
Abstract
The mechanical properties of cell nuclei have been demonstrated to play a fundamental role in cell movement across extracellular networks and micro-channels. In this work, we focus on a mathematical description of a cell entering a cylindrical channel composed of extracellular matrix. An energetic approach is derived in order to obtain a necessary condition for which cells enter cylindrical structures. The nucleus of the cell is treated either (i) as an elastic membrane surrounding a liquid droplet or (ii) as an incompressible elastic material with Neo-Hookean constitutive equation. The results obtained highlight the importance of the interplay between mechanical deformability of the nucleus and the capability of the cell to establish adhesive bonds and generate active forces in the cytoskeleton due to myosin action.








Similar content being viewed by others


References
Ambrosi D, Duperray A, Peschetola V, Verdier C (2009) Traction patterns of tumor cells. J Math Biol 58:163–181
Baumgartner W, Hinterdorfer P, Ness W, Raab A, Vestweber D, Schindler H, Drenckhahn D (2000) Cadherin interaction probed by atomic force microscopy. Proc Natl Acad Sci USA 97:4005–4010
Beadle C, Assanah MC, Monzo P, Vallee R, Rosenfeld SS, Canoll P (2008) The role of myosin II in glioma invasion of the brain. Mol Biol Cell 19:3357–3368
Bonet J, Wood RD (2008) Nonlinear continuum mechanics for finite element analysis. Cambridge University Press, Cambridge
Caille N, Thoumine O, Tardy Y, Meister JJ (2002) Contribution of the nucleus to the mechanical properties of endothelial cells. J Biomech 35(2):177–187
Capito RM, Spector M (2003) Scaffold-based articular cartilage repair. IEEE Eng Med Biol Mag 22:42–50
Chauvière A, Hillen T, Preziosi L (2007a) Modeling the motion of a cell population in the extracellular matrix. Discret Contin Dyn Syst (Supplements), Special Issue: 250–259
Chauvière A, Hillen T, Preziosi L (2007b) Modeling cell movement in anisotropic and heterogeneous network tissues. Netw Heterog Media 2(2):333–357
Chien S, Sung KLP, Skalak R, Usami S, Usami AC (1978) Theoretical and experimental studies on viscoelastic properties of erythrocyte membrane. Biophys J 24:463–487
Dembo M, Wang YL (1999) Stresses at the cell-to-substrate interface during locomotion of fibroblasts. Biophys J 76(4):2307–2316
Evans EA (1973) New membrane concept applied to the analysis of fluid shear and micropipette-deformed red blood cells. Biophys J 13:941–954
Evans EA, Waugh R, Melnik L (1976) Elastic area compressibility modulus of red cell membrane. Biophys J 16:585–595
Evans EA, Kukan B (1984) Passive material behavior of granulocytes based on large deformations and recovery after deformation tests. Blood 64:1028–1035
Evans EA, Yeung A (1989) Apparent viscosity and cortical tension of blood granulocytes determined by micropipet aspiration. Biophys J 56:151–160
Friedl P, Brocker EB (2000) The biology of cell locomotion within three-dimensional extracellular matrix. Cell Mol Life Sci 57(1):41–64
Friedl P, Wolf K (2003) Tumour-cell invasion and migration: diversity and escape mechanisms. Nat Rev Cancer 3(5):362–374
Friedl P, Weigelin B (2008) Interstitial leukocyte migration and immune function. Nat Immunol 9(9):960–969
Friedl P, Wolf K (2009) Proteolytic interstitial cell migration: a five-step process. Cancer Metastasis Rev 28(1–2):129–135
Friedl P, Wolf K, Lammerding J (2011) Nuclear mechanics during cell migration. Curr Opin Cell Biol 23(1):55–64
Gerlitz G, Bustin M (2011) The role of chromatin structure in cell migration. Trends Cell Biol 21(1):6–11
