
Abstract
Pea (Pisum sativum L.) is a valuable grain legume in organic crop rotations. Pea rotations provide nitrogen (N) to the system through N fixation and produce animal feed or human food. Because of the high susceptibility of pea to pests, diseases and weeds and due to low profitability, especially in organic systems, pea cropping intensity decreased in the last 15 years in Austria. Therefore, it is important to find strategies for improving pea cropping systems in organic systems, by increasing yields and providing a positive N balance. The objective of this study was to compare pea genotypes of selected field and fodder pea in pure and mixed pea stands for biomass performance, biological N fixation and pre-crop effect under dry site conditions in a 2-year study in Eastern Austria. Pea N fixation was estimated using the extended N-difference method, with oat as the reference crop. The highest grain yield was found for the leafed field pea, with 2.5 Mg dry mass (DM) ha−1, followed by the semi-leafless field pea with 2.1 Mg DM ha−1 and the pea-mixtures with 2.2 Mg DM ha−1. The field pea cultivars yielded more than the fodder pea cultivars with 1.6 Mg DM ha−1. The average N concentration in pea grains was 3.6 mg g−1. The fodder pea type contained 3.8 mg g−1, significantly more N (p < 0.0001) than the semi-leafless and leafed field pea. Pea N fixation ranged from 53 to 75 kg N ha−1, corresponding to 42–50% of N derived from the atmosphere (% Ndfa). No differences in N fixation were found among cultivars, types and field/fodder pea. The fodder pea exported less N from the field because of their lower grain yield. Therefore, the N balance (N-input − N-output) of fodder pea was positive, with + 3.4 kg N ha−1 compared to the negative N balance of − 0.6 to − 3.6 kg N ha−1 for the leafed field pea types. These differences were not reflected in the following winter wheat crop, where the DM grain yield was 3.6–3.9 Mg ha−1 with no differences between cultivars and ideotypes. The results demonstrate that leafed field pea could have a sufficient grain yield and fodder pea could produce high N concentration in the grains. Because there are no differences regarding the effect of pea types on the yield of the following crop, it can be concluded that all tested pea types are suitable for successful organic pea production under dry site conditions. While there were no negative effects on the subsequent crop, the different ideotypes and mixtures may be selected based on different management goals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The pea (Pisum sativum L.) is a valuable grain legume in organic farming. Pea rotations provide N to the organic system for the succeeding non-leguminous crops via symbiosis with N fixing bacteria, and produce grain that is rich in protein, which is particularly important for organic pig and poultry feed. Generally, the cultivation of pea in organic farming is difficult due to its low tolerance against pests, diseases and weeds and its low profitability, directly attributable to pea perceived unstable yield of pea crops (Moot and McNeil 1995). Reckling et al. (2018) found that the yield instability of grain legumes was comparable to that of other spring-sown crops. In Austria, organic pea cropping decreased from 8824 ha in 2005 to 2581 ha in 2016 (BMLFW 2017); therefore, it is essential to find strategies to achieve high and stable organic pea yields.
Pea genotype has a major influence on the yield and development of the plant. Semi-leafless pea types with leaflets transformed into tendrils and conventional stipules allow additional light and air movement throughout the crop, thus creating less favourable conditions for fungal diseases (Grevsen 2003). In comparison, leafed pea types have normal leaflet and stipule sizes, a higher shading ability and usually exhibit the highest yield potential (Baigorri et al. 1999). Therefore, the postulated competitive ability of leafed pea types against weeds could be higher, but their standing is lower compared with semi-leafless cultivars because the plant has no tendrils (Grevsen 2003). The fodder pea (Pisum sativum L. arvense) may be ideally suited for weed management in organic farming, because of its bigger leaves and therefore higher competitive ability against weeds. However, the nutritive value for animals is higher in field pea (Pisum sativum L. sativum). Grain legumes such as pea generally have a weak competitive ability against weeds. Several studies have demonstrated a reduced weed biomass in crop mixtures combining a cereal and a grain legume, compared to the legume crop alone (Courre-Hellou et al. 2011; Hauggaard-Nielsen et al. 2001). The factors that promote the greater competitive ability of intercrops compared to legume monocultures are not well understood. Crop mixtures with different pea types provide the possibility to combining the advantages of the cultivars, e.g. mixtures of leafed pea types with a higher shading ability together with semi-leafless pea types with tendrils that provide structural stability, and may improve weed suppression and yield compared with monocultures of the same cultivars. The yield of mixtures with different pea leaf type cultivars has been shown to increase compared to single cultivars (Schouls and Langelaan 1994), where the semi-leafless pea improved the lodging resistance of the leafed pea. Syrovy et al. (2015) found an optimal combination of 25% leafed and 75% semi-leafless pea types to improve weed suppression by 19% compared with the less competitive leafed cultivar and a higher total seed yield than single-cultivar crops. Syrovy et al. (2015) stated that combinations of leafed and semi-leafless pea are better able to compete with weeds and yield potential was higher than crops with only one cultivar, but the mechanism is currently not well understood.
