
Abstract
With the rapid development of synthetic biology and metabolic engineering technologies, yeast has been generally considered as promising hosts for the bioproduction of secondary metabolites. Sterols are essential components of cell membrane, and are the precursors for the biosynthesis of steroid hormones, signaling molecules, and defense molecules in the higher eukaryotes, which are of pharmaceutical and agricultural significance. In this mini-review, we summarize the recent engineering efforts of using yeast to synthesize various steroids, and discuss the structural diversity that the current steroid-producing yeast can achieve, the challenge and the potential of using yeast as the bioproduction platform of various steroids from higher eukaryotes.
Similar content being viewed by others
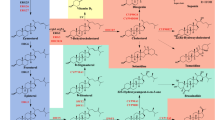


Introduction
Sterols are essential membrane components that regulate membrane fluidity and permeability in eukaryotic organisms [1]. Animals generally synthesize cholesterol, fungi produce ergosterol, and plants utilize an array of sterols with β-sitosterol and campesterol as the most common ones [1]. Despite the essential roles in membrane modulation, sterols also exhibit important bioactivities. For example, due to the structural similarity to cholesterol, phytosterols can reduce cholesterol absorption in digestive system and reduce the risk of heart disease [2, 3]. In higher eukaryotes, sterols are also precursors to a wide array of specialized metabolites with broad structural diversity and biological activities. In insects, cholesterol is converted into insect steroid hormones, such as ecdysteroids that can be potentially utilized as anabolic steroids [4]. In human, cholesterol is the precursor to an array of steroid hormones such as progesterone and vitamin D [5, 6]. In plants, campesterol is the main precursor of the ubiquitous phytohormone brassinosteroids [7]; cholesterol, although exhibits very low abundance in plants, is the precursor to a broad spectrum of phytochemicals, such as the medicinally important steroidal alkaloids and steroidal saponins [8].
Intriguingly, some phytosteroids exhibit promising antiviral activities via different mechanisms, which make them intriguing lead structures for novel antiviral drug discovery and development [9,10,11,12]. Plants also produce ecdysteroid analogs, phytoecdysteroids, as natural defense mechanism against phytophagous insects [4]. Moreover, steroids are an important pharmaceutical medication, with the global steroid market projected to reach $17 billion in 2025 [13]. Steroid drugs generally fall into two categories: anabolic and corticosteroids. Anabolic steroids are involved in the treatment of muscle loss or late puberty [14]; while corticosteroids can be used as anti-inflammatory drugs or allergic treatment [15]. During the recent outbreak of COVID-19, methylprednisolone (median price $16.03/80 mg [16]), a corticosteroid, was reported to have positive effects on a faster improvement of the clinical symptoms caused by COVID-19 [17]. More recently, another corticosteroid, dexamethasone (median price $12.30/4 mg from www.GoodRx.com), has been found to able to reduce the death by roughly one-third of the patients on ventilators [18].
However, most of these specialized steroids are not natively produced at a high level, due to the nature of their biological functions. Similar to other specialized metabolites, the structural complexity also makes the chemical synthesis of these compounds challenging and expensive [19]. Recent development of synthetic biology provides an alternative sourcing strategy of these molecules through biotechnological production in the fast-growing, fermentable microorganisms, such as yeast and Escherichia coli. The structural and biosynthetic similarity of ergosterol in yeast to the other eukaryotic sterols highlights the potential of using yeast as the bioproduction host for sterols and derivatives [20]. The previous metabolic engineering efforts in establishing steroid production in yeast have been very well reviewed in Wriessnegger’s 2013 review [20]. In this review, we will mainly discuss the efforts in engineered steroid bioproduction in yeast since 2013 and focus on the metabolic engineering strategies, and the challenges and potential biosynthetic capacities of yeast-based steroid bioproduction.
Ergosterol biosynthesis in yeast
Ergosterol, similar to cholesterol in animals and phytosterols in plants, is the most abundant sterol in fungi and plays an essential role in maintaining physicochemical properties of plasma membrane and involves in protein sorting and protein receptor regulation [21]. The biosynthesis of ergosterol exists in almost all yeast species and is well characterized, especially in the model organism Saccharomyces cerevisiae. The biosynthesis of ergosterol in S. cerevisiae can be divided into three modules [22]: (1) biosynthesis of mevalonate (MVA) from acetyl-CoA, (2) conversion of MVA into farnesyl pyrophosphate (FPP) and 3) ergosterol biosynthesis from FPP. The third module can be further divided into (3-1) lanosterol biosynthesis from FPP, (3-2) zymosterol biosynthesis from lanosterol and (3-3) ergosterol biosynthesis from zymosterol. Ergosterol, cholesterol, and the major phytosterols, campesterol and β-sitosterol, are different in three aspects: saturation of C7–C8 bond in B-ring, saturation of C22–C23 bond, and the alkane group at C24 in the side chain (Fig. 1).

Ergosterol synthesis and sterols from mammals and plants. The endogenous ergosterol biosynthesis in yeast is highlighted in yellow. The three different sheds of yellow represented three modules of ergosterol synthesis. Major phytosterols that have been synthesized or possibly synthesized in yeast are highlighted in green, and cholesterol is highlighted in blue. The characteristic structural features of sterols from different eukaryotic organisms are highlighted in pink. The enzymes marked in red represent the ones that require NADPH/NADP+, and the ones that require ATP are marked in blue
Sterol biosynthesis is highly conserved in eukaryotes [23]. The synthesis of ergosterol, cholesterol, and phytosterols diverges from lanosterol, and shares a number of downstream intermediates. For example, zymosterol and episterol in the ergosterol synthesis (submodule 3-3) can also be found in animals and plants, respectively [8, 24]. The convergency of the eukaryotic sterol structures and biosynthesis makes it possible to redirect the biosynthesis of ergosterol to cholesterol and phytosterols in yeast. The late-stage biosynthesis of ergosterol from zymosterol (module 3-3) involves five ERG enzymes, ERG2-6. ERG6 is the ∆24-sterol C-methyltransferase, converting zymosterol into fecosterol [25]. Fecosterol is then converted into episterol by ERG2 through the isomerization of C8–C9 double bond to C7–C8 position [25]. ERG3 is the C5 desaturase that converts episterol into ergosta-5,7,24(28)-trienol [25]. ERG5 is the C-22 sterol desaturase that catalyzes the conversion from ergosta-5,7,24(28)-trienol to Ergosta-5,7,22,24(28)-tetraenol [25]. ERG4 is the ∆24-sterol reductase that reduces C24–C28 double bond to synthesize ergosterol [25]. The corresponding genes (erg2-6) are all identified as nonessential genes to yeast viability [26]. Previous investigations on the effects of erg gene deletions suggest that ERG2-6 enzymes can accept a broad range of sterol structures, which indicates the broad substrate promiscuities of ERG2-6 and that these enzymes may not follow a specific order to afford the synthesis of ergosterol [27]. For example, the disruption of erg6 accumulates not only zymosterol, but also cholesta-7,24-dienol, cholesta-5,7,24-trienol and cholesta-5,7,22,24-tetraenol, which indicates that ERG2, ERG3 and ERG5 can keep functioning sequentially on zymosterol in the absence of ERG6 [28]. Different combinations of erg gene deletions result in various sterol compositions. More detailed sterol composition can refer to the review published by Johnston et al. very recently [27]. The versatility of yeast ergosterol biosynthesis implicates the potential of yeast to synthesize diversified sterol structures through ergosterol biosynthesis engineering and heterologous pathway reconstitution.
To utilize yeast as the steroid bioproduction platform, ergosterol biosynthesis needs to be disrupted and thus, it is necessary to understand the effects of erg gene deletions on yeast. Generally, among single deletion of erg2-6, Δerg6 and Δerg4 exhibit the most evident effects on the response of yeast to antifungal drugs (e.g., hygromycin B, tetramethylammonium), cations and heat [29,30,31]. Inactivation of erg6 or erg4 also results in a crucial deficiency on plasma membrane integrity, leading to strong deviation from the wild type in respect of membrane potential and tolerance to LiCl [30, 31]. On the other hand, inactivation of erg5 from S. cerevisiae does not change the membrane potential and the tolerance to cations (such as Li+, K+ and Na+) and hygromycin B as much [30]. The phenotypes of other erg mutants have been carefully summarized and discussed in the recent reviews by Johnston et al. and Sokolov et al. [21, 27].
Steroid bioproduction in yeast through biotransformation
Biotransformation of steroid drugs usually requires hydroxylation and dehydrogenation at different positions of the sterane skeleton, and is normally catalyzed by cytochrome P450s and hydroxysteroid dehydrogenases (HSDs) [32]. Steroid hydroxylation usually involves 7α-, 11α-, 11β-, 14α-hydroxylases, while dehydrogenation is generally catalyzed by 3β-, 5α-HSDs. Biotransformation of androstenedione (AD) to testosterone (TS) by S. cerevisiae was discovered as early as in 1937 [33], with the same function discovered in Schizosaccharomyces pombe later [34]. The biotransformation of steroid drugs by microbial strains was intensively studied in the 1990s, but limited to strain mining without comprehensive enzyme characterizations [32, 35]. However, the wild-type strains are limited to specific types of reactions, which cannot be utilized to catalyze biotransformation beyond native catalytic capacity. For example, although S. cerevisiae was reported to be able to catalyze the reduction at C3, C20 and C17 of C21 and C19 steroids [36], it cannot catalyze the desired hydroxylation of steroids for the synthesis of hydrocortisone, which requires the C11-hydroxylation. It is important to characterize the enzymes responsible for the bioconversion in the corresponding microbial strains, and thus enable the establishment of programmable bioproduction of steroids using biotransformation strategy. More information about the microbial biotransformation in all types of microbes can be found in an earlier comprehensive review by Donova et al. [37]; here, we will only discuss the biotransformation efforts established in yeast (Table 1).
