
Abstract
Tendon injuries occur commonly in both human and equine athletes, and poor tendon regeneration leads to functionally deficient scar tissue and an increased frequency of re-injury. Despite evidence suggesting inadequate resolution of inflammation leads to fibrotic healing, our understanding of the inflammatory pathways implicated in tendinopathy remains poorly understood, meaning successful targeted treatments are lacking. Here, we demonstrate IL-1β, TNFα and IFN-γ work synergistically to induce greater detrimental consequences for equine tenocytes than when used individually. This includes altering tendon associated and matrix metalloproteinase gene expression and impairing the cells’ ability to contract a 3-D collagen gel, a culture technique which more closely resembles the in vivo environment. Moreover, these adverse effects cannot be rescued by direct suppression of IL-1β using IL-1RA or factors produced by BM-MSCs. Furthermore, we provide evidence that NF-κB, but not JNK, P38 MAPK or STAT 1, is translocated to the nucleus and able to bind to DNA in tenocytes following TNFα and IL-1β stimulation, suggesting this signalling cascade may be responsible for the adverse downstream consequences of these inflammatory cytokines. We suggest a superior approach for treatment of tendinopathy may therefore be to target specific signalling pathways such as NF-κB.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tendon injuries are a major cause of musculoskeletal morbidity in equine athletes. In racing Thoroughbreds, tendon injuries account for 46% of all limb injuries on racecourses, with the superficial digital flexor tendon (SDFT) being most frequently affected (Williams et al. 2001). The comparable nature of the equine SDFT and human Achilles tendon, both acting as an elastic energy store for high-speed locomotion, makes the horse an attractive model for the study of equivalent human injury (Patterson-Kane et al. 2012; Sobhani et al. 2013; Cassel et al. 2015; Ellis et al. 2022). Post-injury, the healing response is inadequate leading to the formation of inferior quality scar-like tissue. This compromises the tendons structural integrity leading to prolonged recouperation periods, increased retirement rate and high re-injury rates of up to 67% in equine athletes (Dyson 2004; Dakin et al. 2012). Although tendon injuries pose an important clinical problem, our complete understanding of the mechanisms underpinning dysregulated healing is lacking.
Historically, tendinopathy was described as a degenerative condition devoid of inflammation, as inflammatory cells were rarely identified in histological studies (Alfredson and Lorentzon 2002). However, modern molecular techniques have identified a strong inflammatory component to acute tendinopathy, with upregulation of pro-inflammatory cytokines including IFN-γ (interferon gamma), TNFα (tumour necrosis factor alpha), IL-1β (interleukin 1 beta) and IL-6 (interleukin 6) within the tendon matrix microenvironment (Hosaka et al. 2002; Dakin et al. 2012; Dakin et al. 2014; Millar et al. 2017; Morita et al. 2017). Although moderate levels of inflammation are required to initiate tendon repair, growing evidence suggests inadequate resolution of inflammation contributes to fibrotic healing (Dakin et al. 2012; Abraham et al. 2017; Millar et al. 2017). Previously, in vitro studies have demonstrated that TNFα and IL-1β used individually exhibit detrimental consequences for equine adult tenocyte function by significantly increasing the expression of matrix metalloproteinases (MMPs) and altering the expression of tendon-associated genes (John et al. 2010; McClellan et al. 2019a, b). Additionally, IL-1β has been shown to impair three-dimensional (3-D) collagen gel contraction by tenocytes (McClellan et al. 2019a), a culture technique which more closely resembles the in vivo 3-D environment than plastic adherent monolayer cell culture (Paterson et al. 2020). Nevertheless, the synergistic effect of multiple inflammatory cytokines on tendon function in vitro is unknown.
Despite the evidence implicating inflammation in tendinopathy, there remains a lack of consensus surrounding inflammation-targeting treatments. Currently, the treatments showing the most promise include cellular therapies (Godwin et al. 2012; Gaspar et al. 2015), platelet-rich plasma (PRP) (Filardo et al. 2018), gene therapies (Tang et al. 2016) and pharmaceutical interventions which target specific inflammatory cytokines (Fredberg and Ostgaard 2009; Berkoff et al. 2016). Non-steroidal anti-inflammatory drugs (NSAIDs) which inhibit cyclooxygenase (COX) activity are often used initially following tendinopathy diagnosis to relieve pain (Wang et al. 2019). However, non-specific blockage of the inflammatory response using NSAIDs may inhibit tendon cell migration and proliferation and ultimately impair tendon healing (Marsolais et al. 2003). Therefore, for a tendinopathy treatment to be effective, it must be capable of diminishing the inflammatory response whilst still promoting tendon regeneration.
Bone marrow–derived mesenchymal stromal cells (BM-MSCs) are commonly used to treat equine tendon injuries. Clinical and experimental evidence shows BM-MSCs improve tendon healing and may be beneficial to reduce re-injury rates (Godwin et al. 2012; Smith et al. 2013). This may be due to BM-MSCs having regenerative and immunomodulatory properties (Paterson et al. 2014; Sullivan et al. 2014); however, their specific mechanism of action is largely unknown. The original concept was that BM-MSCs acted by direct differentiation into tenocytes due to their potential to differentiate into lineages of the mesenchyme such as osteoblasts, chondrocytes and adipocytes in vitro (Guest et al. 2008). However, the prolonged survival of BM-MSCs injected into injured equine tendon is poor (< 5% survival 10 days following implantation) (Guest et al. 2010; Becerra et al. 2013), suggesting BM-MSCs instead help tendon healing through trophic effects. BM-MSCs can modulate the inflammatory environment by influencing the activities of resident macrophages (Manning et al. 2015). Other studies suggest BM-MSCs can improve tendon healing through the secretion of soluble factors which protect endogenous cells from inflammation (Viganò et al. 2019). Consequently, it is apparent that further work is required to establish the exact role BM-MSCs have in improving tendon regeneration.
More targeted approaches to reduce inflammation in tendinopathy are under investigation. In vitro, blockage of IL-1β signalling using exogenous IL-1 receptor antagonist protein (IL-1RA) rescued the adverse consequences of IL-1β on 3-D collagen gel contraction by equine tenocytes (McClellan et al. 2019a). Nevertheless, IL-1RA is specific to IL-1β signalling and is unlikely to block the effects of the other inflammatory cytokines known to be upregulated in the injured tendon.
Inflammatory cytokines activate numerous signalling cascades including c-Jun N-terminal kinase (JNK), mitogen-activated protein kinases (MAPKs), nuclear factor kappa B (NF-κB) and signal transducer and activator of transcription (STATs) in musculoskeletal cell types (Tsai et al. 2014; McClellan et al. 2019a, b; Wang et al. 2019; Freedman et al. 2022). Furthermore, several animal models have implicated these inflammatory signalling pathways within tendon extracellular matrix interactions (Gupta et al. 2010; Schwartz et al. 2015; Abraham et al. 2017). Therefore, rather than targeting specific cytokines, it would be more beneficial to dissect the role inflammatory pathways such as NF-κB play in mediating tendon regeneration. This would allow the development of novel targeted therapeutic interventions which enhance tendon healing.
This in vitro study explored the effect a combination of IFN-γ, TNFα and IL-1β has upon equine tenocyte function and identified the key inflammatory signalling pathways which they activate. The effects of IL-1RA and BM-MSCs on attenuating adverse inflammatory responses to this combination of inflammatory cytokines were also determined.
Materials and methods
Study design
An overview of the study design is provided in Fig. 1. Initial experiments measured NF-κB P65 nuclear translocation in equine tenocytes following exposure to different concentrations of IFN-γ, TNFα and IL-1β for various durations (Fig. 1a). The optimal dose and exposure time for maximal P65 translocation were then used to determine if there were any effects on cell viability and proliferation, confirm P65 DNA binding and determine if other signalling pathways (STAT1, JNK and P38 MAPK) were activated by measuring their nuclear translocation (Fig. 1b). Antibody specificity was confirmed using western blot. The effect of the cytokines on tenocyte 3-D collagen gel contraction, 2-D gene expression and IL-6 production was then determined (Fig. 1c). As IL-6 production was found to be increased following exposure to the inflammatory cytokines, the effect of IL-6 on P65 translocation, tenocyte 3-D collagen gel contraction and 2-D gene expression was determined (Fig. 1d). We then determined the effect of IL-1RA and BM-MSCs (co-culture and conditioned media) on attenuating adverse inflammatory responses by measuring their effects on cytokine mediated P65 translocation, tenocyte 3-D collagen gel contraction and 2D gene expression (Fig. 1e).

Experimental study design outlining cytokine optimisation (a), inflammatory pathway activation (b), effect of inflammatory cytokines on equine tenocytes (c), effect of IL-6 on equine tenocytes (d) and BM-MSC or IL-1RA rescue (e). Created with BioRender.com
Tendon cell isolation and culture
Tenocytes were harvested post-mortem from healthy SDFTs of a total of ten adult Thoroughbred horses euthanized for reasons unrelated to this study with approval from the Royal Veterinary College Clinical Research Ethical Review Board (URN 2020 2017–2). The horses used were aged between 2 and 13 years old: eight were males, and two were females. Tenocytes were isolated and cultured as described previously (Barsby and Guest 2013). Briefly, tendon tissue was dissected into pieces and incubated with 1 mg/mL type I collagenase from Clostridium histolyticum (Sigma-Aldrich, Dorset, UK) for 14–16 h at 37 °C. Dissociated cells were cultured in growth media consisting of Dulbecco’s modified Eagle’s medium (DMEM; high glucose [4500 mg/L] with sodium pyruvate [110 mg/L]) with 10% foetal bovine serum (FBS), 1% penicillin–streptomycin (P/S) and 2 mM L-glutamine (LQ) (all Gibco, Thermo Fisher, Hemel Hempstead, UK). Tenocytes were cultured at 37 °C in a humidified atmosphere of 5% CO2 and passaged before reaching maximum confluency every 3–4 days using 0.25% trypsin–EDTA (Sigma-Aldrich).
