et al. 2014a, b). Synergistic effect of surfactin (magainin 2) and detergent octyl glucoside, along with detergent C12EO8 was observed. Also, coordinated effect of fengycin and detergent CHAPS, along with C12EO8 was witnessed. The improved membrane perturbation and peptide penetration was explained as the mechanism (Patel et al. 2014). A critical literature review emphasizes the need of innovative recruitment of the existing antibiotics, in the face of sluggish discovery of new candidates (Claeys et al. 2014).
Bioprospecting of new lipopeptides
Overriding the vicious drug resistance and facilitating the implication of cyclic lipopeptides in therapeutics, warrants the screening of novel candidates from the biodiversity. Of late, many potent lipopeptides have been isolated from unconventional sources and unexplored niches. Pseudomonas fluorescens BD5, an arctic freshwater bacterium produced the biosurfactants, pseudofactin I and II, when grown on 2 % glucose. The lipopeptides were identified to contain palmitic acid connected to the terminal amino group of an octapeptide moiety. Emulsification activity and stability of pseudofactin II was greater than that of the synthetic surfactants Tween 20 and Triton X-100 (Janek et al. 2010). Bacillus subtilis CSY191, a probiotic strain isolated from Korean traditional fermented soybean paste doenjang produced a surfactin. As verified in MTT assay, the surfactin inhibited the growth of human breast cancer MCF-7 cells. IC50 of 10 μg/ml at 24 h incubation was reported and dose-dependent efficacy was obtained (Lee et al. 2012). For the first time, Citrobacter and Enterobacter were detected to liberate iturins, fengycins, kurstakins and surfactin type lipopeptides (Mandal et al. 2013b). A halophilic Bacillus sp. BS3 produced biosurfactant that conferred anticancer effect on the mammary epithelial carcinoma cell (Donio et al. 2013). It heralded that extreme habitats can be sampled for novel lipopeptides. Empirical findings have implied that the structure and function of microbial biosurfactant largely depends on the substrates in the growth medium. Bacillus amylofaciens AR2 when grown in sucrose medium produced the most effective antifungal lipopeptide. In glucose, sucrose and glycerol-fortified minimal salt medium, surfactin, iturin and fengycin were produced; whereas in maltose, lactose and sorbitol-fortified medium only iturin was produced (Singh et al. 2014a). Iturin A production in fed-batch fermentation on amending the growth medium with the amino acids asparagine, glutamic acid and proline was optimized by artificial neural network-genetic algorithm (ANN-GA). The model enhanced the lipopeptide yield by 34.6 % (Peng et al. 2014).
Emerging roles beyond medicine
Bioremediation
Alkanes and polycyclic aromatic hydrocarbons (PAHs) are xenobiotics posing threats for environment (Kanaly and Harayama 2000). Pseudomonas-elaborated lipopeptides and Bacillus subtilis-elaborated surfactins are known to contribute in hydrocarbon degradation (PAHs) and thus, facilitate bioremediation (Das and Chandran 2011; Pacwa-Płociniczak et al. 2014; Xia et al. 2014). Pseudomonas-produced lipopeptide viscosin exerted as strong mineralization potency as the synthetic surfactant Tween 80. However, the utility is limited owing to its rapid degradation and growth-inhibiting properties (Bak et al. 2014). Marine Brevibacterium luteolum strain produced a lipopeptide that possesses bioremediation ability. Reduction of surface tension was detected to be the mechanism (Vilela et al. 2014). Pseudomonas sp. WJ6 elaborated a lipopeptide biosurfactant mixture of surfactin, fengycin and lichenysin, capable of degrading alkanes and PAHs. This strain might be harnessed to clean oil sludge and oil contaminated soil (Xia et al. 2014). Bacillus licheniformis NIOT-AMKV06 isolated from marine sponge synthesized a lipopeptide with surfactant and emulsifying traits which appeared promising in hydrocarbon degradation (Lawrance et al. 2014).
Agronomy is threatened by a range of fungal pathogens. In this context, the prevention and remediation of plant pathogenesis by lipopeptides is well-substantiated. B. amyloliquefaciens lipopeptide isolated from Japanese fermented food natto eliminated Rhizoctonia solani and Fusarium oxysporum (Murata et al. 2013). Iturins compromised the viability of fungal plant pathogen Verticillium dahliae by upsetting the fungal signaling pathways and enhancing host plant immunity (Han et al. 2014). Bacillus sp. derived lipopeptides surfactin and mycosubtilin (a iturin variant) enhanced defiance of grapevine leaves towards necrotrophic fungus Botrytis cinerea (Farace et al. 2014). The augmentation of host innate immunity by intervention of early signaling pathways was recognized to be the underlying mechanism. B. amyloliquefaciens WH1-produced WH1 fungin inhibited a wide spectrum of fungal pathogens on crop plants (Qi et al. 2010).
Issues to overcome and solutions
Despite their versatile potentials, several impediments exist in optimal utilization of bioactive cyclic lipopeptides. The major adverse responses and roadblocks have been discussed below. Some of the lipopeptides are hemolyins, causing disintegration of red blood cell membranes (Dehghan-Noude et al. 2005; Aranda et al. 2005). Cyanobacterial cyclic lipopeptides, anabaenolysin A and B lysed mammalian cells, enhanced permeability and led to necrosis. It was revealed that the anabaenolysins mimicked the plant-derived glycoside digitonin in rupturing the cholesterol-rich mammalian membranes. Also, the enhanced infiltration stimulated the influx of cyanobacterial toxin nodularin (Oftedal et al. 2012). The recommended daptomycin dose is 4–6 mg/kg; however some infections are not responsive to it and may cause resistance development in patients (Gould et al. 2013). At doses of 8–12 mg/kg, it may prove effective against severe sepsis, caused by Staphylococcus aureus. However, it is not suitable to exceed the dose of 6 mg/kg in patients with obesity and creatinine clearance less than 50 ml/min (Gutiérrez Urbón et al. 2013). Daptomycin and statins when administered together caused creatine phosphokinase elevations, muscle pain and muscle weakness (Bland et al. 2014). Even when exposed to daptomycin of 100-fold MIC concentration, some Staphylococcus aureus SA113 survived. The pathogen viability even at very high dose was correlated to de novo biosynthesis of the amino acids and enhanced pace of TCA cycle (Lechner et al. 2014). Higher percentage of cardiolipin, a diphosphatidylglecerol lipid in the bacterial membrane has been documented to negate the effects of daptomycin. Increased cardiolipin content renders the membrane stiff which antagonizes the flexibility imposed by surfactin. Also cardiolipin with its negative charge repel surfactin and prevent pore formation (Seydlová et al. 2013). A liposome model unraveled that cardiolipin directly restrains the membrane impregnation by daptomycin. Cardiolipin prevents the octameric pore formation by daptomycin (Zhang et al. 2014c). Structural characterization of the lipopeptide and determination of the sequence of amino acid residues in the peptide moiety is vital for drawing the link between structure variation and functional diversity. Edman degradation was the conventional technique for the above purpose, but it was riddled with deficiencies.