The contribution of grain legumes to the N supply of the organic crop rotation is generally reduced through the removal of the grain (Haynes et al. 1993), represented by a high NHI (=N harvest index; Carranca et al. 1999). In order to evaluate the N benefit of the legume to organic cropping systems, a simplified N balance of the crop can be estimated. Specifically, the net output of the N removed in the grain is subtracted from the N input of biologically fixed N in the grain legume to obtain N balance (Peoples et al. 1995). As long as the NHI of a legume is less than the Ndfa, a positive N balance can be archieved (Haynes et al. 1993). For instance, Kumar and Goh (2000) reported an unusually high positive N balance (171 kg N ha−1) for pea due to low NHI values (28%) and high Ndfa (N derived from atmosphere) values (70%), including root N in the estimations. The N benefit can be decreased when the major part of the N from vegetative plant parts is remobilized for the seed production (Corre-Hellou and Crozat 2005). Other studies have shown negative N balances because of high NHI values in the grain legume (Corre-Hellou and Crozat 2005, Kelstrup et al. 1996; Armstrong et al. 1997) and not considering root-N and N rhizodeposition. In pea, up to 12% of the plant N may be present in the roots (Armstrong et al. 1994). Consequently, the fixed N in the pea roots and rhizodeposition should be included to the estimation of the N balance (Jensen 1996). Generally, N balance of pea has a wide range, for example − 18 to + 73 kg N ha−1 (Aveline et al. 1998) or − 25 to + 1 kg N ha−1 (Reiter et al. 2002). Legume breeders usually attempt to increase the NHI and protein supply in the grain, reducing the contribution of grain legumes to the soil N (Senaratne and Hardarson 1988). Therefore, maximising both the biological N fixation and optimising the grain production of the pea is an important issue for organic farming. In areas with moist and cool climates, or with irrigation, pea can fix N up to 286 kg N ha−1 (Kumar and Goh 2000; inclusion of root-N). In regions with high temperature and less precipitation, values ranged from 37 (Carranca et al. 1999) to 136 kg N ha−1 (McCauley et al. 2012).
To increase the value of the grain legume in the crop rotation, it is important to identify pea cultivars and mixtures with the desired performance in organic systems. Additionally, information about N fixation capacity of pea cultivars is necessary for breeders and farmers to promote organic farming. Pea mixtures with different seeding proportions of field/fodder pea and foliage types (semi-leafless/leafed) could be a method to optimize organic pea cultivation. The objective of our study was to test the effect of (1) genotypes representing cultivars of selected field/fodder pea and (2) semi-leafless/leafed-types in pure stands and mixtures in two field trials on biomass performance, biological N fixation, N balance and pre-crop effect.
Materials and methods
Site and soil
The experiment was carried out in 2002–2004 in Raasdorf, on the organically cultivated fields of the University of Natural Resources and Life Sciences, Vienna. Raasdorf is located in the Marchfeld, an area of about 100,000 hectares, approx. 5 km east of Vienna (48°14′N, 16°35′E) at an altitude of 150–160 m above sea level. The mean annual air temperature is 10.2 °C and the average precipitation is 520 mm year−1. Air temperature and rainfall during the experimental period are shown in Tab. SM1. Soils are Calcaric Phaeozems (WRB 1998) from loess with a silty loam texture, organic carbon contents of 22 g kg−1, Pcal (calcium acetate lactate extract) of 83 and Kcal of 201 mg kg−1 in the Ap horizon, and a pHCaCl2 value of 7.6.
Experimental set-up and management practice
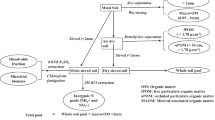
The experiment consisted of two trials, starting in spring 2002 and 2003, respectively. During both years, eight different pea cultivars in pure stands, four pea mixtures and the reference oat crop (Avena sativa L.; seeding density 120 kg ha−1) were sown in March after the pre-crop winter wheat in four replications in a randomized complete block design with plots measuring 6 × 7 m (Tab. SM2). The last legume grown on the experimental site was Lucerne (Medicago sativa) in 1998. The seeding density of pea and oat was 120 and 350 plants m−2 respectively. The number of seedlings in four rows of 1 m length was counted 2–3 weeks after seedling emergence. The actual plant densities were 86–115 plants m−2 in pea monocrop variants and 77–119 plants m−2 in pea mixtures. Each experimental plot was subdivided into four harvest areas (0.5 × 2 m) which were used for retrieving plant samples at harvest. The crops did not receive any fertilizer application; the experimental site was managed according to organic guidelines with no use of herbicides and with weeding by hand 1–2 times during the emergence stage of the pea. The hand weeding was carried out in order to support a successful development of the young pea plants, but could be a limitation in the present study when compared the results with cultivation in practice. During the year, the weed cover in the experimental plots was evaluated several times (from emergence stage to pea harvest; see Table 5) by estimation of weed pressure per area (in % m−2) and at the flowering stage by determining the leaf area index (LAI). After pea harvest in July, the pre-crop effect on the subsequent winter wheat crop was investigated by measuring grain yield and crude protein content. Winter wheat was sown in October (Triticium aestivum; cv. Capo; seeding density 160 kg ha−1) and harvested in July of the following year.
Sampling and analytical methods
Harvest was carried out when the pea crop reached maturity. All plants of four harvest areas per plot (0.5 × 2 m) were collected by hand. Grain and shoot dry matter (DM) yield were determined for pea and oat plants by drying an aliquot at 105 °C to constant weight. In the second year, the root biomass of the plants in two layers (0–30 cm, 30–60 cm) was taken at harvest by using a root auger (10 cm in diameter, 60 cm deep, with four replicates per plot). The roots were subsequently separated from the soil by a hydro pneumatic elutriation system (Gillison’s Variety Fabrication Inc., USA) through a sieve with a mesh of 760 µm. Aliquots of the grain, shoot and root material were dried at 60 °C for 48 h, ground up to a fine powder, and analysed for plant N content with an LECO CN-2000 analyser (Vario Macro Cube, Elementar Analysensysteme GmbH). The N content of wheat was converted to protein content with a factor of 5.7 (Tkachuk 1966).