Immunofluorescence
Three biological replicates of tenocytes (between P3 and P10) were cultured on gelatin-coated glass coverslips (Sigma-Aldrich; 70–80% confluent) prior to stimulation with recombinant human TNFα (25 pg/mL and 0.1, 1, 10 ng/mL) (Cheshire and Baldwin 1997; Lee et al. 2007; Tsai et al. 2014), recombinant human IL-1β (0.0425, 0.17, 1.7 and 17 ng/mL) (Gehwolf et al. 2019; McClellan et al. 2019a; Vinhas et al. 2020) and recombinant equine IFN-γ (1, 100, 200, 500 ng/mL) (Paterson et al. 2014; Yang et al. 2014; McClellan et al. 2019b) (all PeproTech, London, UK) alone or in combination for 20 min, 1 h, 2 h and 24 h. Unstimulated cells served as controls. Following stimulation, coverslips were fixed with 3% paraformaldehyde for 20 min. Fixed cells were permeabilised with 0.1% Triton-X-100 (Sigma-Aldrich) at room temperature for 1 h and then blocked with 2.5% normal horse serum (NHS) (Vector Laboratories, Peterborough, UK) for 20 min. Incubation with primary antibodies (see Table 1) was completed overnight at 4 °C in NHS, before detection with either goat anti-mouse IgG Alexa fluor 594 1:200 or goat anti-rabbit IgG Alexa Fluor 594 1:200 (both Thermo Fisher) in 2.5% NHS for 3 h at room temperature. Negative controls were performed using the secondary antibody only. Coverslips were mounted using Vectashield Hardset with DAPI (4′,6-diamidino-2-phenylindole, Vector Laboratories). Images were acquired using a Nikon Eclipse Ti2 series microscope (Nikon, Surrey, UK). Nuclear fluorescent intensity was quantified by measuring mean grey scale of the nucleus using ImageJ software.
Western blot analysis
To confirm that the antibodies for NF-κB P65 (3 μg/mL), STAT1 (1:10,000) and JNK 1, 2 and 3 (1:1000) could recognise the equine protein, a western blot was performed. Whole cell protein extract (WCE) was isolated from equine skin fibroblasts (P5) by three rounds of freeze-thawing in a whole cell extraction buffer (20 mM Hepes pH 7.9, 450 mM NaCl, 0.4 mM EDTA, 25% glycerol, 1 mM PMSF), with supernatants collected by centrifugation. Twenty micrograms of reduced protein was then run on a TEO-Tricine Precast Gels, RunBlue™ 4–12% (Abcam) and transferred to a PDVF or nitrocellulose membrane. Immunoreactivity was detected using goat anti-mouse IgG H&L (HRP) (Abcam, ab6789, 1:10,000) or goat anti-rabbit IgG H&L (HRP) (Abcam, ab6721, 1:5000) and the ECL plus Western Blotting Substrate (Pierce™, Thermo Fisher).
NF-κB activation assay
A DNA-binding ELISA was performed using three biological replicates of tenocytes between P6 and P10 that had been exposed to TNFα (10 ng/mL) (Cheshire and Baldwin 1997), IL-1β (17 ng/mL) (Tsuzaki et al. 2003; McClellan et al. 2019a) and/or IFN-γ (100 ng/mL) (Paterson et al. 2014) alone or in combination for 1 h. Unstimulated cells were used as controls. WCEs were isolated as described above. Accurate determination of protein concentration was determined using a Pierce BCA Protein Assay Kit (Thermo Fisher) and Tecan plate reader measuring absorbance at 562 nm (Infinite M Plex; Tecan, Switzerland). Protein samples were then subsequently analysed for NF-κB activation using the TransAM™ NF-κB P65 transcription factor assay kit (Active Motif, Belgium), according to the manufacturer’s instructions. Briefly, 20 μg of whole cell protein was added to each well of a 96-well plate containing a consensus NF-κB oligonucleotide and incubated for 1 h to allow the binding of P65 to the NF-κB consensus site. The presence of the resulting complex was detected using an NF-κB primary antibody, followed by a secondary antibody conjugated to horseradish peroxidase (HRP) which provided a sensitive colorimetric readout which could be quantified by spectrophotometry (Infinite M Plex; Tecan).
Population doubling time and cell viability assay
The mean population doubling time (DT) was calculated from tenocytes in 2-D culture following 72 h stimulation with all combinations of TNFα (10 ng/mL), IL-1β (17 ng/mL) and/or IFN-γ (100 ng/mL). DT was calculated using the formula DT = T ln2/ln(Xe/Xb), where T = incubation time, Xb = starting cell number and Xe = ending cell number.
To measure differences in cell viability, tenocytes were seeded onto 96-well tissue culture plates at a concentration of 1.5 × 104 cells/well in growth media and allowed to attach at 37 °C and 5% CO2 for 24 h. After 24 h, the tenocytes were stimulated with combinations of TNFα (10 ng/mL), IL-1β (17 ng/mL) and/or IFN-γ (100 ng/mL) for 72 h. After incubation, culture media was removed and 100 μL of diluted PrestoBlue™ reagent (1:10; Invitrogen, Thermo Fisher) was added to each well and the plates were incubated at 37 °C for 30 min. Fluorescence of each well was measured at an excitation wavelength of 560 nm and an emission of 590 nm on a Tecan plate reader (Infinite M Plex; Tecan, Switzerland). Both DT and cell viability assays were performed using three biological replicates of tenocytes (P4–P11).
Three-dimensional cell culture of tenocytes
Three-dimensional (3-D) cellular culture in collagen gels was performed for 14 days as previously described (Barsby et al. 2014; Bavin et al. 2017; McClellan et al. 2019a, b; Paterson et al. 2020) using four biological replicates of tenocytes (P4–P10). Pairs of 0.2-mm-diameter minutien pins were embedded 15 mm apart into a silicone-coated six-well plate (Sylgard 184 Silicone elastomer; Corning) with each well containing three pairs of pins. Tenocytes (4 × 105 cells/mL) were suspended in a chilled mixture of 2 parts growth medium and 8 parts PureCol (Bovine collagen type I; Advanced Biomatrix, Carlsbad, USA) with the pH adjusted to 7.2–7.6 with 1 M sodium hydroxide. Subsequently, 200 μL of collagen-cell suspension was pipetted between each pair of minutien pins. The plate was then parafilm sealed and kept at 37 °C for 60–90 min to allow the constructs to set. Once set, the 3-D constructs were cultured at 37 °C and 5% CO2, in growth media alone (no cytokine control) or containing combinations of TNFα (10 ng/mL), IL-1β (17 ng/mL), IFN-γ (100 ng/mL) and/or IL-1Ra (100 ng/mL) or IL-6 (10 ng/mL). Media was replenished every 3–4 days with new fresh cytokines as applicable.
The 3-D tendon constructs were photographed daily. Contraction analysis of these images was performed by measuring the average diameter of each gel using ImageJ software (National Institutes of Health, USA). Contraction data was displayed as a percentage of the day 0 value. To measure cell survival, constructs were harvested and digested in 1 mL of growth media with 1 mg/mL type I collagenase from Clostridium histolyticum (Sigma-Aldrich) for 20 min–1 h at 37 °C. Cell counts were performed using a haemocytometer, and results displayed as a percentage of the number of cells seeded on day 0. Collagen gel contraction and cell survival were performed using three to four biological replicates of tenocytes with each replicate containing 1 to 9 tendon-like constructs.
RNA extraction, cDNA synthesis and quantitative PCR
Tenocytes from three biological replicates between P4 and P8 were used in these experiments. RNA was extracted using 1 mL Tri-reagent (Sigma-Aldrich) per 10 cm plate, or 3 wells of a 6-well plate, of 2-D cultured cells. RNA was next purified using the Qiagen RNeasy kit (Qiagen, Manchester, UK) and contaminating genomic DNA removed using a DNA-free™ DNA removal kit (Invitrogen, Thermo Fisher) according to the manufacturer’s instructions. RNA concentrations were calculated using a DeNovix Spectrophotometer (DeNovix, Wilmington, USA), ensuring the 260:280 ratio was between 1.8 and 2.2. cDNA was synthesized from 1 μg of RNA using the sensiFAST™ cDNA synthesis kit (Bioline, London, UK). Equine-specific primers were designed using primer3 (http://primer3.ut.ee) and mfold (http://unafold.rna.albany.edu/?q=mfold) to obtain amplicons with a melting temperature (Tm) of 58–62 °C, lacking a secondary structure at Tm 60 °C, and with an amplicon size of 50–150 bp. Primer sequences can be found in Table 2. Forty nanograms of cDNA was used for each qPCR reaction with a SYBR Green containing supermix (Bioline) on a Bio-Rad C1000 Touch Thermal Cycler (Bio-Rad, Hertfordshire, UK) and performed in duplicate. PCR cycle parameters were as follows: 95 °C (10 min), followed by 45 cycles of 95 °C (15 s), 60 °C (15 s) and 72 °C (15 s). Following this, a melt curve was produced with readings taken every 1 °C from 65 to 95 °C. Analysis of three candidate reference genes (18 s rRNA, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and β-actin) was performed using the comprehensive gene ranking program RefFinder https://www.heartcure.com.au/reffinder/ (Xie et al. 2012) (data not shown). Consequently, relative gene expression levels were normalised with the housekeeping gene 18 s rRNA using the 2−ΔΔCt method (Livak and Schmittgen 2001). qPCR data was presented as fold change in gene expression of the inflammatory cytokine treated cells, compared with untreated controls.
IL-6 ELISA
The IL-6 concentrations in conditioned media obtained from three biological replicates of tenocytes (between P4 and P8) stimulated with or without inflammatory cytokines for 72 h were measured by an equine IL-6 ELISA kit with an additional ancillary kit (both R&D systems) according to the manufacturer’s instructions. Each biological replicate was measured in duplicate on a Tecan plate reader (Infinite M Plex; Tecan). Colorimetric detection was performed at 450 nm with a background correction read at 540 nm. Optical density values were converted to picograms per microliter concentrations utilising a seven-point standard curve and data reduction with four-parameter logistic regression.
IL-6 treatment of equine tenocytes
As IL-6 production was found to be increased following exposure to the inflammatory cytokines, the effect of IL-6 on P65 translocation, tenocyte 3-D collagen gel contraction and 2-D gene expression was determined. Three biological replicates of tenocytes between P2 and P9 were used in these experiments and exposed to 10 ng/mL (John et al. 2010; Martincuks et al. 2017) of recombinant human IL-6 (PeproTech). Unstimulated cells served as controls.
