
Abstract
Relations between vegetation characteristics and eco-hydrological processes were assessed in a pristine mire in the valley of the Ob River (Western Siberia). Along a transect from the terrace scarp to the river, field data were collected on vegetation composition, peat stratigraphy, peat chemistry, hydrology and hydrochemistry. Based on floristic composition, eight vegetation communities were distinguished. Hydraulic head measurements were used to obtain an indication of groundwater flow directions. The water balance of the mire was calculated with a two-dimensional steady-state numerical groundwater model. Water types were defined based on cluster analysis of hydrochemical data. The results revealed that the dominant hydrological factor in the Ob mire is the discharge of groundwater, which supplies about threefold more water than net precipitation. Although the discharge flux decreases with increasing distance from the terrace scarp, high water levels and a “groundwater-like” mire water composition were observed in the major part of the study site. Precipitation and river water play only a minor role. Despite dilution of discharging groundwater with rainwater, spatial differences in pH and solute concentrations of the surficial mire water are small and not reflected in the vegetation composition. Although small amounts of silt and clay were found in the peat in the proximity of the river, indicating the occurrence of river floods in former times, no river-flood zone could be recognized based on hydrochemical characteristics or vegetation composition. A comparison of the Ob mire with well-studied and near-natural mires in the Biebrza River valley (Poland) revealed substantial differences in both vegetation characteristics and the intensity and spatial pattern of eco-hydrological processes. Differences in the origin and ratios of water fluxes as well as a dissimilar land use history would seem to be key factors explaining the differences observed.
Similar content being viewed by others

Introduction
A study of mires reveals close relations between topography, hydrology, hydrochemistry and vegetation. In general, differences in the topographical and hydrological positions of mires give rise to different types of mire water, especially in terms of nutrient and calcium concentrations and acidity. This in turn largely determines the floristic composition and diversity of the vegetation of the mire (Kulczyński 1949; Sjörs 1950; Moore and Bellamy 1974; Succow and Joosten 2001). Mires in river valleys exhibit distinct vegetation zones, both parallel and perpendicular to the river axis (Ratcliffe 1977; Pałczyński 1984), which have been shown to reflect different ratios in the supplies of groundwater, surface (river) water and precipitation (Oświt 1968; Wassen et al. 1992, 2002b; Wassen and Joosten 1996). As in the temperate zones of Europe and European Russia, North America and Eastern Asia (China, Japan), these mires have largely been drained for agriculture (Joosten and Clarke 2002), and few studies have addressed the relations between water flow, water chemistry and vegetation in pristine river valley mires (cf. Wassen et al. 1992; Wassen and Joosten 1996).
In the sub-taiga zone of Western Siberia, vast complexes of largely undisturbed mires still prevail. The objective of this article is to describe the relations between vegetation characteristics and eco-hydrological processes in a pristine mire in the Ob River valley, close to Tomsk. Such a descriptive study contributes to our understanding of the eco-hydrological functioning of mires, which in turn is highly relevant for the conservation or restoration of these systems. Because a comparison with other mires might reveal general patterns, we compare our findings in the Ob mire with the characteristics of the well-studied, anthropogenically little affected mires in the Biebrza River valley in Poland (Wassen et al. 1992; Wassen and Joosten 1996). Similarities and differences are discussed, and hypotheses are formulated for the differences observed.
Methods
Study area
The West-Siberian Plain is the largest peatland region of the world, with mires covering over 10,00,000 km2 (Yefremov and Yefremova 2001; Tanneberger et al. 2003; Smith et al. 2004). The plain is situated between the Ural Mountains to the west, the Yenisey River to the east and the Altai Mountains to the south. The south-eastern part of the West-Siberian Plain consists of an alluvial plain that slightly dips in a north-western direction. Paleozoic rock was covered with layers of loam, silt and clay up to 250 m thick during the Mesozoic and Cenozoic. The area was never glaciated during the Quaternary (Franz 1973), but was dissected by fluvial erosion when the Ob River cut stepwise into the alluvial surface. The total width of the Ob valley reaches 30–60 km and includes four to five river terraces.
The climate of the area is continental, with cold winters (mean January temperature: −19.6°C) and warm summers (mean July temperature: 18.0°C) (Evseeva 2001). Mean annual precipitation is 415 mm, 70% of which falls in the frost-free period (May–September). The average snow cover is 60 cm during a period of 180 days (Atlas of the USSR 1983); permafrost does not occur (Walter and Breckle 1994).
