
Abstract
In the Brassica genus, we find both diploid species (one genome) and allotetraploid species (two different genomes) but no naturally occurring hexaploid species (three different genomes, AABBCC). Although hexaploids can be produced via human intervention, these neo-polyploids have quite unstable genomes and usually suffer from severe genome reshuffling. Whether these genome rearrangements continue in later generations and whether genomic arrangements follow similar, reproducible patterns between different lineages is still unknown. We crossed Brassica hexaploids resulting from different species combinations to produce five F1 hybrids and analyzed the karyotypes of the parents and the F1 hybrids, as well as allele segregation in a resulting test-cross population via molecular karyotyping using SNP array genotyping. Although some genomic regions were found to be more likely to be duplicated, deleted, or rearranged, a consensus pattern was not shared between genotypes. Brassica hexaploids had a high tolerance for fixed structural rearrangements, but which rearrangements occur and become fixed over many generations does not seem to show either strong reproducibility or to indicate selection for stability. On average, we observed 10 de novo chromosome rearrangements contributed almost equally from both parents to the F1 hybrids. At the same time, the F1 hybrid meiosis produced on average 8.6 new rearrangements. Hence, the increased heterozygosity in the F1 hybrid did not significantly improve genome stability in our hexaploid hybrids and might have had the opposite effect. However, hybridization between lineages was readily achieved and may be exploited for future genetics and breeding purposes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Brassica genus belongs to the Brassicaceae family (Warwick et al. 2006) and includes many economically valuable crop species. The Brassica crops can be broadly classified into vegetable, oilseed, fodder, and condiment types. During the 1930s, the chromosome number and genetic relationships between the cultivated Brassica species were established in what we know as the triangle of U (U 1935): the diploid species B. rapa (AA, n = 10), B. nigra (BB, n = 8) and B. oleracea (CC, n = 9) were defined as progenitors of the allotetraploid species B. juncea (AABB, n = 18), B. napus (AACC, n = 19), and B. carinata (BBCC, n = 17), which originated via pairwise spontaneous hybridization between these diploids. The Brassica vegetables, B. oleracea and B. rapa, are characterized by vast diversity in subspecies and varieties (Cheng et al. 2016). In these species, it is possible to find different domesticated morphotypes distinguishable by leaf types, inflorescence types or enlargement of roots or stems. In Brassica species it is also possible to find traits of agronomic interest, such as disease resistance (Chevre et al. 1996; Mei et al. 2011, 2013; Peng et al. 2014; Taylor et al. 2015; Fredua-Agyeman et al. 2019), abiotic stress tolerance (Gill et al. 2011; Hayat et al. 2011; Wilson et al. 2014; Irfan et al. 2014) and pod shatter resistance (Zhang et al. 2016), among other potential traits (reviewed in Katche et al. (2019)).
Commonly, in the Brassica genus, we find both diploid and tetraploid species but no naturally occurring hexaploid species (AABBCC = 2n = 6x = 54). Despite this, it is possible to synthesize this hybrid via human intervention. The three most common cross combinations to produce an allohexaploid are: (i) B. carinata × B. rapa (Fig. 1a), (ii) B. juncea × B. oleracea, and (iii) B. napus × B. nigra (reviewed in (Gaebelein and Mason 2018)), that from now on will be referred to as “carirapa,” “junleracea,” and “naponigra” allohexaploid types (Zou et al. 2010). All of these hybrids are usually colchicine-treated to induce chromosome doubling following hybridization between a diploid and a tetraploid species. A more recent method to produce an allohexaploid is via two-step crossing and relies on unreduced gamete production: the hybridization between B. napus × B. carinata × B. juncea (Mason et al. 2012), referred to as “NCJ” allohexaploid types (Fig. 1a). Until now, many of the allohexaploid genotypes produced were solely used to cross to B. napus, either to introgress genetic diversity (Li et al. 2004, 2006; Jiang et al. 2007; Zou et al. 2010, 2018; Hu et al. 2019), or to transfer specific traits such as yellow seededness (Meng et al. 1998; Rahman 2001) or fungal disease resistance (Sjödin and Glimelius 1989). Despite this, a new allohexaploid hybrid has the potential to become a new species with the advantage of combining all the different traits present in the six U´s triangle species, thus broadening the genetic resources available for breeders (Chen et al. 2011). In such an allohexaploid, it would also be possible to take advantage of “fixed heterosis” (Abel et al. 2005), where the heterosis is present between the subgenomes (A, B, and C) can be maintained in inbreeding lines. In addition, new phenotypes which are not present in the original parents may develop from the different crosses via novel mutations due to the hybridization event (Udall and Wendel 2006; Kaur et al. 2014).

Crossing scheme. The arrows indicate the direction of the crossing. a Crossings involved in the production of the NCJ and carirapa hexaploid lines. b Selfing and selection of the genotypes. c Crossings between NCJ and carirapa genotypes. For the NCJ genotype combination N6C2.J2, two different lineages were used for crossing, indicated by a different shade of red. The number after the genotype combination represents the plant number. d F1 test-cross to another carirapa hexaploid
Allohexaploid Brassica (AABBCC = 2n = 54), as a new polyploid, has to overcome the major challenge of establishing regular meiosis (Pelé et al. 2018). The correct pairing of homologous chromosomes during meiosis is a key factor to ensure correct cross-over and chromosome segregation. In the case of this allohexaploid, meiosis is challenging due to the presence of three ancestrally homologous chromosome sets from different evolutionary lineages, known as homologous chromosomes (Ramsey and Schemske 2002). If nonhomologous pairing during meiosis occurs, it can lead to different chromosomal rearrangements, such as deletions, duplications and translocations (Udall et al. 2005), heavily affecting genome stability. In a synthetic trigenomic hybrid, we usually observe the A and the C genome are more likely to pair during meiosis, compared to the A-B or C-B homoeologs (Mason et al. 2010). In the case of natural allopolyploids, such a B. napus, meiosis progresses normally, although nonhomologous exchanges do still occur at lower frequency (Parkin et al. 1995; Udall et al. 2005; Higgins et al. 2018).
Many different types of cross-combinations have been tested to produce meiotically stable 2n = AABBCC allohexaploids (reviewed in Gaebelein and Mason (2018)). It has been shown that early generation (F2) carirapa hexaploids have low pollen viability and irregular configurations during meiosis (Howard 1942; Iwasa 1964). It has also been seen that fertility and number of 2n = 54 (putatively euploid) plants can increase by selection over successive generations (Tian et al. 2010; Zhou et al. 2016). Genotype combinations seem to play a great role in the success of the hexaploid, as many of the more stable and fertile hexaploids produced are derived from just a few lines (Tian et al. 2010). A specific B. rapa genotype (R01) was also reported to result in a meiotically stable carirapa allohexaploid (Gupta et al. 2016). Carirapa combinations that included this B. rapa genotype had a high frequency of bivalent formation during meiosis, progeny with the expected 54 chromosomes, and no major translocations observed (Gupta et al. 2016). Similarly, genotype-specific effects on fertility and meiotic stability have been observed in NCJ hexaploids (Mwathi et al. 2017).
In the present study we analyzed chromosome and allele segregation resulting from the meiosis of five different F1 hybrids produced between crosses of putatively stable Brassica hexaploids carirapa and NCJ. We hypothesized that the new hybrids might show improved meiotic stability (more regular chromosome segregation and fewer nonhomologous recombination events) compared to their parents. We also aimed to determine if chromosome rearrangements found at high frequency or in independent lineages of the parent hexaploid types (e.g., both in NCJ and carirapa) might be associated with improved meiotic stability, since all lineages underwent strong selective pressure for fertility.