IL-1RA treatment of equine tenocytes
We determined the effect of IL-1RA on attenuating adverse inflammatory responses by measuring their effects on cytokine mediated P65 translocation, tenocyte 3-D collagen gel contraction and 2-D gene expression. Three (four for 3-D culture experiments) biological replicates of tenocytes between P4 and P10 were used in these experiments and exposed to either IL-1β (17 ng/mL) alone or a combination of IL-1β (17 ng/mL), TNFα (10 ng/mL) and IFN-γ (100 ng/mL) in the presence or absence of 100 ng/mL recombinant human IL-1RA (Sigma-Aldrich) (McClellan et al. 2019a). Unstimulated cells and cells treated with IL-1RA alone were used as controls. P65 translocation was measured after 1 h of cytokine exposure, 2-D gene expression after 72 h of cytokine exposure and 3-D collagen gel contraction over 14 days of cytokine exposure (all as described in the sections above).
BM-MSC culture
BM-MSCs were derived from bone marrow aspirates retrieved from a total of four Welsh mountain ponies immediately following euthanasia for reasons unrelated to this study with the approval of the Animal Health Trust Ethical Review Committee (AHT_02_2012). The ponies were aged between 2 and 5 years old. Three were male, and one was female. All BM-MSCs used were formally processed and cryopreserved as described previously (Guest et al. 2008; Paterson et al. 2014). Cells were used with approval of the Royal Veterinary College Clinical Research Ethical Review Board (URN 2021 2035–2). BM-MSCs were cultured in growth media (as detailed above) at 37 °C and 5% CO2. Medium was replenished every 2–3 days, and the cells passaged using 0.25% trypsin–EDTA (Sigma-Aldrich) every 3–7 days prior to reaching maximum confluency. BM-MSC-conditioned media was obtained by culturing the BM-MSCs on 10 cm2 plates until 70–80% confluent. The culture media was then removed and replenished with fresh growth media which was collected after 48 h of incubation. This “BM-MSC-conditioned media” was passed through a 0.22-μm filter (Sigma-Aldrich) to remove cell debris and was used immediately.
Flow cytometry
Flow cytometry was performed using two biological replicates of BM-MSCs between P5 and P7 (Guest et al. 2022). BM-MSCs were fixed with 3% paraformaldehyde for 20 min and then washed three times with phosphate-buffered saline (PBS). Blocking was carried out using 2.5% NHS for 20 min and 1 × 106 cells were incubated with primary antibodies (see Table 3) or corresponding concentrations of isotype control (mouse IgG, Vector Laboratories) for 45 min at 4 °C. Following washing with PBS, the BM-MSCs were incubated with goat-anti mouse FITC (1:200 dilution, ab7064, Abcam) for 45 min at 4 °C and then washed three times with PBS. A maximum of 10,000 events were acquired on a flow cytometer (BD LSRFortessa x-20 4 laser system, BD Biosciences). The data was subsequently analysed using FlowJo (version 10.8.1). Events were gated using forward versus side scatter of unlabelled cells to exclude dead cells and debris (R1). These events were then gated using forward scatter height versus scatter area to remove doublets (R2). Two percent of the remaining events in the isotype control and everything to the right were included in a new gate (R3) by FITC area versus forward scatter height. The overlayed antibody events gated within the corresponding isotype controls R3 gate were considered positive.
BM-MSC and tenocyte co-culture in 2-D
Two-dimensional (2-D) BM-MSC/tenocyte co-culture experiments were performed using three biological replicates of BM-MSCs (P4–P5) and four biological replicates of tenocytes (P3–P10). Six-well transwell culture plates with a 0.4-μm membrane pore size (Corning, Flintshire, UK) were used to physically separate the BM-MSCs and tenocytes. Tendon cells (2 × 105 cells/well) were seeded into each well (outer chamber) of a transwell culture plate and allowed to attach at 37 °C and 5% CO2 for 24 h. For immunofluorescence, each well was lined with gelatin-coated (Sigma-Aldrich) glass coverslips prior to seeding tenocytes. After 24 h, BM-MSCs which had been mitotically inactivated for 2 h with 10 μg/mL mitomycin C (MMC) from Streptomyces caespitosus (Sigma-Aldrich) were seeded into the inner chamber of each well (1 × 105 cells/well). Growth media alone (no cytokine control) or containing TNFα (10 ng/mL), IL-1β (17 ng/mL) and IFN-γ (100 ng/mL) was then added to the inner and outer chamber of each well. For immunofluorescence, tenocytes seeded on glass coverslips were fixed at room temperature using 3% paraformaldehyde (Sigma-Aldrich) after 1 h of adding the BM-MSCs. This time point was optimised in the earlier experiments for immunofluorescent analysis of P65 nuclear translocation (as described earlier). For qPCR, RNA was harvested from tenocytes using Tri-reagent (Sigma-Aldrich) after 72 h of co-culture (as optimised previously (McClellan et al. 2019a) and used in the earlier experiments above). In addition to the no cytokine controls but the presence of BM-MSCs, tenocytes cultured in the absence of BM-MSCs were used as controls (with or without the presence of inflammatory cytokines).
BM-MSC-conditioned media and tenocyte co-culture in 2-D
BM-MSC-conditioned media/tenocyte co-culture experiments were performed using three biological replicates of BM-MSCs (P4–P5) and four biological replicates of tenocytes (P3–P10). Tenocytes (2 × 105 cells/well) were seeded into 6-well plates and allowed to attach at 37 °C and 5% CO2 for 24 h. For immunofluorescence, 24-well plates were lined with gelatin-coated (Sigma-Aldrich) glass coverslips prior to seeding tenocytes. After 24 h, previously prepared 48-h BM-MSC-conditioned media alone (no cytokine control) or containing TNFα (10 ng/mL), IL-1β (17 ng/mL) and IFN-γ (100 ng/mL) was then added to each well of tenocytes. For immunofluorescence, tenocytes seeded on glass coverslips were fixed at room temperature using 3% paraformaldehyde (Sigma-Aldrich) after 1 h of adding the BM-MSC-conditioned media. This time point was optimised in the earlier experiments (see above) for immunofluorescent analysis of P65 nuclear translocation (as described previously). For qPCR, RNA was harvested from tenocytes using Tri-reagent (Sigma-Aldrich) after 72 h of culture. In addition to the no cytokine controls but the presence of conditioned media, tenocytes cultured in the absence of BM-MSC-conditioned media were used as controls (with or without the inflammatory cytokines).
BM-MSC/BM-MSC-conditioned media and tenocyte co-culture in 3-D
3-D co-culture experiments were performed using three biological replicates of BM-MSCs (P4–P5) and four biological replicates of tenocytes (P4–P6). Pairs of minutien pins were inserted into silicone inserts (2 cm × 2 cm square) produced by punching a silicone-coated 10-cm plate with a metal cutter. Individual inserts were placed centrally into each well of a 6-well plate and the prepared collagen-tenocyte suspension seeded as previously described with each insert holding two sets of pins. Once the constructs had set, 4 mL of growth media (with or without cytokines as described above) was added to the wells with or without 2.5 × 105 MMC mitotically inactivated BM-MSCs and maintained at 37 °C and 5% CO2 for 14 days. BM-MSCs attached to the tissue culture surface around the outside of the silicone inserts. BM-MSC morphology was monitored daily, with media replaced every 3–4 days. On day 14, BM-MSCs were fixed with 3% paraformaldehyde (Sigma-Aldrich) for 20 min prior to staining with 1% Crystal Violet solution (Sigma-Aldrich) for 20 min to observe cell distribution. Alternatively, BM-MSC-conditioned media was used, collected as described earlier. In both cases, tenocyte cell viability and the degree of tenocyte collagen gel contraction was measured as described in the previous section “Three-dimensional cell culture of tenocytes”.
Statistical analysis
Statistical analysis was performed using Excel (Microsoft™) and SPSS (version 28.0.0.0.; IBM, UK). All data sets were tested to ensure Gaussian distribution using the Shapiro-Wilks normality test and equal variance using Levene’s test. For comparisons of two groups, independent Student’s t-tests were used. Data with more than two groups was analysed using a one-way ANOVA with post hoc Tukey. Where data was not normally distributed, a non-parametric Kruskal Wallis test was performed followed by Dunn’s pairwise comparisons. In all cases, a p value of < 0.05 was considered statistically significant.
Results
IL-1β and TNFα, but not IFN-γ, activate NF-κB P65 in tenocytes
Initially, dose and time response experiments were carried out to establish optimal inflammatory cytokine conditions (Fig. S1). At these optimal concentrations, there was no significant effect of the cytokines on tenocyte proliferation or viability (Fig. S2). Furthermore, unstimulated tenocytes expressed the receptors for IL-1β (McClellan et al. 2019a), IFN-γ and TNFα at the protein level (Fig. S3). Immunofluorescence demonstrated that IL-1β and TNFα, but not IFN-γ, induced significant P65 nuclear translocation compared to the no cytokine control within 1 h of stimulation (Fig. 2a–a’’’’’ and b). A further significant difference in the amount of nuclear P65 was evident between IL-1β alone and TNFα in combination with IFN-γ, but none of the other combinations (Fig. 2b). To confirm that P65 nuclear translocation resulted in P65 DNA binding activity, a DNA-binding ELISA was performed. IFN-γ failed to induce P65 DNA binding, whereas treatment with TNFα and IL-1β individually, and all 3 cytokines in combination, led to a 1.6- to 1.8-fold induction of P65 DNA binding compared to the no cytokine control (Fig. 2c). Although these increases were not statistically significant, a binding-site competitor oligonucleotide was able to reverse the trend induced by all three cytokines suggesting binding was occurring.

