
Abstract
Using light-level geolocators, eight European Pied Flycatchers (Ficedula hypoleuca) from two breeding sites in Czechia were tracked. We also gathered all available geolocator tracks on 76 individuals from four European populations and compared the timing of annual cycle events and the African non-breeding sites among all populations. Individuals from both Czech breeding sites had overlapping migration events and non-breeding locations. Four individuals resided in the southwestern edge of Mali, two in Burkina Faso, one in Guinea, and the easternmost one in the Ivory Coast. On average, the birds left the Czech breeding grounds on 8 August and took between one to three stopovers during autumn migration. Birds crossed the Sahara on its western edge on average on 13 September. The mean arrival to the African non-breeding grounds was 47.5 days after departure on 2 October (range 10 September to 10 October). One bird showed intra-tropical movement within West Africa when after a 60-day residency it moved approximately 3° westwards. Estimated locations at the African non-breeding grounds overlapped among tracked birds from five European breeding sites. However, statistically, we could detect longitudinal segregation in two clusters. Birds from the British and Finnish breeding populations shared non-breeding grounds and were located in Africa west of the second cluster of the birds from the Czech and Dutch breeding populations. We show considerable population-specific differences in the timing of annual cycle events. Birds from Dutch breeding sites were the first in all three phases—departure from breeding sites, Sahara crossing and arrival to African non-breeding grounds, followed by the British, Czech, and Finnish birds, respectively. All tracked flycatchers so far fill only the western part of the African non-breeding range. For a complete understanding of the migration pattern in the species, we highlight the need for tracking studies from the eastern part of the range.
Zusammenfassung
Zugablauf und afrikanische Überwinterungsgebiete von mit Geolokatoren georteten Trauerschnäppern: ein Vergleich mehrerer Populationen.
Der Langstreckenzug von acht Trauerschnäppern (Ficedula hypoleuca) aus zwei Brutgebieten der Tschechischen Republik wurde mit Hilfe von Geolokatoren verfolgt. Zusätzlich verglichen wir den zeitlichen Ablauf des Jahreszyklus und die Überwinterungsorte aller derzeit verfügbaren getrackten Schnäpper (n = 76) aus vier verschiedenen europäischen Brutpopulationen. Der zeitliche Zugablauf als auch die Überwinterungsorte der Vögel der beiden tschechischen Brutgebiete überschnitten sich. Vier Individuen hielten sich während der Nichtbrutzeit am südwestlichen Rand von Mali, zwei in Burkina Faso, eines in Guinea und der östlichste Vogel in der Elfenbeinküste auf. Die Vögel verliessen ihre tschechischen Brutgebiete im Mittel am 8. August und legten während des Herbstzuges einen bis drei Zwischenstopps ein. Die Sahara wurde im Durchschnitt am 13. September an ihren westlichen Rand überquert. Die Vögel erreichten die afrikanischen Nichtbrutgebiete durchschnittlich nach 47,5 Tagen am 2.Oktober (Spanne: 10.September bis 10.Oktober). Ein Vogel verlagerte nach 60 Tagen seinen Überwinterungsort innerhalb Westafrikas um 3° westwärts (innertropische Bewegung). Die Aufenthaltsorte der Vögel aus fünf verschiedenen europäischen Brutgebieten überschnitten sich im afrikanischen Überwinterungsgebiet weitgehend, wiesen jedoch eine statistisch abgesicherte Clusterformation entlang des Längengrades auf. Vögel der britischen und finnischen Brutpopulationen teilten sich das westlichere Nichtbrutgebiets-Cluster, das zweite östlichere Cluster wurde von Vögeln aus den tschechischen und niederländischen Brutpopulationen gebildet. Wir fanden erhebliche populationsspezifische Unterschiede im Zeitablauf des Jahreszyklus. Vögel aus den niederländischen Brutgebieten waren für alle drei Ereignisse – dem Abflug aus den Brutgebieten, der Überquerung der Sahara und die Ankunft in den afrikanischen Nichtbrutgebieten – die ersten, gefolgt von den britischen, tschechischen und den finnischen Vögeln. Die bisher getrackten Trauerschnäpper überwintern im westlichen Teil des afrikanischen Nichtbrutgebiets der Art. Für ein umfängliches Verständnis der Zugmuster des Trauerschnäppers sind ergänzende Studien aus dem östlichen Teil des Verbreitungsgebiets dringend erforderlich.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Since the beginning of the twenty-first century, advances in bird-tracking devices have unprecedentedly improved our knowledge of the migration ecology of individual small-bodied songbirds. Retrieval of devices from tracked individuals is often labour-intensive, costly and challenging, which results in many studies restricting the fieldwork to single sites and small tracking sample sizes. Spatial replication is, however, critical for a meaningful understanding of the migration ecology of any species. The best approach is to have multi-population studies across a species’ range. In recent years an accumulation in the number of studies has shown the power of multi-population assessments in Great Reed Warblers Acrocephalus arundinaceus (Koleček et al. 2016), Red-backed Shrikes Lanius collurio (Pedersen et al. 2020), Common Rosefinches Carpodacus erythrinus (Lisovski et al. 2021), Northern Wheatears Oenanthe oenanthe (Meier et al. 2022), and non-Passerines (e.g. Finch et al. 2015; Åkesson et al. 2020; Hahn et al. 2020). Such assessments allow for a deeper understanding of migratory corridors and spatiotemporal organization of distant populations across the year.