Soil samples for determining inorganic N in three layers (0–30, 30–60 and 60–90 cm) were taken in July immediately after the pea harvest and in March in the subsequent winter wheat trial. Soil inorganic N (Ninorganic) was extracted from the soil using a 0.0125 M calcium chloride (CaCl2) solution at a 1:4 ratio. Extract NO3-N and NH4-N were measured calorimetrically according to Schlichting et al. (1995) using an UV–VIS-Photometer (measurement of NH4-N at 660 nm wavelength with NaOH, Sodium salicylate, Nitroprusside sodium dehydrate, Dichlorisocyanuric acid as reagents; measurement of NO3-N at 540 nm wavelength with Vanadium(III)-chloride, N-1-Naphthylethylendiamine dihydrochloride, sulfanilic acid, hydrochloric acid 32% as reagents).
Calculations and statistics
The N-difference method (ND) as described by Evans and Taylor (1987) was used for estimating N derived from the atmospheric N2 of the pea variants. In this method, the difference between total plant N and post-harvest soil mineral N of an N fixing legume and a non-fixing reference crop is considered to be N that has been obtained through biological fixation. N fixation (Nfixe = essential) is therefore:
where Ngrain+shoot Leg = N yield grain and shoot of pea; Ngrain+shoot Ref = N yield grain and shoot of oat; Nsoil Leg = Post-harvest soil N in pea plots; Nsoil Ref = Post-harvest soil N in oat plots.
In the second year, the N fixation was estimated with an extended N-difference method attributed to Hauser (first extension, 1992), including consideration of the roots in order to improve the method. N fixation (Nfixr = N fixation total including roots) is therefore:
where Nroot Leg = N yield root of pea; Nroot Ref = N yield root of oat.
The N-fixing efficiency of the pea was estimated by calculating the Ndfa value (Ndfa = N derived from atmosphere), estimating the proportion of plant N gained from the atmosphere:
where Ngrain Leg = N yield grain of pea, Nshoot Leg = N yield shoot of pea.
In the second year, the estimation of Ndfar (= include the roots) is therefore:
where Nroot Leg = N yield root of pea.
To estimate the N benefit of pea for the subsequent crop in the rotation, which is winter wheat, a simplified N balance (Nbal) calculation was used (Schmidtke 1997):
where N-Input = Nfix plus the estimated rhizodeposition of pea (according to Fustec et al. 2010: approx. 11% of total plant N); N-Output = Ngrain, removed from the field.
By application of this calculation, we assume that there is insufficient time and moisture for mineralization and nitrification of the rhizodeposits.
In legumes, total N is partitioned into harvested grain-N and crop residue-N (straw and roots). The proportion of N partitioned into grain is described as N harvest index (NHI) regarding to Myers and Wood (1987):
The dry matter (DM) grain yield of pea mixtures mixed in different seeding ratios were compared to the expected values, based on pure stands following Kübler et al. (2006):
The mixing effect 1 of the DM grain yield is described regarding Kübler et al. (2006):
Data was analysed using the PROC MIXED procedure of SAS (SAS Institute, Inc. Version 9.4) with row and block of experimental trials as random effects and year, type (semi-leafless field pea type, leaf field pea type, fodder pea type, mixtures type) and cultivar (variety) as fixed effects, as well the interaction of fixed effects. Additionally the effect of seeding density (pure stands vs. mixtures) as a fixed effect was tested, also simple linear regressions on NHI versus N balance and Ndfa versus N balance and a multiple linear regression on N balance versus Ndfa between the pea types were performed. Treatment effects were considered significant at p < 0.05. Results are presented as least-square means (lsmeans) and standard error of the means (SEM). Tukey–Kramer test was used for pairwise comparison of the means and significant differences were marked with lower case letters.
Results
Climatic conditions and soil inorganic N
Rainfall through March and April was higher in spring 2002 and lower in spring 2003 than the long-term precipitation mean (Tab. SM1). The mean air temperature was similar to the long-term averages. At pea harvest, there were no significant differences among treatments in soil inorganic N in both July 2002 with 42–66 kg ha−1 and July 2003 with 20–29 kg ha−1 (Fig. 3). However, there was a large difference between the 2 years. In March 2003, the soil under the subsequent crop winter wheat crop contained 137–174 kg ha−1 in the 0–90 cm soil depth at start of wheat tillering, and there were no significant differences between wheat with different pre-crops (data not shown; p = 0.91). In March 2004, there was a significant difference (p = 0.021) in inorganic N under winter wheat with 145–148 kg ha−1 after the pea cultivars Rhea, Sirius, Bohatyr and pea cv. Herold with 88 kg ha−1 (SE 11.6; data not shown).
Grain yield and N content
The grain DM yield of 2.5 Mg ha−1 (SEM 0.11) for leafed field pea was significantly higher than the 2.1 Mg ha−1 (SEM 0.09) for semi-leafless field pea and 2.2 Mg ha−1 (SEM 0.08) in pea-mixtures (Table 1). There were significantly greater grain yields of field pea cultivars when compared to fodder pea cultivars with 1.6 Mg ha−1 (SEM 0.09; Table 1). Both the effect of pea type and the pea cultivar were significant when comparing grain DM yield (Tab. SM3, Fig. 1).

Grain, straw and root DM (dry matter) yield of different pea cultivars/mixtures (mean value over 2 years). PS: Pisum sativum ssp. sativum, field pea; semi-leafless type. PL: Pisum sativum ssp. sativum, field pea; leafed type, PA: Pisum sativum subsp. arvense, fodder pea. MSA: mixture of field and fodder peas, MSL: mixture of semi-leafless and leafy field pea cultivars; Error bars: SEM, standard error of mean; mean values with same letter do not differ significantly (p < 0.05, Tukey–Kramer)
Grain yield of the pure stand leafed field pea cv. Erbi (2.8 Mg ha−1; SEM 0.15) and Bohatyr (2.2 Mg ha−1; SEM 0.15), the semi-leafless cv. Gotik (2.4 Mg ha−1; SEM 0.15), and mixtures with Erbi–Gotik and Gotik–Erbi significantly exceeded the other pure stand cultivars and mixtures (Fig. 1). Grain DM yield (Tab. SM5), grain N content and N yield were not influenced by seeding proportion, when comparing pure stands with mixtures. The EW values were significantly different between the pea mixtures (Tab. SM6), but there was no significant difference regarding the ME1. The negative ME1 values indicated that the mixtures developed less grain yield regarding the expected values.
