
Abstract
Main conclusion
Chitinase family genes were involved in the response of Brassica oleracea to Fusarium wilt, powdery mildew, black spot and downy mildew.
Abstract
Abstract Chitinase, a category of pathogenesis-related proteins, is believed to play an important role in defending against external stress in plants. However, a comprehensive analysis of the chitin-binding gene family has not been reported to date in cabbage (Brassica oleracea L.), especially regarding the roles that chitinases play in response to various diseases. In this study, a total of 20 chitinase genes were identified using a genome-wide search method. Phylogenetic analysis was employed to classify these genes into two groups. The genes were distributed unevenly across six chromosomes in cabbage, and all of them contained few introns (≤ 2). The results of collinear analysis showed that the cabbage genome contained 1–5 copies of each chitinase gene (excluding Bol035470) identified in Arabidopsis. The heatmap of the chitinase gene family showed that these genes were expressed in various tissues and organs. Two genes (Bol023322 and Bol041024) were relatively highly expressed in all of the investigated tissues under normal conditions, exhibiting the expression characteristics of housekeeping genes. In addition, under four different stresses, namely, Fusarium wilt, powdery mildew, black spot and downy mildew, we detected 9, 5, 8 and 8 genes with different expression levels in different treatments, respectively. Our results may help to elucidate the roles played by chitinases in the responses of host plants to various diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cabbage (Brassica oleracea var. capitata L.) is one of the most important leafy vegetables cultivated worldwide and is of strong economic value. However, cabbage is vulnerable to various diseases, such as Fusarium wilt (FW), powdery mildew (PM), black spot (BS), downy mildew (DM) and clubroot, during its natural growth process, which leads to large economic losses. The pathogens that cause FW, PM, BS, DM and clubroot in cabbage are Fusarium oxysporum f. sp. Conglutinans (Liu et al. 2017), Erysiphe cruciferarum (Zhao et al. 2014), Alternaria brassicicola (Javeria et al. 2018), Peronospora parasitica (Verma et al. 2018) and Plasmodiophora brassicae (Ji et al. 2020), respectively. In general, except for P. brassicae, which is a special pathogen between fungi and slime molds (Hawksworth et al. 1996), the other four pathogens are all fungi. Chitin, a polymer of N-acetyl-β-d-glucosamine, is widely observed in insect carapaces and the cell walls of fungi and P. brassicae (Merzendorfer 2011; Thornton et al. 1991). The cell wall determines the shape and strength of the pathogen cells and is a key determinant of cell morphology development. As one of the primary components of the cell wall, chitin plays a very important role in the growth and development of pathogens, as well as the defense against external stress (Kombrink et al. 2011). Chitin is also considered to be a prominent signal to induce the natural immunity of plants for the invasion of pathogens (Pentecost 2013). A previous study suggested that the perception of chitin contributes to enhanced disease resistance in certain crops, such as rice and Arabidopsis (Kishimoto et al. 2010; Wan et al. 2012).
Chitinase is an enzyme system that employs chitin as a substrate and hydrolyzes it to N-acetyl oligosaccharide and glucose. As a subgroup of pathogenesis-related (PR) proteins, chitinase is widely present in various organs of higher plants and can be rapidly produced and accumulated when plants are subjected to pathogen infection or abiotic stress sources, such as heavy metals and drought (Bravo et al. 2003; Wang et al. 2015; Li et al. 2018). Therefore, chitinase plays an important role in protecting plants from a variety of pathogens. The induction of pathogens can enhance the activity of chitinase in plants, which subsequently inhibits spore germination and mycelial growth and even directly degrades the chitin of the fungal cell wall (Roby et al. 1988). Ntui et al. (2011) increased tobacco resistance to FW by transferring chitinase genes into tobacco. The same results were also obtained in tomatoes (Jabeen et al. 2015). Similarly, transgenic grapes carrying the wheat chitinase gene exhibit increased resistance to DM (Nookaraju et al. 2012). Marchant et al. (1998) reported that expression of the chitinase transgene reduced the severity of BS development by 13–43% in rose. Chen et al. (2018b) found that the expression of chitinase reduced the symptoms of clubroot in Chinese cabbage. In addition, exogenous application of chitinase to barley may also inhibit the proliferation of PM pathogens.
Although the function of chitinase has been analyzed in various plant species, such as tomato (Staehelin et al. 1994), potato (Khan et al. 2017), rice (Zhao et al. 2018) and apple (Fan et al. 2015), the role of the chitinase gene family in the response of cabbage to FW, PM, BS, DM and clubroot has not been elucidated. In this study, 20 chitinase genes were identified in cabbage, and their chromosome location, gene structure, collinearity relationships, evolution and cis-acting regulatory elements in promoters were further analyzed. The expression patterns of the chitinase family genes in response to FW, PM, BS, DM and clubroot were also investigated. Our results may help to elucidate the role played by chitinase the responses of plants to various diseases and may establish a foundation for future research investigating the genetic improvement of cabbage.
