
Abstract
The emerging amphibian disease, Batrachochytrium dendrobatidis (Bd), is driving population declines worldwide and even species extinctions in Australia, South and Central America. In order to mitigate effects of Bd on amphibian populations, high-exposed areas should be identified at the local scale and effective conservation measures should be planned at the national level. This assessment is actually lacking in the Mediterranean basin, and in particular in Italy, one of the most relevant amphibian diversity hotspots in the entire region. In this study, we reviewed the available information on Bd in Italy, and conducted a 5-year molecular screening on 1274 individual skin swabs belonging to 18 species. Overall, we found presence of Bd in 13 species and in a total of 56 known occurrence locations for peninsular Italy and Sardinia. We used these occurrence locations and climate data to model habitat suitability of Bd for current and future climatic scenarios. We then employed electric circuit theory to model landscape permeability to the diffusion of Bd, using a resistance map. With this procedure, we were able to model, for the first time, the diffusion pathways of Bd at the landscape scale, characterising the main future pathways towards areas with a high probability of Bd occurrence. Thus, we identified six national protected areas that will become pivotal for a nationally-based strategic plan in order to monitor, mitigate and possibly contrast Bd diffusion in Italy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
As part of the recent "biodiversity crisis," many amphibian populations are declining worldwide (e.g., Blaustein et al. 1994; Wake and Vredenburg 2008; Catenazzi 2015; Scheele et al. 2019). Indeed, the comprehensive analysis by Stuart et al. (2004) indicated that about one-third of all amphibian species is threatened with extinction, while almost half are experiencing regional or local declines due to often interacting causes, such as habitat reduction, pollution, climate change and emerging diseases. Among pathogens, the recently described chytrid fungus Batrachochytrium dendrobatidis (Longcore et al. 1999) is considered the main cause of population declines in different continents, driving species to extinction in Australia, South and Central America (Fisher and Garner 2020; Scheele et al. 2019). Batrachochytrium dendrobatidis (Bd) is a highly virulent pathogen that infects the skin of all the three orders of amphibians [i.e. Anura (frogs), Caudata (salamanders) and Apoda (caecilians)], causing chytridiomycosis, a frequently lethal disease that produces immunosuppression, depletion of plasma electrolytes and cardiac electric dysfunctions in amphibians (Berger et al. 2004, 2016).
The global Bd occurrence and its effects on amphibian populations have been recently reviewed and mapped (Olson et al. 2013; Lötters et al. 2009; Scheele et al. 2019). To date, six different Bd lineages have been identified by multilocus sequence typing, but only the global pandemic lineage (GPL) seems associated to widespread chytridiomycosis outbreaks that caused populations declines (Fisher and Garner 2020). The main factor spreading the pandemic Bd lineage in different parts of the world and in different time periods is the international trade of amphibians and other aquatic animals for food, research, collection or company (Olson et al. 2013).
Various studies, in the last fifteen years, mapped the known distribution of Bd-infected amphibian populations and analysed the possible future consequences of this disease at the global or continental level (e.g. Ron 2005; Becker and Zamudio 2011; Doherty-Bone et al. 2020; Ribeiro et al. 2020). However, this kind of assessment is lacking at the regional level in the Mediterranean biogeographic area, a well-known hotspot of natural and human-adapted ecosystems (Blondel and Aronson 1999; Myers et al. 2000). In particular, the central Mediterranean region has been shown as a potential suitable area for Bd by a variety of global studies (e.g. Fig. 4 in Ron 2005; Fig. 3 in Lötters et al. 2009; Fig. 2 in Liu et al. 2013). At the centre of the Mediterranean basin, Italy represents one of the most relevant hotspots of biodiversity with a high concentration of amphibian species and in particular of endemics (Sindaco et al. 2006). In fact, due to its central geographic position within the Mediterranean, complex geological history, contrasted geomorphology, variable climates and the long-lasting coevolution between rural landscapes and wildlife (Cevasco et al. 2015), Italy hosts a highly diverse and unique amphibian fauna, comprising about half of all amphibian species described in Europe (Table 1; Temple and Cox 2009; Rondinini et al. 2013). Various local studies have been published on Bd occurrence in Italy (see Table 1), but up to now only one mass-mortality event has been observed in Sardinia (Bielby et al. 2013; Tessa et al. 2013). In this island the pandemic lineage GPL has been recorded (Fisher and Garner 2020). Besides, in some areas along the Apennine mountain range the observed decline of the Apennine yellow-bellied toad, Bombina pachypus, was attributed to Bd infection (Stagni et al. 2004; Canestrelli et al. 2013). However, declines are also observed in other areas where Bd has been screened for, but is apparently absent (Canessa et al. 2013a, 2019). In these latter areas, the Apennine yellow-bellied toads’ declines were attributed to major habitat changes rather than pathogens (Canessa et al. 2013b). Therefore, the ecological effects of chytridiomycosis on the conservation status of Italian amphibians remain poorly understood, and in some cases enigmatic. For this reason, is of primary importance to delineate a nationwide monitoring and intervention plan, and model Bd occurrence to forecast possible future outbreaks.
In this research, we reviewed all the available information on Bd occurrence in Italian amphibian populations and we added original data, obtained over a five-year screening on peninsular Italy (Grasselli et al. unpublished data). Then we used both bibliographic and original data to model the present and near-future pathways of Bd diffusion in Italy, by both building habitat suitability models and ecological connectivity models at the landscape scale (McRae et al. 2008; Dickson et al 2019). This approach allowed us to forecast the most probable pathways of Bd diffusion and to better understand the impact of this disease on amphibian populations, within and outside protected areas, and to possibly identify the best mitigation measures to be implemented at the national and regional scale.
Materials and methods
Study rationale and framework
The principal aim of this study was to employ occurrence data of Bd in Italy, both from published research and original data, to model its current and future habitat suitability, to identify possible pathways in order to define a national monitoring network and to drive conservation measures. For that purpose, we collected all the available information on Bd occurrence data from published research that confirmed the presence of Bd by PCR and that possessed a reliable geographic locality. Moreover, we collected new data from 1274 skin swabs obtained between 2015 and 2019. Occurrence data were used to build habitat suitability models for both current and near future climatic conditions (year 2050), considering that climate has been found to have a predominant effect on Bd occurrence (e.g. Liu et al. 2013; Xie et al. 2016; Flechas et al. 2017). These habitat suitability models, in turn, have been used as a basis for building connectivity models by means of electric circuit theory, in order to delineate landscape permeability and to identify actual and future channels of Bd diffusion in Italy. Our aim is to identify which national protected areas will be most likely interested by massive diffusion of Bd in the near future. In this way, we will be able to design a rapid response monitoring network and to close possible knowledge gaps on the diffusion of Bd, concentrating the monitoring effort in those areas that will be interested by a rapid Bd spread (Fig. 1).