The average N concentration in pea grains was 3.65 mg g−1. Fodder pea contained significantly more N than the semi-leafless and leaf field pea (Table 1). There were also significant differences in grain N content among the cv. Rhea, Dora, Erbi, Herold, Herold/Rhea, Sponsor/Rhea, Erbi/Gotik and the other varieties and mixtures (Fig. 2).

N (mg g−1) in grain, straw and roots; NHI (nitrogen harvest index; %, bars) of different pea cultivars/mixtures (mean value over 2 years). PS: Pisum sativum ssp. sativum, field pea; semi-leafless type, PL: Pisum sativum ssp. sativum, field pea; leafed type, PA: Pisum sativum subsp. arvense, fodder pea, MSA: mixture of field and fodder peas, MSL: mixture of semi-leafless and leafy field pea cultivars; Error bars: SEM, standard error of mean; mean values with same letter do not differ significantly (p < 0.05, Tukey–Kramer)
The higher pea grain yield was significant for the leafed field pea (93 kg N ha−1; SEM 4) and the mixtures (81 kg N ha−1; SEM 3), compared to the semi-leafless (76 kg N ha−1; SEM 3) and the fodder pea types (63 kg N ha−1; SEM 3; Table 1). The grain N yield of cv. Erbi (106 kg N ha−1; SEM 6) was significantly greater than that of the cv. Herold (72 kg N ha−1; SEM 6), Sponsor (72 kg N ha−1; SEM 6), Bohatyr (80 kg N ha−1; SEM 6), Dora (50 k g N ha−1; SEM 6) and Rhea (71 kg N ha−1; SEM 6) and also of the mixture Herold/Rhea (69 kg N ha−1; SEM 6) and Sponsor/Rhea (69 kg N ha−1; SEM 6; data not shown). The variety Gotik (82 kg N ha−1; SEM 6) and mixtures with cv. Erbi and Gotik (98 kg N ha−1 Erbi–Gotik, SEM 6; 88 kg N ha−1 Gotik–Erbi, SEM 6) accumulated similar amounts of grain N yield as cv. Erbi.
Dry matter production and N accumulation in crops
No differences were found in straw DM yield among pea leaf-types, fodder and field pea, pure stands and mixtures. In contrast, the pea cultivar had a significant effect on the straw yield (Tab. SM3, Fig. 1), where fodder pea cv. Dora had higher straw biomass (2.9 Mg ha−1; SEM 0.1) than cv. Sirius (2.2 Mg ha−1; SEM 0.1). The N content in pea straw was 1.8–2.1 mg g−1 with no significant differences between cultivars and types (Fig. 2, Table 1).
The straw DM yield, straw N content and N yield were also not influenced by seeding density (Tab. SM5). Pea variants developed similar amounts of root DM, with an average of 0.46 Mg ha−1 for fodder pea types (SEM 0.04) to 0.54 Mg ha−1 (SEM 0.04) for leafed field pea types (Table 1, Fig. 1), and no significantly differences between types and varieties. There were also no significant differences between N content in roots of pea variants, with an average of 1.75 mg g−1 N (Fig. 2, Table 1).
The total crop biomass (grain and straw DM yield) of leafed (cv. Erbi 5.4 Mg ha−1) and semi-leafless (cv. Gotik 5.2 Mg ha−1) pure crops showed no significant differences between the ideotype mixtures (Mix. Erbi–Gotik 5.1 Mg ha−1; Mix. Gotik–Erbi 4.9 Mg ha−1; Fig. 1). In our two consecutive experimental years, the weed pressure in fodder pea plots was significantly lower (13% weed cover) than in field pea plots (18–19%), where the leafed types cv. Bohatyr and Erbi had the highest weed cover (Table 5). The weed pressure at the pea flowering and pod stage was significantly higher in mixtures than in pure stands (Tab. SM5, Table 3); therefore, the LAI at flowering stage was lower in mixtures.
Biological N fixation
The amount of N fixed (mean of 2 years, Nfixe) in pea variants was 53–75 kg N ha−1, respectively, corresponding to 42–50% of N being derived from the atmosphere (Ndfae). No differences were found between different cultivars and types (Tab. SM4).
In the present field study, leafed types (cv. Bohatyr, Erbi) fixed 100 kg N ha−1 on average in 2002 and 49 kg N ha−1 on average in 2003, compared to the semi-leafless types (cv. Gotik, Herold, Sponsor) with 78 kg N ha−1 in 2002 and 37 kg N ha−1 in 2003. The N fixation of field peas was 87 and 42 kg N ha−1 in 2002 and 2003. For fodder peas, average N fixation was 64 and 42 kg N ha−1 in 2002 and 2003 in the two consecutive years. The amount of N fixed in the second year only, calculated with consideration of above- and belowground root N yield (Nfixr), ranged from 20 to 53 kg N ha−1, with no significant differences between pea types and cultivars (data not shown). When considering N fixation of aboveground N only (Nfixe), N fixation ranged from 24 to 57 kg N ha−1. Similarly, aboveground Ndfae was 34%, whereas root Ndfar was 29%. Therefore, the consideration of roots could not improve the estimation of N fixation in our experiment.