To add to this list of multi-population assessments, we tracked the European Pied Flycatcher with light-level geolocators at two breeding sites in Czechia. We aim to provide detailed data on the migration patterns of birds from these Czech sites. In addition, we take the opportunity to summarize the current knowledge of the species’ migration patterns based on published tracking results from four other populations available to date (Ouwehand et al. 2016; Ouwehand and Both 2017; Bell et al. 2022). For all birds sampled across European sites we aim to provide an overview of population-specific non-breeding grounds and timings of annual cycle events. We intend to assess whether there is a role of breeding locations in Europe for the clustering of individuals at African residency areas and whether the timing of annual cycle events is linked to the breeding origin of populations, i.e. northern breeding populations migrate later than the southern ones at all stages of the annual cycle (Briedis et al. 2016; Gow et al. 2019).
Methods
Study sites and geolocators in Czechia
We studied the migration of European Pied Flycatchers at two breeding sites in Czechia. The first study site was in Northern Bohemia (50.62 N, 15.83 E); the second site was in North-eastern Moravia (49.95 N, 17.25E). The great circle distance (the shortest distance measured along the surface of a sphere) between the two sites is 170 km. The first site shows stable numbers of breeding birds in a nest box population. At the second site, there is a steady decline of breeding birds and in some plots with nest boxes, the population underwent local extinction during recent decades.
We deployed light-level geolocators (model GDL2 with 7 mm light stalk, Swiss Ornithological Institute) on adult breeding birds across three different field seasons: 2012, 2014, and 2015. In each season, we deployed 20, 36, and 38 geolocators, respectively (Online Supplement 1, Table S2). All birds (42 males and 52 females) were trapped while they were feeding nestlings (at the age of 6–11 days) in nest boxes. We attached the geolocators on the birds’ backs using leg-loop harnesses made of 1 mm thick silicone. Each device, including the harness, weighed approximately 0.6 g (< 5% of the bird’s body mass). In the years following the deployment, we recovered one, three, and five geolocators, respectively. Due to technical difficulties, one logger failed recording, and seven geolocators from 2014 and 2015 contained data only for autumn migration and parts of the wintering period. However, we could identify the African non-breeding residency sites of all seven birds. The remaining geolocator from 2012 stopped recording on 4 May 2013 shortly after the bird had returned to the breeding area. The overall return rate of logger-tagged birds was 11.5% (6/52) for females and 7.1% (3/42) for males. At both sites, regular control of nest box occupancy by flycatchers and overall nest success was performed but due to a lack of manpower and funding no regular recaptures of nesting birds were done. The second site in Moravia had a small population size which also contributed to the lack of a control group. Thus, we lack a formal control group of ringed-only birds for the two sites. The only relevant data on returns of control birds are from a nearby (13 km to the site in Moravia) long-term study site in Dlouhá Loučka (49.83 N 17.21E). At that site, the return rate during 2005–2019 was 12% for females (3/25) and 13.6% for males (3/22; P. Adamík unpubl. data). There was no significant difference between the overall recapture rates of logger-tagged (9/94) and the above-mentioned untagged birds (6/47; χ2 test, χ2 = 0.27, P = 0.604).
Geolocator data analyses
We used the threshold method (Lisovski and Hahn 2012) to determine the sunrise and sunset times of the recorded light data using ‘GeoLocator’ software (Swiss Ornithological Institute) and setting the light level threshold to 1 unit on an arbitrary scale (i.e., minimum detectable ambient light by the given light sensor). All further analyses were conducted using the R-package ‘GeoLight’ v 2.0.0 following the standard procedures (Lisovski and Hahn 2012; Lisovski et al. 2020). Using the ‘loessFilter’ function, in each dataset we first filtered for outlying twilight events that exceed two interquartile ranges (k = 2) of the residuals from a local polynomial regression. We determined the stationary periods with the ‘changeLight’ function by setting the minimal stationary period to 2.5 days and the probability of change to q = 0.9.
When calculating geographic positions for the stationary periods, we excluded 7 days on either side of the equinox times and later filtered all positions north from 80°N and south from 20°S (more than 30° latitude from breeding and median African non-breeding site latitudes). We estimated the geographic positions of the stationary periods using sun-elevation angles derived from Hill–Ekstrom calibration, but when it was not possible, we used in-habitat calibration from the pre-migratory period (Lisovski et al. 2020, Online Supplement 1, Table S3). However, neither of the two methods worked for three of our datasets. For these three datasets, we developed and used a new calibration method—‘equinox calibration’. This calibration method calculates the appropriate sun-elevation angle for the specified number of days around the equinox time when the day and night length at any given geographic location is just about 12 h long. Thus, any deviation from the 12-h day/night length in the geolocators’ recordings reflect the measurement error due to the sensitivity limits of the light sensor or shading by vegetation, weather, etc. The calibration method finds the appropriate sun-elevation angle that would give the desired 12-h day/night length. R-script for this calibration method is provided in Zenodo (Adamík et al. 2023). Due to technical differences in the sensitivity of the geolocators’ light sensors between devices used in different study years, the estimated sun-elevation angles ranged widely between − 2.15 for the newer generation devices, and + 11.39 for the one from the oldest generation device with lower sensitivity light sensor used in 2012. Raw geolocator files from the eight Czech birds are freely available in the Zenodo data repository (Adamík et al. 2023).