Study rationale and framework: (1) Batrachochytirum dendrobatidis presence data acquisition; (2) modelling Batrachochytirum dendrobatidis habitat suitability; (3) obtaining landscape permeability and forecasting possible spread pathways in present and future climate scenarios
Bd screening in Italy: field sampling and occurrence data
In the present study, we used data from multiple sources: published records and original data of Bd in peninsular and insular Italy. Bibliographic data were obtained from published research spanning from 2004 to 2015 (Table 1) and yielded 33 occurrence locations (Fig. 1; Supplementary Material Appendix 1, Table 1). Original data belong to a nationwide screening, conducted between 2015 and 2019 which mainly occurred in four national protected areas (Grasselli et al. unpublished data). We analysed 1274 amphibian swabs, belonging to 18 species, from 114 sampling locations. Swabbing procedures were standardised (Blooi et al. 2013) and field operators were formed during a 2-days field course held by the University of Genova, dedicated to the formation of national Parks’ staff and provided all participants with written instructions. Each swab was obtained by firmly rubbing its tip on the abdomen, pelvic area and leg pits of the captured individual (Boyle et al. 2004). All locations of Bd presence, both from published research and national screening, were georeferenced on a 1 × 1 km grid.
The extraction of nucleic acids from skin swabs for qPCR was performed as described by Boyle et al. (2004). Briefly, nucleic acids were extracted from swabs using 200 μl PrepMan Ultra along with 30–40 mg of Zirconium/silica beads (Biospec Products) and then incubated in a bead-beater. After centrifugation (1 min at 13 × 103 × g) and incubation in a boiling water bath for 10 min, supernatant was recovered and stored at—80 °C. The Bd SYBR green assay (Canessa et al. 2017) was performed in 15 μl reactions containing 7.5 μl of 2 × iQ™ universal SYBR® Green (Bio-Rad Laboratories, Hercules, CA), PCR primers at a concentration of 200 nM (fwd2: 5ʹ-CCTTGATATAATACAGTGTGCCAT-3ʹ; rev: 5ʹ-AGCCAA GAGATCCGTTGTCAAA-3ʹ) and 5 μl of samples were run in duplicate. Standards from 100, 10, 1, and 0.1 Bd genome equivalents (Bd GE; Verbrugghe et al. 2019) were a kind gift by Prof. An Martel and Frank Pasmans (University of Ghent, Belgium). Amplification occurred in CFX96 real-time PCR System (Bio-Rad Laboratories) with the following conditions: 3 min at 95 °C, followed by 15 s at 95 °C and 30 s at 60 °C for 40 cycles.
Habitat suitability models
Habitat suitability models were fitted using Maxent (version 3.4.0), a machine-learning algorithm that creates a model of habitat suitability (or probability of presence) for a given species, based on gridded predictors at observed species’ occurrence locations (Phillips et al. 2006). It compares environmental data at occurrence locations to the background data, where the presence of the species is unknown (Phillips et al. 2006; Guisan et al. 2017). The obtained model of habitat suitability can then be projected onto a different set of predictors, representing a different landscape or a future condition, thus obtaining a suitability model for another region or for a future scenario for the species of interest (Elith et al. 2011; Merow et al. 2013). This approach has been proved to outperform other methods based on presence-only (or presence background) data (Elith et al. 2006). Moreover, it also may have some advantages for the specific application on Bd data: (i) by relying on presence-only data there is no need for absence data that could be difficult to collect in the case of Bd, given the non-exhaustive sampling, the mixed origin of the data (i.e. bibliographic and original), (ii) Maxent algorithm proved to be reliable with a small amount of presence locations (Pearson et al. 2007; van Proosdij et al. 2016; Bacigalupe et al. 2019), which is the case of Bd occurrence data in the present study.
As environmental predictors, 19 bioclimatic variables representative of current and future (2050) climatic conditions were downloaded from Climatologies at high resolution for the earth’s land surface areas (CHELSA—Karger et al. 2017) website (https://chelsa-climate.org), at a resolution of 1 × 1 km, and cropped at the extent of the occurrence data (Italy borders). In order to reduce collinearity and overfitting in the model (MacNally 2002) we retained only those variables with a Pearson’s correlation coefficient <|0.8| (Dormann et al. 2013; Supplementary Material Appendix 1, Fig. S1). As a result, we retained 9 bioclimatic variables: BIO1—Annual mean temperature, BIO2—Mean temperature diurnal range, BIO5—Max temperature of warmest month, BIO6—Min temperature of coldest month, BIO8—Mean temperature of wettest quarter, BIO9—Mean temperature of driest quarter, BIO12—Annual precipitation, BIO13—Precipitation of wettest month, BIO14—Precipitation of driest month. In addition to bioclimatic variables, we also included in the predictor set a variable representing the density of the hydrographic network (HYD; Ribeiro et al. 2020). For projecting our habitat suitability model on future conditions, we considered two Representative Concentration Pathways (RCP): RCP2.6 and RCP8.5, representing the minimum and the maximum emission pathways for the year 2050 respectively. For each RCP we used projections from four CMIP5 Global Circulation Models (GCM): CCSM4, HadGEM2-CC, IPSL-CM5A-LR and MPI-ESM-LR. Since our main interest was to project Bd habitat suitability and spread to future climatic conditions, we avoided to include land cover, vegetation and anthropogenic variables in the models (despite these variables may contribute to determine Bd occurrence: e.g. Liu et al. 2013; James et al. 2015; Bacigalupe et al., 2019), because they may experience an unpredictable and radical change in a Mediterranean environment by year 2050, and therefore their reliability for future projections is doubtful.
For what concerns habitat suitability model building, availability of environmental conditions was drawn from 10,000 random background points. Model performance was determined using the area under the receiver operating characteristic curve (AUC), which ranges from 0.5 for completely random models to 1.0, for perfectly predictive models, and considering that models with an AUC > 0.8 have a good predictive ability (Merow et al. 2013). Random sampling of 75% of the data were used to fit the model while the remaining 25% were used to evaluate model performance. This procedure has been repeated 20 times and AUC values and habitat suitability model predictions were averaged from these repeated runs (Merow et al. 2013; Bacigalupe et al. 2019). Finally, we projected our habitat suitability model to each one of the four potential future climatic conditions forecasted by selected GCMs, and then we averaged GCMs projection within the same RCP, hence obtaining two habitat suitability model projections for year 2050, for RCP2.6 and RCP8.5 respectively (Guisan et al. 2017; Salas et al. 2017). Finally, we compared the gains and losses of suitability between the current and future bioclimatic conditions.
