
Abstract
The objective of this study was to assess genotype by environment interaction for seed yield in rapeseed cultivars grown in West Poland by the additive main effects and multiplicative interaction model. The study comprised 25 rapeseed genotypes (15 F1 CMS ogura hybrids, their parental lines and two varieties: Californium and Hercules F1), analyzed in five localities through field trials arranged in a randomized complete block design, with four replicates. Seed yield of the tested genotypes varied from 15.9 to 80.99 dt/ha throughout the five environments/localities, with an average of 39.69 dt/ha. In the variance analysis, 69.82 % of the total yield variation was explained by environment, 13.67 % by differences between genotypes, and 8.15 % by genotype by environment interaction. Seed yield is highly influenced by environmental factors. Due to high influence of the environment on yield high adaptability of the genome is required.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Oilseed rape (Brassica napus L.) is the most important oil seeds and oil meal (protein source) plant species in the countries with moderate climate, especially in the European Union (EU). In 2013, winter oilseed rape (canola, rapeseed) as the basic oilseed crop in European Union was grown over a total area of 6.7 M ha with the annual production of 21.0 Mt (FAOSTAT 2015, hhtp://faostat3.fao.org). The major producers of rapeseed in the EU (2013) were: France, Germany and Poland, and in the worldwide—Canada, China, India and Australia. Since the introduction, in the 90s, twentieth century double low oilseed rape cultivars—“zero” erucic acid and low glucosinolate content the growing area systematically increase, and the global production of rapeseed has tripled. Today, erucic acid-free rapeseed oil with a balanced fatty acid composition is considered optimal for nutritional purposes (salad oil, margarine), and for non-food purposes: biofuel production, tensides for detergent, biodegradable plastics and hydraulic oils (Friedt et al. 2007). The meal from oilseed rape after oil extraction also provides a protein-rich animal meal for all livestock and compounds of meal (Wittkop et al. 2009).
The profitability of oilseed rape products food and non-food depends on yielding capacity. Significant increase of production of oilseed rape can be achieve through increase of growing area or introducing high yield open-pollinated and hybrid varieties into production (Bartkowiak-Broda and Ogrodowczyk 2007). The yield potential of hybrid cultivars was reported at the level of 15 % above those of open-pollinated varieties. More than 50 % of the German oilseed rape growing area in 2007/2008 was planted with hybrid varieties and a trend that is reflected in all of the major growing areas for oilseed rape worldwide (Wittkop et al. 2009).
Seed yield is a very complex quantitative trait, which expression is the result of genotype, environment and the genotype × environment interaction. Complexity of this traits is a results different reactions of genotypes on changeable environmental conditions during plant development. Many breeders have understood the importance of genotype × environment interactions in plant breeding programme for a long time (Allard and Bradshaw 1964; Engqvist and Becker 1993). Özer et al. (1999), Sidlauskas and Bernotas (2003) reported that seed yield is one of the main yield components, which is greatly influenced by genotype, environment and complex genotype–environment interactions. Also Bocianowski et al. (2011) indicated that the main effects of cultivar and location were significant for yield and the traits directly influencing the quantity of seed yield. When field trials are carried out in different agroecological conditions, usually 80 % of yield variation is caused by environment, while genotype and genotype by environment interaction cause 10 % of variation each (Yan 2001). Ali et al. (2003) demonstrated also highly significant and positive correlations of seed yield with harvest index and seed weight. In crop production, yield stability is considered the most important socioeconomic category, especially in extreme environmental conditions (Piepho 1998; Ceccareli 1994). Therefore, it is fundamental to grow stable oilseed rape cultivars with good seed and oil yields in diverse agroecological conditions (Moghaddam and Pourdad 2011). Moreover, the results obtained in different study suggested that F1 rapeseed hybrids display a comparatively higher adaptation ability to adverse soil and climatic conditions than open-pollinating lines, besides their high yielding potential in general (Friedt et al. 2003; Seyis et al. 2006; Gehringer et al. 2007).