Materials and methods
Plant material and crossing scheme
The most advanced genotypes available from the two NCJ parental genotype combinations, N1C2.J1 and N6C2.J2 (one and two lineages, respectively) (Mason et al. 2012; Mwathi et al. 2017) and as well as one lineage from each carirapa allohexaploid genotypes C13, C21 (Tian et al. 2010), were selected based on chromosome number and total seeds produced (Fig. 1b, Supplementary Table 1). In total, 12 plants were grown from the selected genotypes, with 6 plants per allohexaploid type: three plants N1C2.J1, two plants N6C2.J2 lineage one, one plant N6C2.J2 lineage two, four plants C13, and two plants C21 (Fig. 1c). Initially, two plants from each of hexaploid genotypes C13 (plants 2 and 3) and N6C2.J2 (lineage one, plants 1 and 2) were grown and crossed under greenhouse conditions at Justus Liebig University, Giessen, Germany, while the rest of the plants were grown under field conditions at Huazhong Agricultural University, Wuhan, China. In the first round of crossings (F1 hybrid production), the NCJ hexaploid plants were used as a female parents, and the carirapa plants were used as the pollen donor to produce eight different populations (Fig. 1c). The cross was done by hand via emasculating flower buds and gently rubbing the anthers over the exposed stigma. The pollinated buds were then labeled and covered with a microperforated bag to prevent contamination from other pollen sources.
The F1 hybrid seeds were collected and grown under field conditions at Huazhong Agricultural University, Wuhan, China. To be able to analyze the meiotic performance and allele segregation from this new F1 hybrid, a test-cross was carried out. Selection of the F1 hybrids was done based on qualitative phenotyping (e.g., high pollen production, relatively normal agronomic phenotype, and plants looked like true hybrids) for individual plants resulting from the crosses between the lines. A total of eight F1 hybrid plants were selected and test-crossed to another carirapa hexaploid (genotypes C21, C28, and C34) (Tian et al. 2010) (Fig. 1d). The test cross-progeny varied in size depending on the population, ranging from 14 to 22 individuals that were grown under field conditions at Huazhong Agricultural University, Wuhan, China.
DNA extraction and SNP analysis
From each plant, a piece of leaf sample was collected, and DNA was extracted using the CTAB method (Doyle and Doyle 1990). For the following plants, the original leaf material was not available, but three different sibling plants were used instead: F1 hybrid population 3 and test-cross parent population 4. In the case of B. carinata and B. rapa parental lines of the carirapa hexaploids, we extracted DNA from two different plants to account for potential heterosis in the material. The DNA was then genotyped using the Illumina Infinium Brassica 90 K SNP array (Illumina, San Diego, CA, USA) following manufacturer’s instructions. A total of 77,970 SNPs distributed across the A (23,482), B (25,822), C (26,731), and unplaced location (1877) Brassica subgenomes were obtained after applying the recommended cluster file for the A and C genomes (Clarke et al. 2016, Supplementary Data set Table 2) and by automated clustering in Genome Studio for the B genome. SNP positions for the A and C genomes were determined by the top hit (highest e-value) based on BLAST to the B. napus Darmor-bzh v. 8.1 reference genome (Bayer et al. 2017). For the B genome, we used the positions provided in the Illumina Infinium 90 K SNP array, which were based on the B. nigra Ni100 short read reference genome assembly (Perumal et al. 2020). The SNP data was initially cleaned by removing non-specific alleles and SNPs with undetermined genomic locations.
The first step in the analysis was to carry out paternity testing to verify the genetic composition of the F1 hybrid between the NCJ and carirapa allohexaploid parents for each of the eight populations. For this, homozygous polymorphic alleles for each parent in the B subgenome were used as diagnostics. If the expected heterozygosity was observed in the F1 plant (e.g. NCJ allele AA × carirapa allele BB → F1 hybrid allele AB), it was considered a true F1 hybrid. The second step was to determine true segregating progeny between the F1 hybrid and the test-cross carirapa allohexaploid parent. To do this, the same approach was used by selecting homozygous polymorphic alleles in the B subgenome between the F1 and test-cross carirapa parent: if the segregating progeny had the expected heterozygous allele, they were considered true test-cross progeny. If the population were true hybrids in both analyzed cases, subsequent analyses were done.
Chromosome count and molecular karyotyping
To determine chromosome presence or absence, we used SNP and Log2 R ratio data (Supplementary Data set Table 3). The absence of both chromosomes was seen across the entire chromosome, or most of it, as “no call” SNPs (NC). If the chromosome was present in the SNP data (at least one copy of the chromosome present), the Log2 R ratio data was used to determine loss or gain of a chromosome copy by assessing the inheritance of the centromeric region of each chromosome (inheritance of the centromere was inferred to mean inheritance of that chromosome, since chromosome fragments cannot be transmitted without a centromere). Experimentally-derived values were used based on comparison to hybrid standards with known haploid and diploid chromosome complements. Log2 R ratios between − 0.5 and − 0.2 were assumed to indicate one missing chromosome copy, Log2 R ratios between 0.2 and 0.5 were assumed to indicate gain of an extra copy and Log2 R ratios of more than 0.5 to indicate more than one extra chromosome copy gained. The approximate centromere locations for the A and C chromosomes were established according to the B. napus Darmor-bzh v8.1 reference genome (Bayer et al. 2017), based on remapping of previous genetic data (Mason et al. 2016). For molecular karyotyping, a similar approach was used. If no-call (NC) SNPs covering ≥ 1 megabases (Mb) in the chromosome was observed, it was categorized as both copies missing in that particular region of the chromosome. A similar size cut-off was used for the Log2 R ratio values. Missing regions covering less than 1 Mb were not considered in the analysis due to SNP distribution and density constraints. If the duplication or deletion event was found in two copies (on both homologous chromosome sets), we classified it as a “fixed” event since this must have resulted from previous meiosis, after which a self-pollination event allowed two gametes with the same rearrangement event to come together in the same plant. On the other hand, if a rearrangement was only present in one of the homologous chromosomes, it was considered a segregating rearrangement. If a rearrangement was not present in either of the parents but was detected in the next generation, it was classified as a de novo event. Putative translocations between the genomes were established based on initially scoring deletions/duplications based on the Log2 R ratio values in combination with the already known primary homology relationships between the A, B, and C genomes (Chalhoub et al. 2014; Perumal et al. 2020) (Fig. 2, Supplementary Table 4). The final karyotype was plotted in RStudio using the R package chromDraw (Janečka and Lysak 2016), and posterior editing was done in GIMP ver. 2.10.20.

Example of the molecular karyotyping workflow. a Genotype for chromosomes A1 and C1 in Brassica hexaploids: AA and BB: homozygous for allele A or B, respectively. NC, no-call. The end of the C1 chromosome is missing. b Log2 R ratio values: values obtained from the SNP chip. Experimentally derived values were used to establish the ranges for each type of copy number variation (details in the legend). c Log R ratio values interpretation. The top of chromosome A1 is present in a single copy, and the end of A1 has duplication. In the case of C1, there is duplication at the beginning of the chromosome and a deletion (both copies missing) at the end of the chromosome. The extra copies do not have a centromere; as chromosome fragments are eliminated in mitosis without a centromere, these fragments must be located in a chromosome. d Synteny between subgenomes: Brassica homology (Chalhoub et al. 2014; Perumal et al. 2020). Homologous chromosomes can pair during meiosis, and translocations can occur. In this case, the fragment sizes and positions of the different copy number variation events suggest translocation events between A1 and C1 chromosomes. e Final drawing of the karyotype based on genotyping, Log2 R ratio, and homology between the different subgenomes
Allele segregation in the F1 hybrid
To analyze the allele segregation from the F1 hybrid, the alleles from each parent were analyzed independently. To analyze the NCJ parent, homozygous polymorphic alleles from the NCJ vs. carirapa parent 1/carirapa test-cross parent were filtered (NCJ allele: AA vs. carirapa parent 1/carirapa test-cross parent: BB), and vice versa for the carirapa parent. In each of the eight populations, the number of test-cross progeny ranged from 14 to 22 (Supplementary Table 1). The 1 : 1 AB:BB observed vs. expected allele segregation was tested using a X2 test with a significance level of p < 0.05.