Materials and methods
Genome-wide identification of the chitinase genes
The Brassica oleracea Genomics Database (www.ocri-genomics.org/bolbase/blast/blast.html) was employed to download the cabbage whole-genome protein sequences. The Hidden Markov Model (HMM) profile of the Chiti-bind domain was downloaded from the Pfam (http://www.sanger.ac.uk/Software/Pfam/) database and used for protein screening in HMMER 3.2.1 (e-value < 0.01) (Finn et al. 2011). The first part of the candidate chitinase family proteins was obtained. To obtain the second part of the candidate gene, the chitinase protein sequences of Arabidopsis and Brassica rapa were downloaded from the NCBI (https://www.ncbi.nlm.nih.gov/) as a query and submitted in a BLASTP (P = 0.001) search. Subsequently, the two candidate sets were combined, the redundant proteins were removed, and their conserved domains were further identified using NCBI-CDD search (https://www.ncbi.nlm.nih.gov/cdd). The subcellular locations were predicted using Cell-PLoc 2.0 (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/).
Construction of the phylogenetic tree
Based on the amino acid sequences of chitinase derived from cabbage, Arabidopsis thaliana and B. rapa, we used MEGA6.0 (Tamura et al. 2013) to construct an unrooted neighbor-joining phylogenetic tree (bootstrap = 1000).
Localization analysis of the chitinase genes
We used MapInspect software to draw gene chromosome location diagrams based on information regarding chitinase genes available in the cabbage genome database (http://plants.ensembl.org/Brassica_oleracea/Info/Index).
Collinearity analysis of chitinase genes
The microsyntenic relationships of the chitinase genes in cabbage and Arabidopsis thaliana were detected using BLAST with an e-value cutoff of 1 × 10–5 against the whole genomes of these species. Next, we collected the physical location of the chitinase genes on each chromosome from the respective databases. The Circos tool (Krzywinski et al. 2009) was employed to visualize the relationships between two species.
Gene structure and conserved motif analyses
We used the MEME program (http://meme-suite.org/index.html) and NCBI-CDD (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) to identify the conserved motif and protein sequences, respectively. TBtools (Chen et al. 2018a) was employed to draw the gene exon–intron structure.
Analysis of cis-acting elements in chitinase genes
The cis-acting elements in the promoters of the chitinase genes were identified by submitting the upstream sequences (1.5 kb) of the initiation codon (ATG) of each chitinase gene to PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html).
Plant materials and treatments
The F. oxysporum employed in this study belongs to race 1, which is the primary race worldwide. Inbred lines 01–20 and 96–100 utilized for inoculation are susceptible and resistant to F. oxysporum, respectively. The roots of seedlings with three real leaves were soaked in a 1 × 106 cfu/ml spore suspension for 15 min, and the seedlings were subsequently transferred to 32-well plugs. Two leaves from each plant of 01–20 and 96–100 at 0, 3, 6 and 9 dai (day after inoculation) were collected (18 individuals per treatment, 6 individuals per replicate) for RNA extraction.
The P. brassicae used in this study belongs to race 4 based on the differential sets of Williams (1966). A resting spore inoculum of 2 × 108 spores/ml was prepared prior to inoculation. Two commercial cabbage cultivars, Xiangan 336 and Jinfeng No. 1, which were resistant and susceptible to P. brassicae, respectively, were sown in 32-well (8 × 4) plugs. When the seedlings grew to two real leaves, we employed a pipette to inject 2 ml of resting spore suspension into the soil around the roots of each seedling. Two kinds of treatments were performed for each cultivar. A treatment without inoculation served as the control. Eight different root tissue samples, including Jingfeng No. 1 not inoculated at 7 days, Jingfeng No. 1 inoculated at 7 days, Xiangan 336 not inoculated at 7 days, Xiangan 336 inoculated at 7 days, Jingfeng No. 1 not inoculated at 28 days, Jingfeng No. 1 inoculated at 28 days, Xiangan 336 not inoculated at 28 days and Xiangan 336 inoculated at 28 days, were collected (24 individuals per treatment, 8 individuals per replicate) for RNA extraction.
The cabbage material used for the PM inoculation experiment was the cabbage inbred line D157. When the seedlings grew to 4–5 real leaves, a resting spore suspension of 1 × 105 spores/ml was sprayed evenly onto the leaves of the plants in the treatment group. The plants of the control group were sprayed with equal amounts of sterile water. During the pod-setting period, one diseased leaf of each plant in the treatment group and one healthy leaf of each plant in the control group were taken for RNA extraction. Three replicates were employed in the treatment group and the control group, and each replicate consisted of 8 plants.
The cabbage material used for the BS inoculation experiment was cabbage inbred line W18. When the seedlings grew to 2 real leaves, a resting spore suspension of 1 × 104 pfu/ml was sprayed evenly onto the leaves of the plants in the treatment group. The plants of the control group were sprayed with equal amounts of sterile water. During the heading stage, one diseased leaf from each plant in the treatment group and one healthy leaf from each plant in the control group were taken for RNA extraction. Twenty-four plants were employed in this experiment, and every eight plants were considered to be one biological repetition.