The localisation and DNA binding of NF-κB P65 in response to inflammatory cytokine stimulation. (a–a’’’’’) Immunofluorescence staining of NF-κB P65 in tenocytes following 1 h of IL-1β, TNFα and/or IFN-γ stimulation. DAPI staining of the nucleus is shown in blue. Images are representative of three biological replicates. Scale bar = 50 µm. (b) Quantification of the relative nuclear fluorescent intensity of NF-κB P65 immunofluorescence following 1 h of cytokine stimulation. Data shown as fold change compared to the no cytokine control. Error bars represent the S.E.M of three measurements from each of three biological replicates. Asterisk indicates the fold change in nuclear fluorescent intensity is significantly different to the no cytokine control and IFN-γ (p < 0.05). Number sign indicates the fold change in nuclear fluorescent intensity is significantly different between conditions (p < 0.05). (c) NF-κB P65 DNA binding ELISA following 1 h of cytokine stimulation. Results are also shown from combining one biological replicate of tenocytes stimulated with all three cytokines with a wild-type consensus oligonucleotide (competitor of DNA binding) and a mutated consensus oligonucleotide (no effect on DNA binding). Error bars represent the S.E.M of three biological replicates. Cells in these experiments were between P3 and P10
Nuclear translocation of STAT1, JNK and P38 MAPK in response to IL-1β, TNFα and IFN-γ stimulation in tenocytes
Immunofluorescent staining showed basal nuclear STAT1 localisation in tenocytes under no cytokine control conditions (Fig. 3a–a’’’’’) which was significantly increased by IFN-γ, but not by TNFα and IL-1β or a combination of all three cytokines (Fig. 3d). In contrast, immunofluorescent staining for JNK did not demonstrate activation by any cytokine combination (Fig. 3b–b’’’’’ and e). The P38 MAPK signalling pathway was also investigated; however, minimal P38 MAPK was visualised in equine tenocytes (Fig. 3c–c’’’’’’). We confirmed the P38 MAPK antibodies cross-reactivity, previously shown to cross-react with equine cells (Gardner et al. 2016), by demonstrating positive staining in spontaneously differentiated equine induced pluripotent stem cells. Additionally, P38 MAPK has formally been shown to be expressed at the mRNA level in equine tenocytes (Paterson et al. 2020). Western blot analysis confirmed that the antibodies used to detect NF-κB P65, JNK and STAT1 recognise the specific equine proteins (Fig. S4).

The localisation and activation of other inflammatory pathways in response to inflammatory cytokine stimulation. Immunofluorescence staining of STAT1 (a–a’’’’’), JNK (b–b’’’’’) and P38 MAPK (c–c’’’’’’) in tenocytes following IL-1β, TNFα and/or IFN-γ stimulation. c’’’’’’ shows staining of equine induced pluripotent stem cells and acts as a positive control. DAPI staining of the nucleus is shown in blue. All images are representative of three biological replicates. Scale bar = 50 µm. Quantification of the relative nuclear fluorescent intensity of STAT1 (d) and JNK (e) following cytokine stimulation is also shown. Data shown as fold change compared to the no cytokine control. Error bars represent the S.E.M of three measurements from each of three biological replicates. Asterisk indicates the fold change in nuclear fluorescent intensity is significantly different to the no cytokine control (p < 0.05). Cells in these experiments were between P4 and P8
IL-1β, TNFα and IFN-γ work synergistically to inhibit 3-D collagen gel contraction by tenocytes
The degree of 3-D collagen gel contraction by tenocytes was significantly impaired by IL-1β, while TNFα and IFN-γ alone showed no significant effect (Fig. 4a). However, when applied in combination, all three cytokines caused the greatest impairment in 3-D collagen gel contraction, with significant differences to the no cytokine control, TNFα alone and IFN-γ alone from day 4 onwards, and to IL-1β from day 11 onwards, suggesting a synergistic effect. Whilst IFN-γ alone had little effect on 3-D collagen gel contraction, we show that it can potentiate the effects of the other cytokines (Fig. 4b and c–c’’’’’’’). In combination, IFN-γ with TNFα demonstrated a significantly greater inhibitory effect on 3-D collagen gel contraction when compared to TNFα alone from day 4 onwards. IFN-γ with IL-1β showed a significantly greater inhibitory effect on 3-D collagen gel contraction when compared to IL-1β alone but from day 11 onwards. The inhibitory effect on collagen gel contraction was also significantly greater when IFN-γ was added to TNFα and IL-1β from day 4 onwards. Differences in 3-D collagen gel contraction were not due to cytokine effects on tenocyte cell survival which show no significant differences at day 14, although there is a trend for the combination of all three cytokines to reduce survival compared to the no cytokine control (Fig. S5a). Furthermore, the contraction ability of the 3-D collagen gels is not due to the collagen itself as cell-free constructs fail to contract over 14 days (Fig. S5b).

The effect of inflammatory stimulation on 3-D collagen gel contraction by tenocytes. (a) Inflammatory cytokines used in combination reduce collagen gel contraction by tenocytes to a greater extent than when used individually. Asterisk indicates the degree of collagen gel contraction is significantly different to the no cytokine control (p < 0.05). Number sign indicates the degree of collagen gel contraction is significantly different to all 3 cytokines in combination (p < 0.05). (b) IFN-γ potentiates the effect of TNFα and IL-1β on 3-D collagen gel contraction by tenocytes. Number sign indicates the degree of collagen gel contraction is significantly different between indicated conditions at p < 0.05. All error bars represent the S.E.M of four independent biological replicates of tenocytes. Representative images of 3-D collagen gel contraction at day 14 are shown in c–c’’’’’’’. Scale bar = 5 mm. Cells in these experiments were between P4 and P10
Inflammatory cytokines alter tendon-associated gene expression in tenocytes
qPCR analysis was performed to examine changes in tendon-associated gene expression in tenocytes following 72 h simulation with IFN-γ, TNFα and/or IL-1β (Fig. 5a). Treatment of tenocytes with IL-1β, TNFα or IFN-γ alone or all three cytokines in combination resulted in significant downregulation of SCX, COL1A1 and COMP expressions. In contrast, TNC was upregulated, although this was only significant for IL-1β. The only significantly potentiating effect of adding IFN-γ was for COL1A1, which was significantly downregulated when IFN-γ was combined with IL-1β, compared to IL-1β alone (data not shown). There was a tendency for all 3 cytokines in combination to have a greater effect in altering some tendon-associated gene expression than when the cytokines were used individually; however, this was not significant for any gene examined.

Inflammatory cytokines alter gene expression in tenocytes. Fold change in tendon-associated (a) and MMP (b) gene expression in tenocytes following stimulation with IFN-γ, TNFα and/or IL-1β compared to the no cytokine control. Asterisk indicates significant difference relative to no cytokine control p < 0.05. Number sign indicates a significant difference between indicated cytokine conditions (p < 0.05). Error bars represent the S.E.M of three biological replicates. Cells in these experiments were between P4 and P8
IL-1β and TNFα, but not IFN-γ, increase MMP gene expression in tenocytes
Stimulation with IFN-γ alone had minimal effects in altering MMP gene expression in tenocytes, whereas stimulation of tenocytes with IL-1β alone, TNFα alone and all three cytokines in combination resulted in substantial increases in MMP expression (Fig. 5b). In general, TNFα produced smaller changes in MMP expression, exhibiting little impact on MMP2 and MMP9 expression and upregulating MMP1, MMP3 and MMP13 to a lesser degree than IL-1β alone and all three cytokines in combination. Despite the MMPs being consistently upregulated in all replicates, due to the variation in the fold increase between biological replicates, not all changes were statistically significant (see Fig. 5 for significantly different levels).
IL-6 production is induced by exposure to IL-1β, but IL-6 alone does not activate NF-κB signalling and has no effect on 3-D collagen gel contraction or gene expression by tenocytes
Secretion of IL-6 by tenocytes was significantly increased by IL-1β alone and all three cytokines in combination, whereas stimulation with TNFα alone and IFN-γ alone caused minimal increases in IL-6 secretion (Fig. 6a). The impact of IL-6 on responses of tenocytes in vitro was also investigated. Immunofluorescence demonstrated 1 h of IL-6 stimulation did not simulate NF-κB P65 nuclear translocation in tenocytes (Fig. 6b–b’’). Further stimulation timepoints were examined but no NF-κB P65 nuclear translocation was seen (data not shown). In 3-D culture, IL-6 exhibited no significant inhibition in the final degree of 3-D collagen gel contraction (Fig. 6c–c’’) or tenocyte survival (Fig. 6d) compared to the no cytokine control. Finally, IL-6 had minimal effects in altering tendon-associated and MMP gene expression (Fig. 6e and f).

IL-6 fails to activate NF-κB signalling and exhibits no effect on 3-D collagen gel contraction or gene expression by tenocytes. (a) Secretion of IL-6 by tenocytes is significantly increased by IL-1β and all three cytokines in combination, but not by TNFα or IFN-γ alone. (b–b’’) Immunofluorescence staining of NF-κB P65 in tenocytes following 1 h of IL-6 stimulation. DAPI staining of the nucleus is shown in blue. Images are representative of three biological replicates. Scale bar = 50 µm. (c) IL-6 has no effect on 3-D collagen gel contraction by tenocytes. Representative images of gel contraction at day 14 are shown (c’–c’’). Scale bar = 5 mm. (d) No significant differences in cell survival were found between IL-6 and no cytokine control gels. Fold change in tendon-associated (e) and MMP (f) gene expression in tenocytes following stimulation with IL-6 compared to the no cytokine control. Each experiment was performed using three biological replicates. Asterisk indicates significant difference compared to the no cytokine control (p < 0.05). Error bars represent the S.E.M of three biological replicates. Cells in these experiments were between P2 and P9
IL-1RA fails to protect tenocytes when IL-1β, TNFα and IFN-γ are used in combination
IL-1RA blocked the effect of IL-1β on 3-D collagen gel contraction by tenocytes (Fig. 7a) as published previously (McClellan et al. 2019a). However, IL-1RA was unable to rescue collagen gel contraction, or nuclear translocation of NF-κB (Fig. 7b–b’’’’), when IFN-γ, TNFα and IL-1β were used in combination. Gene expression analysis demonstrated IL-1RA alone had no significant effect on tendon-associated or MMP gene expression in tenocytes. Although IL-1RA can fully rescue IL-1β induced 3-D contraction by tenocytes, it did not fully rescue gene expression. While a trend for rescue was seen for most genes, this rescue effect was only significant for SCX, MMP8 and MMP13 (Fig. 7c and d). In comparison, IL-1RA demonstrated minimal impact in rescuing the combined effects of all three cytokines on tendon-associated gene expression. However, IL-1RA did appear to reduce the adverse effects a combination of all three cytokines has upon MMP expression, although this effect was not significant for any gene examined.

