
Abstract
In a time of unprecedented ecological change, understanding natural biophysical relationships between reef resilience and physical drivers is of increasing importance. This study evaluates how wave forcing structures coral reef benthic community composition and recovery trajectories after the major 2015/2016 bleaching event in the remote Chagos Archipelago, Indian Ocean. Benthic cover and substrate rugosity were quantified from digital imagery at 23 fore reef sites around a small coral atoll (Salomon) in 2020 and compared to data from a similar survey in 2006 and opportunistic surveys in intermediate years. Cluster analysis and principal component analysis show strong separation of community composition between exposed (modelled wave exposure > 1000 J m−3) and sheltered sites (< 1000 J m−3) in 2020. This difference is driven by relatively high cover of Porites sp., other massive corals, encrusting corals, soft corals, rubble and dead table corals at sheltered sites versus high cover of pavement and sponges at exposed sites. Total coral cover and rugosity were also higher at sheltered sites. Adding data from previous years shows benthic community shifts from distinct exposure-driven assemblages and high live coral cover in 2006 towards bare pavement, dead Acropora tables and rubble after the 2015/2016 bleaching event. The subsequent recovery trajectories at sheltered and exposed sites are surprisingly parallel and lead communities towards their respective pre-bleaching communities. These results demonstrate that in the absence of human stressors, community patterns on fore reefs are strongly controlled by wave exposure, even during and after widespread coral loss from bleaching events.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Coral reef structure, function and resilience are influenced by a combination of environmental and anthropogenic drivers. As reefs worldwide are degrading and shifting to alternative regimes (Pandolfi et al. 2003; Norström et al. 2009), and climate-driven bleaching events are increasing in frequency and severity (Hoegh-Guldberg 1999; Hughes et al. 2018), the understanding of these drivers in shaping reef communities and supporting recovery after disturbances is of vital importance (Hughes et al. 2010; Pandolfi et al. 2011; Page et al. 2019). Environmental drivers of reef communities include wave forcing, temperature, nutrients, primary production, carbonate chemistry and turbidity (Enochs et al. 2015; Robinson et al. 2018; Wedding et al. 2018; Ceccarelli et al. 2020), which are in turn affected by spatial gradients in waves, currents and local bathymetry. Hydrodynamics or ‘physical drivers’ are therefore a dominant force in shaping reef communities.
Spatial gradients in wave energy directly and strongly influence benthic and fish community patterns at the scale of individual islands and coral atolls (Williams et al. 2013; Jouffray et al. 2019; Karkarey et al. 2020). For instance, high wave forcing can reduce overall coral cover and favour wave-tolerant morphologies, such as encrusting corals (Dollar 1982; Storlazzi et al. 2005; Franklin et al. 2013) or may even shift the entire benthic community to a dominance by low-lying algal species, such as turf algae and crustose coralline algae (CCA) (Williams et al. 2013; Gove et al. 2015). Coral vulnerability to high wave energy is mainly determined by colony morphotype and size, with large corymbose or table corals experiencing high mortality through hydrodynamic dislodgement (Madin and Connolly 2006; Madin et al. 2014). After a disturbance, hydrodynamic forces can furthermore affect the rate at which new coral habitat is formed and old coral habitat degrades (Madin et al. 2016), potentially influencing recovery trajectories. Anthropogenic drivers such as fishing and coastal development lead to sedimentation, nutrient enrichment and overfishing of herbivores (McManus et al. 2000; Fabricius 2005) which can initiate shifts to different reef regimes (Hughes 1994; McCook 1999; Jouffray et al. 2019) and decouple natural relationships between reef assemblages and physical drivers (Williams et al. 2015; Ford et al. 2020).
To disentangle effects of anthropogenic and physical drivers, remote reefs are invaluable places to study impacts of natural environmental gradients in the absence of direct human disturbance (Gilmour et al. 2013; Hays et al. 2020). However, remote areas are often associated with access limitations, leading to sparse temporal and spatial data resolution. In order to evaluate the status and recovery potential of reef communities on a meaningful scale, strategic monitoring over large areas and with high spatial resolution is necessary. This study explores if wave exposure structures coral reef benthic community composition and recovery trajectories after a major bleaching event in the remote Chagos Archipelago, Indian Ocean. In 2020, 23 fore reef sites around the entire Salomon atoll were surveyed to address the following question: (1) Does benthic community structure around the atoll differ predictably based on wave exposure? Average community compositions at sheltered and exposed sites were then compared to cover data in 2006 and intervening years to explore the subsequent question: (2) Was reef status before and recovery trajectories after the 2015/2016 disturbance event equally impacted by wave exposure? The results increase our understanding of wave exposure as a driver of reef ecology in remote atolls and its effects on recovery trajectories after major disturbance events.