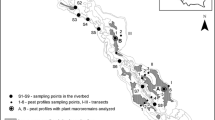
The study site (56°30′N 84°01′E; Fig. 1) is located approximately 10 km south of Melnikovo, 50 km west of Tomsk and 250 km north of Novosibirsk, in the floodplain of the Ob River. Gyttja accumulation started in oxbow lakes from 9000 to 8000 BP; mire development began in 7000 BP (Lapshina 1987; Blyakharchuk 2003). The current surface of the mire, which is bordered by a steep terrace scarp to the west, slopes down over a width of approximately 3 km from the foot of the scarp (ca. 95.0 m a.s.l.) to the banks of the Sargatka River (ca. 91.5 m a.s.l.). The Sargatka River is a left-bank tributary of the Ob River, flowing between the study site and the levees of the Ob River and discharging superfluous mire water (Fig. 1). Spring mounds have formed along the steep scarp (Lapshina 1987). The western part of the study site contains open fen vegetation with sparse growth of shrubs and trees. In the central part, peat ridges of varying length and with a sparse tree cover of Pinus sylvestris, Betula pubescens and Picea obovata run parallel to the Ob River. The mire is forested in the vicinity of the Ob River. In large parts of the mire, the vegetation and root layer almost floats on a watery layer with little dead organic matter. The transition between this watery top layer and the deeper, denser peat cannot be defined clearly everywhere. The uplands are characterized by a mosaic of Betula pendula-Populus tremula forests and agricultural fields or pastures that are largely abandoned since the collapse of the collective farms. In earlier times, floods from the Ob River occasionally reached the terrace scarp (personal communication with residents, 2002). Since 1969, the Novosibirsk hydroelectric power plant reservoir, located 300 km upstream from the study area, has prevented flooding of the site (Lapshina 1987).

Location of the study area and sampling locations (0–9)
Data collection
For detailed study of the relations between vegetation, hydrology and hydrochemistry, a transect (length: 3.2 km) was selected from the terrace scarp to the levee of the Sargatka tributary (Fig. 1), crossing the main variation in vegetation types. Along the transect, surface elevation was measured with a water-level device. To determine peat stratigraphy, we made 23 corings using a chamber peat corer (diameter: 4.5 cm; length: 50 cm) in steps of 200 m (maximum) . Field identification of peat and sediments followed Boden (1996), Stegmann et al. (2001) and Succow and Stegmann (2001). The core from sampling location 1 (Fig. 1) was analysed for macrofossils following Michaelis (2002).
Using a modified Braun-Blanquet scale (Wilmanns 1998), in July 2002 we recorded 105 vegetation relevés (25 m2; cf. Dierschke 1994) that were located within a radius of 25 m from the centres of the sampling locations (indicated as 0–9 on Fig. 1). The nomenclature of the angiosperm species followed Rothmaler (2002) and Krasnoborov (2000) (Salix rosmarinifolia); that of moss species was according to Frahm and Frey (1992). To estimate actual biomass production, we harvested the above-ground standing crop of vascular plants at the peak of the growing season (July 4–12) at sampling locations 0–5. Because representative biomass production samples could not be obtained from sites with substantial tree cover, the forested part of the mire was not sampled (sampling locations 6–9). For each biomass sample, three separate plots were selected of 0.16 m2 each; these were located within a radius of 10 m from the centres of the sampling locations. After removal of the dead parts, the above-ground vascular plant material was stored in polyethylene bags, dried for 48 h at 70°C and weighed immediately after drying. Kjeldahl destruction (Allen 1989) was carried out on the dried and homogenized plant material in order to determine the contents of nitrogen (N) (with an auto-analyser, AA), phosphorus (P) and potassium (K) (with the inductively coupled plasma technique, ICP).
Peat samples were collected from the vegetation relevé sites at a depth of 10–15 cm below the peat surface and stored in polyethylene bags at 5°C. After 2–4 weeks, samples were dried first for 1–2 days under direct sunlight, followed by 12 h in an oven at 105°C. After 20 weeks, samples were analysed for pH and CaCO3 content. pH was determined in rewetted samples in 0.01 M CaCl2 using WTW-pH 96 (Rowell 1997). CaCO3 content (%) was determined in ground samples with 10% HCl using a Scheibler-apparatus (Schlichting et al. 1995). Total carbon (C) and total N were measured by Dumas digestion with a C/N-analyser (Elementar Vario-EL, Germany). Corg/N ratios were calculated after subtracting the inorganic C – determined with the Scheibler-device – from the total C.