IL-1RA fails to protect tenocytes when stimulated with IL-1β, TNFα and IFN-γ in combination. (a) IL-1RA (RA) is able to block the effects of IL-1β alone, but not the effects of a combination of all three cytokines on 3-D collagen gel contraction by tenocytes. Asterisk indicates the final degree of collagen gel contraction is significantly different to the no cytokine control (p < 0.05). Error bars represent the S.E.M of four biological replicates of tenocytes. (b–b’’’’) Immunofluorescence staining of NF-κB P65 in tenocytes following 1 h of IL-1β, TNFα and IFN-γ stimulation with or without IL-1RA. DAPI staining of the nucleus is shown in blue. Images are representative of three biological replicates. Scale bar = 50 µm. Fold change in tendon-associated (c) and MMP (d) gene expression in tenocytes following stimulation with inflammatory cytokines and/or IL-1RA compared to the no cytokine control. Number sign indicates significant rescue effect (p < 0.05). Error bars represent the S.E.M of three biological replicates of tenocytes. Cells in these experiments were between P4 and P10
Equine BM-MSCs are unable to protect tenocytes from the negative consequences of inflammatory cytokines
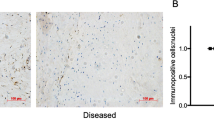
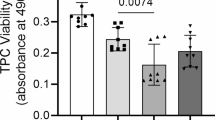
Flow cytometry analysis showed the BM-MSCs were positive for CD90 (98.2%) and CD29 (95.4%), but negative for CD14 (0.82%) and CD45 (0.26%) (Fig. S6). Immunofluorescence demonstrated that neither BM-MSCs in transwell co-culture nor BM-MSC-conditioned media were able to prevent NF-κB P65 nuclear translocation in tenocytes stimulated with IFN-γ, TNFα and IL-1β in combinations compared to the no BM-MSC control (Fig. 8a–a’’’ and b). In 3-D culture, conditioned media taken from actively proliferating BM-MSCs failed to rescue collagen gel contraction by tenocytes following IFN-γ, TNFα and IL-1β stimulations (Fig. 8c–c’). When BM-MSCs were co-cultured with the 3-D cultures of tenocytes, there was a small rescue effect; however, this was highly variable, and the 3-D collagen gels remained significantly different to the no cytokine control. Furthermore, there was no significant difference in the final degree of 3-D collagen gel contraction by tenocytes between BM-MSC culture conditions. Crystal violet staining showed no difference in BM-MSC survival between conditions at day 14 of 3-D co-culture (Fig. S7). In accordance with the 3-D collagen gel contraction data, neither BM-MSC-conditioned media nor BM-MSCs in co-culture were able to rescue adverse alterations in tendon-associated or MMP gene expression resulting from IFN-γ, TNFα and IL-1β stimulations (Fig. 8d–d’).

BM-MSCs are unable to release soluble factors which protect tenocytes from combined inflammatory stimulation. Immunofluorescence demonstrates neither BM-MSC-conditioned media (a’’) nor BM-MSCs in co-culture (a’’’) are able to prevent the nuclear translocation of NF-κB P65 in tenocytes following cytokine stimulation (a–a’’’). DAPI staining of the nucleus is shown in blue. Images are representative of four biological replicates of tenocytes. Scale bar = 50 µm. (b) Quantification of the relative nuclear fluorescent intensity of NF-κB following cytokine stimulation. Data is shown as fold change compared to the no cytokine control. Error bars represent the S.E.M of four biological replicates of tenocytes. Asterisk indicates the fold change in nuclear fluorescent intensity is significantly different to the no cytokine control (p < 0.05). (c–c’) BM-MSC-conditioned media or BM-MSCs in co-culture were unable to rescue the adverse effects inflammatory cytokines have upon 3-D collagen gel contraction by tenocytes. Representative collagen gel images are shown from day 14 (c). Asterisk indicates the degree of collagen gel contraction is significantly different to the no cytokine control (right; p < 0.05). Fold change in tendon-associated (d) and MMP (d’) gene expression in tenocytes following inflammatory cytokine stimulation and culture with no BM-MSCs, BM-MSC-conditioned media or BM-MSCs in co-culture compared to the no cytokine control. Number sign indicates significant difference between the groups (p < 0.05). Error bars represent the S.E.M of four biological replicates of tenocytes. Cells in these experiments were between P3 and P10
Discussion
Inflammation plays a key role in both normal and pathogenic tendon regeneration (Rees et al. 2014). However, our understanding of the inflammatory mechanisms underpinning inferior tendon healing is limited, meaning successful targeted therapeutics are lacking. Our previous work demonstrated IL-1β has adverse effects on equine tenocytes in vitro (McClellan et al. 2019a); however, during tendon injury in vivo, other pro-inflammatory cytokines are also present. Therefore, here we investigated the combined effects of IFN-γ, TNFα and IL-1β on inflammatory pathway activation and tendon cell properties.
Insights into the pathogenic effects of inflammatory cytokines may be obtained by studying specific transcription factors induced by those cytokines. The canonical NF-κB pathway is an important regulator of the inflammatory response, and inappropriate modulation of NF-κB has been implicated in the formation of fibrous tendon tissue (Abraham et al. 2019; Best et al. 2019). Activation of the NF-κB pathway results in the rapid translocation of the NF-κB P65 dimer from the cytoplasm to the nucleus where it modulates gene transcription. Previously, we demonstrated IL-1β stimulation induces the nuclear translocation of NF-κB P65 in equine tenocytes (McClellan et al. 2019a). Here, we demonstrate that NF-κB signalling is also activated by TNFα stimulation, but not by IFN-γ. This is in contrast to studies in HeLa S3 cells, where IFN-γ induced NF-κB DNA binding (Deb et al. 2001). This suggests that the direct activation of NF-κB by IFN-γ is restricted to specific cell lineages or types. In addition to NF-κB, the inflammatory pathways JNK, STAT1 and P38 MAPK were also examined due to their roles in activating genes involved in immune function (Gough et al. 2008; Wang et al. 2019). Despite IFN-γ, TNFα and IL-1β having little effect in the activation of JNK or P38 MAPK, IFN-γ did show upregulation of STAT1 signalling. This is in accordance with previous research, and the importance of STAT1 in IFN-γ biology has been demonstrated, as deficits in immunity in STAT1 knockout mice are notably similar to the phenotypes of IFN-γ knockout mice (Durbin et al. 1996). It is possible that other inflammatory pathways such as STAT3 and STAT5 are involved in the tendon healing response (Tarafder et al. 2017). Therefore, further research examining the phosphorylation profile of a broad array of inflammatory pathway proteins following inflammatory stimuli would be beneficial to identify additional pathways of interest.
Investigation of commonly cited tendon-associated genes revealed significant changes in expression levels following cytokine stimulation. Most notably, the expression of SCX, COL1A1 and COMP was significantly downregulated following stimulation with TNFα and IL-1β. Although not significant, the greatest downregulation was observed when all three cytokines were used in combination, suggesting a synergistic effect. These observed changes in gene expression have previously been suggested to negatively impact tenocytes and their ability to regenerate a healthy tendon matrix (McClellan et al. 2019a). We also identified considerable increases in MMP gene expression in tenocytes following stimulation with TNFα and IL-1β. Metalloproteinases play an integral part in the maintenance and repair of the extracellular matrix (ECM), and evidence suggests alterations to the synthetic-degradative equilibrium of these enzymes underlies tendon degeneration (Jones et al. 2006; Robertson et al. 2012). It has been suggested that changes in MMP gene expression are directly related to NF-κB activation, with studies identifying MMP1 (Vincenti et al. 1998), MMP3 (Borghaei et al. 2004) and MMP9 (Bond et al. 1998) as direct targets of NF-κB signalling. However, to the authors knowledge, it is unknown whether all the genes examined here are direct targets of NF-κB and further work to establish this is warranted.
The ability of tenocytes to contract 3-D collagen gels has widely been used to measure cell-mediated matrix reorganisation (Bell et al. 1979; Grinnell and Petroll 2010; Yang et al. 2015). Previously, we have demonstrated the degree of 3-D collagen gel contraction by tenocytes is significantly impaired by IL-1β (McClellan et al. 2019a). Our results demonstrate multi-cytokine stimulation with IL-1β, TNFα and IFN-γ significantly impairs 3-D collagen gel contraction by tenocytes to a greater extent than when TNFα or IL-1β are used individually. Taken together with the increased MMP gene expression evidenced following cytokine stimulation, these results suggest tenocytes in an inflammatory environment contribute to matrix degeneration rather than regeneration. However, MMP protein activity, tissue inhibitor of metalloproteinase (TIMP) expression, MMP:TIMP ratios and specific collagen type ratios remain to be measured to determine if this is the case.
Of note, our results revealed that IFN-γ, which did not activate NF-κB, had no significant effect on 3-D collagen gel contraction or MMP gene expression. We have previously demonstrated that IFN-γ results in increased expression of MHC I in tenocytes (McClellan et al. 2019b), and this may be mediated by the activation of STAT1 (Lieberman et al. 2004). Additionally, our results further indicate that IFN-γ can synergize with IL-1β and TNFα to cause a greater effect on 3-D collagen gel contraction than when the cytokines are used individually. Previous reports in a vascular endothelium cell line have found that although IFN-γ on its own does not significantly induce NF-κB, it potentiates TNFα-induced NF-κB nuclear translocation (Cheshire and Baldwin 1997). The mechanisms behind this synergistic effect may be due to a cooperative relationship between STAT1 and NF-κB, where certain genes are only expressed when both transcription factors are activated simultaneously (Hiroi and Ohmori 2005).
The secretion of interleukin 6 (IL-6) was significantly upregulated by tenocytes following stimulation with IL-1β and a combination of all three cytokines. Interestingly, IL-6 secretion has been well documented as a downstream effector of NF-κB activation and is elevated during tendon ruptures (Jové et al. 2005; Novotny et al. 2008; Millar et al. 2016; Liu et al. 2017). Nevertheless, the action of IL-6 in tendinopathy is complex. IL-6 is considered as a classical pro-inflammatory cytokine due to its role in macrophage activation (Stolk et al. 2017). However, it is also known to possess anti-inflammatory properties by promoting the secretion of factors such as IL-10 and IL-1RA (Lin et al. 2006). Our results indicate that IL-6 stimulation alone has little effect on tendon-associated gene expression and 3-D collagen gel contraction by tenocytes. Interestingly, like IFN-γ, IL-6 also failed to activate NF-κB signalling. Overall, this suggests IL-6 has limited catabolic effects on the parameters investigated in this in vitro study. This finding is similar to previous research in tenocytes (John et al. 2010), which, however, is in stark contrast to work conducted in bovine articular chondrocytes demonstrating IL-6 upregulates the expression of MMPs (Legendre et al. 2005). Nevertheless, the expression of IL-6 has been shown to be imperative for normal tendon healing (Lin et al. 2006); therefore, it is clear that more detailed analyses into its immunoregulatory role are needed.