Graner F, Glazier JA (1992) Simulation of biological cell sorting using a two-dimensional extended potts model. Phys Rev Lett 69:2013–2016
Guck J, Lautenschläger F, Paschke S, Beil M (2010) Critical review: cellular mechanobiology and amoeboid migration. Integr Biol 2:575–583
Harley BAC, Kim H, Zaman MH, Yannas IV, Lauffenburger DA, Gibson LJ (2008) Microarchitecture of three-dimensional scaffolds influences cell migration behavior via junction interactions. Biophys J 95(8):4013–4024
Hawkins RJ, Piel M, Faure-Andre G, Lennon-Dumenil AM, Joanny JF, Prost J, Voituriez R (2009) Pushing off the walls: a mechanism of cell motility in confinement. Phys Rev Lett 102(5):058103
Helfrich W (1973) Elastic properties of lipid bilayers: theory and possible experiments. Z Naturforsch C 28(11):693–703
Hochmuth RM (1993) Measuring the mechanical properties of individual human blood cells. J Biomech Eng 115(4B):515–519
Hochmuth RM (2000) Micropipette aspiration of living cells. J Biomech 33:15–22
Houchmandzadeh B, Marko JF, Chatenay D, Libchaber A (1997) Elasticity and structure of eukaryote chromosomes studied by micromanipulation and micropipette aspiration. J Cell Biol 139(1):1–12
Ingber DE (1993) Cellular tensegrity: defining new rules of biological design that govern the cytoskeleton. J Cell Sci 104:613–627
Ingber DE (2003) Tensegrity I. Cell structure and hierarchical systems biology. J Cell Sci 116(7):1157–1173
Jones WR, Ting-Beall HP, Lee GM, Kelley SS, Hochmuth RM, Guilak F (1999) Alterations in the Young’s modulus and volumetric properties of chondrocytes isolated from normal and osteoarthritic human cartilage. J Biomech 32(2):119–127
Kole TP, Tseng Y, Jiang I, Katz JL, Wirtz D (2005) Intracellular mechanics of migrating broblasts. Mol Biol Cell 16:328–338
Kumar S, Weaver VM (2009) Mechanics, malignancy, and metastasis: the force journey of a tumor cell. Cancer Metastasis Rev 28:113–127
Kuntz RM, Saltzman WM (1997) Neutrophil motility in extracellular matrix gels: mesh size and adhesion affect speed of migration. Biophys J 72(3):1472–1480
Laadhari A, Misbah C, Saramito P (2010) On the equilibrium equation for a generalized biological membrane energy by using a shape optimization approach. Physica D 239:1567–1572
Landau LLD, Lifschitz EM (1986) Theory of elasticity, vol 7, 3rd edn. Course of Theoretical, Physics
Lämmermann T, Bader BL, Monkley SJ, Worbs T, Wedlich-Söldner R, Hirsch K, Keller M, Förster R, Critchley DR, Fässler R, Sixt M (2008) Rapid leukocyte migration by integrin-independent flowing and squeezing. Nature 453:51–55
Lautenschläger F, Paschke S, Schinkinger S, Bruel A, Beil M, Guck J (2009) The regulatory role of cell mechanics for migration of differentiating myeloid cells. Proc Natl Acad Sci USA 106(37):15696–15701
Legant WR, Miller JS, Blakely BL, Cohen DM, Genin GM, Chen CS (2010) Measurement of mechanical tractions exerted by cells in three-dimensional matrices. Nat Methods 7(12):969–971
Liu K (2006) Deformation behaviour of soft particles: a review. J Phys D Appl Phys 39:189–199
Lowengrub JS, Frieboes HB, Jin F, Chuang YL, Li X, Macklin P, Wise SM, Cristini V (2010) Nonlinear modelling of cancer: bridging the gap between cells and tumours. Nonlinearity 23:R1–R91
Painter KJ (2009) Modelling cell migration strategies in the extracellular matrix. J Math Biol 58:511–543
Panorchan P, Thompson MS, Davis KJ, Tseng Y, Konstantopoulos K, Wirtz D (2006) Single-molecule analysis of cadherin-mediated cell-cell adhesion. J Cell Sci 119:66–74
Peschetola V, Laurent V, Duperray A, Michel R, Ambrosi D, Preziosi L, Verdier C (2013) Time-dependent traction force microscopy for cancer cells as a measure of invasiveness. Cytoskeleton 70:201–214