In field crop trials, this interaction is often analyzed by the additive main effects and multiplicative interaction (AMMI) model. This model was originally developed for analysis in social sciences and physics (Mandel 1961, 1969, 1971; Gollob 1968), and later adjusted for research in plant sciences (Gauch 1988, 1992; Cornelius 1993). The AMMI model is a combines the analysis of variance (ANOVA) with additive parameters and the principal component analysis (PCA) with multiplicative parameters in a single analysis. The AMMI biplot graphic display simultaneously both main and interaction effects for genotypes and environments, and enables a single analysis of the genotype by environment interaction. The AMMI is, therefore, also known as interaction PCA (Gauch and Zobel 1990), and can have several models: AMMI0, which estimates the additive main effect of genotypes and environments, and does not include any principal component axis (IPCA); AMMI1, which combines the additive main effects from AMMI0 with the genotype by environment interaction effects estimated from the first principal component axis (IPCA 1); AMMI2, and so forth, until the full model with all IPCA axis (Gauch 1988).
The objective of this study was to assess genotype by environment interaction for seed yield in oilseed rape grown in West Poland by the additive main effects and multiplicative interaction model.
Materials and methods
Plant material for field trials consisted of 25 winter rapeseed genotypes (Californium—open pollinated variety, Hercules F1—hybrid variety; five restorer lines for Ogura system (Rfo): PN05, PN07, PN17, PN18, PN21, three CMS ogura lines: PN64, PN66, PN68, and 15 hybrids: PN64 × PN17, PN64 × PN18, PN64 × PN21, PN64 × PN05, PN64 × PN07, PN66 × PN17, PN66 × PN18, PN66 × PN21, PN66 × PN05, PN66 × PN07, PN68 × PN17, PN68 × PN18, PN68 × PN21, PN68 × PN05, PN68 × PN07). Hybrids and their parental lines were selected from the rapeseed breeding program of the Plant Breeding and Acclimatization Institute – National Research Institute, Poznań, Poland.
The study was carried out during five locations: Bąków (E1), Borowo (E2), Łagiewniki (E3), Małyszyn (E4) and Zielęcin (E5) (Table 1). The field trials at all locations were arranged in a randomized complete block design, with four replicates. Each genotype was grown in a four row plot of 10.0 m2 (Borowo and Bąków), 12.0 m2 (Zielęcin), 9.6 m2 (Łagiewniki) and 11.2 m2 (Małyszyn) with a 0.30 row distance and a sowing density of 80 seeds/m2. Agricultural practices were optimal for local agroecological conditions in all investigated locations. Plots were harvested using a plot harvester. The field experiment was conducted on typical brown soil (E3, E4, E5) of quality class ranged from IIIa in E3 to IVb in E4 and grey-brown podzolic soil (E1, E2) of quality class IIIa in E1 and IIIB in E2. The topsoil was slightly acidic and ranged from pH 5.7 (E4) to 6.5 (E2). The previous crops were spring barley (E1, E2, E5), spring wheat (E3) and spring triticale (E4). Complex of agricultural usefulness were good rye in E1, E2, E4, E5 and good wheat in E3. In crop seasons 2007/2007, 2007/2008 and 2008/2009, weather conditions were normal for Poland. In 2007, frost occurred during flowering period of winter oilseed rape in Borowo. In 2008, however, abiotic stress occurred, caused by drought, during summer in 2008 in Bąków (germination and sprouting of winter oilseed rape). The sum of rainfall in critical period in autumn (92 mm) was markedly lower when compared with the same period 1957–2009 (128 mm). At the same time some frost damage was observed after the winter. Moreover, just before harvest in July 2009, was the hailstorm. The sum of rainfall was higher (265 mm) when compared with the average rainfall for the period 1957–2009 (212 mm) (Tables 2, 3).
Total grain yield were measured in dt per hectare (dt/ha) adjusted to 91 % dry matter. A two-way fixed effect model was fitted to determine the magnitude of the main effects of variation and their interaction on seed yield. Least–squares means were simultaneously produced for the AMMI model. Genotype main effect (G), environment main effect (E) and genotype by environment (GE) interaction were analyzed by the AMMI model (Gauch and Zobel 1990), represented by:
in which y ge is the yield mean of genotype g in environment e, μ is the grand mean, α g are the genotypic mean deviations (means minus grand mean), β e are the environmental mean deviations, N is the number of PCA axis retained in the adjusted model, λ n is the singular value for PCA axis n, γ gn is the genotype eigenvector for PCA axis n, δ en is the environment eigenvector for PCA axis n, Q ge are the residuals, including AMMI noise and pooled experimental error. Expected distribution of Q ge is normal.