Piezometers were placed at each of sampling locations 1–9, with filters (length: 30 cm) at depths of 0.5, 2 and – when possible – 4 and 6 m below the mire surface. Between June 1 and August 25, 2002, eight sampling campaigns were carried out to measure piezometric heads and surficial mire water levels and to collect water samples. Piezometric heads and surficial mire water levels were measured relative to the mire surface and related to each other with the elevation data of the levelling survey. Water samples were taken from the piezometers, from the surficial mire water at all sampling locations and from the Ob River, using a hand pump connected to a polyethylene Erlenmeyer. Piezometers were emptied and allowed to refill with fresh groundwater once before sampling. Water samples were collected in polyethylene bottles (50 ml) that were filled to the brim to limit aeration. Electrical conductivity (EC) and pH were measured in the field using WTW-LF91 with an Ag/AgCl-electrode and WTW-pH96, respectively. HCO3 concentrations were determined within 24 h after sampling by acidimetric titration to pH 4.3 (Aquamerck Alkalinity field set; Merck, Germany). Samples were conserved by acidification to pH 1 and stored for a maximum of 15 weeks before being analysed for Ca, Fe, K, Mg, Mn, Na, Si and SO4 concentrations using the ICP technique and for concentrations of Cl, NH4, NO3 and PO4 using the AA. Net precipitation in the study area was estimated by measuring water level changes in open-water pans (diameter: 12 cm). Saturated hydraulic conductivity of the peat was determined by slug tests following Hvorslev (1951; in Domenico and Schwarz 1998).
Data processing
Plant communities were classified by ordination of the relevés into non-hierarchical species groups based on resemblances and dissimilarities in floristic composition and dominance of species (Koska et al. 2001); these were then named on the basis of two or three characteristic species. Nutrient limitation was determined by calculating N:P, N:K and K:P ratios in the above-ground biomass of vascular plants (Olde Venterink et al. 2003). Peat and water chemical data of locations characterized by the same floristic vegetation type were combined, and mean values were calculated.
Surficial mire water level measurements and piezometric heads were used to reveal water flow directions in the mire. To determine the water balance of the mire, we constructed a two-dimensional steady-state numerical groundwater model (cell size: 20 m) with the MODFLOW code (Harbaugh et al. 2000). The groundwater body in the terrace plateau was simulated as a constant head boundary at a distance of 700 m from the scarp using estimates of the phreatic water level in a small pond in the plateau to determine the head level. At the eastern model border, the Sargatka River was simulated by applying a constant head of 89.2 m a.s.l. in accordance with the water level recorded during the surface levelling measurements. Precipitation data from a nearby climate station (hydrological year 2001–2002) and evapotranspiration rates calculated according to the Penman-Monteith model were used to determine net precipitation input. The model includes a sand layer (K h = 25 m d−1; K v = 10 m d−1) overlain by a clay-loam layer (K h = K v = 0.1 m d−1), a peat layer (K h = 2 m d−1; K v = 1 m d−1), a root zone layer (K h = 100 m d−1; K v = 50 m d−1) and, on top, an air layer with high conductivity (K h = K v = 50,000 m d−1). For the terrace plateau, conductivity values of K h = 10 m d−1 and K v = 5 m d−1 were assumed. The model was calibrated on piezometric heads and surficial mire water levels. Based on the model calibration, the hydraulic conductivity of the peat layer was set slightly higher than the value measured in the field (i.e. K = 0.695 (±0.96) m d−1; n = 6).
Reliability of the hydrochemical data was tested by charge balance calculations. Deviations from electro-neutrality were within 10% for 92% of the water samples, indicating that the reliability of the data is sufficient to explore differences between main water types (cf. Schot and Van der Wal 1992). Water types were distinguished by agglomerative hierarchical clustering of the water samples using the between-groups linkage method as clustering algorithm with the Euclidian distance as measure for similarity (SPSS ver. 14.0; SPSS, Chicago, Ill.). Since pH was measured less frequently than the other hydrochemical variables, this variable was discarded. Clustering is thus based on Ca, Cl, EC, Fe, HCO3, K, Mg, Mn, Na, NH4, NO3, PO4, Si and SO4.