We also determined the timing of Sahara crossings for all individuals by manually inspecting the daily light patterns recorded by the geolocators. In short, when crossing large ecological barriers like seas and deserts typical nocturnal migrants, including the European Pied Flycatcher, regularly prolong their flights into the day or may fly non-stop (Adamík et al. 2016; Jiguet et al. 2019). Such behaviour is reflected in the geolocator’s light recordings as lengthy periods of uninterrupted maximal light intensities when the light sensor is exposed to the sun as the bird flies (full light pattern, hereafter FLP or Sahara crossing). Due to difficulties in reliable estimates of stopover locations close to equinox periods, data on stopovers are presented only as timings and median longitudinal estimates.
We estimated migration speed as migration distance divided by duration (including stopovers). Distances between the breeding and African non-breeding sites were estimated as a great circle distance. Migration duration is the time (in days) between departure from the breeding site and arrival to the African non-breeding site (duration.migration).
Multipopulation assessment
We collated published data on individually tracked European Pied Flycatchers from European breeding sites. To date, there are available data on geolocator-tracked birds from the UK, the Netherlands, Finland, and Norway (Ouwehand et al. 2016; Ouwehand and Both 2017; Bell et al. 2022). From these studies, we extracted data on departure from breeding sites (variable names in parentheses: autumn.departure), timing of Sahara crossing (inferred from light anomalies, FLP), arrivals to African non-breeding grounds (winter.arrival), median nonbreeding location estimates (wint.longitude, wint.latitude) and egg-laying dates (laying.date). We took the dates of the Sahara crossing for the four Finnish birds from Adamík et al. (2016). The full-collated dataset for 76 individuals is available as an Online Supplement 2, Table S1. All variables related to dates are expressed as days of the year.
To assess whether the five European populations differ in African non-breeding site locations or duration of migration, we ran three linear models (LM) with a country as an explanatory variable (five countries) and non-breeding site longitudinal (winter.longitude) or latitudinal location (winter.latitude) and duration of migration (in days) as response variables. In further three LMs, which always had a single predictor, we explored whether non-breeding longitudes (response variable) can be explained by egg-laying dates, departures from breeding sites, and arrivals to Africa.
In further analyses, we used linear mixed-effects models (LMM) to assess whether latitudinal or longitudinal location estimates in Africa (response variables winter.longitude or winter.latitude) are associated with breeding site longitudes or latitudes (fixed predictors: breeding.longitude, breeding.latitude) while accounting for the fact that multiple individuals originate from the same study site. For this reason, we entered the breeding population (country) as a random effect. For evaluating the strength of relationships between the four consecutive phases of the annual cycle (egg-laying date, departure from breeding site, Sahara crossing, arrival to African non-breeding sites) we fitted LMMs which had always a single fixed predictor and country as a random effect. For clarity, the model syntax is provided with the test statistics in the results. For model fitting we used the R package lme4 (Bates et al. 2015). For model diagnostics, we used the R package performance (Lüdecke et al. 2021). The models were run for the full dataset of 76 individuals, however, the sample size was 66 for Sahara crossing, 74 for arrival to non-breeding grounds, 47 for location estimates of nonbreeding grounds and 41 for egg-laying dates.
Results
Migration of birds from Czech breeding grounds
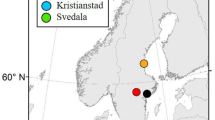
On average, Czech flycatchers departed from their breeding grounds on 8 August (range 24 July to 22 August, Table 1). All flycatchers headed SW towards the Iberian Peninsula (Fig. 1 and Online Supplement 1, Fig. S1). We detected between one to three stopover sites per bird. Stopovers before the Sahara crossing lasted on average 9.7 days (n = 13; range 3–24.5 days) and were located around 4.7°W (range 2.3°E to 9.5°W). Stopovers after the Sahara crossing were slightly shorter, on average 6.8 days (n = 6; range 3.5–11.5 days; Online Supplement 1, Table S4) and were further west at around 12.2°W (range 9.6–14.1° W). On average, birds crossed the Sahara on its western edge on 13 September (range 30 August–30 September). Mean arrival to the African non-breeding grounds was on 2 October (range 10 September to 10 October). One bird showed intra-tropical movement when it arrived at its first African non-breeding site on 11 September where it stayed for 60 days, after which it moved about 3° westwards to its final residency site. Autumn migration lasted on average 47.5 days (range 34–62 days) including stopovers. The African non-breeding residency sites overlapped for the two tracked Czech populations and for both sexes (t-test on longitudes: t = − 0.09, P = 0.928, df = 6; t-test on latitudes: t = 0.53, P = 0.612, df = 6; Fig. 1). Most birds were clustered around the south-western edge of Mali (four individuals), two in Burkina Faso, one in Guinea, and the easternmost one in the Ivory Coast. As birds from both Czech breeding sites showed considerable overlap in both non-breeding locations and their migration phenology, we pooled the data on them for the pan-European comparison of populations.