The cabbage material used for the DM inoculation experiment was the cabbage inbred line 01–20. When the seedlings grew to 2 real leaves, a resting spore suspension of 5 × 104 spores/ml was sprayed evenly to the back of the leaves of the plants in the treatment group. The plants of the control group were sprayed with equal amounts of sterile water. During the heading stage, one diseased leaf from each plant in the treatment group and one healthy leaf from each plant in the control group were taken for RNA extraction. Twenty-four plants were employed in this experiment, and every eight plants were considered to be one biological repetition.
All of the samples were quickly frozen in liquid nitrogen and stored at − 80 °C until RNA extraction.
The inbred line 01–20 was introduced to China from Canada in 1966 by the Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences (IVF-CAAS). Also, 96–100, D157 and W18 are backbone inbred lines cultivated by the cabbage-broccoli research group of IVF-CAAS for many years. Jinfeng No. 1 was developed by China Vegetable Seed Co., Ltd., and Xiangan 336 was developed by Syngenta Seeds. The resistance of 01–20 and 96–100 to FW has been observed by previous researchers (Lv et al. 2014). Similarly, the resistance of Xiangan 336 and Jinfeng No. 1 to clubroot has been reported previously (Ning et al. 2018). The resistance of D157, W18 and 01–20 to PM, BS and DM, respectively, has been identified by researchers from the cabbage-broccoli research group of IVF-CAAS. The voucher specimens of all the above materials have been deposited in a public herbarium in IVF-CAAS.
Total RNA extraction
Total RNA was extracted from cabbage samples using TRIzol following the supplier’s instructions (Transgen, Beijing, China). Then, the RNA quality was assessed using a Nanodrop spectrophotometer (Thermo Fisher Scientific, USA) and 1% formaldehyde gel electrophoresis. The cDNA was reverse transcribed with the HiScript® III 1st Strand cDNA Synthesis Kit (Vazyme, Nanjing, China).
The specific primers for chitinase genes were designed with Premier 3.0 (Table S4). qRT-PCR was carried out using 2X RealStar Green Fast Mixture (GeneStar) in a Bio-Rad CFX96 Real Time PCR System. Each amplification reaction was conducted in a 20-μl reaction volume containing 10 μl KAPA SYBR, 0.5 μl of each primer, 2 μl diluted cDNA and 7 μl ddH2O. The PCR program was set as follows: 95 °C for 2 min followed by 40 cycles of 95 °C for 15 s, 60 °C for 30 s and 72 °C for 30 s. Melting curve analysis was performed from 65 °C to 95 °C with increments of 0.5 °C every 5 s. Three independent biological and technical replicates were performed for each reaction. The housekeeping gene actin was employed as the internal reference gene.
Subcellular localization
The pBWA(V)HS-GLosgfp vector was used for the subcellular localization test and digested with one restriction endonucleases (BsaI) to insert the target genes. The CDS sequences of Bol040748 were amplified with specific primer pairs with homologous arms (F: 5′-cgatGGTCTCacaacatgttcatccacaaggacaatactgcttgtccagcaaatggttt-3′; R: 5′-cagtGGTCTCatacaagcgaagggcctctgattttcacagtccaaattaggcccagttc-3′). The amplification products were recovered using the FastPure Gel DNA Extraction Mini Kit (Vazyme Biotech) and inserted into the pBWA(V)HS-GLosgfp vector, resulting in an N-terminal fusion with GFP under the control of the constitutive CaMV35S promoter. The recombinant plasmids were transferred into Agrobacterium tumefaciens strain GV3101. The fusion constructs were introduced into Nicotiana benthamiana protoplasts as previously described (Liu et al. 2020). The fluorescence signals were detected using confocal laser-scanning microscopy C2-ER (Nikon, Tokyo, Japan).
Genes information and data analysis
The GenBank accession numbers of genes in Arabidopsis were listed in Table S1 and the detailed information about the genes in B. oleracea and B. rapa in this study can be queried through the BRADV3.0 (http://39.100.233.196/#/GeneSequence/).
All samples used for RNA sequencing and qRT-PCR in this study were set with three biological replicates. To test the repeatability among samples, all RNA sequencing data were performed principal component analysis (PCA) by RNA-Seq by Expectation Maximization (RSEM) (http://deweylab.biostat.wisc.edu/rsem/), which used transcripts per million (TPM) as the expression index. The error values of qRT-PCR data were calculated with SPSS Statistics 20.0 (SPSS, Chicago, IL, USA).
Results
Genome-wide identification and phylogenetic analysis of chitinase genes in cabbage
To identify chitin-binding genes in the cabbage genome (02–12), a hidden Markov model was employed to predict chitin recognition proteins in cabbage protein sequences. A total of 20 chitinase genes were identified (Table 1). The lengths of these 20 chitinase proteins ranged from 117 (Bol030012) to 447 (Bol007321) amino acids (aa). Within these 20 chitinase proteins, 13 members shared a similar localization to vacuoles, 1 to extracellular vesicles and 6 to more than one compartment.