Landscape connectivity models
Connectivity models for Bd were built using electric circuit theory (McRae et al. 2008). Electric circuit theory calculates electrical flow between nodes on a resistance surface, with the flow representing dispersal/diffusion pathways across various types of resistances across the landscape (McRae et al. 2008). In particular, animal/pathogen movements are modelled via random walk (i.e. a random process describing a path that consist of subsequent random steps on the space) across all possible movement pathways, assuming that the intensity of current flow between a couple of nodes is proportional to the number of times an individual walks the path under consideration (McRae et al. 2008). In other words, current flow equals the likelihood of movement across the landscape, and therefore it represents landscape connectivity (McRae et al. 2008; Dickson et al. 2019). Specifically, circuit theory analysis produces a map of electric current density, equivalent to the likelihood of use for the given path (McRae et al. 2008), by considering all possible pathways connecting these nodes on a resistance/conductance map.
Circuit-theory connectivity models have been typically built by using occurrence locations or habitat patches as electrical nodes: however, this approach may be restrictive if the objective of the study is to estimate landscape permeability in the whole study area (Koen et al. 2014; Pitman et al. 2017). To overcome this problem, a “wall-to-wall” or “omnidirectional” approach has been applied: by placing current source and sink nodes outside of the study area (Koen et al. 2014; Pelletier et al. 2014; Pitman et al. 2017) or by using a moving-window algorithm (McRae et al. 2016), thus obtaining a landscape-scale connectivity map which is independent of nodes placement. Given the sparseness of Bd data in Italy, we opted to use a “wall-to-wall” method, independent of nodes’ placement.
We built circuit theory connectivity models using Omniscape connectivity algorithm implemented within Julia programming environment (McRae et al. 2016; Landau et al. 2021). This algorithm applies Circuitscape (McRae et al. 2013; Hall et al. 2021) iteratively across the entire map extent using a circular moving window of given radius, and using two raster maps as input: (i) the resistance map, defining the cost of crossing each pixel, and (ii) the source map, defining the amount of current to be injected in every pixel (McRae et al. 2016). Omniscape algorithm evaluates connectivity between every possible pair of pixels within the moving window, and then the resulting maps are summed to obtain a final map of cumulative current flow across the whole landscape.
We converted our habitat suitability models of Bd, for present and future conditions, to resistance maps (1- suitability) in order to model landscape permeability, an approach that has been successfully employed, among others, for frogs (Falaschi et al. 2018), martens (Balestrieri et al. 2019), leopards (Pitman et al. 2017), bears (Zeller et al. 2020) and elephants (Buchholtz et al. 2020). Given the sparseness of Bd data and considering that habitat suitability models also represent probability of presence of the modelled species in a given cell (Guisan et al. 2017), we also employed habitat suitability model values of every cell of the map as source values for the amount of current to be injected in the node. That is, if a cell has a suitability = 0.9, then a corresponding amount of current will be injected trough the corresponding node. We considered this approach to be more informative, rather than using few occurrence locations that are under-representative of the actual distribution of Bd across the study region (McRae et al. 2016), or by arbitrarily placing nodes outside (or randomly inside) of the study area (Koen et al. 2014; Pitman et al. 2017). In our analysis, we set a radius for the moving window equal to 50 km and a block size of 3 (i.e. a parameter coarsening the source surface: it represents the size, in pixels, of each block composing the moving window, at the centre of which the source point is located).
By running Omniscape algorithm, three different outputs for every habitat suitability model (one for current conditions and two for future conditions, RCP2.6 and RCP8.5, respectively) were produced: (i) cumulative current flow (CCF), which is the total current flowing through the landscape, (ii) flow potential (FP), which represents a null model of current flow, assuming movement unconstrained by resistance (i.e. using a constant resistance map), and (iii) normalized current flow (NCF; calculated as CCF/FP) which helps to distinguish between areas where current is impeded (i.e. movement is contrasted by barriers; NCF < 1), diffuse (i.e. no resistance to movement; NCF = 1), intensified (NFC > 1) or channelled (i.e. movement is constrained between barriers and is greater than expected by the null model; NFC > > 1; McRae et al. 2016). Finally, by overlying national protected areas’ borders to CCF and NFC maps, we calculated the mean current flow for each national protected area and identified those areas where the likelihood of Bd diffusion is higher for both present and future conditions (Littlefield et al. 2017; Choe and Thorne 2019).
Results
Bd screening in Italy: field sampling and occurrence data
We found presence of Bd in 71 out of 1274 skin swabs (corresponding to 5.5%), and Bd infection occurred in 13 species out of 18 tested by qPCR (Table 1). The overall Bd prevalence in Italian amphibians was 6% (Bayesian 95% credible intervals 4–7%), with a high variation among species (Table 1). Although, the present research does not focus on individual Bd load, in general the Bd load obtained by qPCR ranged from 1 to 140 Bd GEs per swab, which may be considered as a relatively low infection load. From our screening, we obtained 23 original occurrence locations in different parts of peninsular Italy to be added to bibliographic data, thus yielding to a total of 56 Bd occurrence location, that were employed for building habitat suitability model (Supplementary Material Appendix 1, Table 1).
Habitat suitability models
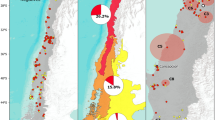
The habitat suitability model built for Bd had a good predictive performance (AUC = 0.879 ± 0.021; mean and standard deviation, calculated over 20 replicated runs). Out of the 10 predictor variables considered, BIO6 (Min temperature of coldest month), BIO8 (Mean temperature of wettest quarter) and BIO13 (Precipitation of wettest month) had the highest relative contribution to the maxent model (39.4%; 14.1%; 8.3% of variable contribution, respectively). At the same time BIO6 is the variable with the highest gain when used in isolation and which determines the highest decrease in gain when omitted from the model. As regards the current bioclimatic conditions, Maxent habitat suitability model shows that maximum habitat suitability for Bd is predicted in the northernmost and southernmost part of the Apennine mountain chain, in Sicily and Sardinia islands, while in the central part of Italy and in the Alps the suitability seems markedly lower (Fig. 2). Habitat suitability model projections on future conditions predict a loss of suitability for the most suitable areas actually identified with current climatic conditions, and conversely an increase of suitability in North-Western Alps and central Italy. This predicted change of suitability is markedly higher for the RCP8.5 with respect to RCP2.6 scenario (Fig. 2).