11α-hydroxylation (Fig. 2, Table 1) of progesterone is an important step in corticosteroid synthesis [38], and the biotransformation has been studied since 1950s [39]. Many microbial strains, such as Rhizopus nigricans and Aspergillus ochraceus [40, 41], have been reported to be able to catalyze this conversion. However, the enzyme responsible for 11α-hydroxylation in Rhizopus was not discovered until 2010 [42]. CYP509C12 from the filamentous fungi Rhizopus oryzae was characterized as the enzyme responsible for the synthesis of 11α-hydroxylation from progesterone [42]. Coexpression of CYP509C12 with its nature redox partner NADPH-cytochrome P450 reductase (RoCPR) in the fission yeast S. pombe 1445 led to 59.7% conversion of progesterone into 11α-hydroxyprogesterone and small amounts of a byproduct 6β-hydroxyprogesterone (~ 0.9%) [42]. CYP509C12 was also found to be able to catalyze the 11α-hydroxylation of TS, 11-deoxycorticosterone (DOC) and cortodoxone (RSS), with DOC exhibiting the highest conversion of progesterone but with low specificity (62% 11α-hydroxylated product and 6.8% 6β-hydroxylated byproduct) [42]. In 2017, another 11α-hydroxylase (11α-SHAoch) from the industrial steroid bioconversion strain A. ochraceus was characterized in S. cerevisiae [43]. Progesterone fed to the 11α-SHAoch-expressing yeast strain was converted into 11α-hydroxyprogesterone as the sole product with a high conversion at 90.9%, without coexpression of the native CPR [43]. In this study, CYP509C12 was expressed in S. cerevisiae as well as a comparison, resulting in a mixture of 11α-hydroxyprogesterone and 6β-hydroxyprogesterone (with the ratio of 7: 1). The amino acid sequences of 11α-SHAoch and CYP509C12 were then compared, which only share 15% identity at whole length and 26% identity in a C-terminus (174 amino acid long), indicating that they belong to different cytochrome P450 families [43].

a Structure of common steroids involved in biotransformation. Based on the biotransformation discussed in the review, they are divided into precursors and products. b Steroid products produced from yeast-based biotransformation through hydroxylation or dehydrogenation mentioned in this review. The functional groups of interests in the review are marked in red
The 11β-hydroxyl (Fig. 2, Table 1) is essential for the anti-inflammatory activity of relating steroid drugs, such as hydrocortisone [37]. The filamentous fungi Absidia orchidis was found to be able to catalyze 11β-hydroxylation on RSS to form hydrocortisone [44]. Recently, the enzymes responsible for the 11β-hydroxylation in A. orchidis were identified, including a cytochrome P450 CYP5311B2 and its native associating CPR and cytochrome b5 (CYB5) from A. orchidis [45]. The three enzymes were then functionally reconstituted in S. cerevisiae to catalyze the synthesis of hydrocortisone from RSS with the productivity of hydrocortisone at up to 22 mg/L day [45]. As a comparison, the mammalian 11β-hydroxylation system with CYP11B1 from Homo Sapiens and the redox partners for P450s, bovine adrenodoxin reductase (ADR) and adrenodoxin (ADX) were expressed in yeast, which produced 3 mg/L day of hydrocortisone, around sixfold lower than the A. orchidis system. The A. orchidis system (CYP5311B2, CPR and CYPB5) was further engineered for enhanced hydrocortisone production. The site-directed mutagenesis of CYP5311B2 was conducted and the R126D/Y398F mutant exhibited 3 times higher productivity of hydrocortisone compared with the wild-type CYP5311B2. To eliminate undesired side reactions in S. cerevisiae, two yeast endogenous enzymes, glycerol dehydrogenase GCY1 and aldosterone reductase YPR1 that can catalyze the C20-keto reduction of RSS, were inactivated. Additionally, various CPRs (CPRs from S. cerevisiae, Arabidopsis thaliana, Vitis vinifera, and A. orchidiswas) were introduced into S. cerevisiae to identify the optimal CPR for the function of CYP5311B2; with CPR from A. orchidiswas exhibiting the best performance. The hydrocortisone productivity was further enhanced to ~ 223 mg/L day upon tuning the expression level and the ratio among CYP5311B2, CPR and CYB5 in the engineered yeast strain, then to ~ 268 mg/L day through overexpression of steroid transporter CDR4 from Cochliobolus lunatus, and ultimately towards ~ 677 mg/L day by increasing the cell density to OD600 ~ 180. Interestingly, the engineered yeast strain exhibits an enhanced regioisomeric ratio (4:1) between the desired 11β-hydrocortisone to 11α-hydrocortisone, compared to 1:1 in A. orchidis, which also encodes 11α-hydroxylase [45]. This study demonstrates the high efficiency and regioselectivity of the engineered steroid biotransformation in S. cerevisiae, indicating yeast as a promising platform for the bioproduction of hydrocortisone through biotransformation [45].
Beside 11-hydroxyl, 14α-hydroxyl (Fig. 2, Table 1) is also an important structural feature and is responsible for the anti-gonadotropic and carcinolytic biological properties of 14α-hydroxysteroid drugs [46], such as 14α-hydroxy-4-androstene-3,6,17-trione (14α-OH-AT) [47]. P450lun from C. lunatus was originally characterized as a bifunctional hydroxylase that catalyzes both 11β- and 14α-hydroxylations of RSS, and has been utilized in the industrial production of hydrocortisone from RSS through 11β-hydroxylation [48]. Recently, the production of 14α-hydroxysteroids was established in S. cerevisiae using P450lun to catalyze the formation of 14α-hydroxyandrostenedione (14α-OH-AD) from androstenedione (AD) (regiospecificity > 99%) [48]. To enhance the efficiency and specificity of P450lun towards the synthesis of 14α-OH-AD, the native redox partner CPRlun was identified and co-expressed with P450lun with enhanced copy numbers in S. cerevisiae. The GCY1 and YPR1 genes were inactivated in the strain to eliminate the side reaction of the C20-keto reduction, same as in the biotransformation of hydrocortisone using CYP5311B2. The final engineered yeast strain synthesized 14α-OH-AD with 60% w/w yield and 99% regioisomeric purity converted from ~ 250 mg/L of fed AD [47, 48].
Boldenone (BD), the precursor to a number of steroid medicines, is normally synthesized from AD through ∆1 dehydrogenation and 17-keto reduction (Fig. 2, Table 1) [49]. A number of yeast strains have been found to be able to catalyze steroid 17β-reduction natively, including Pichia pastoris GS115 [49]. One recent example of yeast-based bioproduction of BD was achieved through overexpressing the 3-ketosteroid-Δ1-dehydrogenase KsdD2 from Rhodococcus rhodochrous in GS115 [49]. Both ∆1 dehydrogenated product androst-1,4-ene-3,17-dione (ADD) from AD and 17-keto reduced product BD from ADD were detected from the engineered yeast strain, indicating the 2-step formation of BD from AD in GS115 [49]. However, the conversion efficiency of AD into BD is low, at around 30% due to the insolubility of steroid in aqueous solution (AD was fed at 2 g/L). Therefore, the cosolvent 2-hydroxypropyl-β-cyclodextrin (HP-β-CD) was supplemented to improve the solubility of steroids in the medium and enhanced the conversion of AD into BD to 56.76% (with molar ratio 1: 1 of HP-β-CD to AD) [49]. The conversion was further improved with the bacterial and yeast co-culture strategy in the follow-up investigations [50, 51]. The bacterial strain Arthrobacter simplex previously identified to exhibit high ∆1-dehydrogenation activity was used to convert AD into ADD; and P. pastoris GS115 was engineered to express 17βHSD from S. cerevisiae to enhance the conversion of ADD into BD [50]. With the optimized medium condition (5.0 g/L AD with molar ratio 1:1 HP-β-CD and 75-g/L glucose), the conversion of AD into BD achieved 83% [50]. The efficient conversion of AD into BD in the bacteria–yeast coculture later on inspired Mycobacterium neoaurum–P. pastoris GS115 coculture, where M. neoaurum can natively convert phytosterols into ADD (with a small amount of AD supplemented) [51]. The two-stage fermentation of KsdD2-overexpressing M. neoaurum and 17βHSD-overexpressing GS115 produced BD at a conversion of 76% from fed phytosterols under the optimal medium condition (50 g/L glucose supplementation and 25 mM β-CD) [51].