Methods
Study site and wave exposure
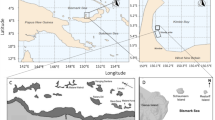
The Chagos Archipelago in the central Indian Ocean consists of five atolls with islands and numerous submerged banks (Fig. 1a). All atolls, except for Diego Garcia, have been uninhabited since the 1970s and have therefore experienced minimal direct or local impacts from fishing, sewage or shoreline modifications for the last 50 yrs (Sheppard et al. 2017). In 2010, the Archipelago and surrounding sea were declared one of the largest no-take marine protected areas, benefitting benthic and pelagic ecosystems alike (Hays et al. 2020). However, despite minimal local impacts, two major global heating events in 1997/1998 and 2015/2016 reduced coral cover values from > 40 to < 10% across the Archipelago and affected reefs down to 25 m water depth (Sheppard et al. 2017; Head et al. 2019). Reefs recovered to pre-bleaching levels 7–10 yrs after the 1997/1998 event, with coral cover reaching a peak in 2012/2013, after which the dominating large table Acropora suffered partial mortality from white band disease (Sheppard et al. 2017). Recovery after the 2015/2016 event is currently ongoing. Due to the remoteness of the Archipelago and related access limitations, research effort in the past decade has strongly focused on repeat monitoring of few sites across the archipelago to build a time-series of observations, rather than undertaking high spatial resolution monitoring. Salomon atoll is located in the northeast of the Chagos Archipelago, covering approximately 38 km2 (Fig. 1a). A reef crest and 10 small islands enclose a shallow lagoon (< 30 m depth), harbouring sheltered backreef and coral knoll habitats. The fore reef terrace surrounds the whole atoll and gently slopes from 3 to ~ 10 m depth before dropping off steeply, with a passage to the lagoon on the northern side (Fig. 1c).

(adapted from Perry et al. 2015)
Location of study sites. a Location of Salomon atoll in the Chagos Archipelago and location of the Chagos Archipelago in the central Indian Ocean (inset); b Modelled wave exposure around Salomon atoll with locations of fore reef sites surveyed in 2020 (green: sites with wave exposure < 1000 J m−3, blue: > 1000 J m−3). Circled numbers indicate sites depicted in Fig. 2; Grey shading displays magnitude of wave exposure on a continuous scale; c Drone image of Salomon atoll from the most northern point looking south (channel into the lagoon on the right), photo courtesy of Robert Dunbar; d Rose diagram showing average annual wind direction, frequency and speed based on hourly wind measurements obtained from Diego Garcia airport (1973–2001)
The seasonally-shifting wind regime in the central Indian Ocean, with the predominant wind direction being from the southeast (Fig. 1d), results in marked spatial variations in wave energy around the atoll. Wave exposure at each site was modelled as a function of wind speed, wind direction, and fetch length (i.e. the distance over open ocean that wind can travel in a specific direction unobstructed by land or reefs) using a model developed for a previous study. Wind speed and direction were obtained from hourly wind measurements at Diego Garcia airport from 1973 to 2001 (n = 219,943). Fetch lengths for 16 compass directions were calculated using the USGS model (Rohweder et al. 2012) and converted into wave energy using linear wave theory and established equations (Ekebom et al. 2003, Chollet and Mumby 2012). A binary raster representing the distribution of land masses and reef crests was generated using the outputs of the Millennium Coral Reef Mapping Project at a spatial resolution of 30 m2 (Andréfouët et al. 2006) (more detailed information in ESM Perry et al. 2015). Based on the model outputs (Fig. 1b) and a natural break in the rank order of data, sites were classified into ‘exposed’ (> 1000 J m−3), encompassing northeast and southeast facing shores, or ‘sheltered’ (< 1000 J m−3), encompassing southwest and northwest facing shores. We suspect that sites at the NE margin might have slightly lower wave exposure than calculated by the model, as the prominent current runs along-shelf and has to cross larger distances over the length of the reef terrace. However, as detailed bathymetry data are not available for the area, the model cannot factor for any such reduction in energy.