On average, N fixation during the first year was 85 kg N ha−1 and 55% Ndfa, and 39 kg N ha−1 and 34% Ndfa during the second year. The reason for these differences was the higher grain N yield of pea in 2002 with an average of 86 kg N ha−1 compared to 68 kg N ha−1 in 2003 and the dryer growth conditions in spring. The lower N fixation in 2003 was visible through the differences in soil inorganic N at pea harvest in both years. In July 2002, average N concentration for pea plots was 56 kg Ninorganic ha−1 was in the 0–90 cm depth, and in July 2003 was reduced to 24 kg Ninorganic ha−1 (Fig. 3).

Soil inorganic N (kg ha−1) in 0–0.9 m of different pea cultivars/mixtures at harvest in July 2002 and 2003. PS: Pisum sativum ssp. sativum, field pea; semi-leafless type, PL: Pisum sativum ssp. sativum, field pea; leafed type, PA: Pisum sativum subsp. arvense, fodder pea, MSA: mixture of field and fodder peas, MSL: mixture of semi-leafless and leafy field pea cultivars; Error bars: SEM, standard error of mean; mean values with same letter do not differ significantly (p < 0.05, Tukey–Kramer)
Contribution to N balance and effect on following crop
The average contribution of the pea crops to the soil N balance was + 15 kg N ha−1 in 2002 and − 17 kg N ha−1 in 2003, although only grain was exported and straw stayed on the field. There was a great variation in the N balance, with a range from + 124 to − 51 kg N ha−1.
In the present field study, NHI of pea leafed types (0.61–0.65; SEM 0.02) and of mixtures (0.61; SEM 0.02) were significantly different from the fodder pea ideotypes with NHI of 0.53 (SEM 0.02; Tab. SM4, Table 2). The NHI of the peas was similar in the years, with 0.59 in 2002, and 0.61 in 2003. The fodder pea cultivars Dora and Rhea, as well as the mixture Sponsor–Rhea, had a significantly lower NHI than the other cultivars/mixtures (Fig. 2).
The N grain yield of the fodder pea types was lower than that of the field pea types (Table 1). Therefore, less N was exported from field and N balance for the fodder pea types was positive with + 3.4 kg N ha−1 compared to the negative N balance of − 0.6 to − 3.6 kg N ha−1 for the leafed field pea types (Table 2). The N balance decreased when N harvest index increased (2002: r2 = 0.51; 0.0 sign., df 1). NHI needs to be below 0.66 (± 24.9 SE) for the N balance to be positive (Fig. 4a; 2003: r2 = 0.40, data not shown). The N balance was positively correlated with the amount of N fixed (2002: r2 = 0.60, 2003: r2 = 0.50, data not shown) and with % Ndfa (2002: r2 = 0.54, data not shown; 2003: r2 = 0.58; 0.0 sign., df 1). Ndfa needs to be above 54.4 (± SE) for the N balance to be positive (Fig. 4b). There were no differences between pea types when comparing the relationship between N balance and Ndfa (p = 0.90).

a Relationships between NHI (nitrogen harvest index; %) and N balance (N-Input − N-Output) for 2002; at NHI = 66% the N balance is zero. b Relationships between Ndfa (Nitrogen derived from atmosphere) and N balance for 2003; at Ndfa = 54.4% the N balance is zero
The differences regarding N balance between the pea ideotypes were not reflected in the following winter wheat crop. The DM grain yield was 3.6–3.9 t ha−1 with no differences between cultivars, ideotypes and seeding proportion (Tab. SM4, Table 2, Tab. SM5). The tendency for higher N fixation of the leafed types compared to the semi-leafless types in both year periods led to higher wheat yields in the following winter wheat (2002/2003: 3.8/3.9 t ha−1 after leafed types compared to 3.5/3.6 t ha−1 after semi-leafless types), but the differences were not significant. The crude protein content was 10.2–10.6 mg g−1, with no differences between cultivars/ideotypes (Tab. SM4, Table 2).
Discussion
Pea biomass performance and weed pressure
The semi-leafless types had a better standing ability than the leafed field pea types; nevertheless the leafed cv. Erbi was the highest yielding cultivar in 2002 (5.4 t ha−1 total crop biomass yield). Pea crops with an early lodging during their development phase are able to raise themselves up again from the upper nodes and produce high grain yields (Aufhammer 1998). Elers (2001) described the leafed type cv. Grana with a low standing ability, nevertheless it had the highest grain DM yield when compared to the other tested semi-leafless pea cultivars. The higher leaf area index of he leafed pea types could reduce weed pressure and therefore support the development of the pea yield.
In our experiment, the weed pressure at emergence, flowering and pod stage of pea mixtures differed from the monocultures (Table 3). At emergence, the weed pressure in fodder pea plots was significantly lower than in field pea plots (Table 4); the reason for this is the higher LAI (leaf area index) of the fodder pea cultivars compared to the field pea cultivars (Table 5), which means the increased leaf area was better able to shade the soil, therefore decreasing overall weed pressure. Regarding weed pressure, our results are in contrast to Syrovy et al. (2015), where an optimal combination of 25% leafed and 75% semi-leafless pea improved weed suppression by 19% compared with the less competitive leafed cultivar. In Syrovy et al. (2015), the semi-leafless monoculture and all mixtures increased total crop biomass when compared with the leafed monoculture. The authors suggested these differences were a result of less complete overlap of resource requirements between different species and an increasing level of lodging resistance as the percentage of semi-leafless pea increased. The design of our present study does not allow the separation of response of each cultivar to monoculture density compared with mixture, because each cultivar was not grown in monoculture of different densities, as suggested by Jolliffe et al. (1984). Nevertheless, our tested pea mixtures were not more successful regarding grain yield compared to the pure stands. Therefore, there was no need for further lodging resistance through our mixtures with semi-leafless types under the trial site conditions.