Results
Preexisting fixed rearrangements in Brassica parental genotypes
Brassica genotypes used to produce the carirapa and NCJ allohexaploids were analyzed for fixed rearrangements in the genome. The parental genotypes used to produce the carirapa hexaploids, B. carinata accession “03,949,” and the B. rapa genotypes “Ankangzhong” and “WulitianYC” did not have any detectable fixed rearrangement events in their corresponding genomes. In the genotype “03949,” we did observe a segregating event at the top of B01, where a single copy was missing between 0 and 13.6 Mb. In the case of the genotypes used in the crossing of NCJ hexaploids, we only observed fixed events in the B. napus genotypes. In the case of the B. napus genotype “Surpass400_024DH” (N1), we detected deletions in chromosomes A01 and A04, located at 2.2–4.7 Mb and 20.7–23.3 Mb, respectively. In the C genome of “Surpass400_024DH,” we identified a deletion at the top of chromosome C01, located between 1.4 and 3.3 Mb. We also found two other deletion events located in chromosome C09, at positions 0–2 and 52.9–60.1 Mb, respectively. For the “Ag-Spectrum” genotype (N6), we observed a deletion on chromosome C02, located between 8–10.7 Mb.
Chromosome inheritance and fixed karyotype changes in NCJ and carirapa allohexaploid types
From the available NCJ and carirapa collection, the most advanced and fertile genotype combinations were selected. From the NCJ type, the genotypes selected were N1C2.J1 and N6C2.J2, and from the carirapa type, C13, and C21. The carirapa plants were not fully homozygous, suggesting that pollen cross-contamination with other genotypes may have occurred during propagation in the field. In the NCJ-type hybrids, residual heterozygosity was present based on the method of producing these hybrids, but no cross-contamination was indicated by the presence of alleles other than those from the parent genotypes. In total, six plants per allohexaploid type were grown and later on crossed to produce F1 hybrids between the lines (Fig. 1c).
First, the relative chromosome number for the 12 allohexaploid parental plants was determined using SNP data and Log2 R ratio values (Supplementary Table 1). For the “carirapa” type, the chromosome numbers were 51 (1 plant), 52 (2 plants), 53 (1 plant), and 54 (2 plants). In the case of the allohexaploid NCJ, the chromosome numbers observed were 50 (1 plant), 51 (1 plant), 52 (1 plant), 53 (1 plant), 54 (1 plant), and 55 (1 plant). The two carirapa plants with 54 chromosomes were aneuploids in terms of genome composition. In contrast, the NCJ plant with 54 chromosomes was an euploid individual (genotype combination N1C2.J1). Overall, in both hexaploid types, the chromosomes A05, A06, A08, A10, C03, C05, C07, B01, B02, B04, B06, and B08 showed no changes (Supplementary Fig. 1). Specifically, in the carirapa plants, no B chromosome changes were observed. Changes in B chromosomes were, however observed in the NCJ plants for chromosomes B03 (two plants), B05 (two plants), and B07 (one plant). For both allohexaploid types, most of the chromosome number changes were observed in the A01–A03, C01, and C02 chromosomes. Also, the chromosomes A01, B05, and B07 were both lost in one carirapa and two NCJ plants, respectively.
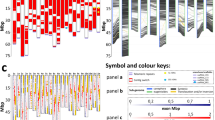
As the NCJ and carirapa allohexaploids have been independently selected by fertility (total seed number) through several generations, we analyzed the presence of fixed rearrangements (rearrangement present in both homologous chromosomes) in all the three genomes. To be able to have a better overview and frequency of all the fixed events, we combined them into one figure (Fig. 3). A single fixed event involving the B genome was detected putatively between B01/A04, where the B01 segment was lost (deletion) and replaced by the end of A04 segment (duplication) for this homologous region. The end of A04 (~ 3 Mb) was frequently lost, as was the case in seven out of 12 allohexaploid parents (four carirapa and three NCJ plants). However, this deletion event was already present in the B. napus cv. “Surpass400_024DH” used in the cross and was inherited and fixed in the three NCJ plants analyzed. In the case of the carirapa plant C21-1 that had this deletion at the end of A04, the region was most likely replaced by the end of chromosome C04, but evidence of this was inconclusive in the NCJ plants analyzed.

Karyotype of fixed events in Brassica allohexaploids NCJ (B. napus × B. carinata × B. juncea) and carirapa (B. carinata × B. rapa) type. The fixed partial or whole chromosome events are colored according to the legend for each of the three Brassica genomes
For chromosomes B05 and B07, both copies were missing in the plants belonging to the two independent N6C2.J2 lineages. Another region that was frequently lost was located at the top of chromosome C03, ranging from approximately 0–1.2 Mb in size: this region was missing in all four carirapa C13 parents (representing four plants of one lineage). The three plants from the genotype combination N1C2.J1 (one lineage) had in common a duplication/deletion event involving an extra copy of the top of chromosome A01 (~ 2 Mb) being putatively translocated into chromosome C01. The deletion of C01 was already present in the Brassica napus cv. “Surpass400_024DH” (N1) used to produce the hexaploid, unlike the duplication of A01, which was a new rearrangement event. In some regions, we found an overlap of events (duplications and deletions) between the genotypes. The region at the end of chromosome A01 was duplicated in two carirapa C13 parents (one lineage), deleted in one carirapa parent (C21), and deleted in one NCJ parent (N6C2.J2). In the homologous region, there was no detectable duplication for any of the plants, hence it was more likely to be only a deletion event. Another region of overlap was found for putative homologous translocations between chromosomes A10 and C09. The end of A10 had three duplication events (N1C2.J1) and one deletion event (C13), while C09 had one duplication (C13) and four deletions located at the end of C09 at ~ 53–60 Mb in three N1C2.J1 and at ~ 52–55 Mb for one C13 parent. For the NCJ plants from the genotype combination N1C2.J1, the putative translocation between C09/A10 was already present in the B. napus cv. “Surpass400_024DH” and was inherited and fixed in the hexaploids. The rest of the events observed in the allohexaploids NCJ and carirapa were more parent-independent (occurred in only one plant). In total, we observed 50 fixed deletion events (32 carirapa, 18 NCJ) and 33 fixed duplication events (23 carirapa and 10 NCJ) involving the A, B, and C genomes of Brassica hexaploids. The fixed events observed in the carirapa hexaploids were not identified in either the B. carinata or the B. rapa accessions used as parents.
Crossing NCJ and carirapa allohexaploids: F1 hybrid and allele segregation
Seven out of eight F1 hybrids were the result of a cross between carirapa and NCJ hexaploids, as expected (Table 1); one hybrid appeared to result from an outcross to an unknown paternal plant. The second step was to analyze the cross between the F1 hybrid and the test-cross parent. To do this, the same approach as in the F1 hybrid was used. In this case, populations 1 and 3 had only 7 and 5 individuals that corresponded to true test-cross progeny, and the remaining individuals resulted from unintended self-pollination to produce F2 progeny seeds. The other five populations were used to analyze allele segregation from the F1 hybrid.
Allele segregation per population
Population 2
The F1 hybrid was the result of a combination between the genotypes N1C2.J1 × C13. This hybrid had 52 chromosomes distributed between the A genome (19), B genome (16), and C genome (17) (Fig. 4a). In the A genome, the chromosomes A01 and A05 were present in only a single copy, with just the chromosome from the carirapa parent present. Chromosome A01 was already a single chromosome in the NCJ parent, unlike A05, where both copies were present in the parent. The other chromosomes (A02–A09) were present in two copies, while chromosome A10 had at least one extra copy from the carirapa parent. In the A genome, putative nonreciprocal translocations between A05/C04, A05/C05, A09/C09, and A10/C09 were present in the chromosomes inherited from the carirapa parent. No translocations were observed in A-genome chromosomes from the NCJ parent. The bottom of chromosome A04 from NCJ was missing 2.5 Mb, corresponding to a known deletion present in the B. napus cv. “Surpass400_024DH” used as a parent in the crossing. The top of chromosome A09 from the NCJ parent was missing ~ 3 Mb that corresponded partially to an extended deletion that was initially inherited from a deletion also present in the B. napus cv. “Surpass400_024DH”. For the B genome, the F1 hybrid contained the expected 16 chromosomes with no translocations or deleted regions detected. In the C genome, the chromosomes C01–C08 were correctly inherited from the carirapa parent. In the case of chromosome C09, just a portion (~ 5 Mb) from the end of the chromosome was present as a translocated region into chromosome A10. Part of the top of chromosome C03 (~ 7 Mb) was lost from the carirapa parent. The chromosomes C01–C09 were correctly inherited from the NCJ parent, despite the presence of only one copy of chromosome C02 in the parent. Putative translocations were observed in chromosomes coming from both parents. In the case of the carirapa parent chromosomes, the translocations present in the F1 were C01/A01 and C02/A02, and in the NCJ parent chromosomes there were C01/A01, C05/A05, C06/A07, and C09/A10. From the above translocations, only two corresponded to putative de novo events identified in the F1 hybrid.