Multi-step biotransformation has also been established in yeast (Table 1). Recently, the early steps of cardenolide formation, from pregnenolone to 5β‐pregnane‐3β,21‐diol‐20‐one, were reconstituted in S. cerevisiae [52], through expressing ∆5 3β-HSD from Digitalis lanata (Dl3β-HSD), ∆5 isomerase from Comamonas testosteronii (Ct3KSI), 5β reductase from Arabidopsis thaliana (AtSt5βR), and a 21-hydroxylase CYP21A1 from Mus musculus (MmCYP21A1). To avoid the esterification of pregnenolone which inhibits its conversion into downstream products, acetyl-CoA:pregnenolone acetyltransferase (APAT) encoded by ATF2 was disrupted. Subsequently, to enhance the performance of the mitochondria cytochrome P450 MmCYP21A1, the redox partners, ADR and ADX were co-expressed. Although the conversion (< 16% from 0.13 mM fed pregnenolone) is not as high as the aforementioned biotransformation using single or double enzymes, this strategy implicates the potential of programming yeast-based biotransformation for the production of more diverse steroid structures using enzymes of different functions and from various organisms [52].
Yeast has also been utilized to characterize several steroid hydroxylases that catalyze special hydroxylation activities. Recently, Thanatephorus cucumeris was found to be able to catalyze 7β-hydroxylation of RSS under neutral or acidic condition (Table 1) [53]. Transcriptome analysis of T. cucumeris was conducted upon RSS exposure at pH 6.0 and pH 9.0. The highly upregulated gene CYP5150AP3 and downregulated gene CYP5150AN1 at pH 6.0 were reconstituted in P. pastoris and presented special hydroxylation activities at C7 and C2. CY5150AP3 catalyzes 7β-hydroxylation on RSS and TS in yeast, with 10% side reaction of 6β-hydroxylation. CYP5150AN1 was identified to form a 2β-hydroxyl on RSS and produced 2β-OH-RSS as the sole product [53]. In addition to the fungal enzymes, CYP71D443 from Ajuga hairy roots was characterized in yeast as a C-22 hydroxylase that functions on 3β-hydroxy-5β-cholestan-6-one (5β-ketone) and 3β-hydroxy-5β-cholest-7-en-6-one (5β-ketol) [54].
Biotransformation of steroid drugs provides an environmentally more friendly approach for the industrial steroid production, compared to chemically catalyzed conversion [55]. The fast growing, genetically manipulatable, and fermentable yeast strains have been considered as a favorable host for biotechnological production [48]. As discussed in this session, different yeast strains have been utilized for the production of various steroids through biotransformation strategy and generally with good conversion efficiency. The medium condition of steroid biotransformation in yeast is relatively simple, usually common yeast rich or minimal medium supplemented with organic solvent, such as ethanol and N,N-dimethylformamide (DMF) to help dissolve the steroid substrates [48, 56]. Addition of β-cyclodextrin often helps the transport of steroid substrates into the cells [50, 57]. Enzymes responsible for steroid hydroxylation and dehydrogenation have been largely identified within the past 20 years, with many characterized in yeast. The massive strains with steroid transformation abilities discovered in the past decades are important resources for novel enzyme discovery; and the discovery of these enzymes set the foundation to use yeast as a versatile bioproduction platform for diverse steroid structures using the biotransformation strategy.
Synthesis of steroids without C24-Alkyl
De novo synthesis of steroids in yeast has gained much attention in the past decades [58]. The complete synthesis of steroid from simple carbon sources exploits the native ergosterol pathway to produce the desired sterol structure by inactivating late stage erg genes. The erg6 gene, encoding ∆24-sterol C-methyltransferase, can be inactivated to form sterols without C24-alkyl, which is a key characteristic of cholesterol. Cholesterol is the crucial component in animal cell membrane and precursor to Vitamin D and many other hormones [5, 6]. Cholesterol can also be found in many plants at a trace amount but is an important precursor to many specialized metabolites such as steroidal glycoalkaloids (SGAs) involving in the plant defense mechanism [8, 59].
The first stable cholesterol-producing S. cerevisiae strain RH6829 was established by Souza et al. in 2011 [60]. Compared to ergosterol, cholesterol does not own C24-methyl and has saturated C7–C8 bond and C22–C23 bond. ERG6 and ERG5 are responsible for the C24 methylation and C22–C23 desaturation, respectively. To redirect ergosterol synthesis to cholesterol, erg6 and erg5 were disrupted, and ∆7-reductase DHCR7 and ∆24(25)-reductase DHCR24, both from Danio rerio, were introduced to S. cerevisiae to generate strain RH6829 (Fig. 3, Table 2). S. cerevisiae RH6829 can stably produce cholesterol from simple carbon source, providing a yeast-based platform to synthesize specialized cholesterol and derivatives [60]. For example, RH6829 was applied to produce 13C-labeled cholesterol by culturing the strain RH6829 with 13C-labeled glucose. The 13C-labeled cholesterol was used to investigate influenza M2 protein–cholesterol interaction by NMR spectroscope [61].

Synthesis of steroids without C24-alkyl yeast. The characteristic C24 of cholesterol is marked in red. Single solid arrow represents reactions that have been validated in yeast, multiple solid arrows indicate multiple steps, and dashed arrows implicate conversions catalyzed by unknown enzyme(s). Compounds that potentially can be synthesized from yeast but not established yet are marked in blue. The endogenous ergosterol biosynthesis in yeast is highlighted in yellow, cholesterol is highlighted in blue, and the steroids derived from cholesterol are highlighted in pink. The enzymes marked in red represent the ones that require NADPH/NADP+, and the ones that require ATP are marked in blue
Another yeast strain P. pastoris has also been engineered to produce cholesterol using the same strategy as RH6829(Table 2) [62]. These cholesterol-producing yeast strains were proposed to be humanized strains and might be suitable hosts for the expression of mammalian membrane proteins. A membrane-binding human protein Na,K-ATPase α3β1 isoform, which requires the interaction with cholesterol and phospholipids to stabilize and enhance the ion transport activity, was used to examine how cholesterol incorporation affects the function of animal membrane proteins. In comparison to the wild-type P. pastoris, Na,K-ATPase α3β1 isoform exhibited longer protein half-life, correct co-localization in plasma membrane, and enhanced ATPase activity in the cholesterol-producing P. pastoris. The enhanced stability and activity of Na,K-ATPase α3β1 isoform in the cholesterol-producing P. pastoris imply the potential of using cholesterol-producing yeast strains to study and express animal membrane proteins [62].
Cholesterol is also the precursor to a number of specialized metabolites in plant, such as saponins and steroidal alkaloids [63]. The cholesterol-producing S. cerevisiae strain RH6829 is, thus, often used for enzyme functional characterization to elucidate the biosynthesis of cholesterol-derived phytochemicals. One recent example is the characterization of the 22R-hydroxylase CYP90B27 involved in the synthesis of polyphyllins in the medicinal herb Paris polyphylla [64]. The first step from cholesterol to saponins and steroidal alkaloids is believed to be C22-hydroxylation and catalyzed by a cytochrome P450. CYP90B27 was identified from functional transcriptome analysis, and the heterologous expression of CYP90B27 led to a descent conversion towards 22(R)-hydroxycholesterol, which confirmed CYP90B27 as the 22α-hydroxylase (Fig. 3, Table 2) [64]. More recently in 2019, RH6829 was utilized in the elucidation of the biosynthesis of diosgenin, the aglycone of a class of spirostanol-type saponins that is synthesized from cholesterol [65]. Similarly, PpCYP90G4-PpCYP94D108 and TfCYP90B50-TfCYP82J17 were identified to catalyze the conversion of cholesterol into diosgenin in P. polyphylla and Trigonella foenum-graecum, respectively, through transcriptome analysis and functional screening using Nicotiana benthamiana-based co-infiltration. The functions of PpCYP90G4, PpCYP94D108, TfCYP90B50, and TfCYP82J17 were further confirmed through functional reconstitutions in RH6829, which also enabled the first reported yeast bioproduction of diosgenin (Fig. 3, Table 2) [65]. Although yeast is mostly used as the characterization platform here instead of robust producers of the cholesterol-derived phytosteroids, these studies demonstrate the potential of using yeast as the platform for the bioproduction of various cholesterol-derived phytosteroids.
In addition to cholesterol-derived compounds, the Δerg6 mutant of S. cerevisiae has also been used to investigate plant cholesterol biosynthesis. Plant cholesterol biosynthesis also involves ∆24(25)-reductase [8]. To provide a platform for the putative Solanum tuberosum ∆24(25)-reductase screening, erg6 was disrupted in S. cerevisiae and StDWF5 (a ∆7-reductase from S. tuberosum) was expressed to accumulate desmosterol (T31) [66]. The expression of SSR2, the putative ∆24 reductase, in this desmosterol-accumulating yeast strain resulted in the production of cholesterol, which confirms that SSR2 functions as a ∆24(25)-reductase and is involved in cholesterol biosynthesis in S. tuberosum (Table 2). Moreover, co-expression of SSR2 with StDWF5 in the Δerg4Δerg5 double mutation and 24-methylenecholesterol-accumulating yeast strain (T21) also resulted in the synthesis of cholesterol. The synthesis of cholesterol in both engineered yeast strains implies that the Δ24(25)-reduction catalyzed by SSR2 could occur before the C-24 methylation catalyzed by ERG6 (Table 2) [66]. The versatility of yeast in sterol biosynthesis and engineering provides a unique platform for the clear elucidation of sterol biosynthesis.