Chitinases are classified into seven classes (Class I–VII), and most of them belong to the first four classes (Neuhaus et al. 1996). In this study, based on the amino acid sequences of the cabbage (20), Arabidopsis (8), and Brassica. rapa (17) chitinase proteins, we constructed the chitinase protein phylogenetic tree of the chitinase family genes using MEGA 6.0 software. The chitinase genes were classified into two groups (class I and IV; Fig. 1), which contained 14 members (ten cabbage, one Arabidopsis and three Brassica. rapa), and 31 members (ten cabbage, seven Arabidopsis and fourteen Brassica. rapa), respectively. All chitinases belong to the glycoside hydrolase 19 (GH-19) family, and they all have an N-terminal chitin-binding domain and a GH-19 catalytic domain (Henrissat 1991). Coincidentally, Davis et al. (2002) reported that class I and class IV chitinase genes in pineapple were induced after inoculation of Fusarium subglutinans f. sp. pini, which may indicate the special role played by these genes in plant resistance to pathogens.

Phylogenetic tree of chitinase genes from cabbage, A. thaliana and B. rapa. The phylogenetic tree was built using the neighbor-joining (NJ) method with 1000 bootstrap replications. Roman numerals (I and IV) represent each gene cluster, which are labeled with different colors
Chromosomal distribution and collinear analysis
The 20 chitinase genes were assigned to six chromosomes of cabbage (Fig. 2). The distribution of the chitinase genes on each chromosome was uneven. The numbers of chitinase genes on each chromosome are as follows: 2 on C01, 8 on C03, 4 on C04, 3 on C05, 2 on C08, and 1 on C09.

Distribution of chitinase genes in B. oleracea. chromosomes. The number on the top of each chromosome represents the cabbage chromosome number. Gene names are indicated on the right sides of each chromosome. The distance (Mb) between genes or genes to the ends of the chromosome is indicated on the left side of each chromosome
Gene duplication is a common phenomenon in the evolution of plants, which is the reason for the formation of homologous genes in different plants. Due to the importance of gene duplications on the evolution of gene families in plants, chitinase gene replication in cabbage and the collinearity between cabbage and Arabidopsis of chitinase genes were analyzed.
The cabbage genome contained 1–5 copies of each chitinase gene (excluding Bol035470) found in Arabidopsis (Fig. 3; Table S2). For example, AT1G56680.1 contained only one homologous gene (Bol029467) in cabbage, while AT2G43590.1 contained up to five homologous genes (Bol004604, Bol039802, Bol021626, Bol030012, and Bol025197) in cabbage. In addition, 32 segmentally duplicated gene pairs were also identified among the 20 chitinase genes in the cabbage genome (Fig. 3; Table S3).

Syntenic relationship of cabbage and A. thaliana chitinase genes shown on the chromosome maps. C01-C09, nine chromosomes of cabbage. 1–5, five chromosomes of A. thaliana. Orange lines, homologous gene between cabbage chromosomes. Green lines, homologous genes between cabbage and A. thaliana chromosomes
Structure and conserved motif analysis of chitinase genes
To further investigate the structural diversity of chitinase genes, the gene structure among 20 chitinase genes was detected (Fig. 4b). Thirteen genes contain two exons, 4 genes contain 3 exons, and 3 genes contain only 1 exon. The lengths of exons in most genes were similar, while the lengths of introns for some genes varied widely. For example, Bol021627 and Bol029469 contained one shorter intron, whereas Bol029467 contained two notably long introns.

Conserved motif (a) and gene structure (b) analysis of chitinase genes. The motifs are indicated in different colored boxes. Exons are represented by boxes, while introns are represented by gray lines
To better understand the structural characteristics of the chitinase genes, the conserved domain and motifs were also detected (Fig. 4a, b). All members contained motifs 2 and 6. Motifs 3, 4 and 9 were uniquely present in members of Class I, while Motifs 7 and 8 were almost present in members of Class IV. Only one member of Class I contains motif 7. Among the 20 chitinase proteins, sixteen members contained motif 1, 15 members contained motif 5 and 14 members contained motif 10. Motifs 2, 3, 6, 1, 4, and 9 displayed in the same order were found in Class I, and motifs 5, 10, 2, 7, 6, 1, and 8 displayed in the same order were found in Class IV. In addition, Bol011420, Bol040748, Bol041024, Bol023322 and Bol035470 have the same motif composition.
As shown in Fig. 4b, most proteins contained a chitinase binding domain and GH19. Only one protein contained a K+ transit domain. The chitinase-like domain was present in 14 genes. The cystatin domain only existed on 1 gene (Bol007321). In addition, most genes in Class I contained lysozyme-like domains, which only existed on the 1 (Bol029467) gene in Class IV.
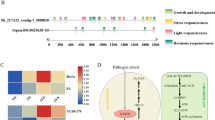
The cis-elements in the promoters of B. oleracea chitinase genes
To further clarify the regulatory mechanism of chitinase genes in cabbage response to FW, clubroot, BS, PM and DM, we identified the cis-elements using the PlantCARE database based on the promoter sequences, and ten types of cis-acting regulatory elements were identified (Fig. 5). All 20 chitinase genes contained 3–17 light-responsive cis-elements. Seven chitinase genes contained gibberellin-responsive cis-elements. Eight chitinase genes contained MYBHv1-binding cis-elements. Salicylic acid-responsive cis-elements were detected in 10 chitinase genes, while cis-elements related to MeJA and auxin responsiveness were detected in 15 and 11 chitinase genes, respectively. In addition, the cis-elements related to defense and stress responsiveness, as well as low-temperature responsiveness, existed in 8 and 9 genes, respectively. The distribution of drought-inducible cis-elements was relatively small and was detected in only 3 genes. The cis-element analysis demonstrated that chitinase genes could respond to different stimuli.