Locations of African non-breeding grounds (blue crosses) for eight geolocator-tracked European Pied Flycatchers from two breeding sites in Czechia. Years of tracking are depicted in different colours. The map also shows stopover timing and location (in red) estimates for one individual, 15 HW. Location estimates are median values ± interquartile range. Background map shows breeding (green) and non-breeding (blue) distributions of the species (BirdLife International and NatureServe 2013)
For the one bird with data available up until spring, the departure from the non-breeding site was after 204 days of residency on 20 April. The bird initiated a crossing of the Sahara on 23 April and made a 12-day stopover around 6.7° E after the desert crossing.
Multipopulation assessment of African non-breeding grounds
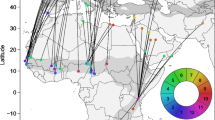
Birds from all five European breeding populations overlapped at their West African non-breeding grounds (Fig. 2). However, there is a statistically significant effect of the breeding origin of the population on longitudinal estimates of non-breeding locations (LM: winter.longitude ~ country, F4,69 = 28.4, P < 0.001, R2 = 0.62). The single Norwegian bird stayed in the most westward location. The four remaining populations created two clusters. Mean location estimates of UK and Finnish birds overlapped around 10.5 and 10.7° W, respectively, and were west of the Czech and Dutch populations. The Czech and Dutch populations overlapped at estimated mean longitudes of 7.14 and 7.32° W. Latitudinally the non-breeding locations largely overlapped across all populations (LM: winter.latitude ~ country, F4,42 = 0.66, P = 0.622, R2 = 0.05). Non-breeding longitude tended to be associated with breeding latitude (LMM: winter.longitude ~ breeding.latitude + (1 | country), b = − 0.37 ± 0.18, t = − 2.0, P = 0.084, marginal R2 = 0.15, conditional R2 = 0.64, n = 74, Online Supplement 1, Fig. S2) but no relationship was found with breeding longitude (LMM: winter.longitude ~ breeding.longitude + (1 | country), b = 0.00 ± 0.11, t = 0.0, P = 0.977, marginal R2 = 0.00, conditional R2 = 0.73, n = 74, Online Supplement 1, Fig. S2).

Location estimates (medians) of African non-breeding grounds for 47 European Pied Flycatcher individuals tracked with geolocators across five European breeding sites. For better visualization, coloured polygons represent spline-smoothed non-breeding regions of each population based on geolocator records. A convex hull of individual non-breeding sites is smoothed (per population) according to their distance from the population mean non-breeding coordinates giving 30% margin of the difference between individual points on the convex hull and population mean non-breeding coordinates. For the single Norwegian bird, a circle with a 1° radius around the median location is drawn. The dots in the inset show locations where the birds were tagged at the breeding sites
Migration timing of European breeding populations
Most birds (54 out of 76) left the breeding sites by the end of the first week of August (range July 15–August 28, Fig. 3). Birds from the Dutch and UK breeding populations were similar in departure timing, with mean departure dates of 1 August and 4 August, respectively. The Czech birds left around 8 August and the Finnish birds departed on average 16 days later. The single Norwegian bird left the breeding site on 16 August. A similar order was found for the Sahara crossing timing, but here the populations differed in the interval between departure from the breeding sites and the Sahara crossing. The shortest interval was in Dutch birds (18 days) and the longest was in the Finnish and Czech birds (36–37 days). Population-specific arrivals to African non-breeding grounds were again in the same order as breeding site departures. Interestingly, the Finish birds had a very short interval (8 days) between Sahara crossing and arrival to non-breeding locations (mean intervals in other populations were in a range of 17–21 days). The Norwegian bird arrived late to the African non-breeding grounds (14 October vs mean for all birds 16 September).

Boxplots of timing of three annual cycle events across four European Pied Flycatcher breeding populations. Populations are ordered by breeding latitude. Data for the single bird from Norway are not visualized. X axis shows day of the year (e.g. 200 = July 19)
Autumn migration ranged from 17 to 85 days, and on average, it took 41.3 days to reach the African non-breeding sites. There was a significant effect of breeding population on the duration of migration (LM: duration.migration ~ country, F4,69 = 4.71, P = 0.002, R2 = 0.21) but this was likely due to the unusually short migration time of the Dutch birds (mean 35.3 days) while birds from other populations had similar durations (except for the one Norwegian bird with migration of 59 days). Birds that left their breeding sites late tended to have shorter migration durations (LMM: duration.migration ~ autumn.departure + (1 | country), b = − 3.31 ± 0.15, t = − 2.2, P = 0.033, marginal R2 = 0.05, conditional R2 = 0.36, n = 74).