The Δerg6 S. cerevisiae, with the lack of 24-alkylsterols, also provides a favorable platform for the characterization of C24-methyltransferase. Since ergosterol cannot be detected in the fungal pathogen Pneumocystis carinii, it is resistant to many common antifungal agents targeting ergosterol biosynthesis, and the S-adenosylmethionine:C-24 sterol methyltransferase (SAM:SMT, AdoMet) has been shown to be a promising target for the development of antifungal drug active against P. carinii [67]. However, P. carinii is difficult to study due to the lack of in vitro culture conditions. Instead, P. carinii SAM:SMT was heterologously expressed in the Δerg6 mutant S. cerevisiae for functional characterization, which revealed the function that this methyl transferase can transfer one or two methyl to C24 to form both 28-carbon and 29-carbon sterols [67].
Although inactivation of erg6 is not lethal to yeast, it affects yeast plasma physical properties. Beside the enhanced hypersensitivity to LiCl and hygromycin B as previously discussed [27], erg6 inactivation is lethal to yeast with tryptophan auxotroph because that the localization of tryptophan transporter TAT2 relies on the interaction with ergosterol in the plasma membrane [68]. The lack of ergosterol results in the mislocation of TAT2 in the vacuole and hinders the uptake of tryptophan. However, the production of cholesterol in Δerg6 mutant can restore the function of TAT2 misroute and recover the tryptophan transport capability [60]. Additionally, ergosterol is very important for the function of PDR12 [69], which is responsible for yeast resistance to weak organic acids. Mutant lacking ergosterol is hypersensitive to weak organic acid. These factors need to be considered during erg6 inactivation related yeast engineering. On the other hand, the Δerg6 mutant shows higher resistance to tomatidine, compared to the wild-type S. cerevisiae [70]. The enhanced resistance is likely due to the accumulation of zymosterol, implying the potential of using Δerg6 yeast mutants for the biosynthesis of steroidal alkaloid and saponins exhibiting antifungal activities [70].
Synthesis of steroids with saturated C7–C8 bond
ERG5 catalyzes C-22 desaturation, which is a unique characteristic of ergosterol, compared with cholesterol and phytosterol. The inactivation of erg5 resulted in the accumulation of ergosta-5,7-dienol-3β-ol, the precursor of Vitamin D4 [71]. The expression of DHCR24 from Homo sapiens in Δerg5 mutant S. cerevisiae led to the accumulate 7-dehydrocholesterol, the precursor of Vitamin D3 [72]. Phytosterols generally have saturated C7–C8 bond in the B ring, which is catalyzed by the ∆7-reductase (DWF5). DWF5 has been previously utilized for the de novo synthesis of hydrocortisone with the function of eight mammalian enzymes in S. cerevisiae [58]. Hydrocortisone was proposed to be converted from campesterol or brassicasterol of saturated C7–C8 in the B ring. The synthesis of hydrocortisone in the engineered S. cerevisiae with the titer up to 11.5 mg/L implies the significance of establishing efficient bioproduction of phytosterols in yeast [58].
Although stable synthesis of campesterol was established in S. cerevisiae as early of 2013 [60], most recent campesterol bioproduction efforts were focusing on engineering of Yarrowia lipolytica. The efficient production of campesterol in Y. lipolytica was first reported in 2016 [73]. 7-dehydrocholesterol reductases (DHCR7s) from Rallus norvegicus, Oryza saliva and Xenopus laevis were examined in Δerg5 mutant Y. lipolytica for the best production of campesterol (Fig. 4, Table 2). Out of three reductases, DHCR7 from X. laevis exhibited the highest campesterol production, up to 453 mg/L under the optimal medium optimization with sunflower seed oil as the carbon source under high-cell density fed-batch fermentation [73]. The campesterol-producing Y. lipolytica strain was further enhanced to 942 mg/L by substituting XlDHCR7 with DHCR7 from D. rerio and expressing peroxisome acyl-CoA oxidase 2 (POX2), an enzyme with high activity on the conversion of C16–18 long-chain fatty acids (the main components in sunflower seed oil) into acetyl-CoA [74]. Subsequently, the de novo production of pregnenolone, the early-stage intermediate for the synthesis of hydrocortisone, was established through coexpression of the sterol side-chain cleavage enzyme CYP11A1 with the redox partners bovine ADX and ADR in Y. lipolytica (Fig. 4, Table 2) [75]. Various CYP11A1 orthologs (from Homo sapiens, Bos taurus, Danio rerio, Lithobates catesbeiana, Gossypium arboretum, Taeniopygia guttata, Gallus gallus, Mus musculus) and redox partners were compared to locate the pair (CYP11A1 from Sus scrofa with ADX/ADR from Bos taurus) that provides the highest titer of pregnenolone, which was further optimized to 78 mg/L through expression level tuning and fermentation engineering [75].

Synthesis of steroids with saturated C7–C8 bond in yeast. The characteristic C7–C8 bond of campesterol is marked in red. Single solid arrow represents reactions that have been validated in yeast, multiple solid arrows indicate multiple steps, and dashed arrows implicate conversions catalyzed by unknown enzyme(s). Compounds potentially can be synthesized from yeast but not established yet are marked in blue. The endogenous ergosterol biosynthesis in yeast is highlighted in yellow, phytosterols are highlighted in green, and steroids synthesized from phytosterols are highlighted in pink. The enzymes marked in red represent the ones that require NADPH/NADP+, and the ones that require ATP are marked in blue
Besides steroid drugs, phytosterols are also precursors to an array of specialized metabolites in plants, such as brassinosteroids and withanolides. As discussed previously, phytosterol-producing yeast strains are useful platforms for the functional characterization of enzymes involved in the biosynthesis of specialized phytosteroids. Brassinosteroids are a group of phytohormones that play an essential role in plant growth, development, and resistance to both biotic and abiotic stress, and are mainly synthesized from campesterol [76]. In our most recent work, S. cerevisiae was engineered to express Δ7-sterol-C5-desaturase (DWF7), DWF5 and Δ24-sterol reductase (DWF1) from A. thaliana to synthesize campesterol. However, campesterol and the precursor 24-methylenecholesterol were found to be vigorously esterified and were merely accessible to the downstream tailoring enzymes such as CYP90B1 or SMT2 [77]. The inactivation of the native sterol acyltransferase genes ARE1 and ARE2 resulted in the synthesis of free phytosterols without much growth deficiency [78]. The introduction of SMT2 to the free campesterol-producing yeast strain led to the synthesis of β-sitosterol, the titer of which was then enhanced through inactivating erg4, the paralog and competing gene of dwf1 (Table 2). However, inactivation of erg4 resulted in dramatically reduced campesterol synthesis and significant growth deficiency, which were partially overcome by adaptive evolution. The free campesterol-producing yeast strain was also utilized as a platform to reconstitute the early part of the brassinolide biosynthetic pathway, using CYP90B1 and CYP90A1, towards the synthesis of 22(S)-hydroxycampest-4-en-3-one (Fig. 4, Table 2) [77]. On the other hand, withanolides are a group of bioactive steroids mainly found in the Solanoideae family, and are believed to be synthesized from 24-methylenecholesterol [79, 80]. The first step converting 24-methylenecholesterol towards the synthesis of withanolides has been very recently characterized to be catalyzed by a sterol Δ24-isomerase (24ISO) (Fig. 4, Table 2) [79]. 24ISO candidates were proposed from transcriptome analysis of different Solanoideae plants, and functionally identified in both plant tissues and the 24-methyldesmosterol-producing S. cerevisiae (strain T21) [79].
Most steroids with saturated C7–C8 and unsaturated C22–C23 bonds are phytosterol related. To date, except for campesterol production, metabolic engineering efforts in pursuing yeast-based bioproduction of phytosterol and phytosteroids are limiting. Yeast is more often utilized as an enzyme functional characterization tool. Although often with low efficiency, the structural diversity of phytosteroids produced in yeast has been broadened through this yeast-based enzyme characterization process, and thus shed lights on the synthetic potential of yeast-based steroid bioproduction.
Future perspective
There are several challenges remaining in establishing efficient steroid production in yeast. First, cytochrome P450s are often involved in steroid biosynthesis, which has been generally considered as a burden in yeast metabolic engineering of specialized metabolites [81]. In several examples discussed in this review, the co-expression and examination of different redox partners have been conducted for the sufficient electron transfer, enhanced conversion efficiency, and higher regioselectivity [45, 48, 75]. Second, steroids, such as campesterol and pregnanolone [75], are normally non-exportable, and the accumulation of heterologous steroids within the cells could be toxic to the yeast cells [82]. Further investigations on steroid transport and storage should be conducted to develop engineered transportation of steroids of cytotoxicity in yeast. Third, many steroids that can be potentially synthesized in yeast require highly complex synthetic pathways (e.g., brassinolide [83], tomatidine [84]), which is a general challenge in engineering de novo synthesis of complex structures in microbial hosts and has been carefully discussed in several recent reviews [81, 85, 86]. Last but not least, heterologous steroid production often requires the alternation of endogenous sterol synthesis, which may cause extra burden to the engineered yeast strains and growth deficiency. These challenges inevitably affect the engineered biosynthesis of steroids and may result in low production of target steroids.