The emerging technologies discussed above can be integrated to resolve the pitfalls in optimum utilization of these versatile microbial secondary metabolites (Mitchell 2011; Lynch and Gill 2012). The feasibility of administering PEGylated liposomal daptomycin against MRSA strains was evaluated. This new strategy exerted more sustained effect than conventional liposomal and solution form of daptomycin. This finding built hope that PEGylated liposome-incorporated daptomycin can be employed to eliminate the resistant infections from bloodstream (Huang et al. 2014a, b). A PEGylated lipopeptide with drug-interactive motifs was designed and its efficacy as drug carrier was evaluated. These micelle formulations furnished superior carrier-drug interaction (Zhang et al. 2014a). In targeted and sustained drug delivery, nanotechnology has created a niche. The feasibility of formulating of nano-sized drug delivery vectors from lipopeptides has been reviewed (Rodrigues 2015).
Conclusions
The literature survey drew attention towards several deficient areas in lipopeptide research that could be explored. How substrate composition in growth medium play decisive role in lipopeptide structure merits in-depth study. If these compounds interfere with quorum sensing and biofilm formation of pathogens can be explored. Most importantly, all mechanisms of pathogen armory circumvention need to be unraveled. Unorthodox potential roles of lipopeptides against wide range of ailments must be pursued. The cross-talk between the lipopeptides and immune system deserves deeper investigation. Genome shuffling of the producing microbe, combinatorial biosynthesis and manipulation of pathways to enhance lipopeptide production seems to be promising area. In silico-driven metabolic engineering of the natural lipopeptides for desired functionality holds immense prospects. Their integration in vaccines to boost adjuvancy of antigens might open up new avenues. Many hitherto unkown aspects are emerging, as technical and bioinformatics boom takes over. Invested ample research initiative, these compounds could prove to be treasure trove of robust antibiotic candidates and beyond. We hope this concise review portrays the latest development in the natural cyclic lipopeptides for plausible exploitation.
References
Ajesh K, Sudarslal S, Arunan C, Sreejith K (2013) Kannurin, a novel lipopeptide from Bacillus cereus strain AK1: isolation, structural evaluation and antifungal activities. J Appl Microbiol 115:1287–1296. doi:10.1111/jam.12324
Aktuganov G, Jokela J, Kivelä H et al (2014) Isolation and identification of cyclic lipopeptides from Paenibacillus ehimensis, strain IB-X-b. J Chromatogr B Analyt Technol Biomed Life Sci 973C:9–16. doi:10.1016/j.jchromb.2014.09.042
Aleti G, Sessitsch A, Brader G (2015) Genome mining: prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Comput Struct Biotechnol J 13:192–203. doi:10.1016/j.csbj.2015.03.003
Alexander DC, Rock J, He X et al (2010) Development of a genetic system for combinatorial biosynthesis of lipopeptides in Streptomyces fradiae and heterologous expression of the A54145 biosynthesis gene cluster. Appl Environ Microbiol 76:6877–6887. doi:10.1128/AEM.01248-10
Alexander DC, Rock J, Gu J-Q et al (2011) Production of novel lipopeptide antibiotics related to A54145 by Streptomyces fradiae mutants blocked in biosynthesis of modified amino acids and assignment of lptJ, lptK and lptL gene functions. J Antibiot (Tokyo) 64:79–87. doi:10.1038/ja.2010.138
Ali S, Hameed S, Imran A et al (2014) Genetic, physiological and biochemical characterization of Bacillus sp. strain RMB7 exhibiting plant growth promoting and broad spectrum antifungal activities. Microb Cell Fact 13:144. doi:10.1186/s12934-014-0144-x
Angelini TE, Roper M, Kolter R et al (2009) Bacillus subtilis spreads by surfing on waves of surfactant. Proc Natl Acad Sci USA 106:18109–18113. doi:10.1073/pnas.0905890106
Aranda FJ, Teruel JA, Ortiz A (2005) Further aspects on the hemolytic activity of the antibiotic lipopeptide iturin A. Biochim Biophys Acta 1713:51–56. doi:10.1016/j.bbamem.2005.05.003
Arnold RS, Thom KA, Sharma S et al (2011) Emergence of Klebsiella pneumoniae carbapenemase-producing bacteria. South Med J 104:40–45. doi:10.1097/SMJ.0b013e3181fd7d5a
Bak F, Bonnichsen L, Jørgensen NOG et al (2014) The biosurfactant viscosin transiently stimulates n-hexadecane mineralization by a bacterial consortium. Appl Microbiol Biotechnol. doi:10.1007/s00253-014-6054-3
Baltz RH (2014) Combinatorial biosynthesis of cyclic lipopeptide antibiotics: a model for synthetic biology to accelerate the evolution of secondary metabolite biosynthetic pathways. ACS Synth Biol 3:748–758. doi:10.1021/sb3000673
Baltz RH, Brian P, Miao V, Wrigley SK (2006) Combinatorial biosynthesis of lipopeptide antibiotics in Streptomyces roseosporus. J Ind Microbiol Biotechnol 33:66–74. doi:10.1007/s10295-005-0030-y
Bassetti M, Cadeo B, Villa G et al (2014) Current antibiotic management of prosthetic joint infections in Italy: the “Udine strategy”. J Antimicrob Chemother 69(Suppl 1):i41–i45. doi:10.1093/jac/dku251
Baumgart F, Kluge B, Ullrich C et al (1991) Identification of amino acid substitutions in the lipopeptide surfactin using 2D NMR spectroscopy. Biochem Biophys Res Commun 177:998–1005