Habitat suitability model for Batrachochytrium dendrobatidis at current conditions (left side of the image) and maps of suitability changes (right side of the image) predicted for future conditions, at year 2050 under two representative concentration patwhays: RCP2.6 upper right and RCP8.5 lower right. Red dots and green diamonds, on the left side panel, represents original and bibliographic occurrence locations, respectively
Landscape connectivity models
Cumulative current flow (CCF) and Normalized current flow (NCF), obtained from the application of Omniscape algorithm, are presented in Figs. 3 and 4, respectively, for both current and future conditions. Cumulative current flow, which is the total current flowing through the landscape, shows a higher connectivity in the northernmost and southernmost portions of the Apennine mountain chain, and for Sardinia and Sicily islands, while landscape connectivity seems to be reduced in the central part of peninsular Italy. For future conditions, CCF seems to be reduced in the northern and southern extremes of the Apennine mountain chain, with respect to the current condition model, but landscape connectivity increases in central Italy, in particular when considering the most severe climate change scenario (RCP8.5—Fig. 3). For what concerns NCF, the flow appears as unconstrained in the major part of the landscape (Fig. 4), while in some areas the flow is channelled by barriers that create some pinch points: e.g. in the southernmost Apennines, Sicily and Sardinia islands and in some Alpine valleys as well. For future conditions NCF seems to be subject to smaller changes with respect to CCF, and as a general rule the amount of landscape portions with channelled flow is reduced in both RCP2.6 and RCP2.8 scenarios, while diffused and intensified flow increases overall (Fig. 4). At current conditions, there are four national protected areas experiencing a potentially high level of Bd diffusion: three in the southernmost part of Italy (Sila, Pollino, Aspromonte National Parks) and one in Sardinia (Golfo di Orosei/Gennargentu National Park; Fig. 5). The current flow in the remaining national protected areas is lower or even negligible. Conversely, for what concerns future bioclimatic conditions, connectivity models for both RCP2.6 and RCP8.5 predict a decrease of potential diffusion in these same protected areas (Sila, Pollino, Aspromonte, Golfo di Orosei/Gennargentu National Parks), while an increase of Bd flow for the Circeo National Park and Dolomiti Bellunesi National Park is expected (Fig. 5).

Cumulative current flow for Batrachochytrium dendrobatidis, derived from circuit theory connectivity model, at current conditions (left side of the image) and future conditions (right side of the image) at year 2050 under two representative concentration patwhays: RCP2.6 upper right and RCP8.5 lower right

Normalized current flow for Batrachochytrium dendrobatidis derived from circuit theory connectivity model, at current conditions (left side of the image) and future conditions (right side of the image) at year 2050 under two representative concentration patwhays: RCP2.6 upper right and RCP8.5 lower right

Mean cumulative current flow for Batrachochytrium dendrobatidis in Italian National Parks, derived from circuit theory connectivity model, at current conditions (red squares) and future conditions at year 2050 under two representative concentration pathways: RCP2.6 (green triangles) and RCP8.5 (blue diamonds)
Discussion
In peninsular Italy, Bd is present all along the Apennine mountain chain and also in the Po plain, with an overall prevalence of 6%. When this value was compared with other European countries, with a reported sample larger than 1000 individuals, a similar prevalence was found in Germany (7%, in 3064 individuals) but a much higher one was observed in Spain (20%, in 1149 individuals; data from supplementary materials in Baláž et al. 2013). However, this latter high prevalence may be explained by an over-sampling of amphibians in areas with well-known Bd outbreaks, such as Central Spain and the island of Majorca (Bosch et al. 2001; Bosch and Martinez-Solano 2006; Walker et al. 2008; Baláž et al. 2013). Concerning the individual Bd load, the Italian data were in the lowest range of those reported by Baláž et al. (2013) that measured up to 4067 GEs, but usually much lower.
The application of species distribution and habitat suitability models to the chytrid amphibian fungus has been widespread in the last 15 years. Several studies modelled the current distribution and habitat suitability of Bd at a global (e.g. Ron 2005; Rödder et al. 2009; Lötters et al. 2009; Liu et al. 2013; Xie et al. 2016) or continental scale (e.g. James et al. 2015; Rahman et al. 2020; Zimkus et al. 2020); while other studies investigated Bd distribution at a smaller, regional scale, focusing on specific amphibian diversity hot-spots (e.g. Puschendorf et al. 2009; Seimon et al. 2015; Flechas et al. 2017; Miller et al. 2018; Bacigalupe et al. 2019). However, ecological relationships among hosts and parasites are complex and the outcomes of their interactions vary in association with global environmental variables but also with the complexity of the biological community at the local scale (Benício et al. 2019, Halliday et al. 2019). Indeed, this relationship seems to be generally non-linear and that high biodiversity may dilute parasite occurrence (Halliday and Rohr 2019). This implies that highly diverse and rich ecosystems could inhibit the diffusion of wildlife diseases at the local scale. Therefore, the role of areas with complex amphibian communities could act as ecological barriers to Bd spread, and should become important areas for the monitoring of the chytrid fungus. Among the studies modelling Bd occurrence at a global scale, several identify Italy as a high-suitability/high-risk area for Bd (Rödder et al. 2009; Lötters et al. 2009; Liu et al. 2013). Within our study, we identified some specific area of high suitability for Bd in Italy (e.g. Sila, Pollino, Aspromonte National Parks), while other areas were predicted to be less suitable at the current climatic conditions (e.g. Foreste Casentinesi National Park). The major part of the studies inferring habitat suitability and distribution of Bd, highlighted an overwhelming effect of climate. For instance: Puschendorf et al. (2009) found that high temperature seems to constrain the distribution of the pathogen at small scale in Costa Rica, Flechas et al. (2017) identified mean temperature and precipitation seasonality as main drivers of Bd in Colombia, while Liu et al. (2013) observed a relationship with annual temperature range at a global scale. In this study, among the bioclimatic variables included in the habitat suitability model, we observed that extremely high or low temperatures, in particular in the wettest quarter of the year, (BIO6 and BIO8) were main predictors of Bd suitability and acted as limiting conditions for his occurrence. These results can be explained by both an increased rate of epidermal renewal, driven by higher temperatures, which may in turn reduce Bd infection (Piotrowski et al. 2004), or alternatively producing physiological stress, which may limit Bd reproductive success (Piotrowski et al. 2004). Besides climate, also vegetation (Liu et al. 2013), land cover (James et al. 2015) and anthropogenic factors have been found to shape the distribution of Bd (Liu et al. 2013; Bacigalupe et al. 2019).