One promising strategy to overcome these challenges is to use non-conventional yeast strains other than S. cerevisiae. Among the studies discussed in this review, multiple yeast strains have been utilized, such as S. cerevisiae, Y. lipolytica and P. pastoris, which are the most common selections and with different properties. S. cerevisiae has been extensively studied and utilized for genetic and metabolic engineering, with the well-established genetic modification tools [87]. Therefore, it is widely used in enzyme characterizations in addition to bioproduction purpose, such as the functional identification of CYP90B27 [64] and 24ISO [79]. Y. lipolytica is an industrial oleaginous strain and is usually used for high-yield lipid production, up to 70% w/w dry cell weight [88]. The high efficiency of acetyl-CoA synthesis and robust lipid storage make Y. lipolytica a favorable host for the hydrophobic steroid production [88,89,90]. P. pastoris can be cultured with methanol and grow at a relatively low pH, making the fermentation process less prone to microorganism contamination [91]. Moreover, unlike S. cerevisiae known to have hyperglycosylation with the addition of 50–150 mannose residues to heterologous proteins on average, P. pastoris exhibits much less glycosylation with an average of ~ 8–10 mannoses, providing a favorable yeast host when certain enzymes are difficult to be functionally reconstituted due to incorrect glycosylation [92]. P. pastoris also encodes an efficient secretion system [93], which may enable extracellular steroid synthesis aided by the efficient export of the steroid synthetic enzymes. In addition, using non-conventional yeast strains for specialized metabolite bioproduction has gained much attention recently [94, 95]. With the genetic engineering tools developed in these strains [96,97,98], there will be more choice of yeast strains that are feasible for the bioproduction of specialized steroids.
Directed evolution has been successfully utilized to engineer yeast strains for enhanced performance [99, 100], such as enhanced thermostability with altered sterol composition [101]. This strategy has also been successfully used to develop yeast adaptive mutants that overcome the growth deficiency caused by altered sterol metabolism in our efforts to develop β-sitosterol and 22(S)-hydroxycampest-4-en-3-one-producing yeast strains. The evolved strain was able to synthesize ~ sixfold more free campesterol and double the cell density at stationary phase, compared to the original free campesterol-producing strain [77]. Although the evolved strain still exhibits certain limitations, adaptive evolution is indispensable for the engineering and reconstitution of downstream metabolic pathways. The system biology and metabolic flux analysis, thus, become important to decipher how yeast was able to adapt its metabolism and growth to the altered sterol composition with excessive free sterols.
The system biology-based approaches have also been successfully utilized to increase the production of target metabolites and overcome the metabolic burdens in different microorganism hosts [102, 103]. The system-guided strategy will provide a promising route to tune the crosstalk between heterologous sterol pathway and the central sterol metabolism for enhanced performance of the engineered steroid-producing yeast strains, which are often of low efficiency. Although the system-level analysis of mevalonate pathway has been thoroughly investigated [104], few studies have been done on yeast endogenous sterol metabolism, as well as the crosstalk with the heterologous sterol-derived pathways, which, thus, become an appealing target for future investigation to better understand and engineer yeast for enhanced sterol production, especially in the case of de novo steroid biosynthesis.
Pathway compartmentalization and metabolite channeling have been successfully applied to enhance the efficiency of many heterologous pathways in yeast [105, 106], mainly through enhancing the local substrate concentrations, separating desired reactions from the competing side reactions, and decreasing the release of the toxic intermediates into the cells [102, 107, 108]. The synthesis of steroids in yeast takes place on endoplasmic reticulum (ER) membrane with excessive sterols generally esterified and stored in the lipid droplets [109]. As described in our recent work, the natural compartmentalization of sterol synthesis and esterification in yeast blocked the access of downstream enzymes (e.g., CYB90B1, SMT2) to the sterol substrates, which leads to the failed synthesis of the target products. Although inactivation of the acyltransferases ARE1 and ARE2 enabled the synthesis of free sterol and downstream target metabolites, it also introduced growth deficiency and decreased overall sterol production, which is possibly due to the cytotoxicity of excessive free sterols [77]. In such a case, compartmentalization and intermediate channeling may provide a unique tool to circumvent the undesired crosstalk between yeast sterol esterification and the heterologous sterol pathway, and decrease the toxicity caused by the presence of heterologous sterols unnatural to yeast.
Last but not least, feeding steroid intermediates as the precursors is also a promising approach for the production of high-value specialized steroids. There are two main challenges to use biotransformation for steroid production: yeast exogenous sterol uptake mechanism prefers anaerobic condition, and many value-added steroids of complex structural features require large numbers of enzymes due to the lack of sterol intermediates that are commercially available. Although the uptake control mutant (UPC2-1) has been shown to enable aerobic uptake of sterols [109], the production of target steroids can also be limited by yeast intracellular capacity of sterols. Thus, the choice of establishing de novo synthesis or biotransformation depends on a number of factors, including but not limited to the availability of substrates, chemical characteristics of the substrates, number of heterologous enzymes required, and type of yeast to be engineered.
Conclusion
To date, steroids that have been synthesized from yeast can be generally divided into 3 groups: (1) C19 or C21 steroid drugs, such as boldenone and hydrocortisone [45, 49], (2) cholesterol and its derivatives [60, 65, 79] and (3) phytosterols and their derivatives [54, 73]. The steroid drug biotransformation in yeast strains has been discovered since early last century, yet it is not until the recent 20 years that enzymes responsible for the steroid biotransformation were characterized and their recombinant forms expressed in yeast. The discovery of steroid transforming enzymes from various microbial hosts enables the engineering of yeast as versatile platforms catalyzing diversified steroidal structural alternations such as 11α-, 11β-, and 14α-hydroxylation [42, 45, 48]. The de novo synthesis of cholesterol in yeast was first reported in 2011 [60], which is a milestone of yeast heterologous sterol synthesis and yeast steroid production has attracted much attention since then [61, 62, 64,65,66]. Cholesterol-producing yeast strains could serve as hosts to express mammalian membrane-bound proteins that require interaction with cholesterol to fold and function [62]. The various sterol-producing yeast strains have also been widely used to characterize enzyme activity and elucidate the biosynthesis of specialized steroids from various species. Elucidation and reconstitution of the biosynthesis of steroids of diverse structures and origins highlight the steroid structural diversity that can be established in yeast. Although the yeast-based synthesis of specialized steroids has just started in the past few years, the broad structural diversity established in yeast suggests the potential of using yeast-based steroid bioproduction as a promising sourcing mechanism for this group of molecules. The past decade of engineering efforts has demonstrated yeast as a unique and promising bioproduction platform for specialized steroids, yet this is just a start, with a multitude of challenges remaining to be resolved.
References
