
Abstract
Differences in litter decomposition patterns among mesic, semiarid, and arid grassland ecosystems cannot be accurately explained by variation in temperature, moisture, and litter chemistry alone. We hypothesized that ultraviolet (UV) radiation enhances decomposition in grassland ecosystems via photodegradation, more so in arid compared to mesic ecosystems, and in litter that is more recalcitrant to microbial decomposition (with high compared to low lignin concentrations). In a 2-year field study, we manipulated the amount of UV radiation reaching the litter layer at three grassland sites in Minnesota, Colorado, and New Mexico, USA, that represented mesic, semiarid, and arid grassland ecosystems, respectively. Two common grass leaf litter types of contrasting lignin:N were placed at each site under screens that either passed all solar radiation wavelengths or passed all but UV wavelengths. Decomposition was generally faster when litter was exposed to UV radiation across all three sites. In contrast to our hypothesis, the contribution of photodegradation in the decomposition process was not consistently greater at the more arid sites or for litter with higher lignin content. Additionally, at the most arid site, exposure to UV radiation could not explain decomposition rates that were faster than expected given climate constraints or lack of N immobilization by decomposing litter. Although photodegradation plays an important role in the decomposition process in a wider range of grassland sites than previously documented, it does not fully explain the differences in decomposition rates among grassland ecosystems of contrasting aridity.
Similar content being viewed by others


Introduction
Decomposition rates in mesic grassland ecosystems are well described by established decomposition models based on temperature, moisture, and litter chemistry, but these models tend to underestimate rates in arid ecosystems (Meentemeyer 1978; Schaefer and others 1985; Whitford and others 1981). Several hypotheses have been proposed to explain faster than predicted litter decomposition in arid ecosystems, including precipitation pulses (Austin and others 2004), consumption by arthropods (Santos and Whitford 1981), and physical abrasion and burial by soil (Throop and Archer 2007). However, tests of these hypotheses have shown that they do not fully explain why decomposition rates are more rapid than expected in arid ecosystems (MacKay and others 1994; Schaefer and others 1985; Whitford and others 1986).
Greater photodegradation in arid compared with mesic grasslands could account for decomposition rates being more rapid in arid grasslands than expected based on climate (Parton and others 2007; Adair and others 2008). Indeed, a growing number of studies in arid and semiarid ecosystems have shown that photodegradation by ultraviolet (UV) radiation (280–400 nm) contributes significantly to the decomposition of surface litter (Austin and Vivanco 2006; Day and others 2007; Brandt and others 2007; Gallo and others 2009). However, whether the contribution of photodegradation to decomposition is in fact greater in arid than in mesic systems has not been tested. A study manipulating precipitation and UV radiation in a semiarid grassland showed that photodegradation increased decomposition rates under dry conditions but not under wet conditions (Brandt and others 2007). Although this study suggests a greater role for photodegradation when conditions are dry, other site-specific conditions besides photodegradation also could contribute to differences between arid and mesic systems, including timing of precipitation, soil type, microbial community structure, and plant canopy cover.
A greater role of photodegradation in arid than mesic systems could explain the lack of a negative correlation between lignin content and mass loss often observed in arid systems (Whitford and others 1981; Moorhead and Callaghan 1994). Lignin absorbs UV radiation, and several studies have shown that exposure to UV radiation increases litter lignin loss (Day and others 2007; Henry and others 2008; Austin and Ballaré 2010). If lignin is the primary carbon fraction that is photodegraded, plant litter with higher lignin content could be more susceptible to photodegradation than low-lignin litter. However, only a few studies manipulating UV radiation have used more than one litter type (Gallo and others 2006; Brandt and others 2007). Although some studies suggest that plant litter chemistry may be an important factor influencing photodegradation rates (Brandt and others 2007), others suggest that litter surface area is more important than chemistry (Gallo and others 2006; Brandt and others 2009).
A greater role of photodegradation in arid than mesic systems is also consistent with differences in litter microbial processes observed between these ecosystems. For example, in contrast to mesic ecosystems, decomposing surface litter in some arid ecosystems does not immobilize nitrogen, and decomposition rates are unrelated to initial N content (Parton and others 2007; Vanderbilt and others 2008; Gallo and others 2009). These patterns suggest that abiotic processes rather than microbial activity are the major drivers of decomposition (Moorhead and Reynolds 1989). If abiotic processes dominate, one would expect to find lower microbial enzyme activity relative to litter that is primarily decomposed by microbes. Alternatively, enzyme activity could increase if photodegradation increases the number of effective binding sites by disrupting the lignocellulose matrix (Gallo and others 2006). However, the potential for increased binding is probably limited because exposure to high levels of UV radiation tends to decrease microbial activity and abundance (Gehrke and others 1995; Duguay and Klironomos 2000; Pancotto and others 2003; Belnap and others 2008).
Consistent with greater photodegradation, an abiotic process, litter decay patterns in arid ecosystems often exhibit linear mass loss over time (Austin and Vivanco 2006; Adair and others 2008; Vanderbilt and others 2008), rather than the exponential decay pattern usually observed for decomposing litter (Olson 1963). An exponential decay model implicitly assumes that microbial metabolism is the principal driver of mass loss, with mass loss proportional to remaining mass. If photodegradation is the principal driver of decomposition, mass loss should be proportional to exposed surface area, rather than to mass remaining, perhaps explaining linear litter decay patterns observed in many decomposition studies in arid ecosystems (Austin and Vivanco 2006; Adair and others 2008; Vanderbilt and others 2008).
In summary, a number of indirect lines of evidence suggest that photodegradation plays a greater role in decomposition in arid than in mesic ecosystems, but this idea has never been tested directly across sites that vary in climatic regime. We designed a 2-year field litter decomposition experiment at three grassland sites that comprise a gradient in UV radiation, precipitation, and temperature. At each site, we manipulated UV radiation using specially designed screens while allowing other site-specific decomposition processes to occur. For the purposes of our study, we defined photodegradation as the net positive direct effects of photolysis and photo-oxidation of organic matter and any indirect positive effects on litter decomposition through changes in litter chemistry that facilitate leaching, fragmentation, or microbial decomposition minus any indirect negative effects of UV radiation on the decomposer community.