Beiras-Fernandez A, Vogt F, Sodian R, Weis F (2010) Daptomycin: a novel lipopeptide antibiotic against Gram-positive pathogens. Infect Drug Resist 3:95–101. doi:10.2147/IDR.S6961
BenMohamed L, Wechsler SL, Nesburn AB (2002) Lipopeptide vaccines–yesterday, today, and tomorrow. Lancet Infect Dis 2:425–431
Bland CM, Bookstaver PB, Lu ZK et al (2014) Musculoskeletal safety outcomes of patients receiving daptomycin with HMG-CoA reductase inhibitors. Antimicrob Agents Chemother 58:5726–5731. doi:10.1128/AAC.02910-14
Chan Tompkins NH, Harnicar SJ (2008) Prescribing trends with daptomycin (cubicin) for the treatment of gram-positive infections. P T 33:282–288
Chen Y-F, Sun T-L, Sun Y, Huang HW (2014) Interaction of daptomycin with lipid bilayers: a lipid extracting effect. Biochemistry 53:5384–5392. doi:10.1021/bi500779g
Chooklin CS, Maneerat S, Saimmai A (2014) Utilization of banana peel as a novel substrate for biosurfactant production by Halobacteriaceae archaeon AS65. Appl Biochem Biotechnol 173:624–645. doi:10.1007/s12010-014-0870-x
Claeys KC, Fiorvento AD, Rybak MJ (2014) A review of novel combinations of colistin and lipopeptide or glycopeptide antibiotics for the treatment of multidrug-resistant Acinetobacter baumannii. Infect Dis Ther. doi:10.1007/s40121-014-0051-9
Cochrane SA, Vederas JC (2014) Lipopeptides from Bacillus and Paenibacillus spp.: a gold mine of antibiotic candidates. Med Res Rev. doi:10.1002/med.21321
Cochrane SA, Findlay B, Vederas JC, Ratemi ES (2014) Key residues in octyl-tridecaptin A1 analogues linked to stable secondary structures in the membrane. ChemBioChem 15:1295–1299. doi:10.1002/cbic.201402024
Curran WV, Leese RA, Jarolmen H et al (2007) Semisynthetic approaches to laspartomycin analogues. J Nat Prod 70:447–450. doi:10.1021/np068062b
Das N, Chandran P (2011) Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol Res Int 2011:941810. doi:10.4061/2011/941810
De Brucker K, Cammue BPA, Thevissen K (2011) Apoptosis-inducing antifungal peptides and proteins. Biochem Soc Trans 39:1527–1532. doi:10.1042/BST0391527
Debono M, Turner WW, LaGrandeur L et al (1995) Semisynthetic chemical modification of the antifungal lipopeptide echinocandin B (ECB): structure–activity studies of the lipophilic and geometric parameters of polyarylated acyl analogs of ECB. J Med Chem 38:3271–3281
DeCrescenzo Henriksen E, Phillips DR, Peterson JBD (2007) Polymyxin E production by P. amylolyticus. Lett Appl Microbiol 45:491–496. doi:10.1111/j.1472-765X.2007.02210.x
Dehghan-Noude G, Housaindokht M, Bazzaz BSF (2005) Isolation, characterization, and investigation of surface and hemolytic activities of a lipopeptide biosurfactant produced by Bacillus subtilis ATCC 6633. J Microbiol 43:272–276
Deleu M, Bouffioux O, Razafindralambo H et al (2003) Interaction of surfactin with membranes: a computational approach. Langmuir 19:3377–3385. doi:10.1021/la026543z
Dey G, Bharti R, Sen R, Mandal M (2014) Microbial amphiphiles: a class of promising new-generation anticancer agents. Drug Discov Today 20:136–146. doi:10.1016/j.drudis.2014.09.006
Dey G, Bharti R, Dhanarajan G et al (2015) Marine lipopeptide Iturin A inhibits Akt mediated GSK3β and FoxO3a signaling and triggers apoptosis in breast cancer. Sci Rep 5:10316. doi:10.1038/srep10316
Dijkmans AC, Wilms EB, Kamerling IMC et al (2014) Colistin: revival of an old polymyxin antibiotic. Ther Drug Monit. doi:10.1097/FTD.0000000000000172
Domalaon R, Yang X, O’Neil J et al (2014) Structure–activity relationships in ultrashort cationic lipopeptides: the effects of amino acid ring constraint on antibacterial activity. Amino Acids 46:2517–2530. doi:10.1007/s00726-014-1806-z
Donio MBS, Ronica SFA, Viji VT et al (2013) Isolation and characterization of halophilic Bacillus sp. BS3 able to produce pharmacologically important biosurfactants. Asian Pac J Trop Med 6:876–883. doi:10.1016/S1995-7645(13)60156-X
Duarte C, Gudiña EJ, Lima CF, Rodrigues LR (2014) Effects of biosurfactants on the viability and proliferation of human breast cancer cells. AMB Express 4:40. doi:10.1186/s13568-014-0040-0
Eeman M, Olofsson G, Sparr E et al (2014) Interaction of fengycin with stratum corneum mimicking model membranes: a calorimetry study. Colloids Surf B Biointerfaces 121:27–35. doi:10.1016/j.colsurfb.2014.05.019
Eschenauer G, Depestel DD, Carver PL (2007) Comparison of echinocandin antifungals. Ther Clin Risk Manag 3:71–97
Falagas ME, Kasiakou SK (2006) Toxicity of polymyxins: a systematic review of the evidence from old and recent studies. Crit Care 10:R27. doi:10.1186/cc3995
Farace G, Fernandez O, Jacquens L et al (2014) Cyclic lipopeptides from Bacillus subtilis activate distinct patterns of defence responses in grapevine. Mol Plant Pathol. doi:10.1111/mpp.12170
Fischer A, Yang S-J, Bayer AS et al (2011) Daptomycin resistance mechanisms in clinically derived Staphylococcus aureus strains assessed by a combined transcriptomics and proteomics approach. J Antimicrob Chemother 66:1696–1711. doi:10.1093/jac/dkr195
Gao Z, Zhao X, Lee S et al (2013) WH1fungin a surfactin cyclic lipopeptide is a novel oral immunoadjuvant. Vaccine 31:2796–2803 doi:10.1016/j.vaccine.2013.04.028.