Dufourc EJ (2008) Sterols and membrane dynamics. J Chem Biol. 1(1–4):63–77. https://doi.org/10.1007/s12154-008-0010-6
Ostlund RE Jr (2004) Phytosterols and cholesterol metabolism. Curr Opin Lipidol 15(1):37–41. https://doi.org/10.1097/00041433-200402000-00008
Piironen V, Lindsay DG, Miettinen TA, Toivo J, Lampi AM (2000) Plant sterols: biosynthesis, biological function and their importance to human nutrition. J Sci Food Agr. 80(7):939–966. https://doi.org/10.1002/(Sici)1097-0010(20000515)80:7%3c939:Aid-Jsfa644%3e3.0.Co;2-C
Festucci-Buselli RA, Contim LAS, Barbosa LCA, Stuart J, Otoni WC (2008) Biosynthesis and potential functions of the ecdysteroid 20-hydroxyecdysone—a review. Botany. 86(9):978–987. https://doi.org/10.1139/B08-049
Li S, He Y, Lin S, Hao L, Ye Y, Lv L, Sun Z, Fan H, Shi Z, Li J, Feng R, Na L, Wang Y, Li Y, Sun C (2016) Increase of circulating cholesterol in vitamin D deficiency is linked to reduced vitamin D receptor activity via the Insig-2/SREBP-2 pathway. Mol Nutr Food Res 60(4):798–809. https://doi.org/10.1002/mnfr.201500425
Tuckey RC (2005) Progesterone synthesis by the human placenta. Placenta 26(4):273–281. https://doi.org/10.1016/j.placenta.2004.06.012
Khripach V, Zhabinskii V, De Groot A (2000) Twenty years of brassinosteroids: steroidal plant hormones warrant better crops for the XXI century. Ann Bot 86(3):441–447. https://doi.org/10.1006/anbo.2000.1227
Sonawane PD, Pollier J, Panda S, Szymanski J, Massalha H, Yona M, Unger T, Malitsky S, Arendt P, Pauwels L, Almekias-Siegl E, Rogachev I, Meir S, Cardenas PD, Masri A, Petrikov M, Schaller H, Schaffer AA, Kamble A, Giri AP, Goossens A, Aharoni A (2016) Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat Plants. 3:16205. https://doi.org/10.1038/nplants.2016.205
Amarelle L, Lecuona E (2018) The antiviral effects of Na, K-ATPase inhibition: a minireview. Int J Mol Sci. https://doi.org/10.3390/ijms19082154(Epub 2018/07/26)
Arthan D, Svasti J, Kittakoop P, Pittayakhachonwut D, Tanticharoen M, Thebtaranonth Y (2002) Antiviral isoflavonoid sulfate and steroidal glycosides from the fruits of Solanum torvum. Phytochemistry. 59(4):459–463. https://doi.org/10.1016/s0031-9422(01)00417-4(Epub 2002/02/07)
Ikeda T, Ando J, Miyazono A, Zhu XH, Tsumagari H, Nohara T, Yokomizo K, Uyeda M (2000) Anti-herpes virus activity of Solanum steroidal glycosides. Biol Pharm Bull. 23(3):363–364. https://doi.org/10.1248/bpb.23.363(Epub 2000/03/22)
Wachsman MB, Castilla V (2012) Antiviral properties of brassinosteroids. Brassinosteroids: practical applications in agriculture and human health, pp 57–71
QY Research (2018) Global Steroids Market: Corticosteroids Segment to Reach Value of US$ 8.6 Bn by 2025 end - QY Research, Inc. Available via PRNewswire. https://www.prnewswire.com/in/news-releases/global-steroids-market-corticosteroids-segment-to-reach-value-of-us-86-bn-by-2025-end—qy-research-inc-693708281.html
Kutscher EC, Lund BC, Perry PJ (2002) Anabolic steroids. Sports Med 32(5):285–296
Barnes PJ (2006) Corticosteroids: the drugs to beat. Eur J Pharmacol 533(1–3):2–14. https://doi.org/10.1016/j.ejphar.2005.12.052
Bernstein JJJ, Holt GB, Bernstein J (2019) Price dispersion of generic medications. PLoS One 14(11):e0225280. https://doi.org/10.1371/journal.pone.0225280
Wang Y, Jiang W, He Q, Wang C, Wang B, Zhou P, Dong N, Tong Q (2020) Early, low-dose and short-term application of corticosteroid treatment in patients with severe COVID-19 pneumonia: single-center experience from Wuhan, China. medRxiv
Mahase E (2020) Covid-19: Low dose steroid cuts death in ventilated patients by one third, trial finds. BMJ 369:m2422. https://doi.org/10.1136/bmj.m2422
Brower V (2008) Back to nature: extinction of medicinal plants threatens drug discovery. J Natl Cancer Inst 100(12):838–839. https://doi.org/10.1093/jnci/djn199
Wriessnegger T, Pichler H (2013) Yeast metabolic engineering–targeting sterol metabolism and terpenoid formation. Prog Lipid Res 52(3):277–293. https://doi.org/10.1016/j.plipres.2013.03.001
Sokolov SS, Trushina NI, Severin FF, Knorre DA (2019) Ergosterol turnover in yeast: an interplay between biosynthesis and transport. Biochemistry (Mosc). 84(4):346–357. https://doi.org/10.1134/S0006297919040023
Liu JF, Xia JJ, Nie KL, Wang F, Deng L (2019) Outline of the biosynthesis and regulation of ergosterol in yeast. World J Microbiol Biotechnol 35(7):98. https://doi.org/10.1007/s11274-019-2673-2
Desmond E, Gribaldo S (2009) Phylogenomics of sterol synthesis: insights into the origin, evolution, and diversity of a key eukaryotic feature. Genome Biol Evol. 1:364–381. https://doi.org/10.1093/gbe/evp036
Russell DW (1992) Cholesterol biosynthesis and metabolism. Cardiovasc Drugs Ther 6(2):103–110. https://doi.org/10.1007/bf00054556
Lees ND, Skaggs B, Kirsch DR, Bard M (1995) Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review. Lipids 30(3):221–226. https://doi.org/10.1007/bf02537824
Palermo LM, Leak FW, Tove S, Parks LW (1997) Assessment of the essentiality of ERG genes late in ergosterol biosynthesis in Saccharomyces cerevisiae. Curr Genet 32(2):93–99. https://doi.org/10.1007/s002940050252
Johnston EJ, Moses T, Rosser SJ (2020) The wide-ranging phenotypes of ergosterol biosynthesis mutants, and implications for microbial cell factories. Yeast 37(1):27–44. https://doi.org/10.1002/yea.3452
Heese-Peck A, Pichler H, Zanolari B, Watanabe R, Daum G, Riezman H (2002) Multiple functions of sterols in yeast endocytosis. Mol Biol Cell 13(8):2664–2680. https://doi.org/10.1091/mbc.e02-04-0186
Barreto L, Canadell D, Petrezselyova S, Navarrete C, Maresova L, Perez-Valle J, Herrera R, Olier I, Giraldo J, Sychrova H, Yenush L, Ramos J, Arino J (2011) A genomewide screen for tolerance to cationic drugs reveals genes important for potassium homeostasis in Saccharomyces cerevisiae. Eukaryot Cell 10(9):1241–1250. https://doi.org/10.1128/EC.05029-11
Kodedova M, Sychrova H (2015) Changes in the sterol composition of the plasma membrane affect membrane potential, salt tolerance and the activity of multidrug resistance pumps in Saccharomyces cerevisiae. PLoS One 10(9):e0139306. https://doi.org/10.1371/journal.pone.0139306
Liu G, Chen Y, Faergeman NJ, Nielsen J (2017) Elimination of the last reactions in ergosterol biosynthesis alters the resistance of Saccharomyces cerevisiae to multiple stresses. Fems Yeast Res. https://doi.org/10.1093/femsyr/fox063
Kristan K, Rizner TL (2012) Steroid-transforming enzymes in fungi. The Journal of steroid biochemistry and molecular biology. 129(1–2):79–91. https://doi.org/10.1016/j.jsbmb.2011.08.012
Charney W, Herzog HL (1967) Microbial transformations of steroids; a handbook, vol xi. Academic Press, New York, p 728
Pajic T, Vitas M, Zigon D, Pavko A, Kelly SL, Komel R (1999) Biotransformation of steroids by the fission yeast Schizosaccharomyces pombe. Yeast 15(8):639–645. https://doi.org/10.1002/(Sici)1097-0061(19990615)15:8%3c639:Aid-Yea408%3e3.0.Co;2-3
Mauersberger S, Novikova LA, Shkumatov VM (2013) Cytochrome P450 expression in Yarrowia lipolytica and its use in steroid biotransformation. In: Barth G (ed) Yarrowia lipolytica. Microbiology Monographs, vol 25. Springer, Berlin, Heidelberg, pp 171–226
Donova MV, Egorova OV, Nikolayeva VM (2005) Steroid 17 beta-reduction by microorganisms—a review. Process Biochem 40(7):2253–2262. https://doi.org/10.1016/j.procbio.2004.09.025
Donova MV, Egorova OV (2012) Microbial steroid transformations: current state and prospects. Appl Microbiol Biotechnol 94(6):1423–1447. https://doi.org/10.1007/s00253-012-4078-0
Hosseinabadi T, Vahidi H, Nickavar B, Kobarfard F (2015) Biotransformation of progesterone by whole cells of filamentous fungi Aspergillus brasiliensis. Iran J Pharm Res. 14(3):919–924
Murray HC, Peterson DH (1952) Oxygenation of steroids by Mucorales fungi US Patent No 2,602,769 8 Jul 1952
Ghosh D, Samanta TB (1981) 11 alpha-Hydroxylation of progesterone by cell free preparation of Aspergillus ochraceus TS. J Steroid Biochem. 14(10):1063–1067. https://doi.org/10.1016/0022-4731(81)90217-x
Volker EJ (1977) The microbial hydroxylation of progesterone. J Chem Educ 54(1):65–66
Petric S, Hakki T, Bernhardt R, Zigon D, Cresnar B (2010) Discovery of a steroid 11alpha-hydroxylase from Rhizopus oryzae and its biotechnological application. J Biotechnol 150(3):428–437. https://doi.org/10.1016/j.jbiotec.2010.09.928