We evaluated the following specific hypotheses: (1) photodegradation will play a larger role in the decomposition process in arid systems than mesic systems; (2) in sites where photodegradation dominates, photodegradation will be greater for high lignin than for low-lignin litter because of preferential loss of the lignin fraction in the litter; (3) as the role of photodegradation increases, litter decay will be increasingly decoupled from N immobilization and extracellular enzyme activity; and (4) as the role of photodegradation increases, mass loss will follow a more linear instead of exponential decay pattern.
Methods
Study Sites
Cedar Creek, Minnesota; Central Plains, Colorado; and Sevilleta, New Mexico represent mesic, semiarid, and arid grasslands, respectively (Table 1), that are part of the U.S. Long-Term Ecological Research (LTER) Network. Sevilleta receives the highest dose of UV radiation (roughly 50% more than Central Plains and 100% more than Cedar Creek, Figure 1C). On average, Cedar Creek receives roughly twice as much annual precipitation as Central Plains and roughly three times as much as Sevilleta (Figure 1B). Sevilleta receives 60% of its precipitation as summer monsoon rains (June–September), with the rest as winter storm fronts. Precipitation patterns at Cedar Creek and Central Plains are not driven by the monsoons, and both sites have relatively low precipitation from November–March and higher precipitation during the growing season (April–October). The sites also differ in mean daily temperature (Figure 1A).

Climate characteristics during the experiment. A Mean daily air temperature by month for each site. B Total monthly precipitation (left y-axis) and cumulative monthly precipitation (right y-axis) for each site. C Mean monthly above-canopy daily erythemal UV-B dose (YES UVB-1 broadband sensor) from the nearest USDA UV-B monitoring station (ftp://uvb.nrel.colostate.edu/UVB). Minnesota: 47.2°N, 93.5°W, 424 m elev.; Colorado: 40.8°N, 104.8°W, 1641 m elev.; New Mexico: 32.6°N, 106.74°W, 1317 m elev. Arrows indicate litterbag collection times (S3 = collection time 3 at Sevilleta).
At all three sites, plots were set up in a 2800-m2 area dominated by grasses. At Cedar Creek, plots were located in a mid-successional old field site dominated by the C3 and C4 grasses and forbs. Poa pratensis (C3 grass) is the dominant grass species, with the C4 grasses Schizachyrium scoparium, Andropogon gerardii, and Eragrostis spectablilis making up a large component. Common forbs are Lychnis alba, Achillea millefolium, and Artemisia ludoviciana. Vegetation cover is relatively complete and uniform with an underlying contiguous litter layer. At Central Plains, plots were located in a shortgrass steppe loamy upland community, dominated by the C4 perennial short grass Bouteloua gracilis. Opuntia polyacantha (low cactus) and Sphaeralcea coccinea (C3 forb) are important components. Plots at Sevilleta were established in Chihuahuan desert grassland, dominated by the two C4 short grasses Bouteloua eriopoda and B. gracilis. Both Central Plains and Sevilleta have patchy vegetative cover, with roughly 25% exposed mineral soil.
Experimental Manipulation of UV Radiation
We experimentally manipulated UV radiation (280–400 nm) over the litter layer at each of the three sites using ten pairs of 75 cm × 150 cm × 20 cm (l × w × h) UV-blocking and UV-passing plastic screens in a randomized complete block design. UV-transparent acrylic (UV pass, which passes 90% of the solar spectrum, including UV-A and UV-B, Solacryl SUVT®, Spartech Polycast, Stamford, Connecticut, USA) or polycarbonate (hereafter UV block, which eliminates 90% of UV-A and UV-B, optically equivalent to Lexan XL-1®, GE, Pittsfield, Massachusetts, USA) was used because it effectively passed or blocked UV radiation without substantially affecting temperature or photosynthetically active radiation (PAR, 400–700 nm, Brandt and others 2007). Although PAR has been demonstrated to play a role in photodegradation (Austin and Vivanco 2006; Brandt and others 2009), we chose not to manipulate PAR because it would have confounding effects on vegetation cover and microclimate. Screens were custom designed and constructed for this study in a louvered design to allow for penetration of precipitation to the litter layer and to avoid excessive heating while still blocking UV except in early morning and late evening when the atmosphere filters a relatively greater proportion of UV than PAR (Figure 2). Louvers (23 per screen, each 6.35 cm wide) were fixed at an angle of 60°. The bottom and top of adjacent louvers lined up vertically but did not overlap. Louvers were replaced on an as-needed basis if they became damaged (it was determined in a previous experiment that UV and PAR transmission properties did not change over a 2-year period, Brandt and others 2007). Use of a central 40 cm × 110 cm area under the screens minimized edge effects.

UV treatment screen design. Frames were custom designed from galvanized steel with removable plastic louvers that either blocked or passed UV radiation. Screens were oriented southward to achieve the greatest possible UV treatment effect. Also shown: litterbags attached to the ground with sod staples; radiometers for measuring solar radiation.
We placed small, data-logging temperature sensors (I-button, Dallas Semiconductor) underneath the litter in filled mock litterbags (n = 3 per treatment) containing A. gerardii litter to record temperature once per hour for 1 year. There was no significant difference in daily mean, minimum, or maximum temperatures between UV treatments. On average, both UV block and UV pass screened plots had 2°C lower daily minimum and 1°C lower mean temperatures than unscreened plots in the winter when covered with snow, and 1°C higher daily minimum temperatures and 0.5°C higher daily mean temperatures when not covered with snow (screen*season: P < 0.0001). There was no significant effect of the screens on daily maximum temperatures at any time of year.
To ensure that our UV treatment was effective, we spot-checked UV-A, UV-B, and PAR at various times of day (usually the 5 h leading up to and including solar noon, depending on season) an average of once per month for 6 months (June–December) at all three sites. We used a UV radiometer (UV-X, UV Products, Upland, California, USA) with separate sensors for UV-A and UV-B. The UV-B sensor (UV-X 31) was calibrated and had a maximum peak at 310 nm with a spectral response curve encompassing 260–370 nm. The UV-A sensor (UV-X 36) was calibrated at 365 nm and had a maximum peak at 360 nm with a spectral response curve encompassing 300–400 nm. Measurements were compared to readings taken outside of the experimental units. UV-A and UV-B were reduced by an average of 86 and 74%, respectively (SE = 1%) under the UV block screens, and by 13 and 14% (SE = 1%) under the UV pass screens. We measured PAR using a quantum sensor calibrated to natural sunlight (Apogee Instruments, Logan, Utah, USA). The UV block treatment received 7% lower PAR (400–700 nm) than the UV pass treatment because of slight differences in transmission properties of the plastic materials in the 400–500 nm range (Brandt and others 2007).