Gao Z, Yang T, Zhao X et al (2014) Immunomodulation therapy of diabetes by oral administration of a surfactin lipopeptide in NOD mice. Vaccine. doi:10.1016/j.vaccine.2014.08.082
Gill KA, Berrué F, Arens JC, Kerr RG (2014) Isolation and structure elucidation of cystargamide, a lipopeptide from Kitasatospora cystarginea. J Nat Prod 77:1372–1376. doi:10.1021/np500122s
Gong J, Zheng H, Wu Z et al (2009) Genome shuffling: progress and applications for phenotype improvement. Biotechnol Adv 27:996–1005. doi:10.1016/j.biotechadv.2009.05.016
Gong A-D, Li H-P, Yuan Q-S et al (2015) Antagonistic mechanism of iturin A and plipastatin A from Bacillus amyloliquefaciens S76-3 from wheat spikes against Fusarium graminearum. PLoS One 10:e0116871. doi:10.1371/journal.pone.0116871
Gould IM, Miró JM, Rybak MJ (2013) Daptomycin: the role of high-dose and combination therapy for Gram-positive infections. Int J Antimicrob Agents 42:202–210. doi:10.1016/j.ijantimicag.2013.05.005
Guo Q, Dong W, Li S et al (2014) Fengycin produced by Bacillus subtilis NCD-2 plays a major role in biocontrol of cotton seedling damping-off disease. Microbiol Res 169:533–540. doi:10.1016/j.micres.2013.12.001
Gupta S, Govil D, Kakar PN et al (2009) Colistin and polymyxin B: a re-emergence. Indian J Crit Care Med 13:49–53. doi:10.4103/0972-5229.56048
Gutiérrez Urbón JM, Linares Mondéjar P, Martin Herranz I (2013) Daptomycin dosing greater than 6 mg/kg/day depending on pharmacokinetic and pharmacodynamic parameters infections by Staphylococcus aureus. Farm Hosp 37:534–538. doi:10.7399/FH.2013.37.6.742
Hajare SN, Subramanian M, Gautam S, Sharma A (2013) Induction of apoptosis in human cancer cells by a Bacillus lipopeptide bacillomycin D. Biochimie 95:1722–1731. doi:10.1016/j.biochi.2013.05.015
Hajfarajollah H, Mokhtarani B, Noghabi KA (2014) Newly antibacterial and antiadhesive lipopeptide biosurfactant secreted by a probiotic strain, Propionibacterium freudenreichii. Appl Biochem Biotechnol. doi:10.1007/s12010-014-1221-7
Han Q, Wu F, Wang X et al (2014) The bacterial lipopeptide iturins induce Verticillium dahliae cell death by affecting fungal signalling pathways and mediate plant defence responses involved in pathogen-associated molecular pattern-triggered immunity. Environ Microbiol. doi:10.1111/1462-2920.12538
Hashizume H, Nishimura Y (2008) Bioactive natural products (Part O). Stud Nat Prod Chem. doi:10.1016/S1572-5995(08)80016-6
Horn JN, Cravens A, Grossfield A (2013) Interactions between fengycin and model bilayers quantified by coarse-grained molecular dynamics. Biophys J 105:1612–1623. doi:10.1016/j.bpj.2013.08.034
Huang E, Yousef AE (2014a) The lipopeptide antibiotic paenibacterin binds to the bacterial outer membrane and exerts bactericidal activity through cytoplasmic membrane damage. Appl Environ Microbiol 80:2700–2704. doi:10.1128/AEM.03775-13
Huang E, Yousef AE (2014b) Paenibacterin, a novel broad-spectrum lipopeptide antibiotic, neutralises endotoxins and promotes survival in a murine model of Pseudomonas aeruginosa-induced sepsis. Int J Antimicrob Agents 44:74–77. doi:10.1016/j.ijantimicag.2014.02.018
Huang E, Guo Y, Yousef AE (2012) Draft genome sequence of Paenibacillus sp. strain OSY-SE, a bacterium producing the novel broad-spectrum lipopeptide antibiotic paenibacterin. J Bacteriol 194:6306. doi:10.1128/JB.01506-12
Huang E, Guo Y, Yousef AE (2014a) Biosynthesis of the new broad-spectrum lipopeptide antibiotic paenibacterin in Paenibacillus thiaminolyticus OSY-SE. Res Microbiol 165:243–251. doi:10.1016/j.resmic.2014.02.002
Huang X-L, Wu J, Su T-T et al (2014b) Long-circulating liposomal daptomycin enhances protection against systemic methicillin-resistant Staphylococcus aureus infection with improved therapeutic potential. Yao Xue Xue Bao 49:701–710
Janek T, Łukaszewicz M, Rezanka T, Krasowska A (2010) Isolation and characterization of two new lipopeptide biosurfactants produced by Pseudomonas fluorescens BD5 isolated from water from the Arctic Archipelago of Svalbard. Bioresour Technol 101:6118–6123. doi:10.1016/j.biortech.2010.02.109
Janek T, Łukaszewicz M, Krasowska A (2012) Antiadhesive activity of the biosurfactant pseudofactin II secreted by the Arctic bacterium Pseudomonas fluorescens BD5. BMC Microbiol 12:24. doi:10.1186/1471-2180-12-24
Janek T, Krasowska A, Radwańska A, Łukaszewicz M (2013) Lipopeptide biosurfactant pseudofactin II induced apoptosis of melanoma A 375 cells by specific interaction with the plasma membrane. PLoS One 8:e57991. doi:10.1371/journal.pone.0057991
Kanafani ZA, Corey GR (2007) Daptomycin: a rapidly bactericidal lipopeptide for the treatment of Gram-positive infections. Expert Rev Anti Infect Ther 5:177–184. doi:10.1586/14787210.5.2.177
Kanaly RA, Harayama S (2000) Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J Bacteriol 182:2059–2067
Kanj SS, Kanafani ZA (2011) Current concepts in antimicrobial therapy against resistant gram-negative organisms: extended-spectrum beta-lactamase-producing Enterobacteriaceae, carbapenem-resistant Enterobacteriaceae, and multidrug-resistant Pseudomonas aeruginosa. Mayo Clin Proc 86:250–259. doi:10.4065/mcp.2010.0674
Kaya S, Yilmaz G, Kalkan A et al (2013) Treatment of Gram-positive left-sided infective endocarditis with daptomycin. J Infect Chemother 19:698–702. doi:10.1007/s10156-012-0546-9