Results
Peat stratigraphy
The cores revealed that the peat body (thickness: 3.5–7.5 m) is underlain by a gyttja (thickness: 0.5–3.0 m) over an undulating layer of silt and clay (Fig. 2). In the western part of the transect (0–1500 m from the scarp) loose, calcareous and slightly decomposed brownmoss peats predominantly formed of Drepanocladus aduncus prevail. These brownmoss peats alternate with radicel peat – i.e. peat predominantly formed of sedge rootlets. Silt deposits with a thickness of up to 1.5 m and thin silt layers intersecting the peat were found near the scarp. In the eastern part of the transect (1500–2800 m from the scarp), brownmoss peats are nearly absent and radicel peats with some wood remains predominate. Wood peats were exclusively found at depths of 0.5–1.5 m in a narrow belt of approximately 250 m adjacent to the river. Radicel peats with living roots and rhizomes form the uppermost part of the peat layer, also at the peat ridges in the central part of the mire. Small amounts of silt and clay were found in the peat in the proximity of the river.

Stratigraphy of the Ob mire transect
Vegetation characteristics and peat chemistry
Eight vegetation communities were distinguished (Table 1). The species-poor community – denoted community A (the Phragmites australis-Angelica archangelica community) – dominated by P. australis lacks many species common to all other vegetation communities. It is located close to the terrace scarp. Community B (the Betula pubescens-Menyanthes trifoliata- Parnassia palustris community) has a well-developed B. pubescens layer similar to communities F, G and H, but the former is distinguished from the latter by the presence of P. australis and Parnassia palustris. Communities C, D and E consist of open vegetation with abundant sedges (e.g. Carex lasiocarpa, C. rostrata), Menyanthes trifoliata and a closed-cover brownmoss layer (mainly Drepanocladus aduncus). Communities C and E also contain dwarf shrubs (e.g.Betula humilis). In addition, community C (Carex lasiocarpa-Betula humilis community) contains Salix rosmarinifolia, whereas in community E (Carex appropinquata-Typha latifolia community) T. latifolia occurs at a high constancy. Community D (Carex lasiocarpa-Parnassia palustris community) contains some Pinus sylvestris. The communities in the eastern part of the study site (communities F, G and H) have a well-developed tree layer (Betula pubescens, Pinus sylvestris). Community F (Betula pubescens-Menyanthes trifoliata-Pinus sylvestris community) is distinguished by the absence of Carex lasiocarpa. Whereas Pinus sylvestris and Menyanthes trifoliata are common in communities F and G (Pinus sylvestris-Carex appropinquata-Calamagrostis stricta community), they do not occur with high abundance in community H (Betula pubescens-Salix pentandra community).
Species richness is highest in communities B, F and G. Community B combines a high number of species with a low productivity, while communities C, D and E show predominantly P-limitation (Table 1). The peat chemical data (Table 1) reveal that the pH is slightly higher near the scarp (communities A and B) and in the open sedge communities (communities C, D and E) than in the forested communities (communities F, G and H). The carbonate content of the peat is also high near the scarp, decreasing from 19.1 (community B) to 1.0% near the river (community H). The Corg/N ratio is lowest near the scarp (13.6), varies between 20.2 and 21.0 in the open sedge vegetation and is highest close to the riverbank (26.8 in community H).
Hydrology and hydrochemistry
High surficial mire water levels were observed in May, probably as a result of snow melt during the spring. Water levels declined during the summer but remained on average above the mire surface (Table 2). Water level fluctuations observed during the summer could be related to precipitation events (Fig. 3). The MODFLOW simulations indicate that precipitation is, however, a minor component in the water balance of the mire, as the amount of net precipitation is a factor of 3 smaller than the amount of groundwater supply. MODFLOW results show that the influx of groundwater from the upland is most manifest near the terrace scarp and decreases gradually in the eastward direction (Fig. 4a). The simulations revealed two zones where net groundwater fluxes are directed downward (Fig. 4a) – i.e. in the central part of the mire at sampling locations 5 and 6 and in a zone bordering the Sargatka River (location 9). For sampling locations 3 and 4, model simulations yielded a net water flux of approximately 0 (Fig. 4a). The model results are in good agreement with the field observations, as the equipotential lines also indicate upward groundwater flow close to the scarp, infiltration at sampling locations 5 and 9 and stagnation of groundwater flow at sampling locations 3 and 4, where a local minimum in hydraulic heads is found (Fig. 4b).