Litter Collection
We compared the decomposition of two C4 grass species that differ in initial litter chemistry (Table 2). A. gerardii (8.1% lignin) is a C4 tall grass and is a significant proportion of plant cover at Cedar Creek. It is not present at the other two sites but is present in the region. B. gracilis (6.6% lignin) is a C4 short grass and is one of the primary species found at Sevilleta and Central Plains. It is not present in the study area at Cedar Creek, although it is present in the region. Litter was collected in late October 2005 following senescence from planted monocultures located in Princeton, Minnesota, USA (45.61°N, 93.58°W), a site that was not part of the experiment. Litter was oven dried at 35°C. Random grab samples (n = 10) of each species were collected for analysis of initial litter chemistry after drying at 55°C.
Litterbag Preparation
Ten grams of dry litter was placed in 1.5-mm aluminum mesh litterbags (15 cm × 15 cm). Aluminum mesh was chosen instead of the more commonly used fiberglass screen material because aluminum fibers are thinner and thus leave more litter exposed to ambient solar radiation (~70 vs. 50% area exposed). Prior to placing litterbags, the ground beneath each experimental unit was manually cleared of vegetation by clipping. Periodic clipping to prevent shading of litterbags continued for the duration of the experiment (with the exception of the canopy treatment described below). Litterbags were deployed at each site between April 22 and May 9, 2006. Litterbags were all placed in the field on the same day within each site, but were staggered by several weeks among sites for logistical reasons. Eight litterbags were placed beneath each experimental unit in a completely randomized design and attached to the ground in two corners with metal sod staples (Figure 2).
Litterbag Collection
Litterbags were collected four times: approximately 2.5, 6, 12, and 24 months after deployment (in essence, summer 2006, fall 2006, summer 2007, and spring 2008). We collected litterbags at all three sites within 3 weeks of each other, with the exception of the third collection date, when Sevilleta was collected in May, and the other two sites were collected in July. Each collected litterbag was sealed in a plastic bag and refrigerated prior to processing. Visible soil, plants, or arthropods were separated from the litter prior to weighing. If litterbags had a quantifiable amount of soil accumulation in the bag, we saved, dried (55°C), and weighed the soil that was removed from each bag. The litter was cut into 1-cm pieces, mixed, and sub-divided for the following analyses: ash content, C and N composition, fiber fraction analysis, and extracellular enzyme assays. Subsamples used for enzyme assays (0.5 g) were placed in air-tight plastic bags and kept frozen at −20°C prior to analysis. The remainder of the litter was oven-dried at 55°C and re-weighed for gravimetric moisture. We then used gravimetric moisture to correct the field-moist sample weight and calculate final dry mass remaining. Subsamples of litter from each bag were ashed at 600°C to calculate ash-free dry mass.
Chemical Analysis
At our central lab at the University of Minnesota, we analyzed litter composition (cell solubles, hemicellulose, cellulose, and lignin) using the forage fiber technique (Van Soest 1967). Subsamples (0.5 g) were ground through a Wiley Mill and subjected to sequential neutral detergent fiber, acid detergent fiber, and sulfuric acid (acid detergent lignin) digestions using an ANKOM fiber analyzer (ANKOM Technology, Macedon, New York, USA). Samples were ash-corrected after the final acid digestion to account for any contamination of mineral soil in the lignin fraction. Percent remaining of each fiber fraction was calculated with respect to the average initial value for each litter type (Table 2).
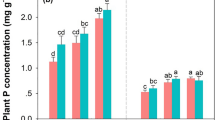
In addition, initial grab samples and subsamples of each litterbag were ground to a powder using a ball mill and then weighed into tin capsules for C and N analysis (Elementar, Mt. Laurel, New Jersey, USA). Nitrogen immobilization and release were calculated as the change in N content in the litter relative to initial values on an ash-free dry (55°C) mass basis. Initial litter samples were also analyzed for P, K, and Fe using inductively coupled plasma spectroscopy (Perkin Elmer, Waltham, Massachusetts, USA, Table 2).
Enzyme Assays
We assayed for activities of six extracellular enzymes using standard protocols (Saiya-Cork and others 2002; Sinsabaugh and others 2008): β-1,4-glucosidase and cellobiohydrolase, which mediate cellulolysis; phenol oxidase and peroxidase, which oxidize lignin and other phenolic compounds; acid phosphatase, which hydrolyzes phosphate from phospholipids and phosphosaccharides; and n-acetyl-glucosaminidase, which hydrolyzes aminosaccharides from chitin. Assay plates (96-well) were incubated at room temperature for 0.5–20 h, depending on the assay. Activity was calculated in μmol h−1 g−1 dry litter.
Canopy Manipulation Experiment
Because plant canopy cover attenuates solar radiation input to the litter layer especially at Cedar Creek, we included a supplemental plant canopy treatment in addition to the UV treatment at that site. Ten UV block and ten UV pass units were added. A canopy treatment (unclipped) was performed by allowing natural vegetation to grow up through the litterbags. The non-canopy treatment (clipped) was performed by hand-clipping as described above, but plots were clipped every 2 weeks throughout the growing season (April–October). Litterbags were placed in the field, collected, and processed at the same time and in the same manner as those described above.
We tested the effectiveness of the canopy treatment by measuring leaf area index (LAI) on July 6, 2006 (near peak standing biomass) using a Li-Cor LAI-2000 sensor. We measured LAI in clipped and unclipped reference plots (n = 4 per treatment). Measurements were taken at dusk on a clear sunny day using the 90° view cap on the sensor in the northeast, southeast, southwest, and northwest corners of each plot, facing inward. The above-canopy reference was taken by holding the sensor directly over the top of the canopy. At peak standing biomass, LAI of the unclipped plots averaged 0.50 m2 m−2 (SE = 0.10), whereas LAI of the clipped plots averaged 0.03 m2 m−2 (SE = 0.005).