Our study also evaluated whether currently used inflammatory targeting treatments were able to protect tenocytes from a multi-cytokine environment in vitro. IL-1RA is a naturally occurring inhibitor of IL-1β and functions by competing with IL-1β for binding to the IL-1 receptor 1 (Fredberg and Ostgaard 2009). IL-1RA-based therapies, such as Anakinra, have been advocated for the treatment of inflammatory disorders such as rheumatoid arthritis (Bresnihan et al. 1998; Hallegua and Weisman 2002; Dragoljevic et al. 2020). Since IL-1β is implicated in tendinopathy (Hosaka et al. 2002; Morita et al. 2017), IL-1RA may have therapeutic potential to improve tendon regeneration (Geburek et al. 2015). Nonetheless, anakinra injection in human patients with chronic Achilles tendinopathy demonstrated little beneficial effect, with no significant reduction seen in pain or intratendinous blood flow, and increased tendon thickness after 12 weeks of treatment (Fredberg and Ostgaard 2009). Our results do suggest some beneficial effects of IL-1RA, but only when IL-1β was used alone. Treatment with IL-1RA was able to fully rescue the adverse effects of IL-1β on 3-D collagen gel contraction by tenocytes but was unable to fully rescue tendon-associated gene expression. This suggests that we were unable to completely block IL-1β signalling, and based on the previous literature, it is possible a higher concentration of IL-1RA may have been needed (Hallegua and Weisman 2002). Nevertheless, our work demonstrates that IL-1RA cannot prevent the adverse effects of multiple cytokine stimulation on tenocytes. Since the primary goal of IL-1RA is to inhibit the inflammatory cascade induced by IL-1β, instead of directly aiding tissue regeneration, future treatment options that directly target the inflammatory cascades activated by both IL-1β and TNFα may be a more effective treatment option.
It is well established that BM-MSCs possess both anti-inflammatory and immunosuppressive properties (Aggarwal and Pittenger 2005; Zhang et al. 2017). However, the detailed mechanisms by which BM-MSCs modulate inflammation to improve tendon regeneration remain unclear. Potential theories include the following: BM-MSCs modulate the activity of macrophages (Kim and Hematti 2009; Manning et al. 2015), BM-MSCs secrete factors which modulate tenocyte activity (van Buul et al. 2012) or that BM-MSCs inactivate pro-inflammatory cytokines directly (Viganò et al. 2019). Using a BM-MSC/tenocyte co-culture model, we demonstrated that BM-MSCs were unable to directly modulate tenocyte activity and protect these cells from inflammatory stimuli. This result is supported by previous studies in mice where MSCs derived from adipose tissue (ASCs) were unable to protect tendon fibroblasts from IL-1β stimulation (Manning et al. 2015). Although the in vitro methods used here provide a well-controlled environment to determine tenocyte responses to specific cytokine combinations and BM-MSC interventions, it does not fully reproduce the complexities of tendinopathy in vivo where multiple cell types, including immune cells, are present and cells have direct cell-to-cell contact. Manning et al. (2015) found that ASCs suppressed the negative effects of M1 (pro-inflammatory) macrophages on tendon fibroblasts by inducing a phenotypic switch to an M2 (anti-inflammatory) macrophage. Therefore, our results are more consistent with BM-MSCs exhibiting their beneficial effects via circulating macrophages rather than the tenocytes themselves. Consequently, immune cell manipulation towards a regenerative phenotype may provide a promising therapeutic option for superior tendon healing.
Under pro-inflammatory conditions, BM-MSCs become “primed” and upregulate major histocompatibility complex (MHC) expression in order to enhance their immunomodulatory properties (Noronha et al. 2019). Here, we did not “prime” the BM-MSCs prior to use, and the conditioned media came from BM-MSCs that had not been exposed to inflammatory cytokines therefore this may have affected our results. However, the BM-MSCs used in co-culture with the tenocytes were in direct contact with IFN-γ, TNFα and IL-1β throughout the experiments. IFN-γ in particular is identified as one of the most potent activators of BM-MSCs immunomodulatory properties (Krampera et al. 2006; De Witte et al. 2016); therefore, we do not afford the lack of “priming” to the lack of protective effects afforded to tenocytes here. Furthermore, we also performed the work using BM-MSC-conditioned media in 10% serum. This was because the tenocytes are unable to contract a 3-D collagen gel over a 2-week period in low serum concentrations. However, the high serum levels we used may have masked the effects of components produced by the BM-MSCs.
There are other limitations to our study. For instance, our gene expression experiments were performed using cells cultured in 2-D monolayer to enable us to make direct comparisons to our previous work on IL-1β (McClellan et al. 2019a). However, we have recently reported that the distinct gene expression profiles of tenocytes are better preserved in a 3-D culture environment (Paterson et al. 2020) and future work in 3-D may enable the capture of differences between experimental groups that may be lost in conventional 2-D culture systems. Additionally, our gene expression experiments were performed using cells up to passage 10, and it has been suggested that cells may change their gene expression profiles with increasing passage. However, both ourselves and other researchers have demonstrated that although changes in tendon-associated gene expression are found to occur between P0 and some of the very early passages, they then remain stable up to passage 10, therefore, we feel our results do provide an accurate in vitro model (Jo et al. 2019; Liao et al. 2020; Paterson et al. 2020). Furthermore, here we added the inflammatory stimulus at the same time as the protective therapy (IL-1RA or BM-MSCs), whereas in clinical cases, this would not be the case. This approach was chosen because the 3-D cultures can only be maintained for 2 weeks (Barsby et al. 2014) and we wanted to provide the maximum opportunity for the protective therapy to have an effect. Future studies aiming to identify novel therapies for tendinopathy should consider adding the therapy subsequently to inflammation. Finally, the inflammatory cytokine concentrations used here are based on previous in vitro studies of a similar type. However, it is unknown if these concentrations correlate to the in vivo environment due to a lack of investigation of cytokine concentrations in equine tendon tissue (Ellis et al. 2022).
In conclusion, we have shown that TNFα and IL-1β both activate NF-κB signalling and together induce negative effects in tenocytes which cannot be rescued by conventional therapies such as IL-1RA or BM-MSCs. A superior approach for treatment of tendinopathy may therefore involve the targeting of specific, shared signalling pathways such as NF-κB.
Data availability
All relevant data are within the manuscript and its supplementary information files.
References
Abraham AC, Shah SA, Golman M, Song L, Li X, Kurtaliaj I, Akbar M, Millar NL, Abu-Amer Y, Galatz LM, Thomopoulos S (2019) Targeting the NF-kB signaling pathway in chronic tendon disease. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aav4319
Abraham AC, Shah SA, Thomopoulos S (2017) Targeting Inflammation in Rotator Cuff Tendon Degeneration and Repair. Tech Shoulder Elbow Surg 18:84–90
Aggarwal S, Pittenger MF (2005) Human mesenchymal stem cells modulate allogeneic immune cell responses. Blood 105:1815–1822
Alfredson H, Lorentzon R (2002) Chronic tendon pain: no signs of chemical inflammation but high concentrations of the neurotransmitter glutamate. Implications for Treatment? Curr Drug Targets 3:43–54
Barsby T, Bavin EP, Guest DJ (2014) Three-dimensional culture and transforming growth factor beta3 synergistically promote tenogenic differentiation of equine embryo-derived stem cells. Tissue Eng - Part A 20:2604–2613
Barsby T, Guest D (2013) Transforming growth factor beta3 promotes tendon differentiation of equine embryo-derived stem cells. Tissue Eng - Part A 19:2156–2165
Bavin EP, Atkinson F, Barsby T, Guest DJ (2017) Scleraxis is essential for tendon differentiation by equine embryonic stem cells and in equine fetal tenocytes. Stem Cells Dev 26:441–450
Becerra P, Valdés Vázquez MA, Dudhia J, Fiske-Jackson AR, Neves F, Hartman NG, Smith RKW (2013) Distribution of injected technetium99m-labeled mesenchymal stem cells in horses with naturally occurring tendinopathy. J Orthop Res 31:1096–1102
Bell E, Ivarsson B, Merrill C (1979) Production of a tissue-like structure by contraction of collagen lattices by human fibroblasts of different proliferative potential in vitro. Proc Natl Acad Sci USA 76:1274–1278
Berkoff DJ, Kallianos SA, Eskildsen SM, Weinhold PS (2016) Use of an IL1-receptor antagonist to prevent the progression of tendinopathy in a rat model. J Orthop Res 34:616–622
Best KT, Lee FK, Knapp E, Awad HA, Loiselle AE (2019) Deletion of NFKB1 enhances canonical NF-κB signaling and increases macrophage and myofibroblast content during tendon healing. Sci Rep 9:1–11