Net precipitation (mm d−1) and levels of surficial mire water (cm). Water levels are averaged for all sampling locations and related to the levels measured on August 25th 2002

Analysis of the water samples revealed that spatial patterns in water composition were more distinct than temporal fluctuations. Cluster analysis resulted in the distinction of three water types (Table 3). One type comprises exclusively samples from the Ob River; the two other types comprise exclusively mire water samples. The Ob River is characterized by low concentrations of solutes and a relatively high pH. The mire water types 1 and 2 are characterized by a typical groundwater composition, with high concentrations of HCO3 and Ca. In comparison to type 1, type 2 contains higher amounts of HCO3, Cl, Na and Si. This type is found exclusively at sampling locations 3, 4 and 5 (Fig. 4c).
Discussion
Palaeo-ecology and palaeo-hydrology
Macrofossil analysis and peat stratigraphy (Fig. 2) indicate that mire development started with the deposition of silt and gyttja in a sub-neutral, carbonate-rich lake. The lake rapidly terrestrialized and changed into a wet, sedge-dominated mire. Brownmoss peats in the western part of the mire, which contain remains of Drepanocladus aduncus, indicate a dominant influence of discharging groundwater (cf. Michaelis and Joosten 2002) and constantly wet conditions (Slobodda 1979). The influence of groundwater is also demonstrated by the presence of carbonate in the peat throughout the mire. Silt layers between peat layers close to the scarp (Fig. 2) indicate periodical influxes from upland soil material, probably transported by overland flow and subsequently buried by new layers of peat. River flooding has led to the deposition of some of silt and clay in the peat in a zone bordering the river. However, the amounts of clastic material are rather small, probably due to the small sediment load of the river.
In general, plant macrofossils and peat stratigraphy compare well with the present vegetation composition, indicating long-term stable site conditions during peat formation. Peat deposits in the western part of the mire consist mainly of brownmoss peats, whereas radicel peats prevail in the eastern part of the mire. The peat underlying the forested eastern part of the study site, however, almost lacks wood peat, indicating that the establishment of a closed forest is a recent phenomenon.
Relations between hydrology, chemistry and vegetation
The chemical and vegetation characteristics of the study site can largely be explained by hydrological processes. Despite a large flux of discharging groundwater close to the terrace scarp, the steep slope of the “marginal mire zone” leads to relatively dry conditions (Table 2). This results in higher mineralization rates and more eutrophic conditions (Grootjans et al. 1985), as illustrated by the Corg/N ratios at sampling locations 0 and 1 being slightly lower than those at the other locations (Table 1). The eutrophic and relatively dry conditions give rise to the development of highly productive, species-poor vegetation dominated by Phragmites australis at sampling location 0 and to a high abundance of Betula pubescens at sampling location 1.
In the “open mire zone”, permanent inundation of the mire surface (Table 2) facilitates the presence of sedges and Menyanthes trifoliata and prevents the establishment of trees and shrubs, as the latter require (at least periodically) water levels below the surface (Moore and Bellamy 1974; Wassen et al. 1992). The high water levels can be ascribed to a large groundwater flux combined with a gentle slope of the mire surface and obstruction of surficial mire water flow by the peat ridges, which run parallel to the Ob River between locations 5 and 6. The obstruction and stagnation of water flow are illustrated by the high iron concentrations of the surficial mire water at locations 5 and 6 (Table 1) as a result of Fe2+ oxidation and the precipitation of iron compounds. Flow stagnation may also be an explanation for the presence of a distinct mire water type at locations 3–5 (Fig. 4c), as the high concentrations of HCO3, Cl, Na and Si might result from evaporation of the stagnating water. In previous research, an average concentration factor of 6.1 has been calculated for open water evaporation (Schot and Van der Wal 1992; Schot and Wassen 1993), which agrees well with the differences in Na and Cl concentrations between the mire water types 1 and 2. Water type 2 was not found at the mire surface, indicating dilution by rainwater. As a consequence of the diminishing groundwater discharge flux with increasing distance from the scarp (Fig. 4a), the influence of rainwater increases in the eastward direction, as is illustrated by the HCO3 and Ca concentrations in the surficial mire water slightly decreasing from sampling location 2 to location 5 (Table 1). This is, however, not reflected in changing vegetation characteristics.