Clipped and unclipped plots were also monitored for temperature and solar radiation as described above. Unclipped plots received an average of 17% lower PAR and UV than clipped plots (P < 0.0001). Litter moisture in collected litterbags was an average of 24% higher in the unclipped plots than clipped plots, but the difference was only statistically significant for the third collection date (time*clipping: P = 0.0016). Clipped plots had daily maximum temperatures that were on average 3°C lower than unclipped plots from May through July (month*clipping: P < 0.0001), which may be attributed to differences in albedo or air circulation.
Data Analysis
Statistical analyses were conducted using JMP software (SAS Institute, Cary, North Carolina). Percent ash-free dry mass remaining, litter N content (% of initial), litter moisture, enzyme activities, and carbon fractions were evaluated using a nested split-plot mixed-effects ANOVA with site as a fixed effect and UV, species, collection time, and their interactions as fixed effects nested within sites. Block and the block × UV interaction were included as random effects. We also analyzed each site separately using the same model (excluding the site effect). We evaluated each effect using a full-factorial model and then simplified the model if interactions were not significant and the simpler model was a better fit according to Akaike’s information criterion (AIC). Data were square root transformed as necessary to improve normality, and models were run with and without outliers.
Decay constants (k) were calculated by fitting ash-free dry mass remaining to single pool exponential decay and linear models using non-linear and linear least-squares fitting procedures, respectively. For the exponential model, mass remaining was fit to the model X/X 0 = e −kt, where X is mass remaining at time t, X 0 is the initial mass, and k is the litter-specific decay constant. For the linear model, mass remaining was fit to the model X/X 0 = −kt + c, where c is a constant. Litterbags of the same species within each plot were treated as replicates (n = 10 for each factorial combination of species, UV treatment, and site). We compared linear and single exponential models using AIC with a second-order correction (AICc) for small sample sizes (Burnham and Anderson 2002). When the difference between the two AICc values (delta r) was greater than 3, the model with the lowest AICc was considered the better fit. If the same model was identified for two or more data sets, their decay constants were then compared using the same mixed-effects model as above, but eliminating time as a factor.
We plotted the N content relative to initial values (% of initial N) versus percent ash-free dry mass remaining to compare results of this study to relationships between N and litter mass loss modeled by Parton and others (2007). For each species, we calculated the predicted percent of initial N based on models derived for mesic and dry grasslands (Parton and others 2007, supporting online material). The mesic grassland model includes initial litter N concentration as a parameter, and therefore differs between A. gerardii and B. gracilis. The dry grassland model does not include initial N concentration as a parameter, and therefore is the same for the two species. We also included a 1:1 linear model (% mass remaining = % of initial N) as a third potential model, which assumes that N and total litter mass are lost at the same rate. We then compared the three models (mesic, dry, and linear) using AICc as described above. We hypothesized that litter would follow the dry grassland model or 1:1 linear model when UV radiation plays a significant role in litter mass loss and would follow the mesic grassland model when litter mass loss is driven by microbial processes.
Results
Litter Decay Models
The linear and single exponential models fit equally well the litter mass loss data (delta r < 3) in the majority of treatment–species–replicate (sample) combinations (76%). In the remaining 24% of cases, a single exponential model was a better fit 88% of the time. Cedar Creek had more samples that fit a single exponential model than the other two sites. There was no trend in model performance between UV treatments or species. For simplicity, the single exponential model was chosen, and the decay constant (k) was used to compare among sites and between treatments. Cedar Creek model fits had the highest average r 2 value of 0.96 (range = 0.75–0.99, SE = 0.003). The mean r 2 values for Central Plains and Sevilleta were 0.79 (range = 0.37–0.99, SE = 0.02) and 0.77 (range = 0.26–0.99, SE = 0.01), respectively.
Effects of UV exposure on single exponential decay constants were generally positive in all three sites and for both species, although UV effects depended on site (Figure 3). Overall, the low C:N B. gracilis litter had a higher decay constant than A. gerardii litter at all three sites (P < 0.0001). Contrary to expectations, the difference in decay constants between the two species was greater at Sevilleta than the other two sites (site*species interaction: P < 0.0001). Sevilleta had the highest decay constants, whereas Central Plains had the lowest (P < 0.0001). There was a significant UV*species interaction (P = 0.0231), nested within site. At Cedar Creek, B. gracilis litter decomposed 17% faster in the UV pass treatment than in the UV block treatment, but A. gerardii was not affected by the UV treatment (Figure 3A). In addition, the clipping treatment did not have a significant effect on decay constants at Cedar Creek (P = 0.2677, data not shown). At Central Plains, A. gerardii litter decomposed 50% faster in the UV pass treatment than the UV block treatment, but the decay constant for B. gracilis did not significantly differ between treatments (Figure 3B). At Sevilleta, both species decomposed faster under the UV pass treatment: A. gerardii showed a 100% increase in decay constant, and B. gracilis showed a 40% increase (Figure 3C).

Mean litter decay constant (k, y−1) of ash-free dry mass from a single exponential decay model for A. gerardii and B. gracilis litter under the UV block or UV pass treatment at A Cedar Creek, B Central Plains, and C Sevilleta. Experimental blocks that failed to fit an exponential decay model (P > 0.05) due to outliers were excluded from the analysis. Standard error shown, n > 7 in all cases.
Influence of Soil Accumulation on Mass Loss Patterns
Mass loss (Figure 4) was relatively steady over time at Cedar Creek and Central Plains, whereas litter at Sevilleta exhibited rapid mass loss between the first and second collection dates (20% of original mass), and between the third and fourth collection dates (35% of original mass), which corresponded with the monsoon seasons (see Figure 1). This pattern was partially reflected in the higher r 2 values of the single exponential model at Cedar Creek and Central Plains compared to Sevilleta. There was also substantial mineral soil accumulation at Sevilleta within some of the litterbags during these periods, and in fact soil accumulation was positively correlated with litter mass loss at Sevilleta (r 2 = 0.29, P < 0.0001). Adding accumulated soil mass as a covariate increased the goodness-of-fit of the model, but did not affect the significance of the UV effect. There was no significant soil accumulation or correlation of soil accumulation with litter mass loss at the other two sites.

Mean (n = 10) percent ash-free dry mass remaining over time in A. gerardii and B. gracilis litter under UV pass or UV block treatments at A Cedar Creek, B Central Plains, and C Sevilleta. Standard error shown.