a Molecular karyotype and b allele segregation for F1 hybrid population 2. Chromosomes are colored based on hexaploid parent: carirapa in purple, NCJ in orange. Rearrangements are colored in blue. De novo translocations (present in the F1 hybrid but not in the parents) are marked with a star in a different color depending on the type (see legend). Chromosome sizes are represented in megabases (Mb). Expected segregation ratio of the alleles (50%) is marked with a red dotted line for each chromosome. Significant allele distortion (X2 test, p < 0.05) is indicated with dark orange (NCJ) or dark purple (carirapa). Seg., segregation
Allele inheritance from the F1 hybrid into the test-cross progeny was also analyzed (Fig. 4b). Segregation distortion was established if the observed allele segregation ratio was significantly different from the expected (X2 test, p < 0.05). This test was performed independently for each of the parents of the F1 hybrid. In population 2, 18 test-cross progeny individuals were used to assess allele segregation (for more details, see materials and methods). The allele segregation from the A01 and C02 chromosomes could not be established due to the lack of enough polymorphic alleles. The segregation distortion observed in the chromosomes A05 and C09 corresponded directly to the presence of only a single chromosome in the carirapa or NCJ parent, respectively. The allele segregation distortion observed in A07, A09, A10, and C05 corresponded to rearrangements present in the F1 hybrid. Allele segregation distortion not directly explained by the karyotype of the F1 parent was observed in chromosomes A02 (extension of a known duplication present in the F1 hybrid, carirapa), A03 (although few markers were represented, carirapa), C01 (NCJ), C04 (NCJ), and B02 (NCJ). Potentially, these events correspond to de novo rearrangements produced during meiosis in the F1.
Population 4
The F1 hybrid resulting from the combination between NCJ N6C2.J2 × carirapa C13 had 51 chromosomes distributed between the A (21), B (15), and C (15) genomes (Supplementary Fig. 2a). In the F1 hybrid, chromosomes A02 and A07 had at least one extra copy from the carirapa and from the NCJ parent, respectively. These chromosomes were also doubled in the corresponding allohexaploid parent. Chromosome A04 was present as a single copy in the F1 hybrid, inherited only from the NCJ parent. In the carirapa parent, chromosome A04 was present as a single copy, and it was not inherited from the F1. In F1, the chromosome A05 inherited from the carirapa parent had a small region missing (1.6 Mb) located at chromosome position ~ 24 Mb, and no potential candidate translocation corresponding to this region was identified. In the B genome, the F1 hybrid had only one B07 chromosome (carirapa origin). Chromosome B07 was completely absent in the NCJ parent. In the B03 chromosome of the F1 hybrid (NCJ origin), a duplicated region was identified, but it was not possible to determine the position in the genome of this extra copy (colored gray, see Supplementary Fig. 2a).
In the C genome, chromosomes C01, C02, and C06 were present in single copies (NCJ). The chromosomes C01 and C02 were also present as a single copy in the carirapa parent, unlike C06, which was present as two copies in the carirapa parent and as a single copy in the NCJ parent. In the A genome, putative translocations between homoeologs A01/C01, A02/C02, A07/C07, and A09/C08 were observed. In the B genome just one putative translocation between B01/A04 (NCJ) was observed. In the C genome, we had putative translocations involving C02/A02, C04/A04, C05/B01, and C07/B02.
In the analysis of the allele segregation, most significant distortions were explained by the translocations described above (Supplementary Fig. 2b). The exceptions were present in chromosomes A02 (carirapa), A07 (carirapa), A10 (carirapa), C06 (NCJ), B01 (carirapa), and B02 (NCJ). In addition, the single B07 chromosome in the F1 hybrid was expected to be present in 50% of the test-cross population, but was only present in 18% of the individuals (3 out of 17).
Population 5
The F1 hybrid (N6C2.J2 × C13) had 54 chromosomes distributed between the A (21), B (15), and C (18) genomes (Supplementary Fig. 3a). Chromosomes with at least one extra copy were present for A04 (NCJ) and C03 (NCJ). Chromosome A04 was already doubled in the NCJ parent, unlike chromosome C03, which resulted from a new chromosome duplication event. Single copies were observed for chromosomes B05 (carirapa) and C09 (NCJ). In the case of B05, both copies were missing in the NCJ parent. Chromosome C09 was present as a single copy in the carirapa parent, and did not get inherited into the F1 hybrid. In the chromosomes from NCJ, putative translocations between A01/C01, A03/C03, A04/C04, B01/A05, C03/A03, and C08/A09 were found. In the case of chromosomes coming from the carirapa parent, we observed translocations involving the chromosomes A02/C02, A05/C04, A09/C09, C01/A01, C02/A10, and C03/A03 (Supplementary Fig. 3a).
In the population segregating for alleles from the F1 hybrid, most of the allelic distortion was explained by CNV events in the parent F1 (Supplementary Fig. 3b). The exceptions to this (putatively novel CNV events) were located in A02 (carirapa), A08 (NCJ and carirapa), B03 (NCJ), B04 (NCJ), B07 (NCJ), C02 (NCJ), C04 (NCJ and carirapa), and C08 (NCJ). Chromosome C09, as mentioned before, had just one copy in the F1 hybrid, and it was present in fewer test-cross individuals than expected (25% presence vs. 50% expected). In the case of B05 as a single chromosome, only a few polymorphic alleles were present with which to make a proper comparison, but it also seemed to be present less often than expected (25% presence vs. 50% expected).
Population 6
The F1 hybrid in this population was a cross between N6C2.J2 × C13 and had 51 chromosomes distributed between the A (19), B (15), and C (17) genomes (Supplementary Fig. 4a). Single-copy chromosomes were A04, B05, and C08. Chromosome B05 was already missing both copies in the NCJ parent. In the case of A04 and C08, both copies were present in the corresponding carirapa and NCJ parents, respectively. No extra chromosomes were observed. Putative translocations between A01/C01 and A09/C08 were detected in NCJ chromosomes. In the case of the carirapa chromosomes, observed possible translocations were located between A09/C09-C08, C01/A01, and C02/A02.
When analyzing the allele segregation for alleles from the F1 hybrid (Supplementary Fig. 4b), most of the distortion was explained by rearrangement events, with the exception of the following: end of A02 (carirapa), A03 (carirapa and NCJ), end of chromosome A04 (NCJ), A09 (carirapa), C01 (NCJ), C04 (NCJ), and C09 (NCJ). The A09 chromosome was a special case, as it was present in two copies in the F1 hybrid, but in the test-cross population, the A09 of carirapa origin was present at a higher frequency than expected (expected 50%, observed 86.6%). Interestingly, this chromosome had two translocations involving chromosomes C9 and C8. The single chromosomes A04 and C08 segregated as expected in the population, unlike B05, which was present in only two out of the 15 test-cross population individuals and not in half of them, as expected. No other changes were observed in the B genome.