Predicted cis-acting elements in chitinase gene promoters
Expression patterns of chitinase genes and qRT-PCR verification
The RNA-Seq dataset (GSM1052958-964) was examined to determine the expression levels of chitinase genes in the leaves, stem, flowers, siliques, buds, calli and roots of cabbage. Most of the chitinase genes exhibited different expression patterns (Fig. 6; Table S5). Eighteen of the genes were expressed in all organs, while the expression levels of two chitinase genes (Bol030015 and Bol007323) were almost undetectable. Some genes were expressed only in one or two organ types, such as Bol030012 in leaves and Bol029469 in siliques and calluses. Conversely, Bol023322 and Bol041024 were highly expressed in all tissues, showing the expression characteristics of housekeeping genes (Lopes-Caitar et al. 2013). The multiple expression patterns of the chitinase genes indicate their extensive biological functions during the growth and development of cabbage.

Expression of chitinase genes in different organs
To investigate the role played by chitinase in the response of cabbage to infection by various pathogens, we inoculated different cabbage materials with five pathogens and extracted plant tissue RNA at a specific period for transcriptome sequencing. Then, two heatmaps were established according to the RNA-seq data (Figs. 6, 7; Table S6).

Expression patterns of chitinase genes analyzed by RNA-Seq. Samples S0, S3, S6 and S9 represented 01–20 inoculated by F. oxysporum at 0, 3, 6 and 9 days, respectively. Samples R0, R3, R6 and R9 represented 96–100 inoculated by F. oxysporum at 0, 3, 6 and 9 days, respectively. J7C, Jingfeng No. 1 not inoculated by P. brassicae at 7 days. J7I, Jingfeng No. 1 inoculated by P. brassicae at 7 days. X7C, Xiangan 336 not inoculated by P. brassicae at 7 days. X7I, Xiangan 336 inoculated by P. brassicae at 7 days. J28C, Jingfeng No. 1 not inoculated by P. brassicae at 28 days. J28I, Jingfeng No. 1 inoculated by P. brassicae at 28 days. X28C, Xiangan 336 not inoculated by P. brassicae at 28 days. X28I, Xiangan 336 inoculated by P. brassicae at 28 days. QBF-S, leaves with PM of D157 during the pod-setting stage. QBF-R, normal leaves of D157 during the pod-setting stage. BP, leaves with BS of W18 during the heading stage. 120-QY, normal leaves of W18 during heading storage. SMB, leaves with DM of 01–20 during heading storage. 120-WY, normal leaves of 01–20 during heading storage
The expression levels of chitinase genes in 01–20 and 96–100 infected by F. oxysporum were notably different. Four genes (Bol010293, Bol007321, Bol021626 and Bol025197) were upregulated in 96–100 compared with 01–20, while five genes (Bol035464, Bol021627, Bol035467, Bol029469 and Bol040748) were downregulated. In different stages, the expression patterns of chitinase genes are also different. Five genes (Bol035464, Bol010293, Bol021627, Bol007321 and Bol035467) and two genes (Bol023322 and Bol041024) were significantly up- and downregulated, respectively, in both 01–20 and 96–100 after inoculation by F. oxysporum. The expression levels of the three genes (Bol029469, Bol030015 and Bol004604) increased first and then decreased in both 01–20 and 96–100 after inoculation. In contrast, the expression levels of the two genes (Bol011420 and Bol040748) decreased first and then increased in 96–100 after inoculation. Taken together, compared with 01–20, five genes (Bol035464, Bol010293, Bol021627, Bol035467 and Bol040748) were significantly downregulated, and five genes (Bol007321, Bol041024, Bol0304604, Bol021626 and Bol025197) genes were significantly upregulated, in 96–100 after inoculation. qRT-PCR was performed to verify the chitinase gene expression patterns under F. oxysporum stress in different inoculation periods of 01–20 and 96–100. As shown in Fig. 8, the seven genes that we detected by qRT-PCR were approximately in keeping with the results of the RNA-seq analysis, except for Bol004604, which further confirmed their expression patterns.

Expression levels of chitinase genes under F. oxysporum stress. Data are presented as the means ± SD
The expression patterns exhibited by the chitinase genes in response to invasion of E. cruciferarum, A. brassicicola and P. parasitica are also different in diseased and normal leaves. Compared with PM leaves, three genes (Bol023322, Bol041024 and Bol035470) and two genes (Bol040748 and Bol021626) were up- and downregulated, respectively, in normal leaves. Similarly, three genes (Bol007321, Bol023322 and Bol041024) and five genes (Bol035470, Bol011420, Bol040748, Bol021626 and Bol025197) were up- and downregulated, respectively, in normal leaves compared to BS leaves. In addition, compared to the leaves with DM, two genes (Bol041024 and Bol035470) and six genes (Bol010293, Bol007321, Bol023322, Bol011420, Bol040748 and Bol021626) were up- and downregulated in normal leaves, respectively.