Hull CM, Warrilow AGS, Rolley NJ, Price CL, Donnison IS, Kelly DE, Kelly SL (2017) Co-production of 11 alpha-hydroxyprogesterone and ethanol using recombinant yeast expressing fungal steroid hydroxylases. Biotechnol Biofuels. https://doi.org/10.1186/s13068-017-0904-z
Herraiz I (2017) Chemical pathways of corticosteroids, industrial synthesis from sapogenins. Methods Mol Biol. 1645:15–27. https://doi.org/10.1007/978-1-4939-7183-1_2
Chen J, Fan FY, Qu G, Tang JL, Xi YY, Bi CH, Sun ZT, Zhang XL (2020) Identification of Absidia orchidis steroid 11 beta-hydroxylation system and its application in engineering Saccharomyces cerevisiae for one-step biotransformation to produce hydrocortisone. Metab En. 57:31–42. https://doi.org/10.1016/j.ymben.2019.10.006
Salvador JA, Carvalho JF, Neves MA, Silvestre SM, Leitao AJ, Silva MM, Sa e Melo ML (2013) Anticancer steroids: linking natural and semi-synthetic compounds. Nat Prod Rep 30(2):324–374. https://doi.org/10.1039/c2np20082a
Andryushina VA, Voishvillo NE, Druzhinina AV, Stytsenko TS, Yaderets VV, Petrosyan MA, Zeinalov OA (2013) 14 alpha-Hydroxylation of steroids by mycelium of the mold fungus Curvularia lunata (VKPM F-981) to produce precursors for synthesizing new steroidal drugs. Pharm Chem J. 47(2):103–108. https://doi.org/10.1007/s11094-013-0905-6
Chen J, Tang JL, Xi YY, Dai ZB, Bi CH, Chen X, Fan FY, Zhang XL (2019) Production of 14 alpha-hydroxysteroids by a recombinant Saccharomyces cerevisiae biocatalyst expressing of a fungal steroid 14 alpha-hydroxylation system. Appl Microbiol Biot. 103(20):8363–8374. https://doi.org/10.1007/s00253-019-10076-x
Tang R, Ji PL, Yu Y, Yang X, Liu MJ, Chen KY, Shen YB, Wang M (2018) Biotransformation to produce boldenone by Pichia pastoris GS115 engineered recombinant strains. Lect Notes Electr En. 444:117–124. https://doi.org/10.1007/978-981-10-4801-2_12
Tang R, Shen YB, Wang M, Zhou HJ, Zhao YQ (2019) Highly efficient synthesis of boldenone from androst-4-ene-3,17-dione by Arthrobacter simplex and Pichia pastoris ordered biotransformation. Bioproc Biosyst Eng. 42(6):933–940. https://doi.org/10.1007/s00449-019-02092-y
Tang R, Shen YB, Xia ML, Tu LN, Luo JM, Geng YH, Gao T, Zhou HJ, Zhao YQ, Wang M (2019) A highly efficient step-wise biotransformation strategy for direct conversion of phytosterol to boldenone. Bioresource Technol. 283:242–250. https://doi.org/10.1016/j.biortech.2019.03.058
Rieck C, Geiger D, Munkert J, Messerschmidt K, Petersen J, Strasser J, Meitinger N, Kreis W (2019) Biosynthetic approach to combine the first steps of cardenolide formation in Saccharomyces cerevisiae. Microbiologyopen. 8(12):e925. https://doi.org/10.1002/mbo3.925
Lu W, Feng J, Chen X, Bao Y-J, Wang Y, Wu Q, Ma Y, Zhu D (2019) Distinct regioselectivity of fungal P450 enzymes for steroidal hydroxylation. Appl Environ Microbiol 85(18):e01182-19. https://doi.org/10.1128/aem.01182-19
Tsukagoshi Y, Ohyama K, Seki H, Akashi T, Muranaka T, Suzuki H, Fujimoto Y (2016) Functional characterization of CYP71D443, a cytochrome P450 catalyzing C-22 hydroxylation in the 20-hydroxyecdysone biosynthesis of Ajuga hairy roots. Phytochemistry 127:23–28. https://doi.org/10.1016/j.phytochem.2016.03.010
Hegazy MEF, Mohamed TA, ElShamy AI, Mohamed AH, Mahalel UA, Reda EH, Shaheen AM, Tawfik WA, Shahat AA, Shams KA, Abdel-Azim NS, Hammouda FM (2015) Microbial biotransformation as a tool for drug development based on natural products from mevalonic acid pathway: a review. J Adv Res. 6(1):17–33. https://doi.org/10.1016/j.jare.2014.11.009
Naumann JM, Zollner A, Dragan CA, Messinger J, Adam J, Bureik M (2011) Biotechnological production of 20-alpha-dihydrodydrogesterone at pilot scale. Appl Biochem Biotech. 165(1):190–203. https://doi.org/10.1007/s12010-011-9243-x
Manosroi J, Salowakhon S, Manosroi A (2008) Enhancement of 17 alpha-hydroxyprogesterone production from progesterone by biotransformation using hydroxypropyl-beta-cyclodextrin complexation technique. J Steroid Biochem. 112(4–5):201–204. https://doi.org/10.1016/j.jsbmb.2008.10.003
Szczebara FM, Chandelier C, Villeret C, Masurel A, Bourot S, Duport C, Blanchard S, Groisillier A, Testet E, Costaglioli P, Cauet G, Degryse E, Balbuena D, Winter J, Achstetter T, Spagnoli R, Pompon D, Dumas B (2003) Total biosynthesis of hydrocortisone from a simple carbon source in yeast. Nat Biotechnol 21(2):143–149. https://doi.org/10.1038/nbt775
Milner SE, Brunton NP, Jones PW, O’Brien NM, Collins SG, Maguire AR (2011) Bioactivities of glycoalkaloids and their aglycones from solanum species. J Agric Food Chem. 59(8):3454–3484. https://doi.org/10.1021/jf200439q
Souza CM, Schwabe TM, Pichler H, Ploier B, Leitner E, Guan XL, Wenk MR, Riezman I, Riezman H (2011) A stable yeast strain efficiently producing cholesterol instead of ergosterol is functional for tryptophan uptake, but not weak organic acid resistance. Metab Eng 13(5):555–569. https://doi.org/10.1016/j.ymben.2011.06.006
Elkins MR, Sergeyev IV, Hong M (2018) Determining cholesterol binding to membrane proteins by cholesterol (13)C labeling in yeast and dynamic nuclear polarization NMR. J Am Chem Soc 140(45):15437–15449. https://doi.org/10.1021/jacs.8b09658
Hirz M, Richter G, Leitner E, Wriessnegger T, Pichler H (2013) A novel cholesterol-producing Pichia pastoris strain is an ideal host for functional expression of human Na, K-ATPase alpha 3 beta 1 isoform. Appl Microbiol Biot. 97(21):9465–9478. https://doi.org/10.1007/s00253-013-5156-7
Yuan CS, Wang CZ, Wicks SM, Qi LW (2010) Chemical and pharmacological studies of saponins with a focus on American ginseng. J Ginseng Res. 34(3):160–167. https://doi.org/10.5142/jgr.2010.34.3.160
Yin Y, Gao L, Zhang X, Gao W (2018) A cytochrome P450 monooxygenase responsible for the C-22 hydroxylation step in the Paris polyphylla steroidal saponin biosynthesis pathway. Phytochemistry 156:116–123. https://doi.org/10.1016/j.phytochem.2018.09.005
Christ B, Xu C, Xu M, Li FS, Wada N, Mitchell AJ, Han XL, Wen ML, Fujita M, Weng JK (2019) Repeated evolution of cytochrome P450-mediated spiroketal steroid biosynthesis in plants. Nat Commun. 10(1):3206. https://doi.org/10.1038/s41467-019-11286-7
Sawai S, Ohyama K, Yasumoto S, Seki H, Sakuma T, Yamamoto T, Takebayashi Y, Kojima M, Sakakibara H, Aoki T, Muranaka T, Saito K, Umemoto N (2014) Sterol side chain reductase 2 Is a key enzyme in the biosynthesis of cholesterol, the common precursor of toxic steroidal glycoalkaloids in potato. Plant Cell 26(9):3763–3774. https://doi.org/10.1105/tpc.114.130096
Kaneshiro ES, Johnston LQ, Nkinin SW, Romero BI, Giner JL (2015) Sterols of Saccharomyces cerevisiae erg6 knockout mutant expressing the Pneumocystis carinii S-Adenosylmethionine: sterol C-24 methyltransferase (vol 62, pg 298, 2015). J Eukaryot Microbiol. 62(4):566. https://doi.org/10.1111/jeu.12236
Gaber RF, Copple DM, Kennedy BK, Vidal M, Bard M (1989) The yeast gene Erg6 is required for normal membrane-function but is not essential for biosynthesis of the cell-cycle-sparking sterol. Mol Cell Biol 9(8):3447–3456. https://doi.org/10.1128/Mcb.9.8.3447
Mollapour M, Fong D, Balakrishnan K, Harris N, Thompson S, Schuller C, Kuchler K, Piper PW (2004) Screening the yeast deletant mutant collection for hypersensitivity and hyper-resistance to sorbate, a weak organic acid food preservative. Yeast 21(11):927–946. https://doi.org/10.1002/yea.1141
Simons V, Morrissey JP, Latijnhouwers M, Csukai M, Cleaver A, Yarrow C, Osbourn A (2006) Dual effects of plant steroidal alkaloids on Saccharomyces cerevisiae. Antimicrob Agents Chemother 50(8):2732–2740. https://doi.org/10.1128/AAC.00289-06
Ma BX, Ke X, Tang XL, Zheng RC, Zheng YG (2018) Rate-limiting steps in the Saccharomyces cerevisiae ergosterol pathway: towards improved ergosta-5,7-dien-3beta-ol accumulation by metabolic engineering. World J Microbiol Biotechnol 34(4):55. https://doi.org/10.1007/s11274-018-2440-9
Su W, Xiao WH, Wang Y, Liu D, Zhou X, Yuan YJ (2015) Alleviating redox imbalance enhances 7-dehydrocholesterol production in engineered Saccharomyces cerevisiae. PLoS One 10(6):e0130840. https://doi.org/10.1371/journal.pone.0130840
Du HX, Xiao WH, Wang Y, Zhou X, Zhang Y, Liu D, Yuan YJ (2016) Engineering Yarrowia lipolytica for campesterol overproduction. PLoS One 11(1):e0146773. https://doi.org/10.1371/journal.pone.0146773