Population 8
The cross between N1C2.J1 (euploid, 2n = 54) × C21 gave rise to an F1 hybrid with 52 chromosomes distributed between the A (19), B (16), and C (17) genomes (Supplementary Fig. 5a). Single chromosomes were observed for A03 and C01. Both chromosomes were present as two copies in the corresponding NCJ and carirapa parents. No chromosomes were doubled. In the NCJ chromosomes, we observed potential translocations between A02/C02, C01/A01, C06/A07, and C09/A10. Chromosomes A04 and A09 inherited from the NCJ parent had a deletion at the bottom and at the top of the chromosome, respectively. These deletions were already present in the parents, and we did not observe a duplicated homologous region that could have replaced this fragment in the F1 hybrid. In the chromosomes from the carirapa parent, we observed putative translocations between A01/C01, C02/A02, C03/A03, and C09/A09.
In the allele segregation from the F1 hybrid (Supplementary Fig. 5b), most of the events were explained by the rearrangements, with the exception of areas on chromosomes A03 (NCJ), A06 (carirapa), B02 (NCJ), B06 (NCJ), B08 (NCJ and carirapa), C08 (NCJ), and C09 (NCJ).
De novo rearrangements and inheritance in the F1 hybrids
Overall, 50 new rearrangement events (22 duplications and 28 deletions) were observed in the F1 hybrids. Out of these events, 28% (6 duplications and 8 deletions) were triggered by a previous event nearby or overlapping the chromosomic location of the new event already present in the hexaploid parent. On average, there were 10 new events per population, affecting mostly the A (52%) and C genome (44%). Both parents contributed almost equally to the de novo rearrangements observed in the F1 hybrids, with the exception of population 5, where the NCJ parent contributed to 10 rearrangements compared to 4 coming from the carirapa parent. As mentioned before, the least affected genome was the B, where just two events in two populations were observed. In the A genome, the chromosome most affected by rearrangements was A09, with a total of 8 events (2 duplications and 6 deletions). In the case of the C genome, the chromosome with more de novo rearrangements was C02, with six events (2 duplications and 4 deletions). We also observed nine de novo events involving whole chromosomes, where six chromosomes were lost, and three were present in an extra copy in the F1 hybrids.
In the F1 hybrids, we also observed that some of the rearrangement events present in the parental hexaploid plants were either inherited in the same size or reduced in size due to cross-overs. In total, 72 rearrangements were inherited from the hexaploid parental plants, with an average of 14.4 rearrangement events per F1 hybrid. Out of the 72 rearrangements, 8.3% had a reduction in size due to a cross-over. Most of the events inherited from the parents corresponded to deletions (66.7%), with seven events involving the loss of a chromosome, affecting mostly the C genome (27 events).
When analyzing the putatively de novo events produced by the F1 hybrids, we observed a total of 43 events (8.6 events on average per population). Out of these events in the F1 hybrids, 25 events were potential duplications and 18 deletions. Interestingly, many of these events also affected the B genome (9 events total, 8 from NCJ and 1 from carirapa origin, respectively). Overall, more events were produced by one meiosis in the F1 hybrid than by meioses coming from the grandparents, although the difference was only significant between the carirapa parent and the F1s (Fig. 5, one way-ANOVA, p < 0.05).

De novo rearrangements produced by meiosis in Brassica hexaploids carirapa (derived from B. carinata crossed with B. rapa), NCJ (derived from crosses between B. napus, B. carinata, and B. juncea) and their F1 hybrids. Per group, there are 5 different meiosis events representing individual gametes produced from the parents and the F1 (red: carirapa, green: NCJ and the F1 hybrid between them: blue). The number of de novo rearrangements (occurring during this specific meiosis and not inherited from a previous meiotic event in the lineage) for each of these five meiotic events for the two parents and their hybrid is indicated as a dot. The average per group is drawn with a black triangle. Significant differences are indicated with letters (Tukey’s HSD, p < 0.05): groups represented with a different letter (a, b) are significantly different, while groups that have a letter in common are not significantly different (a, ab and b, ab)
Discussion
From our study, it is clear that the new F1 hybrids produced by the cross between carirapa and NCJ hexaploid lineages were able to tolerate the inheritance of different chromosome rearrangements, despite the putative impact of these on meiosis. Initially, we hypothesized that meiosis might be more stable in the F1 compared to the parents, as heterozygosity has been linked to crossover frequency in some species (Valenzuela et al. 2012; Rowan et al. 2019), but we observed the opposite. We observed an increase in rearrangements in the F1 hybrid, which was significant compared to the carirapa parent (Fig. 5). An overall comparison may suggest that the more heterozygous the material, the higher the number of rearrangements that were produced, although further studies are needed to confirm this and to eliminate the confounding effect of the different genotypes. We also found more potential new rearrangements in the B genome of the F1 hybrids, mostly affecting alleles inherited from the NCJ parent.
From our analysis, it was also clear that Brassica NCJ and carirapa allohexaploids can also accumulate and tolerate multiple structural rearrangements over generations (Fig. 3). This occurred even under selective pressure for improved fertility (number of total seeds produced), which hypothetically would be expected to select against aneuploid chromosome complements. This is in contrast to observations of Brassica napus synthetic lines in later generations (S1:11), where fertility is majorly reduced in aneuploid lines compared to euploid lines (Xiong et al. 2011). Chromosome loss similarly affected fertility in a Brassica hexaploid mapping population from carirapa origin, where 30 out of 51 plants were infertile if missing one chromosome (Yang et al. 2018). However, Mason et al. (2014) also found no major effects of chromosome rearrangement on fertility in an NCJ allohexaploid-derived F2 population.
The expected chromosome number from the hybrids analyzed in the present study was AABBCC = 2n = 54, but this was rarely observed (Supplementary Table 1). In the case of the carirapa plants, two of them had 54 chromosomes but were not euploid. The only NCJ plant with a euploid 2n = 54 chromosome complement failed to transmit a copy of chromosome A03 to the resulting F1 hybrid, indicating that meiosis was still unstable in this plant. Our results may suggest that meiosis in these hybrids is still highly unstable but that recurrent selection for fertile, viable plants is also acting to eliminate many of these rearrangements so that plants move from euploid to aneuploid and back again, accumulating minor chromosome rearrangements along the way. Aneuploidy and nonhomologous recombination in polyploids, and particularly synthetic polyploids, are not rare events and may, in some cases, even be beneficial due to the potential for creating novel phenotypic variation (reviewed in Schiessl et al. (2019)).
Nonhomologous recombination events reflected known chromosome relationships between the A, B, and C genomes, as expected. Most translocations were observed between homologous chromosome pairs, as was previously observed in both natural and synthetic B. napus (Chalhoub et al. 2014; Samans et al. 2018) and in interspecific Brassica hybrids of various types as well as earlier generation NCJ allohexaploid populations (Mason et al. 2014; Gaebelein et al. 2019). For instance, greater changes were observed for homologous chromosomes A01/C01 and A02/C02, in accordance with results found in other synthetic B. napus lines (Gaeta et al. 2007; Xiong et al. 2011) and natural B. napus (Higgins et al. 2018). The B genome was the least affected by fixed genomic rearrangements (Fig. 3), and all events observed in this genome were present only in the NCJ allohexaploid types. A possible explanation of why the B genome was more affected in the NCJ lines compared to the carirapa lines is the different method used to generate these hybrid types. The NCJ lines were produced using a two-step crossing method, where initially, B. napus was crossed to B. carinata to form hybrids with genome compositions of 2n = CCAB = 36 (Mason et al. 2012). In these hybrids, the C genomes are present as homologous chromosomes, but the A and B genomes are haploid and occasionally pair nonhomologously with each other or with the C genome (Mason et al. 2010). As well, the preferential loss of the B genome over the A and C genome has been observed in other allohexaploid hybrids (Zhou et al. 2016). In the F1 hybrids, we observed very few rearrangements involving the B genome. In the F1 hybrid parent of population 4 (Supplementary Fig. 2a), there were two events involving the introgression of B fragments into the C genome, with a carirapa origin. These events were already present as segregating events in the carirapa parent. Based on homology between the Brassica genomes (Perumal et al. 2020), we identified the most likely position for these introgressions in the C genome. These events were very rare in all of the hexaploids analyzed, but may offer potential for introgression of chromosome fragments from the B genome carrying useful agronomic traits into the A or C genome for Brassica napus (rapeseed) crop improvement.