Poelaert D, Schniewind J, Janssens F (2011) Surface area and curvature of the general ellipsoid. arXiv:1104.5145 [math.CA]
Preziosi L, Vitale G (2011) Mechanical aspects of tumour growth: multiphase modelling, adhesion, and evolving natural configurations. In: Ben Amar M, Goriely A, Mller MM, Cugliandolo LF (eds) New trends in the physics and mechanics of biological systems, Lecture Notes of the Les Houches Summer School, vol 92. Oxford University Press, Oxford, pp 177–228
Renkawitz J, Sixt M (2010) Mechanisms of force generation and force transmission during interstitial leukocyte migration. EMBO Rep 11(10):744–750
Rolli CG, Seufferlein T, Kemkemer R, Spatz JP (2010) Impact of tumor cell cytoskeleton organization on invasiveness and migration: a microchannel-based approach. PLos ONE 5(1):e8726
Rowat AC, Lammerding J, Ipsen JH (2006) Mechanical properties of the cell nucleus and the effect of emerin deficiency. Biophys J 91(12):46494664
Sabeh F, Shimizu-Hirota R, Weiss SJ (2009) Protease-dependent versus -independent cancer cell invasion programs: three-dimensional amoeboid movement revisited. J Cell Biol 185(1):11–19
Sahai E (2007) Illuminating the metastatic process. Nat Rev Cancer 7(10):737–749
Scianna M, Preziosi L, Wolf K (2013) A cellular potts model: simulating cell-and extracellular matrix-derived determinants for cell migration on and in matrix environments. Math Biosci Eng 10(1):235261
Scianna M, Preziosi L (2013) Modelling the influence of nucleus elasticity on cell invasion in fiber networks and microchannels. J Theor Biol 317(21):394–406
Skalak R (1973) Modelling the mechanical behavior of red blood cells. Biorheology 19:229–238
Skalak R, Tozeren A, Zarda RP, Chien S (1973) Strain energy function of red blood cell membrane. Biophys J 13:245–264
Sun M, Graham J, Hegedus B, Marga F, Zhang Y, Forgacs G, Grandbois M (2005) Multiple membrane tethers probed by atomic force microscopy. Biophys J 89:4320–4329
Theret DP, Lvesque ML, Sato M, Nerem RM, Wheeler LT (1988) The application of a homogeneous half-space model in the analysis of endothelial cell micropipette measurements. J Biomech Eng 110:190–199
Tu ZC, Ou-Yang ZC (2004) Geometric theory on the elasticity of bio-membranes. J Phys A Math Gen 37:11407–11429
Tu ZC, Ou-Yang ZC (2008) Elastic theory of low-dimensional continua and its applications in bio- and nano-structures. J Comput Theor Nanosci 5:422–448
Valerius NH, Stendahl O, Hardwig JH, Stossel TP (1981) Distribution of actin-binding protein and myosin in polymorphonuclear leukocytes during locomotion and phagocytosis. Cell 24:195–202
Vaziri A, Mofrad MR (2007) Mechanics and deformation of the nucleus in micropipette aspiration experiment. J Biomech 40(9):2053–2062
Verdier C, Etienne J, Duperray A, Preziosi L (2009) Review: rheological properties of biological materials. C R Phys 10(8):790–811
Vitale G, Preziosi L, Ambrosi D (2012) A numerical method for the inverse problem of cell traction in 3D. Inverse Prob 28:095013
Versaevel M, Grevesse T, Gabriele S (2012) Spatial coordination between cell and nuclear shape within micropatterned endothelial cells. Nat Commun 3. doi:10.1038/ncomms1668
Wang JH-C, Lin J-S (2007) Cell traction force and measurement methods. Biomech Model Mechanobiol 6:361–371
Waugh R, Evans EA (1979) Thermoelasticity of red blood cell membrane. Biophys J 26:115–132
Wolf K, Wu YI, Liu Y, Geiger J, Tam E, Overall C, Stack MS, Friedl P (2007) Multi-step pericellular proteolysis controls the transition from individual to collective cancer cell invasion. Nat Cell Biol 9:893–904