Bond M, Fabunmi RP, Baker AH, Newby AC (1998) Synergistic upregulation of metalloproteinase-9 by growth factors and inflammatory cytokines: an absolute requirement for transcription factor NF-κB. FEBS letters 435:29–34
Borghaei RC, Rawlings PL Jr, Javadi M Woloshin J (2004) NF-κB binds to a polymorphic repressor element in the MMP-3 promoter. Biochem Biophys Res Commun 316:182–188
Bresnihan B, Alvaro-Gracia JM, Cobby M, Doherty M, Domljan Z, Emery P, Nuki G, Pavelka K, Rau R, Rozman B, Watt I, Williams B, Aitchison R, McCabe D, Musikic P (1998) Treatment of rheumatoid arthritis with recombinant human interleukin-1 receptor antagonist. Arthritis Rheum 41:2196–2204
Cassel M, Baur H, Hirschmüller A, Carlsohn A, Fröhlich K, Mayer F (2015) Prevalence of Achilles and patellar tendinopathy and their association to intratendinous changes in adolescent athletes. Scand J Med Sci Sports 25:e310–e318. https://doi.org/10.1111/sms.12318
Cheshire JL, Baldwin AS (1997) Synergistic activation of NF-κB by tumor necrosis factor alpha and gamma interferon via enhanced IκBα degradation and de novo IκBβ degradation. Mol Cell Biol 17:6746–6754
Dakin SG, Dudhia J, Smith RKW (2014) Resolving an inflammatory concept: The importance of inflammation and resolution in tendinopathy. Vet Immunol Immunopathol 158:121–127
Dakin SG, Werling D, Hibbert A, Abayasekara DRE, Young NJ, Smith RKW, Dudhia J (2012) Macrophage sub-populations and the lipoxin A 4 receptor implicate active inflammation during equine tendon repair. PLoS ONE 7:e32333. https://doi.org/10.1371/journal.pone.0032333
De Witte SFH, Franquesa M, Baan CC, Hoogduijn MJ (2016) Toward development of imesenchymal stem cells for immunomodulatory therapy. Front Immunol. https://doi.org/10.3389/fimmu.2015.00648
Deb A, Haque SJ, Mogensen T, Silverman RH, Williams BRG (2001) RNA-dependent protein kinase PKR is required for activation of NF-κB by IFN-γ in a STAT1-independent pathway. J Immunol 166:6170–6180
Dragoljevic D, Lee MKS, Louis C, Shihata W, Kraakman MJ, Hansen J, Masters SL, Hanaoka BY, Nagareddy PR, Lancaster GI, Wicks IP, Murphy AJ (2020) Inhibition of interleukin-1β signalling promotes atherosclerotic lesion remodelling in mice with inflammatory arthritis. Clin Transl Immunol 9:1–9. https://doi.org/10.1002/cti2.1206
Durbin JE, Hackenmiller R, Simon MC, Levy DE (1996) Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 84:443–450
Dyson SJ (2004) Medical management of superficial digital flexor tendonitis: a comparative study in 219 horses (1992–2000). Equine Vet J 36:415–419
Ellis IM, Schnabel LV, Berglund AK (2022) Defining the profile: characterising cytokines in tendon injury to improve clinical therapy. J Immunol Regen Med. https://doi.org/10.1016/j.regen.2022.100059
Fedorka CE, El-Sheikh Ali H, Scoggin KE, Loux SC, Ball BA, Troedsson MHT (2022) Tumor necrosis factor signalling during equine placental infection leads to pro-apoptotic and necroptotic outcomes. J Reprod Immunol 152:103655
Filardo G, Di Matteo B, Kon E, Merli G, Marcacci M (2018) Platelet-rich plasma in tendon-related disorders: results and indications. Knee Surg Sports Traumatol Arthrosc 26:1984–1999
Fredberg U, Ostgaard R (2009) Effect of ultrasound-guided, peritendinous injections of adalimumab and anakinra in chronic Achilles tendinopathy: a pilot study. Scand J Med Sci Sports 19:338–344
Freedman BR, Adu-Berchie K, Barnum C, Fryhofer GW, Salka NS, Shetye S, Soslowsky LJ (2022) Nonsurgical treatment reduces tendon inflammation and elevates tendon markers in early healing. J Orthop Res. https://doi.org/10.1002/jor.25251
Gardner AK, Kelly CS, van Eps AW, Burns TA, Watts MR, Belknap JK (2016) Mitogen-activated kinase pathway activation in epidermal lamellae in the acute stages of carbohydrate overload laminitis models and the effect of regional deep hypothermia on signalling pathways. Equine Vet J 48:633–640
Gaspar D, Spanoudes K, Holladay C, Pandit A, Zeugolis D (2015) Progress in cell-based therapies for tendon repair. Adv Drug Deliv Rev 84:240–256
Geburek F, Lietzau M, Beineke A, Rohn K, Stadler PM (2015) Effect of a single injection of autologous conditioned serum (ACS) on tendon healing in equine naturally occurring tendinopathies. Stem Cell Res Ther 6:1–14
Gehwolf R, Schwemberger B, Jessen M, Korntner S, Wagner A, Lehner C, Weissenbacher N, Tempfer H, Traweger A (2019) Global responses of IL-1β-primed 3D tendon constructs to treatment with pulsed electromagnetic fields. Cells. https://doi.org/10.3390/cells8050399
Godwin EE, Young NJ, Dudhia J, Beamish IC, Smith RKW (2012) Implantation of bone marrow-derived mesenchymal stem cells demonstrates improved outcome in horses with overstrain injury of the superficial digital flexor tendon. Equine Vet J 44:25–32
Gomiero C, Bertolutti G, Martinello T, Van Bruaene N, Broeckx SY, Patruno M, Spaas JH (2016) Tenogenic induction of equine mesenchymal stem cells by means of growth factors and low-level laser technology. Vet Res Commun 40:39–48
Gough DJ, Levy DE, Johnstone RW, Clarke CJ (2008) IFNγ signaling-does it mean JAK-STAT? Cytokine Growth Factor Rev 19:383–394
Grinnell F, Petroll WM (2010) Cell motility and mechanics in three-dimensional collagen matrices. Annu Rev Cell Dev Biol 26:335–361
Guest DJ, Dudhia J, Smith RKW, Roberts SJ, Conzemius M, Innes JF, Fortier LA, Meeson RL (2022) Position statement: minimal criteria for reporting veterinary and animal medicine research for mesenchymal stromal/stem cells in orthopedic applications. Front Vet Sci 9. https://doi.org/10.3389/fvets.2022.817041
Guest DJ, Smith MRW, Allen WR (2010) Equine embryonic stem-like cells and mesenchymal stromal cells have different survival rates and migration patterns following their injection into damaged superficial digital flexor tendon. Equine Vet J 42:636–642
Guest D, Ousey JC, Smith MR (2008) Defining the expression of marker genes in equine mesenchymal stromal cells. Stem Cells and Cloning: Adv Appl 1. https://doi.org/10.2147/sccaa.s3824
Gupta SC, Sundaram C, Reuter S, Aggarwal BB (2010) Inhibiting NF-kB activation by small molecules as a therapeutic stategy. Biochim Biophys Acta 1799:775–787
Hallegua DS, Weisman MH (2002) Potential therapeutic uses of interleukin 1 receptor antagonists in human diseases. Ann Rheum Dis 61:960–967
Hiroi M, Ohmori Y (2005) Transcriptional synergism between NF-κB and STAT1. J Oral Biosci 47:230–242
Hosaka Y, Kirisawa R, Yamamoto E, Ueda H, Iwai H, Takehana K (2002) Localization of cytokines in tendinocytes of the superficial digital flexor tendon in the horse. J Vet Med Sci 64:945–947
Jo CH, Lim HJ, Yoon KS (2019) Characterisation of tendon-specific markers in various human tissues, tenocytes and mesenchymal stem cells. Tissue Eng Regen Med 16:151–159
John T, Lodka D, Kohl B, Ertel W, Jammrath J, Conrad C, Stoll C, Busch C, Schulze-Tanzil G (2010) Effect of pro-inflammatory and immunoregulatory cytokines on human tenocytes. J Orthop Res 28:1071–1077
Jones GC, Corps AN, Pennington CJ, Clark IM, Edwards DR, Bradley MM, Hazleman BL, Riley GP (2006) Expression profiling of metalloproteinases and tissue inhibitors of metalloproteinases in normal and degenerate human Achilles tendon. Arthritis Rheum 54:832–842
Jové M, Planavila A, Laguna JC, Vázquez-Carrera M (2005) Palmitate-induced interleukin 6 production is mediated by protein kinase C and nuclear-factor κB activation and leads to glucose transporter 4 down-regulation in skeletal muscle cells. Endocrinology 146:3087–3095
Kim J, Hematti P (2009) Mesenchymal stem cell-educated macrophages: a novel type of alternatively activated macrophages. Exp Hematol 37:1445–1453
Krampera M, Cosmi L, Angeli R, Pasini A, Liotta F, Andreini A, Santarlasci V, Mazzinghi B, Pizzolo G, Vinante F, Romagnani P, Maggi E, Romagnani S, Annunziato F (2006) Role for interferon-γ in the immunomodulatory activity of human bone marrow mesenchymal stem cells. Stem Cells 24:386–398
Lee CW, Lin CC, Lin WN, Liang KC, Luo SF, Wu CB, Wang SW, Yang CM (2007) TNF-α induces MMP-9 expression via activation of Src/EGFR, PDGFR/PI3K/Akt cascade and promotion of NF-κB/p300 binding in human tracheal smooth muscle cells. Am J Physiol - Lung Cell Mol Physiol. https://doi.org/10.1152/ajplung.00311.2006
Legendre F, Bogdanowicz P, Boumediene K, Pujol JP (2005) Role of interleukin 6 (IL-6)/IL-6R-induced signal tranducers and activators of transcription and mitogen-activated protein kinase/extracellular signal-related kinase in upregulation of matrix metalloproteinase and ADAMTS gene expression in articular chondrocytes. J Rheumatol 32:1307–1316
Liao X, Falcon ND, Mohammed AA, Paterson YZ, Mayes AG, Guest DJ, Saeed A (2020) Synthesis and formulation of four-arm polyDMAEA-siRNA polyplex for transient downregulation of collagen type iii gene expression in TGF-β1 stimulated tenocyte culture. ACS Omega 5:1496–1505
Lieberman LA, Banica M, Reiner SL, Hunter CA (2004) STAT1 plays a critical role in the regulation of antimicrobial effector mechanisms, but not in the development of Th1-type responses during toxoplasmosis. J Immunol 172:457–463
Lin TW, Cardenas L, Glaser DL, Soslowsky LJ (2006) Tendon healing in interleukin-4 and interleukin-6 knockout mice. J Biomech 39:61–69