In the different F1 hybrids analyzed, we observed de novo rearrangements involving different chromosomes depending on the population. In some cases, these de novo rearrangements occurred close to regions where rearrangements were observed in the parental plants. As an example, in population 8, we observed an increase in the size of the deletion at the top of chromosome C03. This deletion was already present in the carirapa parent, but had a size of ~ 6.6 Mb, compared to 16.3 Mb in the F1 hybrid (Supplementary Fig. 5). This suggests an increase in size for some of the rearrangements, although it was not the rule, as other translocations were directly inherited from the parent without size change. However, already-present translocations may lead to additional irregular pairing during meiosis, as recombinant chromosomes enforce close proximity between homologous pairs in translocation heterozygotes, facilitating the production of additional crossover events (Udall et al. 2005), or it might be that those regions in general are more prone to recombine than others (De Muyt et al. 2009). Mwathi et al. (2019) also observed this effect of preexisting translocations in doubled-haploid NCJ allohexaploid lines, but due to the extreme instability of this population, de novo translocation events were found to be much more common (2.2 events per plant on average) than events triggered by preexisting rearrangements (0.8 events per plant on average). In our case, we observed on average 10 de novo rearrangements in the F1 hybrids, and out of these, 2.8 events on average were proximal to or co-located with a preexisting event. The difference observed between the two experiments might be attributed to the fact that, in our case, the analyzed plants underwent more rounds of meiosis (H3–H7) compared to just two rounds for the Mwathi lines (H2) (Mwathi et al. 2019).
Although no fixed pattern of selection was observed across all the allohexaploid genotypes, several events notably seemed to occur in multiple independent lineages or to be preferentially selected for. This was observed in 3 plants belonging to the N1C2.J1 lineage, where the beginning of chromosome A01 was doubled and putatively translocated into C01 (Fig. 3). The region translocated between A01/C01 has a length of ~ 2 Mb, and contains the meiosis gene Cell Division Cycle 20 (CDC20). The gene CDC20 has been found to be crucial in Arabidopsis thaliana meiosis, although it might have a different function in Brassica (Niu et al. 2015).
The end of chromosome A04 was deleted (~ 1.1–2.5 Mb missing) in seven plants belonging to three independent lineages (two carirapa and one NCJ, Fig. 3). In the case of the NCJ plants, the deletion of this region was already present in the parental B. napus cv. “Surpass400_024DH” and in the progenitor line “surpass” (Higgins et al. 2018). No doubling in the corresponding homologous region in C04 was observed for any of these lineages. Although no known meiosis genes fall in this region, the meiosis gene ASYNAPTIC4 (ASY4) is located 2 Mb upstream of the deleted region of A04. In Arabidopsis thaliana, this protein is necessary to complete synapsis, cross-over formation, and normal localization of interacting proteins ASY1 and ASY3 (Chambon et al. 2018). Interestingly, the homozygous presence of an ASY3 allele with a tandem duplication region has been associated with more stable autotetraploid Arabidopsis lyrata lines, as it causes a reduction in multivalent formation (Seear et al. 2020). The homologous region in C04 has recently been associated with fertility in synthetic B. napus, where a deletion of the last 1.5 Mb of chromosome C04 reduced seed number (Ferreira de Carvalho et al. 2021). However, this deletion was not associated with a reduction in the number of nonhomologous recombination events (Ferreira de Carvalho et al. 2021), suggesting that this effect may only be related to selection for fertility and not to increased stabilization of the genome. The top of chromosome C03 (~ 1.2–2.3 Mb) was also missing in four plants from the same carirapa lineage (C13, Fig. 3), most likely as the result of an event fixed very early on in this lineage, although the four plants differed slightly in the size of the deletion. This A03–C03 region has previously been identified in hexaploid lines as a translocation QTL associated with total seed number in NCJ hexaploids, where the top of chromosome C03 was replaced by a copy of A03 (Gaebelein et al. 2019). Within this region, we find a potential meiotic candidate gene called BRCA2. In Arabidopsis, the protein BRCA2 interacts in vitro with DMC1 (Dray et al. 2006) and plays a crucial role in homologous recombination (Seeliger et al. 2012).
Five populations from different crosses were analyzed for bias in allele segregation and chromosome inheritance. The B genome was least affected overall, although it was preferentially lost if present as single chromosomes (B05 and B07 in populations 4, 5, and 6). A similar case was observed for the single C09 chromosome (population 5, Supplementary Fig. 3a), which was also present less often than expected in one test-cross population. However, this was not an effect seen for all chromosomes present in a single copy: the majority (A03, A04, A05, C01, C02, C06, C08, and C09 in the different populations) were inherited as expected in the progeny (50%). Interestingly, in population 6, there was segregation distortion towards retention of chromosome A09 from the carirapa allohexaploid parent, although both chromosomes were present in the hybrid (Supplementary Fig. 4b). This chromosome inherited from the carirapa parent had putative translocations between chromosomes A09/C09 and A09/C08. Additionally, the chromosome A09 from carirapa had a translocated C09 region of 1.7 Mb, containing the meiosis gene X-ray induced 1 (XRI1). This gene has been shown to be involved in meiosis and post-meiotic stages of male gametes development in plants (Dean et al. 2009). In this F1 hybrid, we also just had one C08 chromosome (NCJ), and potentially the retention of an extra A09 occurred as a trisomy compensation of the missing homologous region. Compensatory trisomy of chromosomes has been previously described in resynthesized B. napus (e.g., trisomy of C05 chromosome and single copy A05) (Xiong et al. 2011). The gain of extra chromosomes to compensate the loss of a homologous chromosome can help to prevent further chromosomal instability by providing a partner for the single chromosome (e.g., monosomic-trisomic substitutions) and by preventing the deleterious effects of aneuploidy and changes in gene balance (Xiong et al. 2011).
In the test-cross populations 5 and 6, the end of chromosome A08 had an allele segregation distortion toward the allele inherited from the carirapa parent (Supplementary Fig. 3b and 4b). This region was ~ 1.3 Mb in size and contained the meiosis genes ADA2b and RAD51D. Both populations shared the same carirapa and NCJ genotype combination (C13 and N6C2.J2). In rice, RAD51D has been shown to be associated with the prevention of non-homologous recombination during meiosis (Zhang et al. 2020). The preferential inheritance of an allele putatively related to increased meiotic stability from B. rapa over an allele from B. napus is unexpected, as B. napus is an established allopolyploid species with good meiotic control. However, at least one B. rapa genotype has previously been identified to contain allelic variation relevant for meiotic stability in allohexaploids with B. carinata (Gupta et al. 2016). Further confirmation is needed to support this observation.
In test-cross population 4, allele inheritance distortion was observed at the end of A07, where alleles from the carirapa parent were preferentially inherited over alleles from the NCJ allohexaploids (Supplementary Fig. 2b). The region has a size of ~ 5.6 Mb and has also been previously associated with total seed number in an NCJ population (Gaebelein et al. 2019), although it is hard to point to a meiosis candidate gene as several can be found within this interval.
Conclusions
Our results show that Brassica carirapa and NCJ allohexaploids are highly tolerant of chromosome duplication and deletion events. No reproducible patterns of karyotype change were observed, although certain rearrangement events were present in more than one genotype. At the same time, no strong evidence of subgenome dominance was observed, although fewer rearrangements were found in the B subgenomes, as expected due to the lower degree of homology between the B compared to the A/C subgenomes (Perumal et al. 2020). Although genomic stability has not yet been achieved, the new allohexaploid material could be intercrossed between lineages without major effects on meiosis. This suggests the opportunity to use this material in rapeseed breeding programs, with special interest in the different translocations observed. At the same time, intercrossing between lineages offers great potential to increase genetic diversity in our material and may in future lead to allelic combinations that might be selected for to provide meiotic stability and fertility in a new Brassica allohexaploid crop.