Habitat suitability and distribution models also confirmed the link between epizootic chytridiomycosis and amphibian worldwide decline, highlighting how areas of rapid amphibian decline overlaps with those of higher suitability for Bd at a global scale (Rödder et al. 2009; Lötters et al. 2009; James et al. 2015). The majority of studies modelling Bd distribution only focused on current climatic conditions, while few studies also projected distribution and suitability models on future climatic scenarios, predicting that Bd may decrease globally in some regions by 2100, but with a shift towards higher latitude and altitudes (Xie et al. 2016). This trend has also been confirmed at a smaller spatial scale (e.g. Seimon et al. 2015; Xie et al. 2016; Miller et al. 2018). In the present study we also observed a general contraction of Bd suitable areas by 2050, considering both circulation pathways. In particular, we observed that the suitability loss will mainly occur in southern and coastal Italy, while suitability gain will be observed in mountainous areas, such as central Apennines, western Alps and eastern pre-Alps and Alps. Despite the variety of approaches employed, local studies identified two main issues: (i) the identification of high-risk and refuge area will be of primary importance, (ii) the inadequacy of local strategies to monitor and mitigate Bd expansion (e.g. Flechas et al. 2017; Bacigalupe et al. 2019; Rahman et al. 2020). In Italy, while we addressed the first issue by both collecting new occurrence data and modelling habitat suitability, we also tried to overcome the second problem by modelling diffusion pathways at the landscape scale. Applying this procedure, we identified the national protected areas that should be more involved in monitoring and prevention actions to mitigate Bd diffusion.
Connectivity models based on electric circuit theory, have been widely adopted to model the spread of diseases or pathogens in wildlife populations, such as deer chronic wasting disease (Nobert et al. 2016), or rabies in raccoon populations (Algeo et al. 2017). Notably, among the almost 300 studies using circuit theory biology, ecology and conservation science (Dickson et al. 2019), only one involved the amphibian chytrid pathogen Bd (i.e. Becker et al. 2017). By modelling forest connectivity between populations of an Hylid frog in Brazil, Becker et al. (2017) found that skin microbiome similarity and Bd load are related to landscape connectivity and natural vegetation gradient, but a landscape connectivity model of Bd spread is actually lacking. Therefore, our study, at least to our knowledge, is the first one employing circuit theory to model the diffusion of Bd at the landscape scale. By applying this method, we were able to identify four national protected areas (Pollino, Sila, Aspromonte and Golfo di Orosei and Gennargentu National Parks) that may experience maximum Bd diffusion for the current climatic conditions (i.e. Cumulative Current Flow). These areas of maximum diffusion are also identified by the presence of channelled or intensified movement as predicted by normalized current flow, meaning that in these areas the spread rate is potentially high (McRae et al. 2016).
For these areas, we suggest the development of an intensive molecular screening plan in order to track any possible change in Bd presence, population prevalence and individual load of resident amphibians. Furthermore, three out of four of these areas are in geographic and ecological connectivity (Pollino, Sila and Aspromonte National Parks), as resulted from our model, while the fourth area (Maddalena national Park) is in ecological connection with populations interested by a Bd induced mass-mortality that occurred in Sardinia (Bielby et al. 2013; Tessa et al. 2013). Indeed, according to the Italian national biodiversity strategy, national parks should become “focal points for research and monitoring networks … in terms of biodiversity” (MATTM 2010, page 31). Therefore, national parks identified as areas of high Bd diffusion will represent a fundamental tool to implement monitoring, awareness and mitigation strategies. Moreover, we suggest that a particular monitoring effort should be spent in those area where current and predicted Bd flow is channelled between climatic suitability barriers, within a national park or protected area, such as Dolomiti Bellunesi in the Alpine region and Circeo National Parks in the central region of the Apennine mountain chain. Finally, despite the fact that our study allowed the identification of high priority areas, in order to detect early Bd spread and diffusion; our results are obtained from an incomplete sampling of the national area. Therefore, the identification of National Parks (or other priority areas) involved in the monitoring network could benefit from a more intensive sampling effort or from the application of different techniques: such as the implementation of a national monitoring programme, relying upon environmental DNA sampling and Occupancy modelling (Schmidt et al. 2013).
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Adams JA, Galvan S, Scalera S, Grieco C, Sindaco R (2004) Batrachochytrium dendrobatidis in amphibian populations in Italy. Herpetol Rev 39:324–326
Algeo TP, Slate D, Caron RM, Atwood T, Recuenco S, Ducey MJ, Chipman RB, Palace M (2017) Modeling raccoon (Procyon lotor) habitat connectivity to identify potential corridors for rabies spread. Trop Med Infect Dis 2:44
Bacigalupe LD, Vásquez IA, Estay SA, Valenzuela-Sánchez A, Alvarado-Rybak M, Peñafiel-Ricaurte A, Cunningham AA, Soto-Azat C (2019) The amphibian-killing fungus in a biodiversity hotspot: identifying and validating high-risk areas and refugia. Ecosphere 10:02724
Baláž V, Vörös J, Civiš P, Vojar J, Hettyey A, Sós A, Dankovics R, Jehle R, Christiansen DG, Clare F, Fisher MC, Garner TWJ, Bielby J (2013) Assessing risk and guidance on monitoring of Batrachochytrium dendrobatidis in Europe through identification of taxonomic selectivity of infection. Conserv Biol 28:213–223