Kelesidis T (2014) The interplay between daptomycin and the immune system. Front Immunol 5:52. doi:10.3389/fimmu.2014.00052
Kim S-Y, Kim JY, Kim S-H et al (2007) Surfactin from Bacillus subtilis displays anti-proliferative effect via apoptosis induction, cell cycle arrest and survival signaling suppression. FEBS Lett 581:865–871. doi:10.1016/j.febslet.2007.01.059
Kofla G, Ruhnke M (2011) Pharmacology and metabolism of anidulafungin, caspofungin and micafungin in the treatment of invasive candidosis: review of the literature. Eur J Med Res 16:159–166
Kwa A, Kasiakou SK, Tam VH, Falagas ME (2007) Polymyxin B: similarities to and differences from colistin (polymyxin E). Expert Rev Anti Infect Ther 5:811–821. doi:10.1586/14787210.5.5.811
Lal B, Gund VG, Gangopadhyay AK et al (2003) Semisynthetic modifications of hemiaminal function at ornithine unit of mulundocandin, towards chemical stability and antifungal activity. Bioorg Med Chem 11:5189–5198
Landman D, Georgescu C, Martin DA, Quale J (2008) Polymyxins revisited. Clin Microbiol Rev 21:449–465. doi:10.1128/CMR.00006-08
Laverty G, Gorman SP, Gilmore BF (2011) The potential of antimicrobial peptides as biocides. Int J Mol Sci 12:6566–6596. doi:10.3390/ijms12106566
Lawrance A, Balakrishnan M, Joseph TC et al (2014) Functional and molecular characterization of a lipopeptide surfactant from the marine sponge-associated eubacteria Bacillus licheniformis NIOT-AMKV06 of Andaman and Nicobar Islands, India. Mar Pollut Bull 82:76–85. doi:10.1016/j.marpolbul.2014.03.018
Lechner S, Prax M, Lange B et al (2014) Metabolic and transcriptional activities of Staphylococcus aureus challenged with high-doses of daptomycin. Int J Med Microbiol. doi:10.1016/j.ijmm.2014.05.008
Lee JH, Nam SH, Seo WT et al (2012) The production of surfactin during the fermentation of cheonggukjang by potential probiotic Bacillus subtilis CSY191 and the resultant growth suppression of MCF-7 human breast cancer cells. Food Chem 131:1347–1354. doi:10.1016/j.foodchem.2011.09.133
Lim LM, Ly N, Anderson D et al (2010) Resurgence of colistin: a review of resistance, toxicity, pharmacodynamics, and dosing. Pharmacotherapy 30:1279–1291. doi:10.1592/phco.30.12.1279
Lohans CT, van Belkum MJ, Cochrane SA et al (2014) Biochemical, structural, and genetic characterization of tridecaptin A1, an antagonist of Campylobacter jejuni. ChemBioChem 15:243–249. doi:10.1002/cbic.201300595
Lora-Tamayo J, Parra-Ruiz J, Rodríguez-Pardo D et al (2014) High doses of daptomycin (10 mg/kg/d) plus rifampin for the treatment of staphylococcal prosthetic joint infection managed with implant retention: a comparative study. Diagn Microbiol Infect Dis 80:66–71. doi:10.1016/j.diagmicrobio.2014.05.022
Luo C, Liu X, Zhou H et al (2015) Nonribosomal peptide synthase gene clusters for lipopeptide biosynthesis in Bacillus subtilis 916 and their phenotypic functions. Appl Environ Microbiol 81:422–431. doi:10.1128/AEM.02921-14
Lynch SA, Gill RT (2012) Synthetic biology: new strategies for directing design. Metab Eng 14:205–211. doi:10.1016/j.ymben.2011.12.007
Malina A, Shai Y (2005) Conjugation of fatty acids with different lengths modulates the antibacterial and antifungal activity of a cationic biologically inactive peptide. Biochem J 390:695–702. doi:10.1042/BJ20050520
Mandal SM, Barbosa AEAD, Franco OL (2013a) Lipopeptides in microbial infection control: scope and reality for industry. Biotechnol Adv 31:338–345. doi:10.1016/j.biotechadv.2013.01.004
Mandal SM, Sharma S, Pinnaka AK et al (2013b) Isolation and characterization of diverse antimicrobial lipopeptides produced by Citrobacter and Enterobacter. BMC Microbiol 13:152. doi:10.1186/1471-2180-13-152
Martin NI, Hu H, Moake MM et al (2003) Isolation, structural characterization, and properties of mattacin (polymyxin M), a cyclic peptide antibiotic produced by Paenibacillus kobensis M. J Biol Chem 278:13124–13132. doi:10.1074/jbc.M212364200
McConoughey SJ, Howlin R, Granger JF et al (2014) Biofilms in periprosthetic orthopedic infections. Future Microbiol 9:987–1007. doi:10.2217/fmb.14.64
Meena KR, Kanwar SS (2015) Lipopeptides as the antifungal and antibacterial agents: applications in food safety and therapeutics. Biomed Res Int 2015:473050. doi:10.1155/2015/473050
Miao V, Coëffet-Le Gal M-F, Nguyen K et al (2006) Genetic engineering in Streptomyces roseosporus to produce hybrid lipopeptide antibiotics. Chem Biol 13:269–276. doi:10.1016/j.chembiol.2005.12.012
Mitchell W (2011) Natural products from synthetic biology. Curr Opin Chem Biol 15:505–515. doi:10.1016/j.cbpa.2011.05.017
Mittenbühler K, Loleit M, Baier W et al (1997) Drug specific antibodies: T-cell epitope-lipopeptide conjugates are potent adjuvants for small antigens in vivo and in vitro. Int J Immunopharmacol 19:277–287
Moyle PM, Toth I (2008) Self-adjuvanting lipopeptide vaccines. Curr Med Chem 15:506–516
Murata D, Sawano S, Ohike T et al (2013) Isolation of antifungal bacteria from Japanese fermented soybeans, natto. J Environ Sci (China) 25(Suppl 1):S127–S131. doi:10.1016/S1001-0742(14)60641-0
Naing KW, Lee YS, Nguyen XH et al (2015) Isolation and characterization of an antimicrobial lipopeptide produced by Paenibacillus ehimensis MA2012. J Basic Microbiol. doi:10.1002/jobm.201400505