Benthic community composition 2020
To determine whether benthic community structure around the atoll differs predictably based on wave exposure, a detailed survey of Salomon’s fore reefs was conducted on 22 March 2020 by circumnavigating the atoll in a clockwise direction. The 23 sites (Fig. 1b) were chosen prior to the survey by placing GPS waypoints on a map at 1 km distance from each other. At each site, the depth of the reef terrace was checked with a handheld Echotest 2 depth sounder and surveys were conducted at 6–8 m water depth. Two observers took planar photographs of the reef substrate from the surface and from ~ 3 m distance to the benthos by swimming and duck diving on a parallel transect to the reef crest in opposite directions, with a spacing of > 3 m between photographs (distance covered ~ 50 m). Both observers used Canon Powershot G7X in underwater housings with settings automatic underwater mode and raw image quality. A third observer took short videos of the substrate along the same transect at an oblique angle to evaluate the rugosity at each site, which was rated on a scale of 1–5 (5 = highest complexity) (Johnson et al. 2019).
The photographs taken from approximately 3 m above the reef surface (n = 10/site, 230 total) were uploaded to CoralNet (www.coralnet.ucsd.edu), a web-based tool for coral reef analysis supporting semi-automated annotation of images (Beijbom et al. 2015). In this study, all images were annotated manually. Fifty random points were projected on each photograph (excluding the outer 10% of the image in width and length from each edge to avoid any blurry areas caused by camera distortion) and the substrate directly below was identified to scleractinian coral genus and morphotype level (Acropora table, Acropora branching, Pocillopora branching, Stylophora branching, Other branching, Porites massive, Other massive, All encrusting), or to other categories of benthic substrate (Soft corals, Sponges, Sand, Rubble, Dead Acropora table, Pavement, Halimeda, Other macroalgae). Note that Pavement includes bare substrate, cover of crustose coralline algae (CCA) and fine turf algae, which were not easily distinguishable in the photographs. The per cent cover data for each picture were downloaded and some categories were combined due to consistently small values (< 3%; Pocillopora + Stylophora + Other branching = Other branching; Halimeda + Other macroalgae = Macroalgae). Cover values were averaged over replicates at each site to yield site-level data (accessible at https://doi.org/10.24378/exe.3523).
All statistical analyses were performed using R 4.0.3 (R Core Team 2020). To evaluate differences in community composition we used the beta diversity metric Bray–Curtis on square-root transformed site-level cover data. Hierarchical agglomerative clustering (CLUSTER analysis) and a similarity profile test (SIMPROF) were performed to group sites with similar community composition at 0.1% and 5% significance levels (‘simprof’ function in clustsig package) (Clarke et al. 2008). The resulting a posteriori grouping at p = 0.001 coincided with the a priori grouping into sheltered and exposed sites. Cluster groupings (at p = 0.001: 2 groups; at p = 0.05: 6 groups) were tested for homogeneity of dispersion and differences in community composition using permutational multivariate analysis of variance (PERMDISP and PERMANOVA) (‘betadisper’ and ‘adonis’ functions in vegan) (Oksanen et al. 2020). Subsequently, differences in cover of individual benthic categories were tested using Welch’s t-tests with Holm’s correction of p-values (‘t.test’ and ‘p.adjust’ in stats) to account for unequal variances between groups and multiple testing. Principal component analysis (PCA) was then used to visualize the detected differences in community composition (‘PCA’ in FactoMineR and ‘fviz_pca_biplot’ in factoextra) (Husson et al. 2010). To display which coral and major benthic categories drove the differences, significant correlation vectors (‘envfit’ in vegan with 999 permutations) were overlaid on the PCA plot (all except ‘Other branching’). Grouping of sites according to wave exposure (sheltered vs. exposed) was visualized by adding concentration ellipses at ellipse.level = 0.95. Statistical analyses output tables are provided in Online Resource 1 (ESM 3 and 4).