Balestrieri A, Mori E, Menchetti M, Ruiz-González A, Milanesi P (2019) Far from the madding crowd: tolerance toward human disturbance shapes distribution and connectivity patterns of closely related Martes spp. Pop Ecol 61:289–299
Becker CG, Zamudio KR (2011) Tropical amphibian populations experience higher disease risk in natural habitats. Proc Nat Acad Sci USA 108:9893–9898
Becker CG, Longo AV, Haddad CFB, Zamudio KR (2017) Land cover and forest connectivity alter the interactions among host, pathogen and skin microbiome. Proc R Soc B 284:20170582
Benício RA, Carvalho T, Barbosa MD, Costa JDM, da Silva FC, Fonseca MG (2019) Worrying News for Brazilian Caatinga: prevalence of Batrachochytrium dendrobatidis in Amphibians. Trop Conserv Sci 12:1940082919892626
Berger L, Speare R, Hines H, Marantelli G, Hyatt AD, McDonald KR, Skerratt LF, Olsen V, Clarke JM, Gillespie G, Mahony M, Sheppard N, Williams C, Tyler M (2004) Effect of season and temperature on mortality in amphibians due to chytridiomycosis. Austr Vet J 82:434–439
Berger L, Roberts AA, Voyles J, Longcore JE, Murray CA, Skerratt LF (2016) History and recent progress on chytridiomycosis in amphibians. Fungal Ecol 19:89–99
Bielby J, Bovero S, Angelini C, Favelli M, Gazzaniga E, Perkins M, Sotgiu G, Tessa G, Garner TWJ (2013) Geographic and taxonomic variation in Batrachochytrium dendrobatidis infection and transmission within a highly endemic amphibian community. Divers Distrib 19:1153–1163
Blaustein AE, Wake DB, Sousa WP (1994) Amphibian declines: judging stability, persistence, and susceptibility of populations to local and global extinctions. Conserv Biol 8:60–71
Blondel J, Aronson J (1999) Biology and wildlife of the Mediterranean region. Oxford University Press, Oxford
Blooi M, Pasmans F, Longcore JE, Spitzen-van der Sluijs A, Vercammen F, Martel A (2013) Duplex real-time PCR for rapid simultaneous detection of Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans in Amphibian samples. J Clin Bicrobiol 51:4173–4177
Bosch J, Martinez-Solano I (2006) Chytrid fungus infection related to unusual mortalities of Salamandra salamandra and Bufo bufo in the Peñalara Natural Park, Spain. Oryx 40:84–89
Bosch J, Martinez-Solano I, Garcia-Paris M (2001) Evidence of a chytrid fungus infection involved in the decline of the common midwife toad (Alytes obstetricans) in protected areas of central Spain. Biol Conserv 97:331–337
Boyle DG, Boyle DB, Olsen V, Morgan JAT, Hyatt AD (2004) Rapid quantitative detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using real-time Taqman PCR assay. Dis Aquati Org 60:141–148
Buchholtz EK, Stronza A, Songhurst A, McCulloch G, Fitzgerald LA (2020) Using landscape connectivity to predict human-wildlife conflict. Biol Conserv 248:108677
Canessa S, Martel A, Pasmans F (2013a) No detection of chytrid in first systematic screening of Bombina variegata pachypus (Anura: Bombinatoridae) in Liguria, northern Italy. Acta Herpetol 8:59–63
Canessa S, Oneto F, Ottonello D, Arillo A, Salvidio S (2013b) Land abandonment may reduce disturbance and affect the breeding sites of an endangered amphibian in northern Italy. Oryx 47:280–287
Canessa S, Maggesi M, Salvidio S, Grasselli E (2017) Validation and cost-effectiveness of an alternative method to quantify Batrachochytrium dendrobatidis infection in amphibian samples using real-time PCR. Rend Lincei Sci Fis Nat 28:687–692
Canessa S, Ottonello D, Rosa G, Salvidio S, Grasselli E, Oneto F (2019) Adaptive management of species recovery programs: a real-world application for an endangered amphibian. Biol Conserv 236:202–210
Canestrelli D, Zampiglia M, Nascetti G (2013) Widespread occurrence of Batrachochytrium dendrobatidis in contemporary and historical samples of the endangered Bombina pachypus along the Italian peninsula. PLoS ONE 8:63349
Catenazzi A (2015) State of the World’s Amphibians. Ann Rev Environ Res 40:91–119
Cevasco R, Moreno D, Hearn R (2015) Biodiversification as an historical process: an appeal for the application of historical ecology to bio-cultural diversity research. Biodiv Conserv 24:3167–3183
Choe H, Thorne JH (2019) Omnidirectional connectivity of urban open spaces provides context for local government redevelopment plans. Landsc Ecol Eng 15:245–251
Dickson BG, Albano CM, Anantharaman R, Beier P, Fargione J, Graves TA, Gray ME, Hall KR, Lowler JJ, Leonard PB, Littlefield CE, McClure ML, Novembre J, Schloss CA, Shumaker NH, Shah VB, Teobald DM (2019) Circuit-theory applications to connectivity science and conservation. Conserv Biol 33:239–249
Doherty-Bone TM, Cunningham AA, Fisher MC, Garner TWJ, Ghosh P, Gower DJ, Verster R, Weldon C (2020) Amphibian chytrid fungus in Africa–realigning hypotheses and the research paradigm. Anim Conserv 23:239–244
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, García Marquéz R, Gruber B, Lafourcade B, Leitão P, Münkemüller T, McClean C, Osborne PE, Beineking B, Schröder B, Skidmore A, Zurell D, Lautenbach S (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Elith J, Graham H, Anderson CP, Dudík M, Ferrier S, Guisan A, Hijmans JR, Huettmann F, Leathwick RJ, Lehmann A, Li J, Lohmann LC, Loiselle BA, Manion G, Moritz C, Nakamura M, Nakazawa Y, Jacob MCM, Townsend Peterson OA, Phillips SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Falaschi M, Mangiacotti M, Sacchi R, Scali S, Razzetti E (2018) Electric circuit theory applied to alien invasions: a connectivity model predicting the Balkan frog expansion in Northern Italy. Acta Herpetol 13:33–42
Federici S, Clemenzi S, Favelli M, Tessa G, Andreone F, Casiraghi M, Crottini A (2008) Identification of the pathogen Batrachochytrium dendrobatidis in amphibian populations of a plain area in the Northwest of Italy. Herpetol Notes 1:33–37
Ficetola GF, Valentini A, Miaud C, Noferini A, Mazzotti S, Dejean T (2011) Batrachochytrium dendrobatidis in amphibians from the Po River Delta, Northern Italy. Acta Herpetol 6:297–302