Under P. brassicae infection stress, there was little change in the expression levels of all chitinase genes in the 8 different treatments, which suggesting that chitinase genes played only minor roles in resistance to clubroot. The qRT-PCR results for six chitinase genes were also largely consistent with the results of the RNA-seq analysis (Fig. 9).

Expression levels of chitinase genes under P. brassicae stress. Data are presented as the means ± SD
Subcellular localization of Bol040748
Protein function was always associated with its subcellular localization. To further understand the protein characteristic of chitinase genes, we recombined the pBWA(V)HS-GLosgfp vector with the CDS sequences of Bol040748 and then introduced into tobacco protoplasts. As shown in Fig. 10, The GFP signal of Bol040748 was observed in the cell membrane and nucleus, suggesting that Bol040748 was a cellular membrane and nuclear protein.

Subcellular localization of Bol040748 in tobacco protoplast. a–c and d: Target protein GFP channel, Chloroplast channel, Bright, and Merge. e–h Control GFP channel, chloroplast channel, bright, and merge. Bars = 20 µm
Discussion
Induced resistance means that the stimulus of the pathogen increases the defense of the plant (Van Loon. 1997). Chitinases, representing a subgroup of pathogenesis-related proteins, play important roles in the plant defense against pathogen invasion (Abeles et al. 1971). Chitinases have been identified in previous studies, and their roles in defending against various pathogens in different crops have been discussed (Kasprzewska. 2003; Rasmussen et al. 1992; Zhou et al. 2017). However, the expression patterns of chitinase genes in response to F. oxysporum, P. brassicae, A. brassicicola, E. cruciferarum and P. parasitica in cabbage have not been elucidated to date. In this study, 20 chitinase genes were identified, and their phylogenetic relationship, collinearity, structures, chromosomal locations, cis-elements and expression patterns in response to the invasion of different pathogens in cabbage were reported. This study provides comprehensive information to characterize the chitinase gene family in cabbage.
Whole genome duplication (WGD) is an important event in plant evolution (Adams et al. 2005). Li et al. (2016) studied the duplicated genes of almost 40 different flowering plants that experienced WGD and found that most of the genes quickly returned to a single-copy status; however, some genes were observed to be consistently present in multiple copies, which belong to gene families that are involved in conditional responses to biotic and abiotic stress and are important for local adaptation (Van de Peer et al. 2017). Arabidopsis thaliana has undergone three paleopolyploidy events and two more recent tetraploidy events a and b shared with other members of the order Brassicales (Bowers et al. 2003). In this study, we found that the number of chitinase genes in B. rapa (17) and B. oleracea (20) nearly doubled or tripled compared with the number in A. thaliana (8) (Figs. 1, 3), which suggests that the chitinase genes may be subjected to natural selection pressure and were biased to retain multiple copies after the triploidization event (Cheng et al. 2012).
In this study, we analyzed the expression patterns of 20 chitinase genes under 5 different disease stresses. Specifically, we investigated the chitinase gene members that might play crucial roles in disease resistance. Under FW stress, we detected four genes (Bol010293, Bol007321, Bol021626 and Bol025197) and five genes (Bol035464, Bol021627, Bol035467, Bol029469 and Bol040748) that were up- and downregulated significantly in 96–100 compared with 01–20. The results of qRT-PCR analysis showed that Bol004604 and Bol040748 had completely opposite expression patterns in 96–100 and 01–20, which may indicate that these two genes play an important role in the defense against FW in cabbage. For PM, BS and DM, there were 5, 8 and 8 genes with differential expression in different treatments, respectively. Among these genes, all Bol023322, Bol041024, Bol035470, Bol040748 and Bol021626 had different expression levels between diseased and normal leaves under the stress of three diseases, which suggests their pivotal effects on the interaction between cabbage and fungi. In addition, Bol007321 and Bol011420 also warranted further study, although they were only differentially expressed between diseased and normal leaves under the stress of BS and DM.
Under the stress of P. brassicae, almost all the chitinase genes did not exhibit clear differences in expression, which may suggest that the defense effect of chitinase against clubroot disease was not significant in this study. Coincidentally, from the taxonomic perspective, P. brassicae is different from the other four fungi, which may be the reason for the different expression patterns exhibited by the chitinase genes under the pressure of infection by P. brassicae and fungi.
Conclusions
In this study, a genome-wide analysis of B. oleracea chitinase genes was performed, and 20 chitinase genes were confirmed. Subsequently, analyses of chitinase genes on gene structures, phylogeny, chromosomal location, gene duplication and gene expression patterns were conducted based on bioinformatic analysis and qRT-PCR. These genes were expressed in various tissues and organs. In addition, there were 9, 5, 8 and 8 genes with differential expression levels in different treatments under the four respective stresses of FW, PM, BS and DM. This study provides comprehensive information for further research investigating the role of chitinase in host plant responses to various diseases.
Author contribution statement
SJL and MZ conceived and designed research. YYZ and HHL conducted experiments. JLJ and XLH collected the public dataset and performed bioinformatics analysis. MZZ wrote the manuscript. ZYF, YW and LMY reviewed the manuscript. All authors read and approved the manuscript.