Carbon Fraction Loss
Loss of both hemicellulose and cellulose fractions was always higher in B. gracilis than A. gerardii (Table 3, P < 0.0001). Lignin fractions tended to increase initially, possibly due to a buildup of “lignin-like” microbial by-products. Effects of UV radiation on particular carbon fractions in the litter depended on species and site, and therefore they were evaluated in separate statistical models (Table 4).
At Cedar Creek, lignin loss was not significantly different between UV treatments in A. gerardii but was twice as high under the UV pass compared to the UV block treatment in B. gracilis litter (Tables 3 and 4). At Central Plains, accumulation of “lignin-like” products in the UV pass treatment was half that of the UV block treatment on the first collection date, but there was no significant UV effect at any other collection date. Lignin loss at Sevilleta was closer to predicted patterns: A. gerardii litter lost 52 and 133% more lignin under the UV pass than UV block treatments at the third and fourth collection dates, respectively. This effect was marginally significant because there was insufficient sample remaining to analyze lignin content on the full ten replicates. There was a trend toward greater lignin loss for the UV pass treatment in B. gracilis for the first through third collection dates, but effects were not significant. There were insufficient replicates of B. gracilis to statistically analyze UV effects at Sevilleta at the fourth collection date due to the large amounts of mass loss.
UV radiation did not affect cellulose loss but did increase hemicellulose loss (Table 4). At Cedar Creek and Central Plains, UV exposure significantly increased loss of the hemicellulose fraction an average of 14% in both species (Table 3). At Sevilleta, UV exposure significantly increased loss of the hemicellulose fraction by 63% in A. gerardii at the fourth collection date. UV exposure did not affect loss of the hemicellulose fraction in B. gracilis at Sevilleta, although sample sizes were too small to statistically analyze for the final collection date.
Nitrogen Dynamics
Although there was a trend toward lower N immobilization in the UV pass treatment, there was no significant effect of UV radiation on N immobilization in A. gerardii at either Cedar Creek or Central Plains (Figure 5A, B). At Sevilleta, A. gerardii immobilized N after the first collection date, but lost N by the fourth collection date, more so in the UV pass treatment than UV block treatment (Figure 5C). At Sevilleta, B. gracilis lost N over the entire period (Figure 5C). There was a trend toward greater N loss in B. gracilis in the UV pass treatment than the UV block treatment at Cedar Creek, but the effect was not significant.

Mean (n = 10) litter nitrogen (% of initial) over time in A. gerardii and B. gracilis litter under UV pass or UV block treatments at A Cedar Creek, B Central Plains, and C Sevilleta. Values over 100% indicate net N immobilization, and values below 100% indicate net N loss. Standard error shown.
The relationship between N dynamics and litter mass loss was generally consistent with the Parton and others (2007) models (Figure 6) and, in contrast to expectations, did not differ between UV treatments (data not shown). In A. gerardii litter, the mesic system model was the best fit for Cedar Creek and Central Plains (lowest AICc) but tended to slightly underestimate litter N content at a given mass loss. At Sevilleta, the relationship between N content and litter mass loss followed the dry grassland and linear models equally well (delta r < 3). The models more closely fit measured N dynamics in B. gracilis at all three sites: the mesic system model was the best fit at Cedar Creek (lowest AICc); the linear model was the best fit at Central Plains and Sevilleta (lowest AICc).

Percent ash-free dry mass remaining versus litter nitrogen (% of initial) for A A. gerardii and B B. gracilis at the three sites, for all treatments. Lines represent modeled relationships based on Parton and others (2007) for dry grassland (dashed) and mesic (solid) sites. Dotted lines show a 1:1 N to litter mass loss relationship.
Extracellular Enzymes
We did not detect any oxidative enzyme activity (phenol oxidase or peroxidase) over the course of the experiment. Hydrolytic enzymes (phosphatase, β-1,4-glucosidase, n-acetyl-glucosaminidase, and cellobiohydrolase) were not affected by the UV treatment (P > 0.4 for all enzymes, data not shown). Activity of all enzymes did differ significantly between species (P < 0.0001) and among sites (P < 0.0001, Figure 7). These patterns did not differ significantly over time (data not shown). Phosphatase, β-1,4-glucosidase, n-acetyl-glucosaminidase, and cellobiohydrolase activities were positively correlated with one another in both species and all sites (P < 0.0001).

Activity of extracellular enzymes: A phosphatase, B β-glucosidase, C n-acetyl-glucosaminidase, and D cellobiohydrolase averaged across collection dates and treatments. Effects of site and species were statistically significant (P < 0.0001); A, A. gerardii and B, B. gracilis. The UV effect was not significant, and UV treatments were pooled for analysis. Means (n = 20) and standard error shown.
Discussion
Photodegradation and Litter Decomposition Rates
The results of this study provide further evidence that exposure to UV radiation increases litter decomposition rates and mass loss via photodegradation in arid grassland ecosystems and indicate that photodegradation is important in a wider range of grasslands, including more mesic sites such as Cedar Creek. We had expected that higher canopy cover would impede penetration of solar radiation to the litter layer at Cedar Creek. Even at peak standing biomass, however, canopy cover at Cedar Creek reduced transmission of solar radiation by only 17%. Canopy cover was not sufficient to reduce the effect of photodegradation as both clipped and unclipped plots exhibited the same positive effects of UV radiation on decomposition. We had also expected that Cedar Creek’s higher latitude and lower elevation than the other two sites, which reduces the annual UV dose, would also impede photodegradation. However, unlike the other two sites, litter at Cedar Creek did not become buried by soil and thus had a greater surface area that was exposed to solar radiation. This factor could have led to a larger role of photodegradation at Cedar Creek than originally predicted, and it highlights the importance of site-specific influences on the role of photodegradation in decomposition. Additionally, because extracellular enzyme activity at Cedar Creek was very high, it is possible that microbial breakdown of the litter made it more susceptible to photodegradation, a mechanism that has been supported in studies examining the photodegradation of dissolved organic carbon (for example, Amado and others 2007).
A primary goal of this study was to test if photodegradation plays a larger role in arid than in mesic grassland systems. This hypothesis was partially supported by our results. In A. gerardii litter, the role of photodegradation increased with site aridity and ambient UV radiation as expected. Exposure to UV radiation resulted in no significant increase in mass loss or higher decay constants at Cedar Creek, a 50% higher decay constant at Central Plains, and doubled decay constants at Sevilleta. Patterns in B. gracilis litter did not fit expectations as clearly: UV exposure had the greatest effect at Sevilleta as expected, but played a larger role at Cedar Creek than at Central Plains. The driver of this pattern in B. gracilis litter decomposition remains unclear. However, as mentioned above, greater microbial decomposition at Cedar Creek than the other two sites could have facilitated photodegradation in B. gracilis.