Fisher MC, Garner TWJ (2020) Chytrid fungi and global amphibian declines. Nat Rev Microbiol 18:332–334
Flechas SV, Paz A, Crawford AJ, Sarmiento C, Acevedo AA, Arboleda A, Bolívar-García W, Echeverry-Sandoval CL, Franco R, Mojica C, Muñoz A, Palacios-Rodríguez P, Posso-Terranova AM, Quintero-Marín P, Rueda-Solano LA, Castro-Herrera F, Amézquita A (2017) Current and predicted distribution of the pathogenic fungus Batrachochytrium dendrobatidis in Colombia, a hotspot of amphibian biodiversity. Biotropica 49:685–694
Guisan A, Thuiller W, Zimmermann NE (2017) Habitat suitability and distribution models: with applications in R. Cambridge University Press, Cambdrige
Hall KR, Anantharaman R, Landau VA, Clark M, Dickson BG, Jones A, Platt J, Edelman A, Shah VB (2021) Circuitscape in julia: empowering dynamic approaches to connectivity assessment. Land 10:301
Halliday FW, Rohr JR (2019) Measuring the shape of the biodiversity-disease relationship across systems reveals new findings and key gaps. Nat Commun 10:5032
Halliday FW, Heckman RW, Wilfahrt PA, Mitchell CE (2019) Past is prologue: host community assembly and the risk of infectious disease over time. Ecol Lett 22:138–148
James TY, Toledo LF, Rödder D, da Silva LD, Belasen AM, Betancourt-Román CM, Jenkinson TS, Soto-Azat C, Lambertini C, Longo AV, Ruggeri J, Collins JP, Burrowes PA, Lips KR, Zamudio KR, Longcore JE (2015) Disentangling host, pathogen, and environmental determinants of a recently emerged wildlife disease: lessons from the first 15 years of amphibian chytridiomycosis research. Ecol Evol 5:4079–4097
Karger DN, Conrad O, Böhner J, Kawohl T, Kreft H, Soria-Auza RW, Zimmermann NE, Linder HP, Kessler M (2017) Climatologies at high resolution for the earth’s land surface areas. Sci Data 4:170122
Koen EL, Bowman J, Sadowski C, Walpole AA (2014) Landscape connectivity for wildlife: development and validation of multispecies link- age maps. Methods Ecol Evol 5:626–633
Landau VA, Shah VB, Anantharaman R, Hall KR (2021) Omniscape. jl: Software to compute omnidirectional landscape connectivity. J Open Source Softw 6:2829
Littlefield CE, McRae BH, Michalak JL, Lawler JJ, Carroll C (2017) Connecting today’s climates to future climate analogs to facilitate movement of species under climate change. Conserv Biol 31:1397–1408
Liu X, Rohr JR, Li Y (2013) Climate, vegetation, introduced hosts and trade shape a global wildlife pandemic. Proc Royal Soc B 280:20122506
Longcore JE, Pessier AP, Nichols DK (1999) Batrachochytrium dendrobatidis gen. et sp. nov., a chytrid pathogenic to amphibians. Mycologia 91:219–227
Lötters S, Kielgast J, Bielby J, Schmidtlein S, Bosch J, Veith M, Walker SF, Fisher MC, Rödder D (2009) The link between rapid enigmatic amphibian decline and the globally emerging chytrid fungus. EcoHealth 6:358–372
MacNally R (2002) Multiple regression and inference in ecology and conservation biology: further comments on identifying important predictor variables. Biodivers Conserv 11:1397–1401
MATTM (2010) Italian National Biodiverisy Strategy. www.minambiente.it accessed on November 2020.. Accessed Nov 2020
McRae BH, Dickson BG, Keitt TH, Shah VB (2008) Using circuit theory to model connectivity in ecology, evolution, and conservation. Ecology 89:2712–2724
McRae BH, Shah VB, Mohapatra TK (2013) Circuitscape 4 user guide. The Nature Conservancy, Fort Collins, CO
McRae BH, Popper K, Jones A, Schindel M, Buttrick S, Hall K, Unnasch B, Platt J (2016) Conserving nature’s stage: mapping omnidirectional connectivity for resilient terrestrial landscapes in the pacific northwest. The Nature Conservancy, Portland, OR
Merow C, Smith MJ, Silander JA (2013) A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter. Ecography 36:1058–1069
Miller CA, Tasse Taboue GC, Ekane MMP, Robak M, Sesink Clee PR, Richards-Zawacki C, Fokam EB, Fuashi NA, Anthony NM (2018) Distribution modeling and lineage diversity of the chytrid fungus Batrachochytrium dendrobatidis (Bd) in a central African amphibian hotspot. PLoS ONE 13:0199288
Myers N, Mittermeier RA, Mittermeir CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Nobert BR, Merrill EH, Pybus MJ, Bollinger TK, Hwang YT (2016) Landscape connectivity predicts chronic wasting disease risk in Canada. J Appl Ecol 53:1450–1459
Olson DH, Aanensen DM, Ronnenberg KL, Powell CI, Walker SF, Bielby J, Garner TWJ, Weaver G, Bd Mapping Group, Fisher MC (2013) Mapping the global emergence of Batrachochytrium dendrobatidis, the amphibian chytrid fungus. PLoS ONE 8:56802
Pearson RG, Raxworthy CJ, Nakamura M, Townsend Peterson A (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117
Pasmans F, Van Rooij P, Blooi M, Tessa G, Bogaerts S, Sotgiu G, Garner TWJ, Fisher MC, Schmidt BR, Woeltjes T, Beukema W, Bovero S, Adriaensen, C, Oneto F, Ottonello D, Martel A, Salvidio S (2013) Resistance to Chytridiomycosis in European Plethodontid Salamanders of the Genus Speleomantes. PLoS ONE 8:63639
Pelletier D, Clark M, Anderson MG, Rayfield B, Wulder MA, Cardille JA (2014) Applying circuit theory for corridor expansion and management at regional scales: tiling, pinch points, and omnidirectional connectivity. PLoS ONE 9:84135
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Modell 190:231–259
Piotrowski JS, Annis SL, Longcore JE (2004) Physiology of Batrachochytrium dendrobatidis, a chytrid pathogen of amphibians. Mycologia 96:9–15
Pitman RT, Fattebert J, Williams ST, Williams KS, Hill RA, Hunter LTB, Robinson H, Power J, Swanepoel L, Slotow R, Balme GA (2017) Cats, connectivity and conservation: incorporating data sets and integrating scales for wildlife management. J Appl Ecol 54:1687–1698
Puschendorf R, Carnaval AC, VanDerWal J, Zumbado-Ulate H, Chaves G, Bolaños F, Alford RA (2009) Distribution models for the amphibian chytrid Batrachochytrium dendrobatidis in Costa Rica: proposing climatic refuges as a conservation tool. Divers Distrib 15:401–408
Rahman M, Badhon MK, Salauddin M, Rabbe F, Islam S (2020) Chytrid infection in Asia: how much do we know and what else do we need to know? Herpetol J 30:99–111