References
Abeles FB, Bosshart RP, Forrence LE, Habig WH (1971) Preparation and purification of glucanase and chitinase from bean leaves. Plant Physiol 47:129–134
Adams KL, Wendel JF (2005) Polyploidy and genome evolution in plants. Curr Opin Plant Biol 8:135–141
Bowers JE, Chapman BA, Rong J, Paterson AH (2003) Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 422:433–438
Bravo JM, Campo S, Murillo I, Coca M, Segundo BS (2003) Fungus-and wound-induced accumulation of mRNA containing a class II chitinase of the pathogenesis-related protein 4 (PR-4) family of maize. Plant Mol Biol 52:745–759
Chen J, Piao Y, Liu Y, Li X, Piao Z (2018) Genome-wide identification and expression analysis of chitinase gene family in Brassica rapa reveals its role in clubroot resistance. Plant Sci 270:257–267
Chen C, Chen H, He Y, Xia R (2018a) TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface. BioRxiv 289660.
Cheng F, Wu J, Fang L, Sun S, Liu B, Lin K, Bonnema G, Wang X (2012) Biased gene fractionation and dominant gene expression among the subgenomes of Brassica rapa. PLoS ONE 7:e36442
Davis JM, Wu H, Cooke JE, Reed JM, Luce KS, Michler CH (2002) Pathogen challenge, salicylic acid, and jasmonic acid regulate expression of chitinase gene homologs in pine. Mol Plant Microbe Interact 15:380–387
Fan XJ, Mi YX, Ren H, Zhang C, Li Y, Xian XX (2015) Cloning and functional expression of a chitinase cDNA from the apple leaf miner moth Lithocolletis ringoniella. Biochemistry 80:242–250
Finn RD, Clements J, Eddy SR (2011) HMMER web server: interactive sequence similarity searching. Nucleic Acids Res 39:W29–W37
Hawksworth DL, Kirk PM, Sutton BC, Pegler DN (1996) Ainsworth & Bisby’s dictionary of the fungi. Rev Inst Med Trop Sao Paulo 38:272–272
Henrissat B (1991) A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem J 280:309–316
Jabeen N, Chaudhary Z, Gulfraz M, Rashid H, Mirza B (2015) Expression of rice chitinase gene in genetically engineered tomato confers enhanced resistance to Fusarium wilt and early blight. Plant Pathol J 31:252
Javeria S, Deep S, Prasad L, Sharma P (2018) Vegetative compatibility grouping of Alternaria brassicicola causing black leaf spot in cauliflower. Biochem J 71:43–47
Ji R, Gao S, Bi Q, Wang Y, Lv M, Ge W, Feng H (2020) The salicylic acid signaling pathway plays an important role in the resistant process of Brassica rapa L. ssp. pekinensis to Plasmodiophora brassicae woronin. J Plant Growth Regul 1–18
Kasprzewska A (2003) Plant chitinases-regulation and function. Cell Mol Biol Lett 8:809–824
Khan A, Nasir IA, Tabassum B, Aaliya K, Tariq M, Rao AQ (2017) Expression studies of chitinase gene in transgenic potato against Alternaria solani. Plant Cell Tissue Organ Cult 128:563–576
Kishimoto K, Kouzai Y, Kaku H, Shibuya N, Minami E, Nishizawa Y (2010) Perception of the chitin oligosaccharides contributes to disease resistance to blast fungus Magnaporthe oryzae in rice. Plant J 64:343–354
Kombrink A, Sánchez-Vallet A, Thomma BPHJ (2011) The role of chitin detection in plant–pathogen interactions. Microbes Infect 13:1168–1176
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA (2009) Circos: an information aesthetic for comparative genomics. Genome Res 19:1639–1645
Li Z, Defoort J, Tasdighian S, Maere S, Van de Peer Y, De Smet R (2016) Gene duplicability of core genes is highly consistent across all angiosperms. Plant Cell 28:326–344
Li P, Zhang Y, Wu X, Liu Y (2018) Drought stress impact on leaf proteome variations of faba bean (vicia faba L.) in the qinghai-tibet plateau of China. Biotech 8:110.