Consecutive phases of the annual cycle were strongly correlated, even when controlling for substantial variation within the breeding populations (Fig. 4). The strongest relationship was found between phases that were closer together (i.e. timing of the Sahara crossing and arrival to African non-breeding grounds (LMM: winter.arrival ~ FLP + (1 | country), b = 0.79 ± 0.12, t = 6.5, P < 0.001, marginal R2 = 0.52, conditional R2 = 0.48, n = 64)). Similarly, a strong relationship was observed between the departure from the breeding sites and the Sahara crossing (LMM: FLP ~ autumn.departure + (1 | country), b = 0.59 ± 0.09, t = 6.9, P < 0.001, marginal R2 = 0.21, conditional R2 = 0.78, n = 66). A weaker relationship was found between breeding site departure and arrival to African non-breeding grounds (LMM: winter.arrival ~ autumn.departure + (1 | country), b = 0.51 ± 0.17, t = 3.1, P = 0.003, marginal R2 = 0.10, conditional R2 = 0.40, n = 74). There was no relationship between egg-laying dates and departures (LMM: autumn.departure ~ laying.date + (1 | country), b = 0.18 ± 0.17, t = 1.0, P = 0.303, marginal R2 = 0.02, conditional R2 = 0.53, n = 41, Online Supplement 1, Fig. S3).

Relationships between annual cycle events in the European Pied Flycatcher. The timing of events is expressed as days of the year
There was a weak tendency for birds residing at more westerly non-breeding grounds to breed later (LM: wint.longitude ~ laying.date, b = − 0.07 ± 0.04, F1,39 = 3.82, P = 0.058, R2 = 0.09; Fig. 5). Longitudinal location in Africa was not associated with individual departure from breeding sites (LM: wint.longitude ~ autumn.departure, b = − 0.03 ± 0.03, F1,72 = 1.09, P = 0.300, R2 = 0.01) but it had an effect on arrival, with birds residing further east arriving earlier (LM: wint.longitude ~ winter.arrival, b = − 0.04 ± 0.01, F1,72 = 8.81, P = 0.004, R2 = 0.11; Fig. 5).

Relationships between non-breeding site longitudinal estimates and laying dates, arrivals to non-breeding grounds and departures from breeding sites
Discussion
In this study, we brought new European Pied Flycatcher autumn migration data from two subpopulations in a central European region. We did not find any substantial differences in migration schedules and locations at the African non-breeding grounds for these birds. But we should be cautious as the sample size was small and we did not have access to full-year tracking data. Interestingly, from atlas mapping we see that the two subpopulations show regionally contrasting population trajectories, the one in Bohemia being stable and the second in Moravia declining (Šťastný et al. 2021). It would be valuable to know where and at which time of year the main drivers of population dynamics act in these two regional populations. For Dutch birds, there is evidence for mechanisms at breeding sites (Both et al. 2006), while across the UK the trends in populations are driven by changes in survival and immigration which probably act outside the breeding season (Nater et al. 2023). The fact that we found overlaps in non-breeding locations of our sample of birds does not necessarily mean that they cannot differ in habitat use at a finer scale, which is below the resolution of geolocation by light.
We were able to detect between one to three stopovers during the autumn migration, usually one or two stops before the Sahara crossing and one after it. Stops before the Sahara crossing were slightly longer, nearly 10 days, while those after it lasted on average almost 7 days. Pied Flycatchers tracked from the southwest UK usually have two stops during the autumn migration, one before and one after the Sahara crossing (Bell et al. 2022). Interestingly, for the UK-tracked birds’ stopover durations were slightly longer after the barrier crossing. Longitudinal estimates of stopovers after the Sahara crossing were in a similar range between the two populations (CZ: 9.6–14.1° W vs UK: 9.8–16.2° W), albeit birds from the Czech breeding population stopped on average 1.1° further east of the UK birds. The increased number of stopovers detected in our sample of birds are likely a consequence of the longer migration distances faced by the Central European birds. This fits with the findings of Fourcade et al. (2022) who estimated stopover durations at a fuelling site in south-western France near the Atlantic coast. At their site the body masses were lower than those from study sites located further South on the Iberian Peninsula (Bibby and Green 1980; Goffin et al. 2020), indicating that one additional stopover was needed for fuelling before the barrier crossing. Only a few individuals at the French stopover site had sufficient fuel loads to be able to cross the Sahara without additional refuelling (Fourcade et al. 2022).
Multi-population assessment
By collating available geolocator tracks from 76 individuals from five populations in Europe, we provide a comprehensive overview of the locations of African non-breeding residency sites and the timing of autumn migration for individual European Pied Flycatchers. Except for the single Norwegian individual, birds from the other four European breeding populations showed considerable overlap in location estimates at the African non-breeding grounds. Statistically, we could detect longitudinal segregation in two clusters: birds from the UK and Finnish populations overlapped and were west of the second cluster of birds from the Czech and Dutch populations. There was no evidence of any latitudinal segregation of the populations. However, one has to be careful with the interpretation of latitudinal estimates inferred from light-level geolocators. By default, they have considerable uncertainty, while there are also issues with different calibration approaches, and whether birds from different populations use similar habitats (Lisovski et al. 2012; Lisovski et al. 2018). We failed to find clear support for the role of European breeding locations in the clustering of subsequent African residency areas. Only breeding latitude was very weakly associated with non-breeding site longitude estimates. However, the effect was weaker than in the first study on European Pied Flycatcher tracking by Ouwehand et al. (2016). This could be purely a consequence of the sampling effect. Finch et al. (2017) found strong support for a positive link between breeding and non-breeding longitudes but no link between latitudes in several populations of European Rollers (Coracias garrulus). In Common Swifts (Apus apus) tracked across several European populations, breeding latitudes were positively correlated with non-breeding latitudes, clear evidence for a chain migration pattern (Åkesson et al. 2020). The fact that we failed to find strong support for links in the European Pied Flycatcher might simply reflect the scale of the contemporary study. This is a critical issue in any study on migratory connectivity. In an ideal situation, birds would have to be sampled across the entire species´ breeding range.