Ribeiro JW Jr, Siqueira T, DiRenzo G, Lambertini C, Lyra ML, Toledo LF, Haddad CFB, Becker G (2020) Assessing amphibian disease risk across tropical streams while accounting for imperfect pathogen detection. Oecologia 193:237–248
Rödder D, Kielgast J, Bielby J, Schmidtlein S, Bosch J, Garner TWJ, Veith M, Walker S, Fisher MC, Lötters S (2009) Global amphibian extinction risk assessment for the panzootic chytrid fungus. Diversity 1:52–66
Ron SR (2005) Predicting the distribution of the amphibian pathogen Batrachochytrium dendrobatidis in the New World 1. Biotropica 37:209–221
Rondinini C, Battistoni A, Peronace V, Teofili C (2013) Lista Rossa IUCN dei Vertebrati Italiani. Comitato Italiano IUCN, Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Roma
Salas EAL, Seamster VA, Harings NM, Boykin KG, Alvarez G, Dixon KW (2017) Projected future bioclimate-envelope suitability for reptile and amphibian species of concern in South Central USA. Herpetol Conserv Biol 12:522–547
Scheele BC, Pasmans F, Skerratt LF et al (2019) Amphibian fungal panzootic causes catastrophic and ongoing loss of biodiversity. Science 363:1459–1463
Schmidt BR, Kery M, Ursenbacher S, Hyman OJ, Collins JP (2013) Site occupancy models in the analysis of environmental DNA presence/absence surveys: a case study of an emerging amphibian pathogen. Met Ecol Evol 4:646–653
Seimon TA, Ayebare S, Sekisambu R, Muhindo E, Mitamba G, Greenbaum E, Menegon M, Pupin F, McAloose D, Ammazzalorso A, Meirte D, Lukwago W, Behangana M, Seimon A, Plumptre AJ (2015) Assessing the threat of amphibian chytrid fungus in the Albertine Rift: Past, present and future. PLoS ONE 10:0145841
Simoncelli F, Fagotti A, Dall’Olio R, Vagnetti D, Pascolini R, Di Rosa I (2005) Evidence of Batrachochytrium dendrobatidis infection in water frogs of the Rana esculenta complex in central Italy. EcoHealth 2:307–312
Sindaco R, Doria G, Razzetti E, Bernini F (2006) Atlante degli Anfibi e Rettili d’Italia—Atlas of Amphibians and Reptiles in Italy. SHI, Edizioni Polistampa, Firenze
Stagni G, Dall’Olio R, Fusini U, Mazzotti S, Scoccianti C, Serra A (2004) Declining populations of apennine yellow-bellied toad Bombina pachypus in the northern apennines (italy): is Batrachochytrium dendrobatidis the main cause? Ital J Zool 71:151–154
Stuart SN, Chanson JS, Cox NA, Young BE, Rodrigues ASL, Fischman DL, Waller RW (2004) Status and trends of amphibian declines and extinctions worldwide. Science 306:1783–1786
Temple HJ, Cox NA (2009) European Red List of Amphibians. Office for Official Publications of the European Communities, Luxembourg
Tessa G, Angelini C, Bielby J, Bovero S, Giacoma C, Sotgiu G, Garner TWJ (2013) The pandemic pathogen of amphibians, Batrachochytrium dendrobatidis (Phylum Chytridiomycota), in Italy. It J Zool 80:1–11
van Proosdij ASJ, Sosef MSM, Wieringa JJ, Raes N (2016) Minimum required number of specimen records to develop accurate species distribution models. Ecography 39:542–552
Verbrugghe E, Pasmans F, Martel A (2019) Reference gene screening of Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans for quantitative real-time PCR studies. Sci Rep 9:18534
Wake DB, Vredenburg VT (2008) Are we in the midst of the sixth mass extinction? A view from the world of amphibians. Proc Natl Acad Sci USA 105:11466–11473
Walker SF, Bosch J, James TY, Litvintseva AP, Oliver Valls JA, Pina S, Garcia G, Rosa GA, Cunningham AA, Hole S, Griffith R, Fisher MC (2008) Invasive pathogens threaten species recovery programs. Curr Biol 18:853–854
Xie GY, Olson DH, Blaustein AR (2016) Projecting the global distribution of the emerging amphibian fungal pathogen, Batrachochytrium dendrobatidis, Based on IPCC Climate Futures. PLoS ONE 11:0160746
Zampiglia M, Canestrelli D, Chiocchio A, Nascetti G (2013) Geographic distribution of the chytrid pathogen Batrachochytrium dendrobatidis among mountain amphibians along the Italian peninsula. Dis Aquat Organ 107:61–68
Zimkus BM, Baláž V, Belasen AN, Bell CR, Channing A, Doumbia J, Fokam EB, Gonwouo LM, Greenbaum E, Gvoždík C, Petersen M, Jackson K, James TY, Kusamba C, Larson JG, Mavoungou L-B, Rödel M-O, Zassi-Boulou AG, Penner J (2020) Chytrid Pathogen (Batrachochytrium dendrobatidis) in African amphibians: a continental analysis of occurrences and modeling of its potential distribution. Herpetologica 76:201–215
Zeller KA, Wattles DW, Destefano S (2020) Evaluating methods for identifying large mammal road crossing locations: black bears as a case study. Lands Ecol 35:1799–1808
Acknowledgements
We are grateful to two anonymous Reviewers for their valuable comments. Bd DNA standards were kindly provided by An Martel and Frank Pasmans (Ghent University, Belgium). We thank Marco Maggesi for technical assistance. We thank the staff of Parco Nazionale del Circeo, Parco Nazionale della Maiella and Parco Nazionale del Pollino for assisting in swab collection.
Funding
Open access funding provided by Università degli Studi di Genova within the CRUI-CARE Agreement. This study was partially funded by the Parco Nazionale delle Cinque Terre (Azione di Sistema “Monitoraggio delle specie di habitat umidi-acquatici).
Author information
Authors and Affiliations
Contributions
AC, SS and EG conceived the ideas and designed methodologies; GR and SS collected the data; EG, LD, GA and NBSM performed molecular analyses; AC, SS, and EG analysed the data and drafted the manuscript; LD, GA, NBSM and GR contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interests.
Ethical approval
The Italian Ministry of Environment provided permits for swabbing amphibians.
Additional information
Communicated by Dirk Sven Schmeller.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Costa, A., Dondero, L., Allaria, G. et al. Modelling the amphibian chytrid fungus spread by connectivity analysis: towards a national monitoring network in Italy. Biodivers Conserv 30, 2807–2825 (2021). https://doi.org/10.1007/s10531-021-02224-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-021-02224-5