In the “forested mire zone”, the steeper slope of the mire surface and the proximity of the Sargatka River cause draining conditions. This is indicated by the equipotential lines, which reveal a predominantly eastward groundwater flow direction in this part of the mire (Fig. 4b), and illustrated by a surficial mire water flow velocity of approximately 0.013 m d−1 in the direction of the Sargatka River (incidental measurement with coloured tracer; June 2002). The draining conditions prevent constant inundation of the mire surface at sampling locations 6, 7 and 8, illustrated by the lower levels of surficial mire water (Table 2), and enable trees and shrubs to establish in this zone. Lower PO4 concentrations in the surficial mire water of locations 7, 8 and 9 (Table 1) may indicate P-fixation by Ca- and Fe-complexes at more aerobic conditions (Boyer and Wheeler 1989). Due to aeration and root respiration, the surficial mire water becomes enriched in CO2, resulting in the formation of carbonic acid (Schot and Wassen 1993). This may cause dissolution of calcium carbonate in the underlying peat, which is illustrated by the pH and CaCO3 content of the peat being lower in the “forested mire zone” than in the “open mire zone”. The enrichment in HCO3 and Ca in the surficial mire water at sampling locations 7, 8 and 9 (Table 1) could result from the same process. Differences in peat and surficial mire water chemistry are, however, small and not reflected in vegetation characteristics.
Comparison with Biebrza mires
In the mires in the Biebrza River valley (Poland; 53°30′–53°75′N, 22°30′–23°60′E), a gradient in surficial mire water chemistry is observed from the valley edge to the river, from a groundwater type via a rainwater type to a river water type (Wassen et al. 1992). This gradient in water chemistry corresponds to a distinct vegetation zonation with respectively rich fen dominated by sedges and brownmosses, transitional fen with Sphagnum spp., Betula spp. and other species indicating acid nutrient-poor conditions and, finally, a high-productive, species-poor vegetation of tall growing sedges and grasses in a zone that is inundated by river water in spring (Wassen et al. 1992, 2002a). Contrastingly, in the major part of the Ob mire a “groundwater-like” mire water composition is observed (Fig. 4c, Table 3). The mire lacks both a fen fed by river water and a transitional fen, where the influence of rainwater is reflected in vegetation characteristics.
The absence of a distinct river-flood zone in the Ob mire, despite apparent flooding in former years, might be explained by river characteristics (infrequent and/or short lasting floods) or limited infiltration of river water during flood events due to saturation of the peat with groundwater and melt water. Furthermore, pH and nutrient concentrations do not differ greatly between the water of the Ob River and that of the mire (Table 3), so even if prolonged infiltration of river water would occur, this would not necessarily result in changes in vegetation composition. The absence of significant rainwater lenses and corresponding transitional fen vegetation in the Ob mire – despite net precipitation amounts comparable to those of the Biebrza area – can be ascribed to the larger amount of groundwater discharging into the mire. In the Biebrza mires, the groundwater flux is generally less than 0.001 m d−1 at the valley edge (Mioduszewki and Querner 2002), whereas our groundwater modelling results indicate a much larger groundwater flux to the Ob mire (Fig. 4a). This can be attributed to differences in catchment properties, such as the size of the connected groundwater recharge area, relief and the position of aquifers and aquitards. Peat stratigraphy data of the Biebrza mires, however, indicate that rainwater lenses and transitional fen vegetation have evolved only recently, as Sphagnum peat occurs merely locally and with a thickness of up to 20 cm only (Wassen and Joosten 1996). Radicel and brownmoss peats also prevail in the Biebrza mires, indicating that conditions during peat formation were similar in both areas. The recent evolvement of rainwater lenses in the Biebrza mires might be associated with anthropogenic influences (Wassen and Joosten 1996), including deepening of the riverbed, upstream dam construction, and increased groundwater extraction. An alternative explanation could be the degree of human use. The uppermost part of the peat in the Biebrza mires is more compact than the loosely structured peat in the Ob mire, possibly as a result of almost 1000 years of low intensity exploitation of the former, such as mowing and grazing (cf. De Klerk et al. submitted). More compact, less permeable peat facilitates the formation of rainwater lenses (Schot et al. 2004) and leads to stronger water level fluctuations. As long as hay making and grazing were practised in the Biebrza mires, the formation of rainwater lenses was probably prevented as tread regularly brought the surface in contact with buffering groundwater (cf. Segal 1966; Pałczyński 1985), which is close to the surface, especially in summer (Wassen and Joosten 1996). Furthermore, the regular biomass removal prevented the establishment of trees and shrubs that would result from the stronger fluctuating water levels. After the cessation of mowing (Pałczyński 1985; Okruszko 1990), the formation of rainwater lenses and massive tree and shrub encroachment took place in the naturally open Biebrza mires, possibly as a result of this compacted peat surface, even in areas where the hydrological conditions were unchanged (Joosten and Clarke 2002).