Migration timing was considerably different among populations. Dutch birds were first in all three phases—departure from breeding sites, Sahara crossing and arrival to African non-breeding grounds—followed by the UK, Czech and Finnish birds, respectively. Interestingly, of all populations, the Finnish birds had the shortest interval of only eight days between the Sahara crossing and arrival to non-breeding grounds. This could indicate that they undertook considerable refuelling prior to the barrier crossing, performing a long endurance flight with arrival close to the African residency sites. The birds could skip refuelling after desert crossing or their stops were very short, below the resolution set for stationary periods in the GeoLight package (given the data quality for detecting short stopovers). Autumn migration ranged from 17 to 85 days and it was similar for Czech, UK and Finnish birds. In contrast, the Dutch birds had the shortest migration of only about 35 days. We also found a negative relationship between breeding site departures and duration of migration, i.e. the later a bird departed the shorter time it was en route. This is a similar pattern to Collared Flycatcher (Ficedula albicollis) in which later departing individuals migrated at faster speeds towards African residency sites (Briedis et al. 2018a). Very likely, late individuals are trying to catch up with the early ones. Whether such behaviour is innate or the birds adjust it according to seasonal changes in available food resources is unknown. Another interesting finding was that birds residing further west in Africa arrived there later. The effect was much stronger than in the previous study by Ouwehand et al. (2016). In contrast to Ouwehand et al. (2016) we did not confirm the relationship between breeding site departures and non-breeding longitudes. The difference between these two studies is likely attributable to the sampling effect.
Interestingly, we did not find a significant effect of the timing of breeding on the subsequent phases of the annual cycle, even though the phases were positively correlated with each other, a pattern regularly found in other songbirds (e.g. Mitchell et al. 2012; van Wijk et al. 2017; Gow et al. 2019). As we clearly see large differences in the timings of events in the studied populations, we would expect a strong effect of seasonality. However, egg-laying dates were available for only 41 of the 76 tracked individuals. A similar lack of effect on the timing of breeding for subsequent annual cycle events was found in the Collared Flycatcher (Briedis et al. 2018b). Thus, there must be other factors than just egg-laying that explain the timing of subsequent events. No doubt, there must be a significant role of the photoperiod at the breeding sites that sets the pace for the timing of the circannual rhythms (Gwinner 1996). Briedis et al. (2020) found a strong effect of seasonality shaping the timing of avian annual cycles. Thus, further exploration with a larger number of study sites or experimental translocations would be desirable to explore the role of, for example, site-specific phenology and photoperiod in explaining the variability in the timing of annual cycle events among populations.
In our study, the overall return rate of tagged birds was 9.6%, with males having slightly lower return rates. Unfortunately, for various reasons, we did not have control groups of ringed-only birds at both sites. Our only available data are from a nearby study site with an overall return rate of 12.8%. We know that the true return rate on logger-tagged birds must have been higher, but our study coincided with two seasons of cold and rainy weather at the time of nestlings. In addition, we experienced very high nest mortality due to dormice and marten predation (Adamík and Král 2008). As a result, we often could not catch the adult breeders and control them for geolocators. No matter of this, we have to admit that for these two particular sites we cannot be sure about the tagging effect on return rates. The available published studies report no general tagging effect on return rates (Brlík et al. 2020), and Bell et al. (2017) report no negative effects in British flycatchers. In the Dutch flycatcher population, there was no overall tagging effect, but the type of harness did affect return rates (Ouwehand and Both 2017). Return rates of logger-tracked birds in other populations used in our comparative study were in the range of 4–42% and of the control group in a range of 10–56% (Ouwehand et al. 2016). We think that in our case the loggers did not affect between-population differences in migratory behaviour and the differences found across populations are not related to tags per se. However, this is beyond the scope of our study, and we still know very little for how tagging impacts on behaviour of migratory birds across the years.