Biological N fixation, N balance and pre-crop effect
Pea fix approximately 50–300 kg N ha−1 with an average of 100 kg N ha−1 (Freyer et al. 2005), dependent on environmental conditions and agricultural management, and use almost the entire amount of fixed N in grain production. At least part of the cause for the low N credit from grain legumes is the fact that these legumes have been bred for yield, but not for N fixation. In an experimental study in France (Cousin 1997), the N fixation period of different pea genotypes was equivalent (beginning 1 month after sowing and lasting up to 2 months), but large differences in cumulative N fixation rates between the pea varieties were observed. Armstrong et al. (1994) studied six contrasting pea genotypes in Australia and found considerable variations in Ndfa ranging from 60 to 91 Ndfa % between genotypes and sites due to differences in the proportional dependence on fixation. Siegmund (1994) studied 18 different pea leaf types in a pot experiment and found an increase for N fixation dependent on the leaf area (leafed types > semi-leafless types > leafless types). There seems to be a connection between leaf type and photosynthetic efficiency, which is important for the energy consuming N fixation process. Also, it is well known that N fixation is strongly influenced by soil water content; the symbiotic process is more sensitive to water stress than biomass production and N assimilation for a number of legume crops (Serraj et al. 1999).
Yearly fluctuations in weather patterns influence legume growth and fixation activity. The higher soil inorganic N content during the harvest 2002 showed that environmental conditions were more favourable for the symbiotic N fixation process in this year, as high plant-available soil N resources had not been taken up by the plants. The rainfall in spring 2002 compared to spring 2003 is indicative of these conditions, as demonstrated by the higher precipitation rates in March and April of 2002 (see Tab. SM1). It has been reported that N fixation is very sensitive to moisture stress with drought severely inhibiting nitrogenase activity and nodulation (Ledgard and Steele 1992). Also, it has been reported that nodule biomass of established pea nodules declined under water stress (Frechilla et al. 2000; Mahieu et al. 2009). Consequently, we link yearly variation in nitrogen fixation to precipitation and soil moisture in spring.
Generally, through harvest residues (straw, roots) and rhizodeposition, grain legumes usually provide a positive N balance in the soil (Cuttle et al. 2003). When the N fixation is limited by environmental conditions, such as moisture deficits and subsequent drought stress in pea, N benefits can be very negative (Jensen 1996). Carranca et al. (1999) found, that the cultivation of pea has little benefit for soil N, removing on average 22 kg N ha−1. A small average positive contribution to the soil N balance, less 10 kg N ha−1, was reported by Corre-Hellou and Crozat (2005). Jensen (1996) reported a large variation from − 19 to + 45 kg N ha−1 for the N benefit, including the fixed N in the roots. Evans et al. (1989) reported ranges from − 32 to 96 kg N ha−1 for pea, dependent on biomass accumulation and Ndfa of 65% or greater. The average contribution of our tested pea crops for the N benefit was − 17 to + 15 kg N ha−1. However, the simplified N balance calculation in our study, where we considered the N content of the roots at harvest plus the estimated rhizodeposition of pea roots, underestimates plant contribution to N balance because the turnover of fine roots during the vegetation period and the nodules are not accounted for. Previous studies have shown harvest index of pea genotypes varying from 0.30 to 0.60 (Martin et al. 1994; Uzun and Acikgöz 1998). Legumes with a high NHI can take a high amount of N from the field at harvest. Combined with a reduced N fixation, the cultivation of the legume can therefore lead to a negative N balance. If the conditions for seed-filling are optimal, a major part of the N from vegetative plant parts is remobilized for seeds and the NHI will be high (Jensen 1996). In order to optimize the contribution of N fixing legumes, and therefore maximize the N supply in the crop rotation, among others, it is important to grow legume types which leave more N in the harvest residues on the field, e.g. by a lower N harvest index or higher N rhizodeposition (Schmidtke 2013). Also, the maximisation of the yield is important, because with increasing yield the symbiotic N fixation activity of the legume increases also (Köpke 1996).
Corre-Hellou and Crozat (2005; r2 = 0.67) and Wichmann et al. (2006; r2 = 0.52 or 0.56) found a similar relationship of NHI and N balance. The N balance was positively correlated with the amount of N fixed (2002: r2 = 0.60, 2003: r2 = 0.50, data not shown) and with % Ndfa (2002: 0.54, data not shown; 2003: 0.58; Fig. 4). A high positive contribution to the soil N balance was recorded with combined high % Ndfa and a low N harvest index. On the other hand, breeders usually attempt to increase the N harvest index and the protein supply in the grain, therefore reducing the contribution of the legume to the soil N.
Conclusion
Pea genotypes influence the performance of the crop and the effect on the following crop in the rotation. Leafed pea types developed the highest yield and N fixation potential, caused by a better photosynthetic efficiency. For organic farms in dry regions the leafed pea types should be favored because of their higher shading ability, and therefore better competitiveness against weeds, as well as lower soil evaporation compared to leafless types. Nevertheless, these pea type differences were not reflected in the following winter wheat. Fodder pea had a high competitive availability against weeds and developed a higher straw yield (including the leaves). Although the grain yield of the fodder pea was lower than that of the field pea, the amount of N fixation was similar; the N balance was more positive for fodder pea but not statistically significant, because of their low N-harvest index. Generally, crop mixtures with different pea types combine the advantages of the used cultivars. The pea mixtures in our experiment showed no advantages compared to the pea monocultures with respect to weed pressure and grain yield.