Although photodegradation played the largest role at the most arid site, it could not completely account for the rapid decay observed at Sevilleta. Even when UV radiation was blocked from reaching the litter layer, decay constants for B. gracilis at Sevilleta were twice those at Cedar Creek, and rates of A. gerardii litter decay were equal between Cedar Creek and Sevilleta. This pattern was consistent across years and was not due to any anomalous climate characteristics during the experiment (Brandt 2009). The rapid mass loss during the summer monsoon seasons coupled with the strong correlation between litter mass loss and soil accumulation in the litterbags at Sevilleta lend support to the alternative hypotheses that precipitation pulses and soil burial may be partially driving high rates of litter decay in arid ecosystems such as Sevilleta (Austin and others 2004; Throop and Archer 2007). The synchronization of precipitation pulses with high temperatures during the monsoons could have led to periods of rapid microbial activity as well, which could have been further facilitated by greater contact with the soil microbial community once litter became buried. Because both UV exposure and soil burial had positive effects on litter mass loss at Sevilleta, it is possible that high mass loss at the site is due to a combination of several abiotic and biotic factors, including, but not limited to, photodegradation.
Litter Decay Models
We hypothesized that when photodegradation plays a larger role in litter decomposition, litter mass loss should follow a linear decay model instead of an exponential decay model. Previous field and laboratory experiments separating photochemical from microbial decay have found support for this hypothesis (Austin and Vivanco 2006; Brandt and others 2009). In this study, however, linear and exponential decay models fit equally well in most cases. Many studies that have found linear patterns of decay in arid ecosystems have been conducted for 5 years or more (Adair and others 2008; Vanderbilt and others 2008). Thus, our study may have been too limited in duration and number of collection points to enable us to distinguish between alternative models. In addition, photochemical decay was also allowed to occur simultaneously with microbial decay in this study. Thus, even in cases where photodegradation played a large role in litter decomposition in this study, it was never solely responsible for mass loss. Therefore, the linear pattern of photodegradation may be masked by other decay patterns attributable to microbial or other abiotic processes.
Carbon Fraction Loss Attributed to Photodegradation
In agreement with our hypothesis, photodegradation via UV radiation played a larger role in decomposition of A. gerardii (8.1% lignin) than B. gracilis (6.6% lignin) at both Central Plains and Sevilleta. However, greater impacts of UV exposure were seen in B. gracilis than A. gerardii at Cedar Creek. These results suggest that the impacts of photodegradation on a particular litter type may not be tightly correlated with initial lignin content (but see Austin and Ballaré 2010). The species we chose, however, did not differ greatly in lignin content, so this relationship cannot be ruled out entirely. It is not surprising that lignin loss was not tightly coupled to total litter mass loss from photodegradation because lignin makes up a small fraction of initial litter mass (less than 10%) in both species and is generally low in grasses. Previous research has shown that UV exposure leads to increased lignin loss (Rozema and others 1997; Day and others 2007; Henry and others 2008; Austin and Ballaré 2010), but in general these effects have been small and did not account for the entirety of litter mass loss.
UV exposure tended to increase loss of the hemicellulose fraction, consistent with a few previous studies (Rozema and others 1997; Brandt and others 2007). Hemicellulose makes up a much larger proportion of the initial litter mass (32% in A. gerardii, 38% in B. gracilis) than lignin does; therefore, it has a much greater potential to influence total litter mass loss if it is photochemically susceptible. Photodegradation could weaken the lignocellulose matrix, making hemicellulose more accessible to hydrolytic enzymatic decay through an increase in binding site availability (Gallo and others 2009). Alternatively, hemicellulose could be broken down through indirect photolysis, in which the absorption of UV radiation by lignin leads to the production of free radicals, which break bonds in other compounds, such as hemicellulose, in the lignocellulose matrix (Schade and others 1999). Further research is needed to test whether either of these two potential mechanisms is responsible for hemicellulose loss.
Photodegradation and Nitrogen Dynamics
We hypothesized that mass loss from photodegradation would be decoupled from N dynamics because photodegradation, unlike microbial decomposition, does not require N. However, the relationship between litter N and mass loss was not affected by UV exposure as we would have predicted if N dynamics and litter mass loss were decoupled in the presence of photodegradation. Despite the fact that overall N dynamics were largely unaffected by exposure to UV radiation, N dynamics at Sevilleta appeared to be driven by other abiotic processes. Nitrogen and litter mass loss from both litter types closely followed a 1:1 linear model, indicating that litter did not immobilize N. Because total litter mass and N loss occurred at the same rate, it is unlikely that N loss was due to microbial mineralization. The C:N ratio of B. gracilis remained at initial values throughout the experiment, and the C:N ratio of A. gerardii remained above 115, well above critical stoichiometric ratios necessary for microbial mineralization (Manzoni and others 2008). This result suggests that unusual N dynamics observed in arid systems might be due to abiotic drivers other than photodegradation such as precipitation pulses, which could cause rapid leaching of N, or soil burial, which could abrade litter and cause rapid apparent decomposition via fragmentation (Austin and others 2004; Throop and Archer 2007).
Interactions Between Enzymatic and Photochemical Processes
Extracellular enzyme activity was equal between UV treatments in all sites regardless of whether there was a significant UV effect on litter decay rates, indicating that UV effects on decomposition could be independent of effects on microbial activity (Brandt and others 2009). Alternatively, equal enzyme activity between UV treatments could indicate decreased “enzymatic turnover activity,” the quantity of enzymatic activity needed to degrade a cohort of litter (Gallo and others 2009). If decomposition rates increase but enzyme activity remains constant with UV exposure, the activity to mass loss ratio decreases, meaning less energy is required by the microbial community to degrade a particular substrate. This alternative hypothesis would suggest that photodegradation could indeed be facilitating microbial breakdown of litter. However, we did not find support for this hypothesis in a previous study (Brandt and others 2009).