With our multi-population assessment, we try to fill a gap by comparing the timing of migration of Western European populations of European Pied Flycatcher. In addition, we show that there is considerable mixing at the African non-breeding grounds of birds from various breeding origins in the western Palaearctic. Across species, various breeding populations seem to frequently mix at African non-breeding sites (Finch et al. 2017). This might have interesting consequences for population dynamics at the breeding grounds as depending on the scale of factors operating at the non-breeding sites they might have (de)synchronising effects. However, with the available tracking studies we could cover only a small part of the populations from the extensive breeding range of the species. This comparative study only covered western and central Europe and part of Fennoscandia. The species´ breeding range extends far into the East (up to 90°E) and there is no tracking study from the European part of Russia and further east to western Siberia. The few ringing recoveries available from the eastern part of the range suggest the general heading during migration is towards the Iberian Peninsula (Spina et al. 2022). Thus, in autumn even birds from the Asian breeding populations pass via Iberia and the western fringe of the Sahara (Chernetsov et al. 2008). From a conservation perspective, this means that the entire population passes through a particular region. Any changes in such bottleneck, that could affect stopover behaviour, for example via fuelling rates, might represent a critical point for different populations (Runge et al. 2014). While the African non-breeding range stretches all the way east to the Central African Republic and the northeastern part of DR Congo, there is not a single recovery connecting these areas to the breeding grounds (Spina et al. 2022). Similarly, all tracked flycatchers so far fill only the western part of the African non-breeding range. We suggest that non-breeding populations further east in central Africa originate from breeding sites at the eastern part of the breeding range. As such, further studies from the eastern part of the breeding range are needed, not only for European Pied Flycatchers but also for a wide range of other species. Furthermore, to get a thorough picture of the patterns of migratory connectivity, we need tracking studies from the African non-breeding sites to find the breeding origins of wintering birds (cf. Blackburn et al. 2017). This may be particularly valuable for the European Pied Flycatcher—a model species for climate change research.
Data availability
The dataset used in the current study is available in supplementary online materials. Raw geolocator data from the eight Czech birds are available at Zenodo: https://doi.org/10.5281/zenodo.7434786.
References
Adamík P, Král M (2008) Nest losses of cavity nesting birds caused by dormice (Gliridae, Rodentia). Acta Theriol 53:185–192
Adamík P et al (2016) Barrier crossing in small avian migrants: individual tracking reveals prolonged nocturnal flights into the day as a common migratory strategy. Sci Rep 6:21560. https://doi.org/10.1038/srep21560
Adamík P, Bureš S, Oatley G, Hahn S, Briedis M (2023) Timing of migration and African non-breeding grounds of geolocator-tracked Pied Flycatchers: a multi-population assessment (1.0). Zenodo. https://doi.org/10.5281/zenodo.7434786
Åkesson S et al (2020) Evolution of chain migration in an aerial insectivorous bird, the common swift Apus apus. Evolution 74:2377–2391
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Bell SC, El Harouchi M, Hewson CM, Burgess MD (2017) No short- or long-term effects of geolocator attachment detected in Pied Flycatchers Ficedula hypoleuca. Ibis 159:734–743
Bell F, Bearhop S, Briedis M, El Harouchi M, Bell SC, Castelo J, Burgess M (2022) Geolocators reveal variation and sex-specific differences in the migratory strategies of a long-distance migrant. Ibis 164:451–467
Bibby CJ, Green RE (1980) Foraging behaviour of migrant Pied Flycatchers, Ficedula hypoleuca, on temporary territories. J Anim Ecol 49:507–521
BirdLife International and NatureServe (2013) Bird species distribution maps of the world. BirdLife International, Cambridge, UK and NatureServe, Arlington
Blackburn E et al (2017) Low and annually variable migratory connectivity in a long-distance migrant: Whinchats Saxicola rubetra may show a bet-hedging strategy. Ibis 159:902–918
Both C et al (2006) Climate change and population declines in a long-distance migratory bird. Nature 441:81–83
Briedis M, Hahn S, Gustafsson L, Henshaw I, Träff J, Král M, Adamík P (2016) Breeding latitude leads to different temporal but not spatial organization of the annual cycle in a long-distance migrant. J Avian Biol 47:743–748
Briedis M, Hahn S, Krist M, Adamík P (2018a) Finish with a sprint: evidence for time-selected last leg of migration in a long-distance migratory songbird. Ecol Evol 8:6899–6908
Briedis M, Krist M, Král M, Voigt CC, Adamík P (2018b) Linking events throughout the annual cycle in a migratory bird—non-breeding period buffers accumulation of carry-over effects. Behav Ecol Sociobiol 72:1–12
Briedis M et al (2020) Broad-scale patterns of the Afro-Palaearctic landbird migration. Glob Ecol Biogeogr 29:722–735
Brlík V, Koleček J, Burgess M, Hahn S et al (2020) Weak effects of geolocators on small birds: a meta-analysis controlled for phylogeny and publication bias. J Anim Ecol 89:207–220