Wolf K, Alexander S, Schacht V, Coussens LM, von Andrian UH, van Rheenen J, Deryugina E, Friedl P (2009) Collagen-based cell migration models in vitro and in vivo. Semin Cell Dev Biol 20(8):931–941
Wolf K, Friedl P (2011) Extracellular matrix determinants of proteolytic and non-proteolytic cell migration. Trends Cell Biol 21(12):736–744
Yannas IV, Lee E, Orgill DP, Skrabut EM, Murphy GF (1989) Synthesis and characterization of a model extracellular matrix that induces partial regeneration of adult mammalian skin. Proc Natl Acad Sci USA 86:933–937
Yeung A, Evans E (1989) Cortical shell-liquid core model for passive flow of liquid-like spherical cells into micropipets. Biophys J 56:139–149
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1: Micropipette models applied to cell migration inside channels
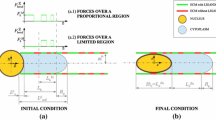
For sake of completeness, we consider here the case in which nucleus entry obeys the classical relations (2.2) or (2.5), deforming the initial spherical nucleus into a cigar-like shape, with the assumption that \(L_p\) in (2.2) and (2.5) represents the length of the deformed nucleus, i.e., \(L_p=L_{n,\mathrm{cigar}}^{\mathrm{fin}}=2(h+R_p)\) with \(h\) given by Eq. (4.3). We define the critical pressure as the value of \(\Delta P\) for which \(L_p=L_{n,\mathrm{cigar}}^{\mathrm{fin}}\) and we assume that a proper representation for \(\Delta P\) in Eqs. (2.2) and (2.5) is \(\dfrac{F_{\mathrm{active}}^Z}{\pi R_p^2}\), where \(F_{\mathrm{active}}^Z\) is the \(Z\)-component of the active force given either by Eq. (3.8) or (3.9) or (3.10). Then, assuming that a pressure above the critical one makes the cell move inside the pipette, it is possible to obtain the relation between mechanical and active properties that should hold for the cell to enter the channel, depending on the geometrical properties (i.e., \(R_{n},R_{c}\) and \(R_{p}\)).
The inequalities that should be satisfied in each case are summarized in Table 2, as a function of the diameter ratio \(\tilde{R}_p=\dfrac{R_p}{R_n}\). On the left-hand side of each relation, we have characteristic parameters representing the ratio between active properties and mechanical properties of cell nucleus. In particular, we identify
On the right-hand side of each relation, we have the critical value of the characteristic number (indicated with \(\overline{G}_{i}^{j}\), with \(i=\left\{ \gamma , E \right\} ,j= \left\{ k,F \right\} \)), which is a function of the diameter ratio, and we have set
with \(\tilde{L}_n^0=1-\sqrt{1-\tilde{R}_p^2}\) and \(\tilde{R}_c=R_c/R_n\).
The critical characteristic numbers are plotted in Fig. 9 as a function of the diameter ratio of the channel. The graphs represent the minimum value that each constant should assume in order to have the cell totally inside the channel, according to Chien’s criterion (Fig. 9a) and Theret’s one (Fig. 9b). Results obtained with the linearized Chien’s equation (2.4) are comparable with the ones obtained with the more complex formula (2.2). In Fig. 9, the dashed line represents results obtained using constant forces over a bounded domain (where we set \(\tilde{L}_b^M=5\)). It is possible to see that for big \(\tilde{R}_p,\overline{G}_{\gamma }^F\) and \(\overline{G}_{E}^F\) are obviously not influenced by the assumption on the boundedness of the contact region in which integrins are expressed (i.e., the red-dashed curve and the black-solid one overlap). Indeed, it exists an \(\tilde{R}_p^*\) such that \(L_b^*=L_b\) for \(\tilde{R}_p \ge \tilde{R}_p^*\), whereas \(L_b^*=L_b^M\) for \(\tilde{R}_p < \tilde{R}_p^*\). Therefore, the active work is influenced by the boundedness assumption only for \(\tilde{R}_p < \tilde{R}_p^*\).