The PCA analysis level of significance was tested with the F test according to Gollob (1968). In the biplot, which is the easiest way to represent the AMMI1 model, genotype by environment interactions are placed on the vertical axis (IPCA 1), while genotype and environment averages are placed on the horizontal axis. Applied analysis procedures, as well as the interpretation of results, were based on the protocol by Gauch and Zobel (1990). For the AMMI analysis, statistical package GenStat v. 17 was used.
Results and discussion
Effect of environmental conditions on biological and commercial traits of oilseed rape is very significant. The results of field trials demonstrated the impact of weather conditions (especially influence of temperature and precipitation in development of plants- germination, overwintering, flowering and level of seed yield), environment and genotypes on the seed yield of winter oilseed rape cultivars and lines. Unpredictable weather events have created a provocative experience conditions that influenced the development of plant and seed yield. Seed yield of the tested genotypes varied from 15.9 to 80.99 dt/ha throughout the five seasons, with an average of 39.69 dt/ha (Table 4). The PN7 (G25) line had the highest average yield, and the line PN18 (G04) had the lowest. The average yield per location also varied from 25.15 dt/ha, in Borowo, to 60.53 dt/ha in Małyszyn. All tested genotypes had the highest average yield in Małyszyn. During 2006–2009, wheather conditions were typical for Poland. However, in 2007 and 2009 there were strong abiotic stresses caused by a ground frost during flowering period in 2007 (April, May in Borowo) and frost damage in winter 2008/2009 and hailstorm just before harvest of winter oilseed rape in summer 2009 in Bąków.
The three sources of variation were highly significant. In the analysis of variance, the sum of squares for environment main effect represented 69.82 % of the total, and this factor had the highest effect on seed yield (Table 5). The differences between genotypes explained 13.67 % of the total yield variation, while the effects of GE interaction explained 8.15 %. Values for the three principal components were also highly significant. The three principal components of GE interaction accounted jointly for 93.75 % of the whole effect it had on the variation of seed yield. The first principal component (IPCA 1) accounted for 64.77 % of the variation caused by interaction, while IPCA 2 and IPCA 3 accounted for 16.45 and 12.53 %, respectively. The obtained lines have the best adaptability to the environmental conditions for which they were bred, which explains why 69.82 % of the variation in seed yield data was due to the main effects of locations.
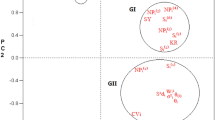
Traditional, statistical methods (ANOVA, PCA and linear regression) are often not effective for understanding and evaluating complex data from multi-environments yield trails. In contrast to the standard statistical analyses, AMMI incorporates ANOVA and PCA into a single model and enables simple visual interpretation of the GE interaction. AMMI model is usually constructed from the first two IPCA axes. Measuring GE interaction is very important to determine an optimum breeding strategy for releasing genotypes with an adequate adaptation to target environments (Fox et al. 1997). Among the tested genotypes, the line PN7 had the highest IPCA 1 value of 2.52, while the highest value of IPCA 1 was 3.39 in Małyszyn (Fig. 1).

Biplot for genotype by environment interaction of rapeseed (Brassica napus L.) lines in five environments, showing the effects of primary and secondary components (IPCA 1 and IPCA 2, respectively)
The clustering of some of the tested genotypes according to their IPCA 1 values and average yield on biplot (Fig. 1) also explains their similarities in yield per plant variations (Shafii et al. 1992). In general, environments with scores near zero have little interaction across genotypes and provide low discrimination among genotypes (Anandan et al. 2009); however, in this study, this pattern was not observed in any of the locations.
Genotype stability is considered as non significant reaction to changing environmental conditions, agronomic factors, weather conditions, biotic and abiotic stresses. In this study, climatic conditions were the source of this variation component. The stability of tested genotypes can be evaluated according to biplot for seed yield (Fig. 2). Experimental lines interacted differently with climate conditions in the observed locations. The lines G01, G02, G03, G04, G05, G06, G07, and G08 interacted positively with the E1 and E2 locations, but negatively with the E4 and E5 (Figs. 1, 2). The lines G10, G14, G21, and G25 interacted positively with the E4 and E5 locations, but negatively with the E1 and E2 locations. The analysis showed that some genotypes have high adaptation; however, most of them have specific adaptability (Yan and Hunt 1998; Atanasova et al. 2009). The lines G16 and G17 had stable and high average seed yield, with the IPCA 1 value closest to zero. Similar IPCA 1 values were found in the lines G12, G16, and G17. This kind of genotype is considered highly desirable in rapeseed breeding.