Nasir MN, Laurent P, Flore C et al (2013) Analysis of calcium-induced effects on the conformation of fengycin. Spectrochim Acta A Mol Biomol Spectrosc 110:450–457. doi:10.1016/j.saa.2013.03.063
Nerurkar AS (2010) Structural and molecular characteristics of lichenysin and its relationship with surface activity. Adv Exp Med Biol 672:304–315
Ochi K, Hosaka T (2013) New strategies for drug discovery: activation of silent or weakly expressed microbial gene clusters. Appl Microbiol Biotechnol 97:87–98. doi:10.1007/s00253-012-4551-9
Oftedal L, Myhren L, Jokela J et al (2012) The lipopeptide toxins anabaenolysin A and B target biological membranes in a cholesterol-dependent manner. Biochim Biophys Acta 1818:3000–3009. doi:10.1016/j.bbamem.2012.07.015
Ongena M, Jacques P (2008) Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol 16:115–125. doi:10.1016/j.tim.2007.12.009
Ortíz-López FJ, Monteiro MC, González-Menéndez V et al (2015) Cyclic colisporifungin and linear cavinafungins antifungal lipopeptides isolated from Colispora cavincola. J Nat Prod. doi:10.1021/np500854j
Pacwa-Płociniczak M, Płaza GA, Poliwoda A, Piotrowska-Seget Z (2014) Characterization of hydrocarbon-degrading and biosurfactant-producing Pseudomonas sp. P-1 strain as a potential tool for bioremediation of petroleum-contaminated soil. Environ Sci Pollut Res Int 21:9385–9395. doi:10.1007/s11356-014-2872-1
Pan H, Zhao X, Gao Z, Qi G (2014) A surfactin lipopeptide adjuvanted hepatitis B vaccines elicit enhanced humoral and cellular immune responses in mice. Protein Pept Lett 21:901–910
Pappas G, Ierodiakonou V, Falagas ME (2009) Lost in translation: differences in antimicrobial indication approval policies between the United States and Europe. Clin Ther 31:1595–1603. doi:10.1016/j.clinthera.2009.06.016
Patel H, Huynh Q, Bärlehner D, Heerklotz H (2014) Additive and synergistic membrane permeabilization by antimicrobial (lipo)peptides and detergents. Biophys J 106:2115–2125. doi:10.1016/j.bpj.2014.04.006
Pathak KV, Keharia H (2013) Identification of surfactins and iturins produced by potent fungal antagonist, Bacillus subtilis K1 isolated from aerial roots of banyan (Ficus benghalensis) tree using mass spectrometry. 3. Biotech 4:283–295. doi:10.1007/s13205-013-0151-3
Peel TN, Buising KL, Choong PFM (2012) Diagnosis and management of prosthetic joint infection. Curr Opin Infect Dis 25:670–676. doi:10.1097/QCO.0b013e32835915db
Peng W, Zhong J, Yang J et al (2014) The artificial neural network approach based on uniform design to optimize the fed-batch fermentation condition: application to the production of iturin A. Microb Cell Fact 13:54. doi:10.1186/1475-2859-13-54
Qazi MA, Kanwal T, Jadoon M et al (2014) Isolation and characterization of a biosurfactant-producing Fusarium sp. BS-8 from oil contaminated soil. Biotechnol Prog 30:1065–1075. doi:10.1002/btpr.1933
Qi G, Zhu F, Du P et al (2010) Lipopeptide induces apoptosis in fungal cells by a mitochondria-dependent pathway. Peptides 31:1978–1986. doi:10.1016/j.peptides.2010.08.003
Qian C-D, Liu T-Z, Zhou S-L et al (2012a) Identification and functional analysis of gene cluster involvement in biosynthesis of the cyclic lipopeptide antibiotic pelgipeptin produced by Paenibacillus elgii. BMC Microbiol 12:197. doi:10.1186/1471-2180-12-197
Qian C-D, Wu X-C, Teng Y et al (2012b) Battacin (Octapeptin B5), a new cyclic lipopeptide antibiotic from Paenibacillus tianmuensis active against multidrug-resistant Gram-negative bacteria. Antimicrob Agents Chemother 56:1458–1465. doi:10.1128/AAC.05580-11
Raaijmakers JM, De Bruijn I, Nybroe O, Ongena M (2010) Natural functions of lipopeptides from Bacillus and Pseudomonas: more than surfactants and antibiotics. FEMS Microbiol Rev 34:1037–1062. doi:10.1111/j.1574-6976.2010.00221.x
Rautela R, Singh AK, Shukla A, Cameotra SS (2014) Lipopeptides from Bacillus strain AR2 inhibits biofilm formation by Candida albicans. Antonie Van Leeuwenhoek 105:809–821. doi:10.1007/s10482-014-0135-2
Reder-Christ K, Schmidt Y, Dörr M et al (2012) Model membrane studies for characterization of different antibiotic activities of lipopeptides from Pseudomonas. Biochim Biophys Acta 1818:566–573. doi:10.1016/j.bbamem.2011.08.007
Robbel L, Marahiel MA (2010) Daptomycin, a bacterial lipopeptide synthesized by a nonribosomal machinery. J Biol Chem 285:27501–27508. doi:10.1074/jbc.R110.128181
Rodrigues LR (2015) Microbial surfactants: fundamentals and applicability in the formulation of nano-sized drug delivery vectors. J Colloid Interface Sci. doi:10.1016/j.jcis.2015.01.022
Roongsawang N, Washio K, Morikawa M (2010) Diversity of nonribosomal peptide synthetases involved in the biosynthesis of lipopeptide biosurfactants. Int J Mol Sci 12:141–172. doi:10.3390/ijms12010141
Rückert C, Szczepanowski R, Albersmeier A et al (2014) Complete genome sequence of the actinobacterium Actinoplanes friuliensis HAG 010964, producer of the lipopeptide antibiotic friulimycin. J Biotechnol 178:41–42. doi:10.1016/j.jbiotec.2014.03.011
Sagar SM, Yance D, Wong RK (2006) Natural health products that inhibit angiogenesis: a potential source for investigational new agents to treat cancer-Part 1. Curr Oncol 13:14–26
Sakoulas G, Moise PA, Casapao AM et al (2014) Antimicrobial salvage therapy for persistent staphylococcal bacteremia using daptomycin plus ceftaroline. Clin Ther 36:1317–1333. doi:10.1016/j.clinthera.2014.05.061