Critical value of the characteristic numbers obtained applying a Chien’s model and b Theret’s model, under either a linear force (blue) or a constant force (black) or a constant force over a bounded region (red dashed) assumption
For instance, Fig. 10 explains how these graphs can be interpreted (for the particular case of Chien model): The bar charts below the graph represent the range of \(\tilde{R}_p\) for which a cell characterized by either a given \(G_{\gamma }^{k}\) or a given \(G_{\gamma }^{F}\) can enter the channel.

Interpretation of the results: bar charts represents the range for which a cell, with a given \(G_{\gamma }^{k}\) or \(G_{\gamma }^{F}\) can enter the channel, for the different hypotheses of bond forces
In the figure, ‘cell 1’ (orange) is characterized by higher \(G_{\gamma }^{k}\) or \(G_{\gamma }^{F}\) than ‘cell 2’ (violet). This means that we are considering either a softer cell (i.e., smaller \(\gamma \)) or a cell that is able to establish a higher number of adhesive bonds (i.e., higher \(\rho _b \alpha _{\mathrm{ECM}}\)) or a cell with better contractile capabilities (i.e., bigger \(k_b\) or \(F_b^M\)). In any case, the range for which ‘cell 1’ can enter the pipette is bigger than for ‘cell 2’ (orange bars vs. violet bars), according to what we expect from biological observations. Moreover, using the constant force assumption, it is possible to see that the range for which cells can enter the pipette is bounded both from below and from above. On the other hand, using the linear force assumption, we do not have any inferior limit, in contrast with biological observation. This contradictory result is due to the hypothesis used in the representation of forces. Indeed, in this case, the more the cytoplasm of the cell spreads inside the channel (small \(\tilde{R}_p\)), the more the traction force (which is related to the adhesive region) can pull the nucleus inside. In particular, even though the force required to deform the nucleus grows as \(\tilde{R}_p^{-3}\), as \(\tilde{R}_p \rightarrow 0\), the adhesive-dependent active force raises faster, since \(\tilde{L}_b^2= \mathcal O \left( \tilde{R}_p^{-4}\right) \). On the other hand, when a constant force assumption is used, for small \(\tilde{R}_p\), the length for which bonds are formed augments \( \left( \tilde{L}_b= \mathcal O \left( \tilde{R}_p^{-2}\right) \, \text{ for } \, \tilde{R}_p \rightarrow 0 \right) \). Thus, the total integrin-dependent traction force exerted on the nucleus increases, but it is not sufficient to compensate the greater deformation required by the nucleus, which goes like \(\tilde{R}_p^{-3} \, \text{ for } \, \tilde{R}_p \rightarrow 0 \). Conversely, introducing the boundedness assumption on \(L_b\), the active force is limited.
In particular, we have that for \(\tilde{R}_p \rightarrow 0, \overline{G}^{\gamma }_{F}\) goes like \(\tilde{R}_p^{-\alpha }\) (with \(\alpha =1\) for unbounded \(L_b\) and \(\alpha =3\) when the adhesive region is limited) and \(\overline{G}^{\gamma }_{k}\) grows linearly.
On the other hand, when the radius of the pipette is very big, the entry of the cell into the channel is limited due to the decrease in the contact area between the cell and the channel wall, where adhesive bonds are formed. It is likely that, in this case, the force exerted by actomyosin is not equal to the maximum executable force. Thus, a linear force can better describe the physiological behaviour. Therefore, a good choice for the bond force relation could be a ramp force on a bounded adhesive region, which is also the most conservative case.
In Theret’s model, it is possible to see that, for \(\tilde{R}_p \rightarrow 0, \overline{G}_{E}^{k}= \mathcal O \left( \tilde{R}_p^2\right) \) and \(\overline{G}_{E}^{F}= \mathcal O \left( 1 \right) \) when the constant force assumption with unbounded adhesive region is implemented. Thus, neither the constant force assumption nor the linear force one can account for the inferior limit in pipette calibres. Only enforcing the boundedness of the adhesive region, the capability of cells to enter very small channels is prevented.