In summary, substantial differences in both vegetation characteristics and the intensity and spatial pattern of eco-hydrological processes are present between the Ob mire and the near-natural mires along the Biebrza River. Differences in the origin and ratios of water fluxes as well as the dissimilar land use history seem to be key factors for explaining these differences observed.
References
Allen SE (1989) Chemical analysis of ecological materials. Blackwell Scientific Publications, Oxford
Atlas of the USSR (1983) Atlas SSSR. Glavnoe upravlenie geodesii I kartografii pri sovete ministrov SSSR. Moscow
Blyakharchuk TA (2003) Four new pollen sections tracing the Holocene vegetational development of the southern part of the West Siberian Lowland. Holocene 13:715–731
Boden AG (1996) Bodenkundliche Kartieranleitung. Schweizerbart, Stuttgart
Boyer MLH, Wheeler BD (1989) Vegetation patterns in spring-fed calcareous fens: calcite precipitation and constraints on fertility. J Ecol 77:597–609
Dierschke H (1994) Grundlagen und Methoden der Pflanzensoziologie. Ulmer, Stuttgart
Domenico PA, Schwartz W (1998) Physical and chemical hydrogeology. Wiley, New York
Evseeva NS (2001) Geografiya Tomskoy oblasti (Geography of Tomsk Oblast). Izdatelstvo Tomskogo Universiteta, Tomsk
Frahm JP, Frey W (1992) Moosflora. Ulmer, Stuttgart
Franz HJ (1973) Physische Geographie der Sowjetunion. VEB Hermann Haack/Geographisch-Kartographische Anstalt, Gotha/Leipzig
Grootjans AP, Schipper PC, Van der Windt HJ (1985) Influence of drainage on N-mineralisation and vegetation response in wet meadows. I: Calthion palustris stands. Acta Oecol 6:403–417
Harbaugh AW, Banta ER, Hill MC, McDonald MG (2000) MODFLOW-2000, the U.S. Geological Survey modular ground-water model – user guide to modularization concepts and the ground-water flow process. U.S. Geological Survey Open-File Report 00–92, Washington D.C.
Joosten H, Clarke D (2002) Wise use of mires and peatlands – background and principles including a framework for decision-making. International Mire Conservation Group/International Peat Society, Saarijärvi
Koska I, Succow M, Clausnitzer U (2001) Vegetation als Komponente landschaftsökologischer Naturraumkennzeichnung. In: Succow M, Joosten H (eds) Landschaftsökologische Moorkunde. Schweizerbart, Stuttgart
Krasnoborov IM (2000) Opredelitel rasteniy Novosibirskoy oblasti (Plant identification guide Novosibirsk oblast). Nauka, Novosibirsk
Kulczyński S (1949) Torfowiska Polesia (Peat bogs of Polesie.) Mem Acad Polon Sci Lett-Sci Math Nat B 15:1–359
Lapshina ED (1987) Struktura I dinamika bolot poymy reki Obi (na yuge Tomskoy oblasti) [Structure and dynamics of Ob floodplain mires (in the Southern Tomsk province)] PhD dissertation, Tomsk State University, Tomsk
Michaelis D (2002) Die spät- und nacheiszeitliche Entwicklung der natürlichen Vegetation von Durchströmungsmooren in Mecklenburg-Vorpommern am Beispiel der Recknitz Dissertationes Botanicae 365. Cramer, Berlin
Michaelis D, Joosten H (2002) Peatlands in ice marginal valleys and their relation to sea level changes. Greifswalder Geograp Arbeit 27:95–99
Mioduszewski W, Querner EP (2002) Hydrological system analysis in the valley of Biebrza River. Wydawnictwo IMUZ, Falenty
Moore PD, Bellamy DJ (1974) Peatlands. Elek Science, London
Okruszko H (1990) Wetlands of the Biebrza valley. Their value and future management. Polish Academy of Sciences, Warsaw
Olde Venterink H, Wassen MJ, Verkroost AWM, De Ruiter PC (2003) Species richness-productivity patterns differ between N, P and K limited wetlands. Ecology 84:2191–2199
Oświt J (1968) Zonal arrangement of plant communities as a reproduction of water conditions in the lower Biebrza River Valley. Zeszyty Problemowe Postepow Nauk Rolniczych 83:217–232
Pałczyński A (1984) Natural differentiation of plant communities in relation to hydrological conditions of the Biebrza valley. Pol Ecol Stud 10:347–385