Salunke DB, Connelly SW, Shukla NM et al (2013) Design and development of stable, water-soluble, human Toll-like receptor 2 specific monoacyl lipopeptides as candidate vaccine adjuvants. J Med Chem 56:5885–5900. doi:10.1021/jm400620g
Schneider T, Gries K, Josten M et al (2009) The lipopeptide antibiotic Friulimicin B inhibits cell wall biosynthesis through complex formation with bactoprenol phosphate. Antimicrob Agents Chemother 53:1610–1618. doi:10.1128/AAC.01040-08
Schneider T, Müller A, Miess H, Gross H (2014) Cyclic lipopeptides as antibacterial agents—potent antibiotic activity mediated by intriguing mode of actions. Int J Med Microbiol 304:37–43. doi:10.1016/j.ijmm.2013.08.009
Seydlová G, Fišer R, Cabala R et al (2013) Surfactin production enhances the level of cardiolipin in the cytoplasmic membrane of Bacillus subtilis. Biochim Biophys Acta 1828:2370–2378. doi:10.1016/j.bbamem.2013.06.032
Shao C, Liu L, Gang H et al (2015) Structural diversity of the microbial surfactin derivatives from selective esterification approach. Int J Mol Sci 16:1855–1872. doi:10.3390/ijms16011855
Sharma D, Mandal SM, Manhas RK (2014) Purification and characterization of a novel lipopeptide from Streptomyces amritsarensis sp. nov. active against methicillin-resistant Staphylococcus aureus. AMB Express 4:50. doi:10.1186/s13568-014-0050-y
Shen K-Y, Chang L-S, Leng C-H, Liu S-J (2015) Self-adjuvanting lipoimmunogens for therapeutic HPV vaccine development: potential clinical impact. Expert Rev Vaccines 14:383–394. doi:10.1586/14760584.2015.966696
Shiono Y, Tsuchinari M, Shimanuki K et al (2007) Fusaristatins A and B, two new cyclic lipopeptides from an endophytic Fusarium sp. J Antibiot (Tokyo) 60:309–316. doi:10.1038/ja.2007.39
Shoemaker DM, Simou J, Roland WE (2006) A review of daptomycin for injection (Cubicin) in the treatment of complicated skin and skin structure infections. Ther Clin Risk Manag 2:169–174
Silverman JA, Perlmutter NG, Shapiro HM (2003) Correlation of daptomycin bactericidal activity and membrane depolarization in Staphylococcus aureus. Antimicrob Agents Chemother 47:2538–2544
Singh AK, Rautela R, Cameotra SS (2014a) Substrate dependent in vitro antifungal activity of Bacillus sp. strain AR2. Microb Cell Fact 13:67. doi:10.1186/1475-2859-13-67
Singh LS, Sharma H, Talukdar NC (2014b) Production of potent antimicrobial agent by actinomycete, Streptomyces sannanensis strain SU118 isolated from phoomdi in Loktak Lake of Manipur, India. BMC Microbiol 14:278. doi:10.1186/s12866-014-0278-3
Sivapathasekaran C, Das P, Mukherjee S et al (2010) Marine bacterium derived lipopeptides: characterization and cytotoxic activity against cancer cell lines. Int J Pept Res Ther 16:215–222. doi:10.1007/s10989-010-9212-1
Song C, Aundy K, van de Mortel J, Raaijmakers JM (2014) Discovery of new regulatory genes of lipopeptide biosynthesis in Pseudomonas fluorescens. FEMS Microbiol Lett 356:166–175
Steenbergen JN, Alder J, Thorne GM, Tally FP (2005) Daptomycin: a lipopeptide antibiotic for the treatment of serious Gram-positive infections. J Antimicrob Chemother 55:283–288. doi:10.1093/jac/dkh546
Steinhagen F, Kinjo T, Bode C, Klinman DM (2011) TLR-based immune adjuvants. Vaccine 29:3341–3355. doi:10.1016/j.vaccine.2010.08.002
Strano CP, Bella P, Licciardello G et al (2015) Pseudomonas corrugata crpCDE is part of the cyclic lipopeptide corpeptin biosynthetic gene cluster and is involved in bacterial virulence in tomato and in hypersensitive response in Nicotiana benthamiana. Mol Plant Pathol. doi:10.1111/mpp.12207
Strieker M, Tanović A, Marahiel MA (2010) Nonribosomal peptide synthetases: structures and dynamics. Curr Opin Struct Biol 20:234–240. doi:10.1016/j.sbi.2010.01.009
Sun H, Bie X, Lu F et al (2009) Enhancement of surfactin production of Bacillus subtilis fmbR by replacement of the native promoter with the Pspac promoter. Can J Microbiol 55:1003–1006. doi:10.1139/w09-044
Sun H, Liu Z, Zhao H, Ang EL (2015) Recent advances in combinatorial biosynthesis for drug discovery. Drug Des Devel Ther 9:823–833. doi:10.2147/DDDT.S63023
Takeuchi M, Ashihara E, Yamazaki Y et al (2011) Rakicidin A effectively induces apoptosis in hypoxia adapted Bcr-Abl positive leukemic cells. Cancer Sci 102:591–596. doi:10.1111/j.1349-7006.2010.01813.x
Tang J, Gao H, Dai Y et al (2008) Progress on the studies of cyclic lipopeptides. Yao Xue Xue Bao 43:873–883
Tareq FS, Lee MA, Lee H-S et al (2014) Gageotetrins A–C, noncytotoxic antimicrobial linear lipopeptides from a marine bacterium Bacillus subtilis. Org Lett 16:928–931. doi:10.1021/ol403657r
Thasana N, Prapagdee B, Rangkadilok N et al (2010) Bacillus subtilis SSE4 produces subtulene A, a new lipopeptide antibiotic possessing an unusual C15 unsaturated beta-amino acid. FEBS Lett 584:3209–3214. doi:10.1016/j.febslet.2010.06.005
Thies S, Santiago-Schübel B, Kovačić F et al (2014) Heterologous production of the lipopeptide biosurfactant serrawettin W1 in Escherichia coli. J Biotechnol 181:27–30. doi:10.1016/j.jbiotec.2014.03.037
Tirilomis T (2014) Daptomycin and its immunomodulatory effect: consequences for antibiotic treatment of methicillin-resistant Staphylococcus aureus wound infections after heart surgery. Front Immunol 5:97. doi:10.3389/fimmu.2014.00097