Because there are no differences regarding the effect of pea types on the yield of the following crop, it can be concluded that all tested pea types are suitable for successful organic pea production under dry site conditions. In order to improve the value of pea in organic crop rotations in semi-arid regions, consequently, additional information about the performance of pea cultivars and types is necessary (e.g. crop water stress index, new cultivars). Due to peas perceived unstable yield, research on pea pests, diseases and weeds should be conducted in the future.
References
Armstrong EL, Pate JS, Unkovich MJ (1994) N balance of field pea crops in South Western Australia, studied using the 15N natural abundance technique. Aust J Plant Physiol 21:533–549
Armstrong EL, Heenan DP, Pate JS, Unkovich MJ (1997) N benefits of lupin, field pea, and chickpea to wheat production in south-eastern Australia. Aust J Agric Res 48:39–47
Aufhammer W (1998) Getreide- und andere Körnerfruchtarten. Verlag Eugen Ulmer, Stuttgart
Aveline A, Domenach AM, Carrouée B, Crozat Y (1998) Compared effects of pea and wheat cultivation on the soil N balance: results of the follow-up of a long term trial. In: Proceedings of the 3rd European conference on grain legumes, Valladolid, pp 134–135
Baigorri H, Antolin MC, Sánchez-Diaz M (1999) Reproductive response of two morphologically different pea culitvars to drought. Eur J Agron 10:119–128
Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft - BMLFUW (2017). Grüner Bericht 2017. http://www.gruenerbericht.at/. Accessed 18 Sept 2017
Carranca C, de Varennes A, Rolston D (1999) Biological N fixation by fababean, pea and chickpea, under field conditions, estimated by the 15N isotope dilution technique. Eur J Agron 10:49–56
Corre-Hellou G, Crozat Y (2005) N2 fixation and N supply in organic pea (Pisum sativum L.) cropping systems as affected by weeds and peaweevil (Sitona lineatus L.). Eur J Agron 22:449–458
Courre-Hellou G, Dibet A, Hauggaard-Nielsen H, Crozat Y, Gooding M, Ambus P, Dahlmann C, von Fragstein P, Pristeri A, Monti M, Jensen ES (2011) The competitive ability of pea-barley intercrops against weeds and the interactions with crop productivity and soil N ability. Field Crops Res 11:264–272
Cousin R (1997) Peas (Pisum sativum L.). Field Crops Res 53:111–130
Cuttle S, Shepherd M, Goodlass G (2003) A review of leguminous fertility-building crops, with particular refence to N fixation and utilization. Defra Project OF0316 ‘The development of improved guidance on the use of fertility-building crops in organic farming’. https://de.scribd.com/document/147086691/A-review-of-leguminous-fertility-building-crops-with-particular-reference-to-nitrogen-fixation-and-utilisation. Accessed 17 Aug 2017
Elers B (2001) Einfluss der Sorte und Bestandesdichte auf Ertrag und Unkrautbesatz bei Futtererbsen. In: Reent HJ (ed) 6. Wissenschaftstagung zum Ökologischen Landbau „Von Leitbildern zu Leit-Linien“, Freising-Weihenstephan, 6.-8.3.2001. Verlag Dr. Köster, Berlin
Evans J, Taylor AC (1987) Estimating dinitrogen (N2) fixation and soil accretion of N by grain legumes. J Aust Inst Agric Sci 53:78–82
Evans J, O’Connor GE, Turner GL, Coventry DR, Fettell N, Mahoney J, Armstrong EL, Walsgott DN (1989) N2 fixation and its value to soil N increase in lupin, field pea and other legume in South-Eastern Australia. Aust J Agric Res 40:791–805
Frechilla S, Gonzalez EM, Royuela M, Minchin FR, Aparicio-Tejo PM, Arrese-Igor C (2000) Source of N nutrition (N fixation or nitrate assimilation) is a major factor involved in pea response to moderate water stress. J Plant Physiol 157:609–617
Freyer B, Pietsch G, Hrbek R, Winter S (2005) Futter- und Körnerleguminosen: Im biologischen Landbau. Österreichischer Agrarverlag, Wien
Fustec J, Lesuffleur F, Mahieu S, Cliquet JB (2010) N rhizodeposition of legumes. A review. Agron Sustain Dev 30:57–66
Grevsen K (2003) Weed competitive ability of green peas (Pisum sativum L.) affected by seeding rate and genotype characteristics. Biol Agric Hortic Int J Sustain Prod Syst 21(3):247–261
Hauggaard-Nielsen H, Ambus P, Jensen ES (2001) Interspecific competitions, N use and interference with weeds in pea-barley intercropping. Field Crops Res 70:101–109
Hauser S (1992) Estimation of symbiotically fixed N using extended N difference methods. In: Muolongoy K, Gueye M, Spencer DSC (eds) Biological N fixation and sustainability of tropical agriculture. Wiley-Sayce Co-Publication, West Sussex
Haynes RJ, Martin RJ, Goh KM (1993) N fixation, accumulation of soil N and N balance for some field-grown legume crops. Field Crops Res 35:85–92
Jensen ES (1996) Effect of pea cultivation on the soil N balance. Grain Legum 14:16–17
Jolliffe PA, Minjas AN, Runecles VC (1984) A reinterpretation of yield relationships in replacement series experiments. J Appl Ecol 21:227–243
Kelstrup L, Rowarth JS, William PH, Ronson C (1996) N fixation in peas (Pisum sativum L.) lupins (Lupinus angustifolius L.) and lentils (Lens culinaris Medik.). Proc Agron Soc NZ 26:71–74