Chernetsov N, Kishkinev D, Gashkov S, Kosarev V, Bolshakov CV (2008) Migratory programme of juvenile pied flycatchers, Ficedula hypoleuca, from Siberia implies a detour around Central Asia. Anim Behav 75:539–545
Finch T et al (2015) A pan-European, multipopulation assessment of migratory connectivity in a near-threatened migrant bird. Divers Distrib 21:1051–1062
Finch T, Butler SJ, Franco AM, Cresswell W (2017) Low migratory connectivity is common in long-distance migrant birds. J Anim Ecol 86:662–673
Fourcade JM, Fontanilles P, Demongin L (2022) Fuel management, stopover duration and potential flight range of pied flycatcher Ficedula hypoleuca staying in South-West France during autumn migration. J Ornithol 163:61–70
Goffin B, Felgueiras M, Hof AR (2020) Increased stopover duration and low body condition of the Pied Flycatcher (Ficedula hypoleuca) at an autumn stopover site. Animals 10:1–13
Gow EA et al (2019) A range-wide domino effect and resetting of the annual cycle in a migratory songbird. Proc R Soc B 286(1894):20181916
Gwinner E (1996) Circadian and circannual programmes in avian migration. J Exp Biol 199:39–48
Hahn S et al (2020) Range-wide migration corridors and non-breeding areas of a northward expanding Afro-Palaearctic migrant, the European Bee-eater Merops apiaster. Ibis 162:345–355
Jiguet F et al (2019) Desert crossing strategies of migrant songbirds vary between and within species. Sci Rep 9:1–12
Koleček J et al (2016) Cross-continental migratory connectivity and spatiotemporal migratory patterns in the great reed warbler. J Avian Biol 47:756–767
Lisovski S, Hahn S (2012) GeoLight—processing and analysing lightbased geolocator data in R. Methods Ecol Evol 3:1055–1059
Lisovski S et al (2012) Geolocation by light: accuracy and precision affected by environmental factors. Methods Ecol Evol 3:603–612
Lisovski S et al (2018) Inherent limits of light-level geolocation may lead to over-interpretation. Curr Biol 28:R99–R100
Lisovski S et al (2020) Light-level geolocator analyses: a user’s guide. J Anim Ecol 89:221–236
Lisovski S et al (2021) The Indo-European flyway: Opportunities and constraints reflected by Common rosefinches breeding across Europe. J Biogeog 48:1255–1266
Lüdecke D et al (2021) performance: an R Package for assessment, comparison and testing of statistical models. J Open Source Softw 6(60):3139
Meier CM, Rime Y, Lisovski S, Buchmann M, Liechti F (2022) Locally adapted migration strategies? Comparing routes and timing of northern wheatears from alpine and lowland European populations. J Avian Biol 2022:e02932
Mitchell GW, Newman AEM, Wikelski M, Ryan Norris D (2012) Timing of breeding carries over to influence migratory departure in a songbird: an automated radiotracking study. J Anim Ecol 81:1024–1033
Nater CR, Burgess MD, Coffey P, Harris B, Lander F, Price D, Reed M, Robinson RA (2023) Spatial consistency in drivers of population dynamics of a declining migratory bird. J Anim Ecol 92:97–111
Ouwehand J, Both C (2017) African departure rather than migration speed determines variation in spring arrival in pied flycatchers. J Anim Ecol 86:88–97
Ouwehand J, Ahola MP, Ausems ANMA, Bridge ES, Burgess M, Hahn SA, Hewson CM, Klaassen RHG, Laaksonen T, Lampe HM, Velmala W, Both C (2016) Light-level geolocators reveal migratory connectivity in European populations of pied flycatchers Ficedula hypoleuca. J Avian Biol 47:69–83
Pedersen L et al (2020) Remarkably similar migration patterns between different red-backed shrike populations suggest that migration rather than breeding area phenology determines the annual cycle. J Avian Biol 51:e02475
Runge CA, Martin TG, Possingham HP, Willis SG, Fuller RA (2014) Conserving mobile species. Front Ecol Environ 12:395–402
Spina F, Baillie, SR, Bairlein F, Fiedler W, Thorup K (2022) The Eurasian African Bird Migration Atlas. https://migrationatlas.org. EURING/CMS
Šťastný K, Bejček V, Mikuláš I, Telenský T (2021) Atlas hnízdního rozšíření ptáků v České republice 2014–2017. Aventinum, Praha
van Wijk RE, Schaub M, Bauer S (2017) Dependencies in the timing of activities weaken over the annual cycle in a long-distance migratory bird. Behav Ecol Sociobiol 71:73. https://doi.org/10.1007/s00265-017-2305-5
Acknowledgements
We thank M. Dusík, L. Harmáčková and J. Fuchs for their help with fieldwork and the two reviewers for their constructive comments on the manuscript. M. Dusík kindly provided access to some of the nest box sites.
Funding
Open access publishing supported by the National Technical Library in Prague. PA received support from the Czech Science Foundation (project 20-00648S) while writing the manuscript.
Author information
Authors and Affiliations
Contributions
The study was conceptualized by PA with input from SH, MB and SB. SB provided resources and with GO did the field work. PA performed statistical analyses, while MB analyzed geolocator data. PA wrote the original draft of the manuscript, and MB wrote part of the methods on geolocation. SH and MB critically edited and revised the manuscript. All authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
None.
Ethics approval
Deployment of geolocators was granted by a ringing licence of the National Museum issued to S. Bureš.
Additional information
Communicated by N. Chernetsov.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Adamík, P., Bureš, S., Hahn, S. et al. Timing of migration and African non-breeding grounds of geolocator-tracked European Pied Flycatchers: a multi-population assessment. J Ornithol 164, 875–886 (2023). https://doi.org/10.1007/s10336-023-02081-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-023-02081-9