Pałczyński A (1985) Succession trends in plant communities of the Biebrza valley. Pol Ecol Stud 11:4–20
Ratcliffe D (1977) A nature conservation review, vol 1. Cambridge University Press, Cambridge
Rothmaler W (2002) Exkursionsflora. Band 4. Spektrum, Berlin
Rowell DL (1997) Bodenkunde – Untersuchungsmethoden und ihre Anwendungen. Springer, Berlin
Schlichting E, Blume HP, Stahr K (1995) Bodenkundliches Praktikum. Pareys Studientexte 81. Blackwell, Berlin
Schot PP, Van der Wal J (1992) Human impact on regional groundwater composition through intervention in natural flow patterns and changes in land use. J Hydrol 134:297–313
Schot PP, Wassen MJ (1993) Calcium concentrations in wetland groundwater in relation to water sources and soil conditions in the recharge area. J Hydrol 141:197–217
Schot PP, Dekker SC, Poot A (2004) The dynamic form of rainwater lenses in drained fens. J Hydrol 293:74–84
Segal S (1966) Ecological studies of peat-bog vegetation in the north-western part of the province Overijssel (The Netherlands). Wentia 15:109–141
Sjörs H (1950) On the relation between vegetation and electrolytes in North Swedish mire waters. Oikos 2:241–258
Slobodda S (1979) Die Moosvegetation ausgewählter Pflanzengesellschaften des NSG "Peenewiesen bei Gützkow" unter Berücksichtigung der ökologischen Bedingungen eines Flusstalmoor-Standortes. Feddes Repetitorium 90:481–518
Smith LC, MacDonald GM, Velichko AA, Beilman DW, Borisova OK, Frey KE, Kremenetski KV, Sheng Y (2004) Siberian peatlands: a net carbon sink and global methane source since the Early Holocene. Science 303:353–356
Stegmann H, Succow M, Zeitz J (2001) Muddearten. In: Succow M, Joosten H (eds) Landschaftsökologische Moorkunde. Schweizerbart, Stuttgart
Succow M, Joosten H (eds) (2001) Landschaftsökologische Moorkunde. Schweizerbart, Stuttgart
Succow M, Stegmann H (2001) Nährstoffökologisch-chemische Kennzeichnung. In: Succow M, Joosten H (eds) Landschaftsökologische Moorkunde. Schweizerbart, Stuttgart
Tanneberger F, Hahne W, Joosten H (2003) Wohin auch das Auge blicket: Moore, Moorforschung und Moorschutz in Westsibirien. Telma 33:209–229
Walter H, Breckle SW (1994) Ökologie der Erde Band 3: Spezielle Ökologie der gemässigten und arktischen Zonen Euro-Nordasiens. Gustav Fischer, Stuttgart
Wassen MJ, Joosten JHJ (1996) In search of a hydrological explanation for vegetation changes along a fen gradient in the Biebrza Upper Basin (Poland). Plant Ecol 124:191–209
Wassen MJ, Barendregt A, Pałczyński A, De Smidt AJT, De Mars H (1992) Hydro-ecological analysis of the Biebrza mire (Poland). Wetlands Ecol Manage 2:119–134
Wassen MJ, Bleuten W, Bootsma MC (2002a) Linking hydrology to ecology; Biebrza as a geographical reference. Ann Warsaw Agric Univ 33:27–47
Wassen MJ, Peeters WHM, Olde Venterink H (2002b) Patterns in vegetation, hydrology and nutrient availability in an undisturbed river floodplain in Poland. Plant Ecol 165:27–43
Wilmanns O (1998) Ökologische Pflanzensoziologie. Quelle and Meyer, Wiesbaden
Yefremov SP, Yefremova TT (2001) Stocks and forms of deposited carbon and nitrogen in bog ecosystems of West Siberia. In: Vasiliev SV, Titlyanova AA, Velichko AA (eds) West Siberian Peatlands and carbon cycle: past and present. OOO Agentstvo Sibprint, Novosibirsk
Acknowledgements
We thank the scientists and students of Tomsk State University and the Institute of Soil Science and Agrochemistry, RAS, Novosibirsk, who helped us in the field and in the laboratory in Siberia, as well as the staff from Utrecht and Greifswald University for their help with the laboratory work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Schipper, A.M., Zeefat, R., Tanneberger, F. et al. Vegetation characteristics and eco-hydrological processes in a pristine mire in the Ob River valley (Western Siberia). Plant Ecol 193, 131–145 (2007). https://doi.org/10.1007/s11258-006-9253-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-006-9253-x