Köpke U (1996) Symbiotische Stickstoff-Fixierung und Vorfruchtwirkung von Ackerbohnen (Vicia faba L.). Schriftenreihe Institut für Organischen Landbau 6:1–113. Verlag Dr. Köster, Berlin, Germany
Kübler E, Aufhammer W, Piepho HP (2006) Mischungseffekte in Getreide-Körnerleguminosen-Beständen auf den Kornertrag in Abhängigkeit vom Mischungsverhältnis. Die Bodenkultur 121:121–130
Kumar K, Goh KM (2000) Biological N fixation, accumulation of soil N and N balance for white clover (Trifolium repens L.) and field pea (Pisum sativum L.) grown for seed. Field Crops Res 68:49–59
Ledgard SF, Steele KW (1992) Biological N fixation in mixed legume grass pastures. Plant Soil 141:137–153
Mahieu S, Germon F, Aveline A, Hauggaard-Nielsen H, Ambus P, Jensen ES (2009) The influence of water stress on biomass and N accumulation, N partitioning between above and below ground parts and on N rhizodeposition during reproductive growth of pea (Pisum sativum L.). Soil Biol Biochem 41:380–387
Martin I, Tenorio JL, Ayerbe I (1994) Yield, growth and water-use of conventional and semi-leafless peas in semiarid environments. Crop Sci 34:1576–1583
McCauley AM, Clain AJ, Miller PR, Burgess MH, Zabinski CA (2012) N fixation by pea and lentil green manures in a semi-arid agroecoregion: effect of planting and termination timing. Nutr Cycl Agroecosyst 92:305–314
Moot DJ, McNeil DL (1995) Yield components, harvest index and plant type in relation to yield differences in field pea genotypes. Euphytica 86:31–40
Myers RJK, Wood IM (1987) Food legumes in the N cycle of farming systems. In: Wallis ES, Blyth DE (eds) Food legume improvement for Asian farming systems, ACIAR proceedings no. vol 18, pp 46–52. Australian Centre for International Agricultural Research, Canberra, Australia
Peoples MB, Herridge DF, Ladha JK (1995) Biological N fixation: an efficient source of N for sustainable agricultural production? Plant Soil 174:3–28
Reckling M, Döring TF, Stoddard FL, Bergkvist G, Watson CA, Seddig S, Chmielewski FM, Bachinger J (2018) Stability of grain legume yields is a matter of scaling. Landscape 2018, 12–16 March 2018, Berlin
Reiter K, Schmidtke K, Rauber R (2002) The influence of long-term tillage systems on symbiotic N2 fixation of pea (Pisum sativum L.) and red clover (Trifloium pratense L.). Plant Soil 238:41–55
Schlichting E, Blume H-P, Stahr K (1995) Bodenkundliches Praktikum—Eine Einführung in pedologisches Arbeiten für Ökologen, insbesonders Land- und Forstwirte und für Geowissenschaftler. 2., neubearbeitete Auflage. Blackwell Wissenschafts-Verlag, Berlin, Wien
Schmidtke K (1997) Schätzverfahren zur Ermittlung der N-Flächenbilanz bei Leguminosen. VDLUFA-Schriftenreihe 46. Kongreßband 1997:659–662
Schmidtke K (2013) How to optimize symbiotic N fixation in organic crop rotations. ISOFAR conference ‘Organic Agriculture in Asia’, Dankook University, Republic of Korea, 13–14 March 2008. http://orgprints.org/13272/. Accessed 10 May 2017
Schouls J, Langelaan JG (1994) Lodging and yield of dry peas (Pisum sativum L.) as influenced by various mixing ratios of a conventional and a semi-leafless cultivar. J Agron Crop Sci 172:207–214
Senaratne R, Hardarson G (1988) Estimation of residual N effect of faba bean and pea on two succeeding cereals using 15N methodology. Plant Soil 110:81–89
Serraj R, Sinclair TR, Purcell LC (1999) Symbiotic N2 fixation response to drought. J Exp Bot 50(331):143–155
Siegmund (1994) Einfluss des Pflanzenwuchstyps auf die N2-Fixierung von Körnererbsen (Pisum sativum L.). Dissertation Botanicae, Band 233, J. Cramer Verlag, Berlin Stuttgart
Syrovy LD, Banniza S, Shirtliffe SJ (2015) Yield an agronomic advantages of pea leaf type mixtures under organic management. Org Agric Agroecol 107(1):113–120
Tkachuk R (1966) Note on the N-to-protein conversion factor for wheat flour. Cereal Chem 43:223–225
Uzun A, Acikgöz E (1998) Effect of sowing season and seeding rate on the morphological traits and yields in pea cultivars of differing leaf types. J Agron Crop Sci 181:215–222
Wichmann S, Loges R, Taube F (2006) Kornerträge, N2-Fixierungsleistung und N-Flächenbilanz von Erbsen, Ackerbohnen und Schmalblättrigen Lupinen in Reinsaat und im Gemenge mit Getreide. Pflanzenbauwissenschaften 10(1):2–5
Acknowledgements
Open access funding provided by University of Natural Resources and Life Sciences Vienna (BOKU). We kindly acknowledge the staff of the research station of the University of Natural Resources and Life Sciences in Gross-Enzersdorf for assistance with field work at the experiment site. The technical assistance of Christoph Gabler and Erich Lang are also gratefully acknowledged, thanks are also extended to Benedikt Blankenhorn, who proofread the English text. We kindly acknowledge the Austrian ‘Federal Ministry of Sustainability and Tourism’ for supporting this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gollner, G., Starz, W. & Friedel, J.K. Crop performance, biological N fixation and pre-crop effect of pea ideotypes in an organic farming system. Nutr Cycl Agroecosyst 115, 391–405 (2019). https://doi.org/10.1007/s10705-019-10021-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-019-10021-4