Toth I, Simerska P, Fujita Y (2008) Recent advances in design and synthesis of self-adjuvanting lipopeptide vaccines. Int J Pept Res Ther 14:333–340. doi:10.1007/s10989-008-9147-y
Velkov T, Roberts KD, Nation RL et al (2013) Pharmacology of polymyxins: new insights into an “old” class of antibiotics. Future Microbiol 8:711–724. doi:10.2217/fmb.13.39
Vilela WFD, Fonseca SG, Fantinatti-Garboggini F et al (2014) Production and properties of a surface-active lipopeptide produced by a new marine Brevibacterium luteolum strain. Appl Biochem Biotechnol. doi:10.1007/s12010-014-1208-4
Walsh CT (2002) Combinatorial biosynthesis of antibiotics: challenges and opportunities. ChemBioChem 3:125–134
Wang CL, Ng TB, Cao XH et al (2009) CLP induces apoptosis in human leukemia K562 cells through Ca(2+) regulating extracellular-related protein kinase ERK activation. Cancer Lett 276:221–227. doi:10.1016/j.canlet.2008.11.007
Wang F, Ren N-N, Luo S et al (2014) DptR2, a DeoR-type auto-regulator, is required for daptomycin production in Streptomyces roseosporus. Gene 544:208–215. doi:10.1016/j.gene.2014.04.044
Xia W, Du Z, Cui Q et al (2014) Biosurfactant produced by novel Pseudomonas sp. WJ6 with biodegradation of n-alkanes and polycyclic aromatic hydrocarbons. J Hazard Mater 276:489–498. doi:10.1016/j.jhazmat.2014.05.062
Xing Y, Wang W, Dai S et al (2014) Daptomycin exerts rapid bactericidal activity against Bacillus anthracis without disrupting membrane integrity. Acta Pharmacol Sin 35:211–218. doi:10.1038/aps.2013.159
Yamanaka K, Reynolds KA, Kersten RD et al (2014) Direct cloning and refactoring of a silent lipopeptide biosynthetic gene cluster yields the antibiotic taromycin A. Proc Natl Acad Sci U S A 111:1957–1962. doi:10.1073/pnas.1319584111
Yang S-Z, Wei D-Z, Mu B-Z (2006) Determination of the amino acid sequence in a cyclic lipopeptide using MS with DHT mechanism. J Biochem Biophys Methods 68:69–74. doi:10.1016/j.jbbm.2006.03.008
Yang H, Chen G, Hu L et al (2014a) In vivo activity of daptomycin/colistin combination therapy in a Galleria mellonella model of Acinetobacter baumannii infection. Int J Antimicrob Agents. doi:10.1016/j.ijantimicag.2014.10.012
Yang H-J, Huang X-Z, Zhang Z-L et al (2014b) Two novel amphomycin analogues from Streptomyces canus strain FIM-0916. Nat Prod Res 28:861–867. doi:10.1080/14786419.2014.886210
Yoshino N, Endo M, Kanno H et al (2013) Polymyxins as novel and safe mucosal adjuvants to induce humoral immune responses in mice. PLoS One 8:e61643. doi:10.1371/journal.pone.0061643
Yu Z, Qin W, Lin J et al (2015) Antibacterial mechanisms of polymyxin and bacterial resistance. Biomed Res Int 2015:679109. doi:10.1155/2015/679109
Yuste JR, Quesada M, Díaz-Rada P, Del Pozo JL (2014) Daptomycin in the treatment of prosthetic joint infection by Enterococcus faecalis: safety and efficacy of high-dose and prolonged therapy. Int J Infect Dis 27:65–66. doi:10.1016/j.ijid.2014.05.034
Zaman M, Toth I (2013) Immunostimulation by synthetic lipopeptide-based vaccine candidates: structure–activity relationships. Front Immunol 4:318. doi:10.3389/fimmu.2013.00318
Zeriouh H, Romero D, Garcia-Gutierrez L et al (2011) The iturin-like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Mol Plant Microbe Interact 24:1540–1552. doi:10.1094/MPMI-06-11-0162
Zhang A, Sun H, Wang P et al (2012) Modern analytical techniques in metabolomics analysis. Analyst 137:293–300. doi:10.1039/c1an15605e
Zhang B, Dong C, Shang Q et al (2013a) New insights into membrane-active action in plasma membrane of fungal hyphae by the lipopeptide antibiotic bacillomycin L. Biochim Biophys Acta 1828:2230–2237. doi:10.1016/j.bbamem.2013.05.033
Zhang T, Muraih JK, Mintzer E et al (2013b) Mutual inhibition through hybrid oligomer formation of daptomycin and the semisynthetic lipopeptide antibiotic CB-182,462. Biochim Biophys Acta 1828:302–308. doi:10.1016/j.bbamem.2012.10.008
Zhang P, Lu J, Huang Y et al (2014a) Design and evaluation of a PEGylated lipopeptide equipped with drug-interactive motifs as an improved drug carrier. AAPS J 16:114–124. doi:10.1208/s12248-013-9536-9
Zhang T, Muraih JK, MacCormick B et al (2014b) Daptomycin forms cation- and size-selective pores in model membranes. Biochim Biophys Acta 1838:2425–2430. doi:10.1016/j.bbamem.2014.05.014
Zhang T, Muraih JK, Tishbi N et al (2014c) Cardiolipin prevents membrane translocation and permeabilization by daptomycin. J Biol Chem 289:11584–11591. doi:10.1074/jbc.M114.554444
Zhao J, Li Y, Zhang C et al (2012) Genome shuffling of Bacillus amyloliquefaciens for improving antimicrobial lipopeptide production and an analysis of relative gene expression using FQ RT-PCR. J Ind Microbiol Biotechnol 39:889–896. doi:10.1007/s10295-012-1098-9
Zhao J, Cao L, Zhang C et al (2014) Differential proteomics analysis of Bacillus amyloliquefaciens and its genome-shuffled mutant for improving surfactin production. Int J Mol Sci 15:19847–19869. doi:10.3390/ijms151119847
Conflict of interest
We declare no conflict of interest in submission of this manuscript.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Patel, S., Ahmed, S. & Eswari, J.S. Therapeutic cyclic lipopeptides mining from microbes: latest strides and hurdles. World J Microbiol Biotechnol 31, 1177–1193 (2015). https://doi.org/10.1007/s11274-015-1880-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11274-015-1880-8