Biplot for the primary component of interaction (IPCA 1) and average rapeseed (Brassica napus L.) seed yield (dt/ha). Vertical line at the centre of biplot is the general grand mean
Genotypes on the highest point in certain sections of the graph have the best results in environments located in the same section (Fig. 2) (Yan 2001). Line G09, with average seed yield close to the general mean of 39.69 dt/ha, is distinguished on the biplot. This line had the highest stability. A group of lines: G11, G14, G16, G21, G24, G25 had the highest averages of yield, but with different adaptations (Figs. 1, 2): G25 showed specific adaptation to the conditions of Małyszyn, G10 to Zielęcin, and G16 showed general adaptation.
A graphically represented AMMI analysis enables selection of stable and high-yielding cultivars for this location, as well as cultivars with specific adaptability. The AMMI analysis is adequate in characterizing G × E interaction for seed yield in rapeseed. Marinković et al. (2011) reached the same conclusion in their study on sunflower under similar agroecological conditions in Southeastern Europe.
The AMMI model was often used in study of many species (Vargas et al. 1999; Hristov et al. 2010; Mehari et al. 2015). Results obtained from such analyses are very important for developing and recommending best cultivars for production in a specific area, as a selection criteria for further genetic improvements and can enable objective estimation of experimental genotypes and hence developing best possible varieties for official testing by national registration authorities (Mijić et al. 2007).
Conclusions
-
1.
The genotype and environment main effects and genotype by environment interaction effect were significant for oilseed rape genotypes studied in West Poland.
-
2.
The line G25 is recommended for further inclusion in the breeding program due to its high seed yield, G09 is recommended because of its stability and high seed yield.
-
3.
In addition to standard and commonly used data analysis techniques, biplot offer additional possibilities, preferably in the part of visual displaying and understanding of important interactions which are omni-present in the datasets from seed science research.
References
Ali N, Javidfar F, Elmira JY, Mirza MY (2003) Relationship among yield components and selection criteria for yield improvement in winter rapeseed (Brassica napus L.). Pak J Bot 35(2):167–174
Allard RW, Bradshaw AD (1964) Implications of genotypes-environment interactions in applied breeding. Crop Sci 4:503–508
Anandan A, Sabesan T, Eswaran R, Rajiv G, Muthalagan N, Suresh R (2009) Appraisal of environmental interaction on quality traits of rice by additive main effects and multiplicative interaction analysis. Cereal Res Commun 37(1):131–140
Atanasova D, Dochev V, Tsenov N, Todorov I (2009) Influence of genotype and environments on quality of winter wheat varieties in Northern Bulgaria. Agric Sci Technol 1(4):121–125
Bartkowiak-Broda I, Ogrodowczyk M (2007) Strategic tendencies in oilseed crops breeding. Monogr Disertation Plant Breed Acclim Inst 27:99–113 (in Polish)
Bocianowski J, Kozak M, Liersch A, Bartkowiak-Broda I (2011) A heuristic method of searching for interesting markers in terms of quantitative traits. Euphytica 181:89–100
Ceccareli S (1994) Specific adaptation and breeding for marginal conditions. Euphytica 77:205–219
Cornelius PL (1993) Statistical tests and retention of terms in the additive main effects and multiplicative interaction model for cultivar trials. Crop Sci 33:1186–1193
Engqvist MG, Becker HC (1993) Correlation studies for agronomic characters in segregating families of spring oilseed rape (Brassica napus). Hereditas 118:211–216
FAOSTAT 2015, http://faostat3.fao.org
Fox PN, Crossa J, Ramagosa I (1997) Multienvironment testing and genotype × environment interaction. In: Kempton RA, Fox PN (eds) Statistical methods for plant variety evaluation. Chapman & Hall, London, pp 117–138