Liu X, Ling J, Xiao Z, Xie B, Fang Z, Yang L, Zhang Y, Lv H, Yang Y (2017) Characterization of emerging populations of Fusarium oxysporum f. sp. conglutinans causing cabbage wilt in China. J Phytopathol 165:813–821
Liu X, Zhao C, Yang L, Zhang Y, Wang Y, Fang Z, Lv H (2020) Genome-wide identification, expression profile of the TIFY gene family in Brassica oleracea var. capitata, and their divergent response to various pathogen infections and phytohormone treatments. Genes 11:127
Van Loon LC (1997) Induced resistance in plants and the role of pathogenesis-related proteins. Eur J Plant Pathol 103:753–765
Lopes-Caitar VS, de Carvalho MC, Darben LM, Kuwahara MK, Nepomuceno AL, Dias WP, Abdelnoor RV, Marcelino-Guimarães FC (2013) Genome-wide analysis of the Hsp 20 gene family in soybean: comprehensive sequence, genomic organization and expression profile analysis under abiotic and biotic stresses. BMC Genomics 14:577
Lv H, Wang Q, Yang L, Fang Z, Liu Y, Zhuang M, Zhang Y, Yang Y, Xie B, Wang X (2014) Breeding of cabbage (Brassica oleracea L. var. capitata) with fusarium wilt resistance based on microspore culture and marker-assisted selection. Euphytica 200:465–473
Marchant R, Davey MR, Lucas JA, Lamb CJ, Dixon RA, Power JB (1998) Expression of a chitinase transgene in rose (Rosa hybrida L.) reduces development of blackspot disease (Diplocarpon rosae Wolf). Mol Breed 4:187–194
Merzendorfer H (2011) The cellular basis of chitin synthesis in fungi and insects: common principles and differences. Eur J Cell Biol 90:759–769
Neuhaus JM, Fritig B, Linthorst HJM, Meins FJRFM, Mikkelsen JD, Ryals J (1996) A revised nomenclature for chitinase genes. Plant Mol Biol Rep 14:102–104
Ning Y, Wang Y, Fang Z, Zhuang M, Zhang Y, Lv H, Liu Y, Li Z, Yang L (2018) Identification and characterization of resistance for Plasmodiophora brassicae race 4 in cabbage (Brassica oleracea var. capitata). Austral Plant Pathol 47:531–541
Nookaraju A, Agrawal DC (2012) Enhanced tolerance of transgenic grapevines expressing chitinase and β-1,3-glucanase genes to downy mildew. Plant Cell Tissue Organ Cult 111:15–28
Ntui VO, Azadi P, Thirukkumaran G, Khan RS, Chin DP, Nakamura I, Mii M (2011) Increased resistance to fusarium wilt in transgenic tobacco lines co-expressing chitinase and wasabi defensin genes. Plant Pathol 60:221–231
Van de Peer Y, Mizrachi E, Marchal K (2017) The evolutionary significance of polyploidy. Nat Rev Genet 18:411
Pentecost JWG (2013) Potential application of chitin signaling in engineering broad-spectrum disease resistance to fungal and bacterial pathogens in plants. Adv Crop Sci Tech 1:e103
Rasmussen U, Bojsen K, Collinge DB (1992) Cloning and characterization of a pathogen-induced chitinase in Brassica napus. Plant Mol Biol 20:277–287
Roby D, Toppan A, Esquerré-Tugayé MT (1988) Systemic induction of chitinase activity and resistance in melon plants upon fungal infection or elicitor treatment. Physiol Mol Plant Pathol 33:409–417
Staehelin C, Granado J, Müller J, Wiemken A, Mellor RB, Felix G, Regenass M, Broughton WJ, Boller T (1994) Perception of Rhizobium nodulation factors by tomato cells and inactivation by root chitinases. Proc Natl Acad Sci USA 91:2196–2200
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) Molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30:2725–2729
Thornton CR, Jarvis BC, Cooke RC (1991) A chitin assay for the enumeration of Plasmodiophora brassicae resting spores in clubroot tissue. Mycol Res 95:879–882
Verma A, Singh Y (2018) Inheritance of downy mildew resistance and its relationship with biochemical traits in cauliflower (Brassica oleracea L. var. botrytis). Crop Prot 106:132–138
Wan J, Tanaka K, Zhang XC, Son GH, Brechenmacher L, Stacey NG (2012) Lyk4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis. Plant Physiol 160:396–406
Wang LY, Wang YS, Zhang JP, Gu JD (2015) Molecular cloning of class III chitinase gene fromavicennia marinaand its expression analysis in response to cadmium and lead stress. Ecotoxicology 24:1697–1704
Williams PH (1966) A system for the determination of races of Plasmodiophora brassicae that infect cabbage and rutabaga. Phytopathology 56:624–626
Zhao X, Situ G, He K, Xiao H, Su C, Li F (2018) Functional analysis of eight chitinase genes in rice stem borer and their potential application in pest control. Insect Mol Biol 27:835–846
Zhao HH, Xing HH, Liang C, Yang XY, Cho SE, Shin HD (2014) First report of powdery mildew caused by Erysiphe cruciferarum on Chinese cabbage in China. Plant Dis 98:421–421
Zhou K, Zhou F, Huang J, Yang Q, Jiang S, Qiu L, Yang L, Zhu C, Jiang S (2017) Characterization and expression analysis of a chitinase gene (PmChi-4) from black tiger shrimp (Penaeus monodon) under pathogen infection and ambient ammonia nitrogen stress. Fish Shellfish Immunol 62:31–40
Acknowledgements
The work was supported by the Youth Science Fund Project (Grant No. 31801876) and the National Key Research and Development Program of China (Grant No. 2017YFD0101804).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Communicated by Anastasios Melis.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
425_2021_3596_MOESM2_ESM.xlsx
Supplementary file 2 Table S2. Chitinase gene homologs in the genomes of Arabidopsis thaliana and B. oleracea. (XLSX 13 KB)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhu, M., Lu, S., Zhuang, M. et al. Genome-wide identification and expression analysis of the Brassica oleracea L. chitin-binding genes and response to pathogens infections. Planta 253, 80 (2021). https://doi.org/10.1007/s00425-021-03596-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00425-021-03596-2