Liu T, Zhang L, Joo D, Sun SC (2017) NF-κB signaling in inflammation. Signal Transduct Target Ther. https://doi.org/10.1038/sigtrans.2017.23
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25:402–408
Manning CN, Martel C, Sakiyama-Elbert SE, Silva MJ, Shah S, Gelberman RH, Thomopoulos S (2015) Adipose-derived mesenchymal stromal cells modulate tendon fibroblast responses to macrophage-induced inflammation in vitro. Stem Cell Res Ther 6:1–14
Marsolais D, Côté CH, Frenette J (2003) Nonsteroidal anti-inflammatory drug reduces neutrophil and macrophage accumulation but does not improve tendon regeneration. Lab Invest 83:991–999
Martincuks A, Andryka K, Küster A, Schmitz-Van de Leur H, Komorowski M, Müller-Newen G (2017) Nuclear translocation of STAT3 and NF-κB are independent of each other but NF-κB supports expression and activation of STAT3. Cell Signal 32:36–47
McClellan A, Evans R, Sze C, Kan S, Paterson Y, Guest D (2019a) A novel mechanism for the protection of embryonic stem cell derived tenocytes from inflammatory cytokine interleukin 1 beta. Sci Rep 9:1–12. https://doi.org/10.1038/s41598-019-39370-4
McClellan A, Paterson YZ, Paillot R, Guest DJ (2019b) Equine fetal, adult, and embryonic stem cell-derived tenocytes are all immune privileged but exhibit different immune suppressive properties in vitro. Stem Cells and Dev 28:1413–1423
Millar NL, Akbar M, Campbell AL, Reilly JH, Kerr SC, McLean M, Frleta-Gilchrist M, Fazzi UG, Leach WJ, Rooney BP, Crowe LAN, Murrell GAC, McInnes IB (2016) IL-17A mediates inflammatory and tissue remodelling events in early human tendinopathy. Sci Rep 6:1–11. https://doi.org/10.1038/srep27149
Millar NL, Murrell GAC, Mcinnes IB (2017) Inflammatory mechanisms in tendinopathy - towards translation. Nat Rev Rheumatol 13:110–122
Morita W, Dakin SG, Snelling SJB, Carr AJ (2017) Cytokines in tendon disease: a systematic review. Bone Jt Res 6:656–664
Noronha NDC, Mizukami A, Caliári-Oliveira C, Cominal JG, Rocha JLM, Covas DT, Swiech K, Malmegrim KCR (2019) Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res Ther. https://doi.org/10.1186/s13287-019-1259-0
Novotny NM, Markel TA, Crisostomo PR, Meldrum DR (2008) Differential IL-6 and VEGF secretion in adult and neonatal mesenchymal stem cells: Role of NFkB. Cytokine 43:215–219
Paterson YZ, Cribbs A, Espenel M, Smith EJ, Henson FMD, Guest DJ (2020) Genome-wide transcriptome analysis reveals equine embryonic stem cell-derived tenocytes resemble fetal, not adult tenocytes. Stem Cell Res Ther. https://doi.org/10.1186/s13287-020-01692-w
Paterson YZ, Rash N, Garvican ER, Paillot R, Guest DJ (2014) Equine mesenchymal stromal cells and embryo-derived stem cells are immune privileged in vitro. Stem Cell Res Ther. https://doi.org/10.1186/scrt479
Patterson-Kane JC, Becker DL, Rich T (2012) The pathogenesis of tendon microdamage in athletes: the horse as a natural model for basic cellular research. J Comp Pathol 147:227–247
Rees JD, Stride M, Scott A (2014) Tendons - time to revisit inflammation. Br J Sports Med 48:1553–1557
Robertson CM, Chen CT, Shindle MK, Cordasco FA, Rodeo SA, Warren RF (2012) Failed healing of rotator cuff repair correlates with altered collagenase and gelatinase in supraspinatus and subscapularis tendons. Am J Sports Med 40:1993–2001
Schwartz AJ, Sarver DC, Sugg KB, Dzierzawski JT, Gumucio JP, Mendias CL (2015) P38 MAPK signaling in postnatal tendon growth and remodeling. PLoS ONE. https://doi.org/10.1371/journal.pone.0120044
Smith RKW, Werling NJ, Dakin SG, Alam R, Goodship AE, Dudhia J (2013) Beneficial effects of autologous bone marrow-derived mesenchymal stem cells in naturally occurring tendinopathy. PLoS ONE. https://doi.org/10.1371/journal.pone.0075697
Sobhani S, Dekker R, Postema K, Dijkstra PU (2013) Epidemiology of ankle and foot overuse injuries in sports: a systematic review. Scand J Med Sci Sports 23:669–686
Spaas JH, De Schauwer C, Cornillie P, Meyer E, Van Soom A, Van De Walle GR (2013) Culture and characterisation of equine peripheral blood mesenchymal stromal cells. Vet J 195:107–113
Stolk M, Klatte-Schulz F, Schmock A, Minkwitz S, Wildemann B, Seifert M (2017) New insights into tenocyte-immune cell interplay in an in vitro model of inflammation. Sci Rep. https://doi.org/10.1038/s41598-017-09875-x
Sullivan CB, Porter RM, Evans CH, Ritter T, Shaw G, Barry F, Murphy JM (2014) TNFα and IL-1β influence the differentiation and migration of murine MSCs independently of the NF-κB pathway. Stem Cell Res Ther. https://doi.org/10.1186/scrt492
Tang JB, Wu YF, Cao Y, Chen CH, Zhou YL, Avanessian B, Shimada M, Wang XT, Liu PY (2016) Basic FGF or VEGF gene therapy corrects insufficiency in the intrinsic healing capacity of tendons. Sci Rep. https://doi.org/10.1038/srep20643
Tarafder S, Chen E, Jun Y, Kao K, Sim KH, Back J, Lee FY, Lee CH (2017) Tendon stem/progenitor cells regulate inflammation in tendon healing via JNK and STAT3 signaling. FASEB J 31:3991–3998
Tsai CL, Chen WC, Hsieh HL, Chi PL, Der HL, Yang CM (2014) TNF-α induces matrix metalloproteinase-9-dependent soluble intercellular adhesion molecule-1 release via TRAF2-mediated MAPKs and NF-κB activation in osteoblast-like MC3T3-E1 cells. J Biomed Sci. https://doi.org/10.1186/1423-0127-21-12
Tsuzaki M, Guyton G, Garrett W, Archambault JM, Herzog W, Almekinders L, Bynum D, Yang X, Banes AJ (2003) IL-1b induces COX2, MMP-1, -3 and -13, ADAMTS-4, IL-1 p and IL-6 in human tendon cells. J Orthop Res 21:256–264
van Buul GM, Villafuertes E, Bos PK, Waarsing JH, Kops N, Narcisi R, Weinans H, Verhaar JAN, Bernsen MR, van Osch GJVM (2012) Mesenchymal stem cells secrete factors that inhibit inflammatory processes in short-term osteoarthritic synovium and cartilage explant culture. Osteoarthr Cartil 20:1186–1196
Viganò M, Lugano G, Perucca Orfei C, Menon A, Ragni E, Colombini A, De Luca P, Randelli P, De Girolamo L (2019) Autologous microfragmented adipose tissue reduces the catabolic and fibrosis response in an in vitro model of tendon cell inflammation. Stem Cells International. https://doi.org/10.1155/2019/5620286
Vincenti MP, Coon CI, Brinckerhoff CE (1998) Nuclear factor κB/p50 activates an element in the distal matrix metalloproteinase 1 promoter in interleukin-1β-stimulated synovial fibroblasts. Arthritis Rheum 41:1987–1994
Vinhas A, Rodrigues MT, Gonçalves AI, Reis RL, Gomes ME (2020) Magnetic responsive materials modulate the inflammatory profile of IL-1β conditioned tendon cells. Acta Biomater 117:235–245
Wang Y, He G, Tang H, Shi Y, Kang X, Lyu J, Zhu M, Zhou M, Yang M, Mu M, Chen W, Zhou B, Zhang J, Tang K (2019) Aspirin inhibits inflammation and scar formation in the injury tendon healing through regulating JNK/STAT-3 signalling pathway. Cell Prolif. https://doi.org/10.1111/cpr.12650
Williams RB, Harkins LS, Hammond CJ, Wood JLN (2001) Racehorse injuries, clinical problems and fatalities recorded on British racecourses from flat racing and National Hunt racing during 1996, 1997 and 1998. Equine Vet J 33:478–486
Xie F, Xiao P, Chen D, Xu L, Zhang B (2012) miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol Biol 80:75–84
Yang S, Yu M, Sun L, Xiao W, Yang X, Sun L, Zhang C, Ma Y, Yang H, Liu Y, Lu D, Teitelbaum DH, Yang H (2014) Interferon-γ-induced intestinal epithelial barrier dysfunction by NF-κB/HIF-1α pathway. J Interferon Cytokine Res 34:195–203
Yang TH, Thoreson AR, Gingery A, Larson DR, Passe SM, An KN, Zhao C, Amadio PC (2015) Collagen gel contraction as a measure of fibroblast function in an animal model of subsynovial connective tissue fibrosis. J Orthop Res 33:668–674
Zhang Y, Ge XH, Guo XJ, Guan S, Bin Li XM, Gu W, Xu WG (2017) Bone marrow mesenchymal stem cells inhibit the function of dendritic cells by secreting galectin-1. Biomed Res Int. https://doi.org/10.1155/2017/3248605
Acknowledgements
The authors are especially grateful to Heather Harris for processing the flow cytometry samples at the Royal Veterinary College (RVC).
Funding
The Horserace Betting Levy Board (VET/RS/257 and VET/PRJ/782) and Petplan Charitable Trust (S20-921–960 and S19-747–786) provided funding for this work.
Author information
Authors and Affiliations
Contributions
DJG, JD: conceptualisation: EJS, REB, AM: methodology; EJS, REB, AM, CS, LH: validation; EJS, REB, AM, CS, LH, DJG: formal analysis; EJS, REB, AM, CS, EPL, LH: investigation; DJG: resources; EJS, DJG: writing — original draft preparation; EJS, DJG, JD, RKS: writing — review and editing; DJG, JD: supervision; DJG: project administration; DJG, JD, RKS, EJS: funding acquisition.
Corresponding authors
Ethics declarations
Ethical approval
The equine cells used in this study were isolated with approval from the Royal Veterinary College Clinical Research Ethical Review Board (URN 2020 2017–2) and the Animal Health Trust Ethical Review Committee (AHT_02_2012).
Informed consent
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Smith, E.J., Beaumont, R.E., McClellan, A. et al. Tumour necrosis factor alpha, interleukin 1 beta and interferon gamma have detrimental effects on equine tenocytes that cannot be rescued by IL-1RA or mesenchymal stromal cell–derived factors. Cell Tissue Res 391, 523–544 (2023). https://doi.org/10.1007/s00441-022-03726-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00441-022-03726-6