We did not detect oxidative enzyme activity in either litter species at any site or time during the decomposition process. Gallo and others (2006) suggest that a lack of oxidative enzyme activity in high light environments could indicate that photochemical processes negate the need for microbial lignin degradation. Although this is possible, the fact that no oxidative enzyme activity was observed in any of the litter, including the UV block treatment at Cedar Creek, may indicate that other processes are at work. Oxidative enzyme activity from surface soils collected at similar grassland sites has been observed and is often the highest in arid environments (Zeglin and others 2007). A study at Cedar Creek found that oxidative activity in litter is relatively low or undetectable, even in forested systems with little light penetration to the litter layer (Keeler and others 2009). It is possible that litter at this stage in the decomposition process simply may not be sufficiently decomposed to the point where oxidative enzymes are required to access energy-rich substrates (Moorhead and Sinsabaugh 2006). Modeling and field studies suggest that until litter reaches a lignocellulose index (LCI, the ratio of lignin:(lignin + cellulose)) of 0.4, microbial decay of the lignin portion will not occur (Aber and others 1984; Moorhead and Sinsabaugh 2006). LCI in this study never reached that threshold, and in fact remained much lower (mean LCI at final collection date = 0.14, max = 0.39). Coupled with the fact that hydrolytic enzyme activity remained high throughout the 2 years of decomposition, the most likely explanation for the lack of detectable oxidative enzyme activity in this study is simply the early stage in the decomposition process.
The results of these enzyme assays are in contrast to previous research demonstrating negative effects of UV exposure on decomposers (Gehrke and others 1995; Duguay and Klironomos 2000; Pancotto and others 2003). Much of this previous work has examined effects of supplemental UV radiation on microbial communities that are not adapted to high levels of solar radiation (Gehrke and others 1995; Duguay and Klironomos 2000; Pancotto and others 2003). Here we examined the effects of ambient UV radiation on microbial communities that are adapted to high light environments and may have protective pigments and other mechanisms to shield them from UV-induced DNA damage (Gallo and others 2009). Recent evidence suggests that UV radiation may lead to shifts in microbial community composition even in arid environments (Belnap and others 2008; Gallo and others 2009). However, these shifts in community composition do not necessarily translate into decreased extracellular enzyme activity (Gallo and others 2009).
Conclusion
This study shows that photodegradation contributes to litter mass loss in a wider range of grassland ecosystems than previously documented. Litter mass loss via photodegradation appears to be independent of factors that influence rates of biological decomposition, including litter lignin and N content and microbial extracellular enzyme activity. Photodegradation does seem to play a larger role in arid ecosystems where biological decomposition is lower, and therefore can partially explain decomposition patterns in arid and semiarid grasslands. However, the higher role of photodegradation in arid systems cannot completely account for higher than predicted litter decomposition rates and atypical N dynamics in arid ecosystems. Further empirical and modeling studies of interactions between photodegradation and other abiotic and biotic controls on decomposition are needed to more completely explain differences in litter dynamics between arid and mesic grassland ecosystems.
References
Aber JD, McClaugherty CA, Melillo JM. 1984. Litter decomposition in Wisconsin forests: mass loss, organic-chemical constituents and nitrogen. Report #R3284, School of Natural Resources, University of Wisconsin, Madison, Wisconsin, USA.
Adair EC, Parton WJ, Del Grosso SJ, Silver WL, Harmon ME, Hall SA, Burke IC, Hart SC. 2008. Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Glob Change Biol 14:2636–60.
Amado AM, Cotner JB, Suhett AL, de Assis Esteves F, Bozelli RL, Farjalla VF. 2007. Contrasting interactions mediate dissolved organic matter decomposition in tropical aquatic ecosystems. Aquat Microb Ecol 49:25–34.
Austin AT, Ballaré CL. 2010. Dual role of lignin in plant litter decomposition in terrestrial systems. Proc Natl Acad Sci 107:4618–22.
Austin AT, Vivanco L. 2006. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442:555–8.
Austin AT, Yahdjian L, Stark JM, Belnap J, Porporato A, Norton U, Ravetta DA, Schaeffer SM. 2004. Water pulses and biogeochemical cycles in arid and semiarid ecosystems. Oecologia 141:221–35.
Belnap J, Phillips SL, Flint S, Money J, Caldwell M. 2008. Global change and biological soil crusts: effects of ultraviolet augmentation under altered precipitation regimes and nitrogen additions. Glob Change Biol 14:670–86.
Brandt LA. 2009. The role of photodegradation in plant litter decomposition in grassland ecosystems. Ph.D. Dissertation. University of Minnesota. 144 p.
Brandt LA, Bohnet C, King JY. 2009. Photochemically induced carbon dioxide production as a mechanism for carbon loss from plant litter in arid ecosystems. J Geophys Res 114:G02004. doi:10.1029/2008JG000772.
Brandt LA, King JY, Milchunas DG. 2007. Effects of ultraviolet radiation on litter decomposition depend on precipitation and litter chemistry in a shortgrass steppe ecosystem. Glob Change Biol 13:2193–205.
Burnham KP, Anderson DR. 2002. Model selection and multimodel inference: a practical information-theoretic approach. 2nd edn. New York: Springer-Verlag. p 488.
Day T, Zhang E, Ruhland C. 2007. Exposure to solar UV-B radiation accelerates mass and lignin loss of Larrea tridentata litter in the Sonoran Desert. Plant Ecol 193:185–94.
Duguay KJ, Klironomos JN. 2000. Direct and indirect effects of enhanced UV-B radiation on the decomposing and competitive abilities of saprobic fungi. Appl Soil Ecol 14:157–64.
Gallo ME, Porras-Alfaro A, Odenbach KJ, Sinsabaugh RL. 2009. Photoacceleration of plant litter decomposition in an arid environment. Soil Biol Biochem 41:1433–41.
Gallo ME, Sinsabaugh RL, Cabaniss SE. 2006. The role of ultraviolet radiation in litter decomposition in arid ecosystems. Appl Soil Ecol 34:82–91.
Gehrke C, Johanson U, Callaghan TV, Chadwick D, Robinson CH. 1995. The impact of enhanced ultraviolet-B radiation on litter quality and decomposition processes in Vaccinium leaves from the Subarctic. Oikos 72:213–22.
Henry HAL, Brizgys K, Field C. 2008. Litter decomposition in a California annual grassland: Interactions between photodegradation and litter layer thickness. Ecosystems 11:545–54.
Keeler B, Hobbie S, Kellogg L. 2009. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition. Ecosystems 12:1–15.