Friedt W, Lühs W, Müller M, Ordon F (2003) Utility of winter oilseed rape (Brassica napus L.) cultivars and new breeding lines for low-input cropiping systems. German J Agron 2:49–55
Friedt W, Snowdon R, Ordon F, Ahlemeyer J (2007) Plant breeding: assessment of genetic diversity in crop plants and its exploitation in breeding. Prog Bot 168:152–177
Gauch HG (1988) Model selection and validation for yield trials with interaction. Biometrics 44:705–715
Gauch HG (1992) Statistical analysis of regional yield trials. AMMI analysis of factorial designs. Elsevier, New York, 278 p
Gauch HG, Zobel RW (1990) Imputing missing yield trial data. Theor Appl Genet 79:753–761
Gehringer A, Snowdon R, Spiller T, Basunanda P, Friedt W (2007) New oilseed rape (Brassica napus) hybrids with high levels of heterosis for seed yield under nutrient-poor conditions. Breed Sci 57:315–320
Gollob HF (1968) A statistical model which combines features of factor analytic and analysis of variance techniques. Psychometrika 33:73–155
Hristov N, Mladenov N, Djuric V, Kondic-Spika A, Marjanovic-Jeromela A, Simic D (2010) Genotype by environment interactions in wheat quality breeding programs in southeast Europe. Euphytica 174:315–324
Mandel J (1961) Non-additivity in two-way analysis of variance. J Am Stat Assoc 56:878–888
Mandel J (1969) The partitioning of interaction in analysis of variance. J Res Nat Bureau Stand Sect B 73:309–328
Mandel J (1971) A new analysis of variance model for nonadditive data. Technometrics 13:1–8
Marinković R, Jocković M, Marjanović-Jeromela A, Jocić S, Ćirić M, Balalić I, Sakač Z (2011) Genotype by environment interaction for seed yield and oil content in sunflower (H. annuus L.) using AMMI model. Helia 34:79–88
Mehari M, Tesfay M, Yirga H, Mesele A, Abebe T, Workineh A, Amare B (2015) GGE biplot analysis of genotype-by-environment interaction and grain yield stability of bread wheat genotypes in South Tigray, Ethiopia. Commun Biometry Crop Sci 10:17–26
Mijić A, Krizmanić M, Liović I, Zdunić Z, Marić S (2007) Response of sunflower hybrids to growing in different environments. Cereal Res Commun 35(2):781–784
Moghaddam MJ, Pourdad SS (2011) Genotype x environment interactions and simultaneous selection for high oil yield and stability in rainfed warm areas rapeseed (Brassica napus L.) from Iran. Euphytica 180(3):321–335
Özer H, Oral E, Doğru Ü (1999) Relationship between yield and yield components on currently improved spring rapeseed cultivars. Turkish J Agric For 23:603–607
Piepho HR (1998) Method for comparing the yield stability of cropping systems. J Agron Crop Sci 180:193–213
Seyis F, Friedt W, Lühs W (2006) Yield of Brassica napus L. hybrids developed using resynthesized rapeseed material sown at different locations. Field Crops Res 96:176–180
Shafii B, Mahler KA, Price WJ, Auld DL (1992) Genotype × environment interaction effects on winter rapeseed yield and oil content. Crop Sci 32:922–927
Sidlauskas G, Bernotas S (2003) Some factors affecting seed yield of spring rapeseed (Brassica napus L.). Agron Res 1:229–243
Vargas W, Crossa J, van Eeuwijk FA, Ramirez E, Sayre K (1999) Using partial least squars regression, factorial regression and AMMI models for interpreting genotype-by-environment interaction. Crop Sci 39:955–967
Wittkop B, Snowdon R, Friedt W (2009) Status and perspectives of breeding for enhanced yield and quality of oilseed crops for Europe. Euphytica 170:131–140
Yan W (2001) GGE biplot: a windows application for graphical analysis of multienvironment trial data and other types of two-way data. Agron J 93:1111–1118
Yan W, Hunt LA (1998) Genotype by environment interaction and crop yield. Plant Breed Rev 16:135–178
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Nowosad, K., Liersch, A., Popławska, W. et al. Genotype by environment interaction for seed yield in rapeseed (Brassica napus L.) using additive main effects and multiplicative interaction model. Euphytica 208, 187–194 (2016). https://doi.org/10.1007/s10681-015-1620-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-015-1620-z