Both Chien’s and Theret’s models, with the assumption of active forces over a bounded region, provide evidence for a biphasic cell migratory behaviour that reveals most optimal migration at pore sizes at nuclear and subnuclear diameters and diminishes at gaps greatly bigger or smaller than the cell nucleus diameter.
However, even though the results obtained by applying the classical models above seem promising, especially when adhesion is active on a bounded domain, they cannot account for the finite boundaries of the nucleus. Indeed, Chien’s model refers to an infinite 2D membrane, whereas Theret’s one was derived for a 3D half-space aspired inside a pipette, only for a small portion. Therefore, these criteria cannot be applied to describe the total entry of the cell into a pipette. The consequence of this assumption is evident in Fig. 9, where, for \(\tilde{R}_p=1\), the force needed to deform the nucleus does not vanish.
Appendix 2: Influence of bending
In Sect. 4.2, we considered only the contribution to the surface deformation energy due to the stretching of the nuclear membrane, but we disregarded the energy contribution associated with bending. In order to introduce in the model the bending of the nuclear membrane, we refer to Helfrich’s work on lipid bilayers (Helfrich 1973). Helfrich introduced a model (Helfrich 1973) in which the bending energy of a membrane is given by
where \(H=\dfrac{1}{2} (\textit{k}_1+ \textit{k}_2)\) is the mean curvature of the membrane surface, \(S,\textit{k}_1\) and \(\textit{k}_2\) are the principal curvatures, \(K= \textit{k}_1 \textit{k}_2\) is the Gaussian curvature, and \(c_0\) represents the spontaneous curvature that describes the asymmetric effect of the membrane. We remark that, from the Gauss-Bonnet theorem, the second term in the Helfrich energy is a topological invariant, and thus, in this work, it can be omitted (Laadhari et al. 2010). The total energy of deformation of the nuclear membrane, considering both the contribution due to stretching and the one due to bending, is Landau and Lifschitz (1986)
where \(\mathcal W _{\mathrm{bending}}^{\mathrm{sphere}}= 8 \pi k_c\). For the cigar-shaped deformed configuration, we define
whereas, for a nucleus deformed into an ellipsoid, we have
where we referred to Poelaert et al. (2011), in which the principal curvatures of the general ellipsoid are derived, i.e.,
and \(h_e=R_n \dfrac{R_n^2}{R_p^2}\) is defined in (4.2). The integral (6.4) is solved numerically.
Once \(\mathcal W _{\mathrm{tot}}^S\) is known, it is possible to obtain an expression for the dimensionless parameters \(G_{\lambda }^{k}\) and \(G_{\lambda }^{F}\). The results are reported in Table 3, where \( \tilde{k}_c= \dfrac{k_c}{\lambda R_n^4}, Q(R_p)= \int _0^{\pi } (2 H )^2 \mathcal A \hbox {d} \Theta , G_{\lambda }^{k}\) and \(G_{\lambda }^{F}\) have the same definition given in Sect. 5.
The results are reported in Fig. 11 for the cigar-shaped nucleus and in Fig. 12 for the ellipsoidal nucleus. In order to observe the bending contribution, the ratio \(k_c/\lambda \) should be larger than \(10\mu \hbox {m}^4\). However, from Helfrich (1973), we, know that for lipid bilayer membrane of a nucleus of radius \(4\mu \)m, we have \(k_c/\lambda = 4 \cdot 10^{-5} \mu \hbox {m}^4 \). Thus, under the assumption that the surface of the nucleus increases, the major contribution is due to stretching.

Critical value of \(G_{\lambda }^k\) and \(G_{\lambda }^F\) for different values of \(k_c\), assuming a cigar-shaped deformation

Critical value of \(G_{\lambda }^k\) and \(G_{\lambda }^F\) for different values of \(k_c\), assuming an ellipsoidal deformation
Rights and permissions
About this article
Cite this article
Giverso, C., Grillo, A. & Preziosi, L. Influence of nucleus deformability on cell entry into cylindrical structures. Biomech Model Mechanobiol 13, 481–502 (2014). https://doi.org/10.1007/s10237-013-0510-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10237-013-0510-3