Lane DR, Coffin DP, Lauenroth WK. 2000. Changes in grassland canopy structure across a precipitation gradient. J Veg Sci 11:359–68.
MacKay WP, Loring SJ, Zak JC, Silva SI, Fisher FM, Whitford WG. 1994. Factors affecting loss in mass of creosotebush leaf-litter on the soil surface in the Northern Chihuahuan Desert. Southwest Nat 39:78–82.
Manzoni S, Jackson RB, Trofymow JA, Porporato A. 2008. The global stoichiometry of litter nitrogen mineralization. Science 321:684–6.
Meentemeyer V. 1978. Macroclimate and lignin control of litter decomposition rates. Ecology 59:465–72.
Moorhead DL, Callaghan T. 1994. Effects of increasing ultraviolet B radiation on decomposition and soil organic matter dynamics: a synthesis and modeling study. Biol Fertil Soils 18:19–26.
Moorhead DL, Reynolds JF. 1989. The contribution of abiotic processes to buried litter decomposition in the northern Chihuahuan Desert. Oecologia 79:133–5.
Moorhead DL, Sinsabaugh RL. 2006. A theoretical model of litter decay and microbial interaction. Ecol Monogr 76:151–74.
Olson JS. 1963. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 44:322–31.
Pancotto VA, Sala OE, Cabello M, Lopez NI, Robson TM, Ballare CL, Caldwell MM, Scopel AL. 2003. Solar UV-B decreases decomposition in herbaceous plant litter in Tierra del Fuego, Argentina: potential role of an altered decomposer community. Glob Change Biol 9:1465–74.
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC, Fasth B. 2007. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–4.
Reich PB, Peterson DW, Wedin DA, Wrage K. 2001. Fire and vegetation effects on productivity and nitrogen cycling across a forest–grassland continuum. Ecology 82:1703–19.
Rozema J, Tosserams M, Nelissen JM, van Heerwaarden L, Broekman RA, Flierman N. 1997. Stratospheric ozone reduction and ecosystem processes: enhanced UV-B radiation affects chemical quality and decomposition of leaves of the dune grassland species Calamagrostis epigeios. Plant Ecol 128:284–94.
Saiya-Cork KR, Sinsabaugh RL, Zak DR. 2002. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol Biochem 34:1309–15.
Santos PF, Whitford WG. 1981. The effects of micro arthropods on litter decomposition. Ecology 62:654–63.
Schade GW, Hofmann RM, Crutzen PJ. 1999. CO emissions from degrading plant matter: (I). Measurements. Tellus 51B:889–908.
Schaefer D, Steinberger Y, Whitford WG. 1985. The failure of nitrogen and lignin control of decomposition in a North American desert. Oecologia 65:382–6.
Shore G. 1997. Sevilleta LTER vegetation plot LAI data. Sevilleta LTER Database. http://www.sevilleta.unm.edu/data/archive/plant/vegmap/classified/lai. June 16, 2005.
Sinsabaugh RL, Lauber CL, Weintraub MN, Ahmed B, Allison SD, Crenshaw C, Contosta AR, Cusack D, Frey S, Gallo M, Gartner TB, Hobbie SE, Holland K, Keeler BL, Powers JS, Stursova M, Takacs-Vesbach C, Waldrop M, Wallenstein M, Zak DR, Zeglin LH. 2008. Stoichiometry of soil enzyme activity at global scale. Ecol Lett 11:1252–64.
Throop HL, Archer SR. 2007. Interrelationships among shrub encroachment, land management, and litter decomposition in a semidesert grassland. Ecol Appl 17:1809–23.
Van Soest PJ. 1967. Development of a comprehensive system of feed analyses and its application to forages. J Anim Sci 26:119–28.
Vanderbilt KL, White CS, Hopkins O, Craig JA. 2008. Aboveground decomposition in arid environments: results of a long-term study in central New Mexico. J Arid Environ 72:696–709.
Whitford WG, Meentemeyer V, Seastedt TR, Cromack K Jr, Crossley DA Jr, Santos P, Todd RL, Waide JB. 1981. Exceptions to the AET model: deserts and clear-cut forest. Ecology 62:275–7.
Whitford WG, Steinberger Y, MacKay W, Parker LW, Freckman D, Wallwork JA, Weems D. 1986. Rainfall and decomposition in the Chihuahuan Desert. Oecologia 68:512–15.
Zak DR, Tilman D, Parmenter RR, Rice CW, Fisher FM, Vose J, Milchunas D, Martin CW. 1994. Plant production and soil microorganisms in late-successional ecosystems: a continental-scale study. Ecology 75:2333–47.
Zeglin L, Stursova M, Sinsabaugh R, Collins S. 2007. Microbial responses to nitrogen addition in three contrasting grassland ecosystems. Oecologia 154:349–59.
Acknowledgements
We thank S. Graves, A. Thone, K. Crooker, Y. Sorokin, K. Hill, F. Walosin, C. Bohnet, M. Gallo, W. Eddy, C. Buyarski, and M. Stursova for their assistance in the field and lab. We thank L. Kinkel and J. McFadden for their assistance in the experimental design and for their helpful comments on the manuscript. We thank the Sevilleta National Wildlife Refuge and LTER (DEB-8811906, DEB-0080529, IBN-9411976), the USDA-ARS Central Plains Experimental Range and Shortgrass Steppe LTER (NSF DEB-0217631, 0823405), and the Cedar Creek Ecosystem Science Reserve and LTER (DEB-0080382) for use of field facilities, meteorological data, and technical support. This project was funded by NSF Ecosystem Science (DEB-0542935, DEB-0935984). Additional funding to L. Brandt was provided by an EPA STAR Fellowship, a University of Minnesota Dissertation Research Grant, the University of Minnesota Carolyn Crosby Fellowship, and the Dayton-Wilkie Natural History Funds.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
LB and JK conceived of and designed the study with contributions from DM, SH, and RS; all authors contributed to performing the research; LB analyzed data and wrote the paper with input from JK, DM, SH, and RS.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Brandt, L.A., King, J.Y., Hobbie, S.E. et al. The Role of Photodegradation in Surface Litter Decomposition Across a Grassland Ecosystem Precipitation Gradient. Ecosystems 13, 765–781 (2010). https://doi.org/10.1007/s10021-010-9353-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-010-9353-2