Data availability
All data associated with this manuscript is available in the Supplementary Data files.
References
Abel S, Möllers C, Becker HC (2005) Development of synthetic Brassica napus lines for the analysis of “fixed heterosis” in allopolyploid plants. Euphytica 146:157–163. https://doi.org/10.1007/s10681-005-3364-7
Bayer PE, Hurgobin B, Golicz AA et al (2017) Assembly and comparison of two closely related Brassica napus genomes. Plant Biotechnol J 15.https://doi.org/10.1111/pbi.12742
Chalhoub B, Denoeud F, Liu S et al (2014) Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 80-(345):950–953. https://doi.org/10.1126/science.1253435
Chambon A, West A, Vezon D et al (2018) Identification of ASYNAPTIC4, a component of the meiotic chromosome axis. Plant Physiol 178:233–246. https://doi.org/10.1104/pp.17.01725
Chen S, Nelson MN, Chèver AM et al (2011) Trigenomic bridges for Brassica improvement. CRC Crit Rev Plant Sci 30:525–547. https://doi.org/10.1080/07352689.2011.615700
Cheng F, Sun R, Hou X et al (2016) Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat Genet 48:1218–1224. https://doi.org/10.1038/ng.3634
Chevre AM, Eber F, This P et al (1996) Characterization of Brassica nigra chromosomes and of blackleg resistance in B. napus - B. nigra addition lines. Plant Breed 115:113–118. https://doi.org/10.1111/j.1439-0523.1996.tb00884.x
De Muyt A, Mercier R, Mézard C, Grelon M (2009) Meiotic recombination and crossovers in plants. Genome Dyn 5:14–25. https://doi.org/10.1159/000166616
Dean PJ, Siwiec T, Waterworth WM et al (2009) A novel ATM-dependent X-ray-inducible gene is essential for both plant meiosis and gametogenesis. Plant J 58:791–802. https://doi.org/10.1111/j.1365-313X.2009.03814.x
Doyle JJ, Doyle JL (1990) Isolation of plant DNA from fresh tissues. Focus (madison) 12:13–15
Dray E, Siaud N, Dubois E, Doutriaux MP (2006) Interaction between Arabidopsis Brca2 and its partners Rad51, Dmc1, and Dss1. Plant Physiol 140:1059–1069. https://doi.org/10.1104/pp.105.075838
Ferreira de Carvalho J, Stoeckel S, Eber F et al (2021) Untangling structural factors driving genome stabilization in nascent Brassica napus allopolyploids. New Phytol nph.17308. https://doi.org/10.1111/nph.17308
Fredua-Agyeman R, Hwang SF, Strelkov SE et al (2019) Identification of Brassica accessions resistant to ‘old’ and ‘new’ pathotypes of Plasmodiophora brassicae from Canada. Plant Pathol 68:708–718. https://doi.org/10.1111/ppa.12980
Gaebelein R, Mason AS (2018) Allohexaploids in the genus Brassica. CRC Crit Rev Plant Sci 37:422–437. https://doi.org/10.1080/07352689.2018.1517143
Gaebelein R, Schiessl SV, Samans B et al (2019) Inherited allelic variants and novel karyotype changes influence fertility and genome stability in Brassica allohexaploids. New Phytol 223:965–978. https://doi.org/10.1111/nph.15804
Gaeta RT, Pires JC, Iniguez-Luy F et al (2007) Genomic changes in resynthesized Brassica napus and their effect on gene expression and phenotype. Plant Cell Online 19:3403–3417. https://doi.org/10.1105/tpc.107.054346
Gill SS, Khan NA, Tuteja N (2011) Differential cadmium stress tolerance in five Indian mustard (Brassica juncea L.) cultivars. Plant Signal Behav 6:293–300. https://doi.org/10.4161/psb.6.2.15049
Gupta M, Atri C, Agarwal N, Banga SS (2016) Development and molecular-genetic characterization of a stable Brassica allohexaploid. Theor Appl Genet 129:2085–2100. https://doi.org/10.1007/s00122-016-2759-2
Hayat S, Mir BA, Wani AS et al (2011) Screening of salt-tolerant genotypes of Brassica juncea based on photosynthetic attributes. J Plant Interact 6:53–60. https://doi.org/10.1080/17429145.2010.521592
Higgins EE, Clarke WE, Howell EC et al (2018) Detecting de novo homoeologous recombination events in cultivated Brassica napus using a genome-wide SNP array. G3 Genes. Genomes, Genet 8:2673–2683. https://doi.org/10.1534/g3.118.200118
Howard HW (1942) The effect of polyploidy and hybridity on seed size in crosses between Brassica chinensis, B. carinata, amphidiploid B. chinensis-carinata and auto-tetraploid B. chinensis. J Genet 43:105–119. https://doi.org/10.1007/BF02982749
Hu D, Zhang W, Zhang Y et al (2019) Reconstituting the genome of a young allopolyploid crop, Brassica napus, with its related species. Plant Biotechnol J 17:1106–1118. https://doi.org/10.1111/pbi.13041
Irfan M, Ahmad A, Hayat S (2014) Effect of cadmium on the growth and antioxidant enzymes in two varieties of Brassica juncea. Saudi J Biol Sci 21:125–131. https://doi.org/10.1016/J.SJBS.2013.08.001
Iwasa S (1964) Cytogenetic studies on the artificially raised trigenomic hexaploid hybrid forms in the genus Brassica. J Fac Agric Kyushu Univ 13:309–352
Janečka J, Lysak MA (2016) chromDraw: an R package for visualization of linear and circular karyotypes. Chromosome Res 24:217–223. https://doi.org/10.1007/s10577-015-9513-5
Jiang Y, Tian E, Li R et al (2007) Genetic diversity of Brassica carinata with emphasis on the interspecific crossability with B. rapa. Plant Breed 126:487–491. https://doi.org/10.1111/j.1439-0523.2007.01393.x
Katche E, Quezada-Martinez D, Katche EI et al (2019) Interspecific hybridization for Brassica crop improvement. Crop Breeding, Genet Genomics 1:e190007
Kaur H, Gupta S, Kumar N et al (2014) Progression of molecular and phenotypic diversification in resynthesized Brassica juncea (L) gene pool with determinate inflorescence. Euphytica 199:325–338. https://doi.org/10.1007/s10681-014-1133-1
Li M, Qian W, Meng J, Li Z (2004) Construction of novel Brassica napus genotypes through chromosomal substitution and elimination using interploid species hybridization. Chromosome Res 12:417–426. https://doi.org/10.1023/B:CHRO.0000034722.66981.94
Li M, Chen X, Meng J (2006) Intersubgenomic heterosis in rapeseed production with a partial new-typed containing subgenome A from and C from. Crop Sci 46:234. https://doi.org/10.2135/cropsci2004.0759
Mason AS, Huteau V, Eber F et al (2010) Genome structure affects the rate of autosyndesis and allosyndesis in AABC, BBAC and CCAB Brassica interspecific hybrids. Chromosome Res 18:655–666. https://doi.org/10.1007/s10577-010-9140-0
Mason AS, Yan G, Cowling WA, Nelson MN (2012) A new method for producing allohexaploid Brassica through unreduced gametes. Euphytica 186:277–287. https://doi.org/10.1007/s10681-011-0537-4
Mason AS, Nelson MN, Takahira J et al (2014) The fate of chromosomes and alleles in an allohexaploid Brassica population. Genetics 197:273–283. https://doi.org/10.1534/genetics.113.159574
Mei J, Qian L, Disi JO et al (2011) Identification of resistant sources against Sclerotinia sclerotiorum in Brassica species with emphasis on B. oleracea. Euphytica 177:393–399. https://doi.org/10.1007/s10681-010-0274-0
Mei J, Ding Y, Lu K et al (2013) Identification of genomic regions involved in resistance against Sclerotinia sclerotiorum from wild Brassica oleracea. Theor Appl Genet 126:549–556. https://doi.org/10.1007/s00122-012-2000-x
Meng J, Shi S, Gan L et al (1998) The production of yellow-seeded Brassica napus (AACC) through crossing interspecific hybrids of B. campestris (AA) and B. carinata (BBCC) with B. napus. Euphytica 103:329–333. https://doi.org/10.1023/A:1018646223643