Zhang Y, Wang Y, Yao M, Liu H, Zhou X, Xiao W, Yuan Y (2017) Improved campesterol production in engineered Yarrowia lipolytica strains. Biotechnol Lett 39(7):1033–1039. https://doi.org/10.1007/s10529-017-2331-4
Zhang R, Zhang Y, Wang Y, Yao M, Zhang J, Liu H, Zhou X, Xiao W, Yuan Y (2019) Pregnenolone overproduction in Yarrowia lipolytica by integrative components pairing of the cytochrome P450scc system. ACS Synth Biol. 8(12):2666–2678. https://doi.org/10.1021/acssynbio.9b00018
Oklestkova J, Rarova L, Kvasnica M, Strnad M (2015) Brassinosteroids: synthesis and biological activities. Phytochem Rev 14(6):1053–1072. https://doi.org/10.1007/s11101-015-9446-9
Xu S, Chen C, Li Y (2020) Engineering of phytosterol-producing yeast platforms for functional reconstitution of downstream biosynthetic pathways. bioRxiv. https://doi.org/10.1101/2020.06.08.139279
Zweytick D, Leitner E, Kohlwein SD, Yu C, Rothblatt J, Daum G (2000) Contribution of Are1p and Are2p to steryl ester synthesis in the yeast Saccharomyces cerevisiae. Eur J Biochem 267(4):1075–1082
Dhar N, Razdan S, Rana S, Bhat WW, Vishwakarma R, Lattoo SK (2015) A decade of molecular understanding of withanolide biosynthesis and in vitro studies in Withania somnifera (L.) dunal: prospects and perspectives for pathway engineering. Front Plant Sci 6:1031. https://doi.org/10.3389/fpls.2015.01031
Knoch E, Sugawara S, Mori T, Poulsen C, Fukushima A, Harholt J, Fujimoto Y, Umemoto N, Saito K (2018) Third DWF1 paralog in Solanaceae, sterol Delta(24)-isomerase, branches withanolide biosynthesis from the general phytosterol pathway. Proc Natl Acad Sci USA 115(34):E8096–E8103. https://doi.org/10.1073/pnas.1807482115
Li SJ, Li YR, Smolke CD (2018) Strategies for microbial synthesis of high-value phytochemicals. Nat Chem. 10(4):395–404. https://doi.org/10.1038/s41557-018-0013-z
Csaky Z, Garaiova M, Kodedova M, Valachovic M, Sychrova H, Hapala I (2020) Squalene lipotoxicity in a lipid droplet-less yeast mutant is linked to plasma membrane dysfunction. Yeast 37(1):45–62. https://doi.org/10.1002/yea.3454
Fujioka S, Yokota T (2003) Biosynthesis and metabolism of brassinosteroids. Annu Rev Plant Biol. 54:137–164. https://doi.org/10.1146/annurev.arplant.54.031902.134921
Itkin M, Heinig U, Tzfadia O, Bhide AJ, Shinde B, Cardenas PD, Bocobza SE, Unger T, Malitsky S, Finkers R, Tikunov Y, Bovy A, Chikate Y, Singh P, Rogachev I, Beekwilder J, Giri AP, Aharoni A (2013) Biosynthesis of antinutritional alkaloids in solanaceous crops is mediated by clustered genes. Science 341(6142):175–179. https://doi.org/10.1126/science.1240230
Pyne ME, Narcross L, Martin VJJ (2019) Engineering plant secondary metabolism in microbial systems. Plant Physiol 179(3):844–861. https://doi.org/10.1104/pp.18.01291
Tatsis EC, O’Connor SE (2016) New developments in engineering plant metabolic pathways. Curr Opin Biotechnol. 42:126–132. https://doi.org/10.1016/j.copbio.2016.04.012
Siddiqui MS, Thodey K, Trenchard I, Smolke CD (2012) Advancing secondary metabolite biosynthesis in yeast with synthetic biology tools. FEMS Yeast Res 12(2):144–170. https://doi.org/10.1111/j.1567-1364.2011.00774.x
Adrio JL (2017) Oleaginous yeasts: promising platforms for the production of oleochemicals and biofuels. Biotechnol Bioeng 114(9):1915–1920. https://doi.org/10.1002/bit.26337
Tai M, Stephanopoulos G (2013) Engineering the push and pull of lipid biosynthesis in oleaginous yeast Yarrowia lipolytica for biofuel production. Metab Eng. 15:1–9. https://doi.org/10.1016/j.ymben.2012.08.007
Worland AM, Czajka JJ, Li Y, Wang Y, Tang YJ, Su WW (2020) Biosynthesis of terpene compounds using the non-model yeast Yarrowia lipolytica: grand challenges and a few perspectives. Curr Opin Biotechnol. 64:134–140. https://doi.org/10.1016/j.copbio.2020.02.020
Looser V, Bruhlmann B, Bumbak F, Stenger C, Costa M, Camattari A, Fotiadis D, Kovar K (2015) Cultivation strategies to enhance productivity of Pichia pastoris: a review. Biotechnol Adv 33(6 Pt 2):1177–1193. https://doi.org/10.1016/j.biotechadv.2015.05.008
Lehle L (1992) Protein glycosylation in yeast. Antonie Van Leeuwenhoek 61(2):133–134. https://doi.org/10.1007/bf00580620
Ahmad M, Hirz M, Pichler H, Schwab H (2014) Protein expression in Pichia pastoris: recent achievements and perspectives for heterologous protein production. Appl Microbiol Biotechnol 98(12):5301–5317. https://doi.org/10.1007/s00253-014-5732-5
Fatma Z, Schultz C, Zhao H (2020) Recent advances in domesticating non-model microorganisms. Biotechnol Prog. https://doi.org/10.1002/btpr.3008
Thorwall S, Schwartz C, Chartron JW, Wheeldon I (2020) Stress-tolerant non-conventional microbes enable next-generation chemical biosynthesis. Nat Chem Biol 16(2):113–121. https://doi.org/10.1038/s41589-019-0452-x
Cai P, Gao J, Zhou Y (2019) CRISPR-mediated genome editing in non-conventional yeasts for biotechnological applications. Microb Cell Fact 18(1):63. https://doi.org/10.1186/s12934-019-1112-2
Lobs AK, Schwartz C, Wheeldon I (2017) Genome and metabolic engineering in non-conventional yeasts: current advances and applications. Synth Syst Biotechnol. 2(3):198–207. https://doi.org/10.1016/j.synbio.2017.08.002
Wagner JM, Alper HS (2016) Synthetic biology and molecular genetics in non-conventional yeasts: current tools and future advances. Fungal Genet Biol. 89:126–136. https://doi.org/10.1016/j.fgb.2015.12.001
Cakar ZP, Turanli-Yildiz B, Alkim C, Yilmaz U (2012) Evolutionary engineering of Saccharomyces cerevisiae for improved industrially important properties. FEMS Yeast Res 12(2):171–182. https://doi.org/10.1111/j.1567-1364.2011.00775.x
Dragosits M, Mattanovich D (2013) Adaptive laboratory evolution—principles and applications for biotechnology. Microb Cell Fact. 12:64. https://doi.org/10.1186/1475-2859-12-64
Caspeta L, Chen Y, Ghiaci P, Feizi A, Buskov S, Hallstrom BM, Petranovic D, Nielsen J (2014) Biofuels. Altered sterol composition renders yeast thermotolerant. Science. 346(6205):75–78. https://doi.org/10.1126/science.1258137
Wu G, Yan Q, Jones JA, Tang YJ, Fong SS, Koffas MAG (2016) Metabolic burden: cornerstones in synthetic biology and metabolic engineering applications. Trends Biotechnol 34(8):652–664. https://doi.org/10.1016/j.tibtech.2016.02.010
Wu SG, He L, Wang Q, Tang YJ (2015) An ancient Chinese wisdom for metabolic engineering: Yin-Yang. Microb Cell Fact. 14:39. https://doi.org/10.1186/s12934-015-0219-3
Martien JI, Amador-Noguez D (2017) Recent applications of metabolomics to advance microbial biofuel production. Curr Opin Biotechnol. 43:118–126. https://doi.org/10.1016/j.copbio.2016.11.006
Avalos JL, Fink GR, Stephanopoulos G (2013) Compartmentalization of metabolic pathways in yeast mitochondria improves the production of branched-chain alcohols. Nat Biotechnol 31(4):335–341. https://doi.org/10.1038/nbt.2509
Thomik T, Wittig I, Choe JY, Boles E, Oreb M (2017) An artificial transport metabolon facilitates improved substrate utilization in yeast. Nat Chem Biol 13(11):1158–1163. https://doi.org/10.1038/nchembio.2457
Huttanus HM, Feng X (2017) Compartmentalized metabolic engineering for biochemical and biofuel production. Biotechnol J. https://doi.org/10.1002/biot.201700052
Thodey K, Galanie S, Smolke CD (2014) A microbial biomanufacturing platform for natural and semisynthetic opioids. Nat Chem Biol. 10:837–844. https://doi.org/10.1038/nchembio.1613
Jacquier N, Schneiter R (2012) Mechanisms of sterol uptake and transport in yeast. J Steroid Biochem Mol Biol 129(1–2):70–78. https://doi.org/10.1016/j.jsbmb.2010.11.014
Acknowledgements
We thank Tiffany Chiu, Anqi Zhou, and Dr. Sheng Wu for their valuable feedback in the preparation of the manuscript. This work was supported by the National Science Foundation (grant to YL, 1748695) and Cancer Research Coordinating Committee Research Award (grant to YL, CRN-20-634571).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest regarding the publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Xu, S., Li, Y. Yeast as a promising heterologous host for steroid bioproduction. J Ind Microbiol Biotechnol 47, 829–843 (2020). https://doi.org/10.1007/s10295-020-02291-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10295-020-02291-7