Mwathi MW, Gupta M, Atri C et al (2017) Segregation for fertility and meiotic stability in novel Brassica allohexaploids. Theor Appl Genet 130:767–776. https://doi.org/10.1007/s00122-016-2850-8
Mwathi MW, Schiessl SV, Batley J, Mason AS (2019) “Doubled-haploid” allohexaploid Brassica lines lose fertility and viability and accumulate genetic variation due to genomic instability. Chromosoma 128:521–532. https://doi.org/10.1007/s00412-019-00720-w
Niu B, Wang L, Zhang L et al (2015) Arabidopsis cell division cycle 20.1 is required for normal meiotic spindle assembly and chromosome segregation. Plant Cell 27:3367–3382. https://doi.org/10.1105/tpc.15.00834
Parkin IA, Sharpe AG, Keith DJ, Lydiate DJ (1995) Identification of the A and C genomes of amphidiploid Brassica napus (oilseed rape). Genome 38:1122–1131. https://doi.org/10.1139/g95-149
Pelé A, Rousseau-Gueutin M, Chèvre AM (2018) Speciation success of polyploid plants closely relates to the regulation of meiotic recombination. Front Plant Sci 9:907
Peng G, Falk KC, Gugel RK et al (2014) Sources of resistance to Plasmodiophora brassicae (clubroot) pathotypes virulent on canola. Can J Plant Pathol 36:89–99. https://doi.org/10.1080/07060661.2013.863805
Perumal S, Koh CS, Jin L et al (2020) A high-contiguity Brassica nigra genome localizes active centromeres and defines the ancestral Brassica genome. Nat Plants 6.https://doi.org/10.1038/s41477-020-0735-y
Rahman MH (2001) Production of yellow-seeded Brassica napus through interspecific crosses. Plant Breed 120:463–472. https://doi.org/10.1046/j.1439-0523.2001.00640.x
Rowan BA, Heavens D, Feuerborn TR et al (2019) An ultra high-density Arabidopsis thaliana crossover map that refines the influences of structural variation and epigenetic features. Genetics 213:771–787. https://doi.org/10.1534/genetics.119.302406
Samans B, Snowdon R, Mason AS (2018) Homoeologous exchanges and gene losses generate diversity and differentiate the B. napus genome from that of its ancestors. Springer, Cham, pp 131–148
Schiessl SV, Katche E, Ihien E et al (2019) The role of genomic structural variation in the genetic improvement of polyploid crops. Crop J 7:127–140
Seear PJ, France MG, Gregory CL et al (2020) A novel allele of ASY3 is associated with greater meiotic stability in autotetraploid Arabidopsis lyrata. PLOS Genet 16:e1008900. https://doi.org/10.1371/journal.pgen.1008900
Seeliger K, Dukowic-Schulze S, Wurz-Wildersinn R et al (2012) BRCA2 is a mediator of RAD51- and DMC1-facilitated homologous recombination in Arabidopsis thaliana. New Phytol 193:364–375. https://doi.org/10.1111/j.1469-8137.2011.03947.x
Sjödin C, Glimelius K (1989) Brassica naponigra, a somatic hybrid resistant to Phoma lingam. Theor Appl Genet 77:651–656. https://doi.org/10.1007/BF00261238
Taylor A, Coventry E, Jones JE, Clarkson JP (2015) Resistance to a highly aggressive isolate of Sclerotinia sclerotiorum in a Brassica napus diversity set. Plant Pathol 64:932–940. https://doi.org/10.1111/ppa.12327
Tian E, Jiang Y, Chen L et al (2010) Synthesis of a Brassica trigenomic allohexaploid (B. carinata × B. rapa) de novo and its stability in subsequent generations. Theor Appl Genet 121:1431–1440. https://doi.org/10.1007/s00122-010-1399-1
U N (1935) Genome-analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Japanese J Bot 7:389–452
Udall JA, Wendel JF (2006) Polyploidy and crop improvement. Crop Sci 46:3–14. https://doi.org/10.2135/cropsci2006.07.0489tpg
Udall JA, Quijada PA, Osborn TC (2005) Detection of chromosomal rearrangements derived from homeologous recombination in four mapping populations of Brassica napus L. Genetics 169:967–979. https://doi.org/10.1534/genetics.104.033209
Valenzuela NT, Perera E, Naranjo T (2012) Dynamics of rye chromosome 1R regions with high or low crossover frequency in homology search and synapsis development. PLoS ONE 7:e36385. https://doi.org/10.1371/journal.pone.0036385
Warwick SI, Francis A, Al-Shehbaz IA (2006) Brassicaceae : species checklist and database on CD-Rom. Plant Syst Evol 259:249–258. https://doi.org/10.1007/s00606-006-0422-0
Wilson RA, Sangha MK, Banga SS et al (2014) Heat stress tolerance in relation to oxidative stress and antioxidants in Brassica juncea. J Environ Biol 35:383–387
Xiong Z, Gaeta RT, Pires JC (2011) Homoeologous shuffling and chromosome compensation maintain genome balance in resynthesized allopolyploid Brassica napus. Proc Natl Acad Sci 108:7908–7913. https://doi.org/10.1073/pnas.1014138108/-/DCSupplemental.www.pnas.org/cgi
Yang S, Chen S, Zhang K et al (2018) A high-density genetic map of an allohexaploid Brassica doubled haploid population reveals quantitative trait loci for pollen viability and fertility. Front Plant Sci 9:1–18. https://doi.org/10.3389/fpls.2018.01161
Zhang Y, Shen YY, Wu XM, Wang JB (2016) The basis of pod dehiscence: anatomical traits of the dehiscence zone and expression of eight pod shatter-related genes in four species of Brassicaceae. Biol Plant 60:343–354. https://doi.org/10.1007/s10535-016-0599-1
Zhang F, Shen Y, Miao C et al (2020) OsRAD51D promotes homologous pairing and recombination by preventing nonhomologous interactions in rice meiosis. New Phytol 227:824–839. https://doi.org/10.1111/nph.16595
Zhou J, Tan C, Cui C et al (2016) Distinct subgenome stabilities in synthesized Brassica allohexaploids. Theor Appl Genet 129:1257–1271. https://doi.org/10.1007/s00122-016-2701-7
Zou J, Zhu J, Huang S et al (2010) Broadening the avenue of intersubgenomic heterosis in oilseed Brassica. Theor Appl Genet 120:283–290. https://doi.org/10.1007/s00122-009-1201-4
Zou J, Hu D, Mason AS et al (2018) Genetic changes in a novel breeding population of Brassica napus synthesized from hundreds of crosses between B. rapa and B. carinata. Plant Biotechnol J 16:507–519. https://doi.org/10.1111/pbi.12791
Funding
Open Access funding enabled and organized by Projekt DEAL. The authors gratefully acknowledge the technical assistance of Liane Renno. This project was funded by the Sino-German Centre and the German Research Council (DFG grant MA6473/3–1, awarded to AM). The Mason lab is partially funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) under Germany’s Excellence Strategy – EXC 2070 – 390732324.
Author information
Authors and Affiliations
Contributions
DQM carried out the experimental analyses, prepared the results figures and tables, and drafted the manuscript. ASM and JZ conceptualized the experiment. JZ and JM generated the experimental material. JZ managed all plants in the field and glasshouse, including selection, crossing and self-pollination, and DNA extraction. WSZ managed the crossing, self-pollination, harvest in the field, and DNA extraction. ASM supervised DQM, ASM, JB, and JZ revised the manuscript. JB managed the generation of SNP array data for the experimental population.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Quezada-Martinez, D., Zou, J., Zhang, W. et al. Allele segregation analysis of F1 hybrids between independent Brassica allohexaploid lineages. Chromosoma 131, 147–161 (2022). https://doi.org/10.1007/s00412-022-00774-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00412-022-00774-3