Pre-bleaching status and recovery trajectories
To determine whether pre-bleaching status and recovery trajectories after the 2015/2016 disturbance event were impacted by wave exposure, we compared the 2020 data to several previous surveys. A similar complete assessment of Salomon’s fore reefs was conducted in 2006, taking benthic photographs from the surface at 22 sites (n = 1/site) around the atoll at similar locations (Online Resource 1, ESM 1) and depth (6–8 m). These pictures were analysed in CoralNet as described above and benthic cover (accessible at https://doi.org/10.24378/exe.3523) was averaged over sheltered (n = 12) and exposed sites (n = 10) according to location on the fore reef terrace. As detailed above, a PERMANOVA was used to detect differences in community composition comparing 2006 and 2020 for both sheltered and exposed sites Online Resource 1, ESM 5), followed by Welch’s t-tests with Holm’s correction of p-values.
Additionally, cover data for several years between 2006 and 2020 were extracted from published and unpublished datasets to assist analysis of trajectories of coral recovery at both sheltered and exposed sites. These data were collected from a smaller subset of sites (Online Resource 1, ESM 1) and using different methods, but are able to give a broad and general indication of benthic trajectories. Benthic categories assigned in these datasets were the same (coral to genus and growth form level, rubble, sand etc.) or adjusted to fit photograph analysis categories (e.g. combining ‘bare rock’ and ‘turf’ to ‘pavement’). Data for 2010 and 2019 were collected using Point-Intercept transects (n = 4/site, 50 m transects, 100 points/transect) in 8 m depth (Graham et al. 2013; Benkwitt and Graham unpublished data). Data for 2016 were extracted from video transects (n = 3/site, 30 m transect, 60 still images/transect, 10 points/image) in 8–10 m depth (Head et al. 2019). Data for 2018 and 2019 were collected along 3D line-intercept transects (n = 4/site, 10 m transect, continuous cover along reef contour) in 8 m depth (Lange and Perry 2019; Lange unpublished data). Comparing data from different sites and using different methodologies introduces some uncertainty, but all surveys were conducted at similar depths (8–10 m) and the different benthic survey methods have been shown to yield comparable data in other reef settings (e.g. Beenaerts and Berghe 2005, Jokiel et al. 2015). Also, at each point in time the data were generated using the same method, therefore clearly illustrating the difference in community composition between sheltered and exposed sites independent of methodological differences.
Due to the variation in site number and location, cover data were averaged over all available replicates of sheltered or exposed sites each year (accessible at https://doi.org/10.24378/exe.3523) before coral community trajectories were visualized using non-metric multidimensional scaling (nMDS) (‘metaMDS’ function in vegan package). The metaMDS function applied square root transformation and Wisconsin double standardization of cover data before calculating Bray–Curtis dissimilarity as recommended for large abundance class scales (Oksanen et al. 2020). Correlation vectors were overlaid on the nMDS plot, with significant groups indicated on the plot (‘envfit’ function with 999 permutations).
Results and discussion
Our study demonstrates that wave exposure is a significant driver of coral reef benthic community composition and recovery trajectories at a remote and uninhabited atoll in the Indian Ocean. In 2020, sheltered reefs along the western shore had significantly higher coral cover and rugosity than exposed reefs along the eastern shore, which were characterized by flat pavement and high boring sponge cover. Temporal patterns indicate distinct exposure-driven assemblages with high live coral cover in 2006, extensive coral mortality after the 2015/2016 bleaching event, and ongoing parallel recovery trajectories towards their respective pre-bleaching communities at both sheltered and exposed sites.
Benthic community composition and influence of wave exposure
We found a strong influence of wave exposure on site-level benthic community composition, with sheltered versus exposed sites forming distinct groups both a priori (PCA, Fig. 2b) and a posteriori (CLUSTER/SIMPROF at p = 0.001; PERMANOVA: F1,21 = 21.449, p = 0.001). Differences were driven by higher cover of Porites spp., other massive corals, encrusting corals, soft corals, rubble and dead table corals at sheltered sites (variables contributing 9–14% to PC1) versus higher cover of pavement (11%) and sponges (6%) at exposed sites. Within this broad separation, there were additional sub-groups of statistically distinct benthic communities (CLUSTER/SIMPROF at p = 0.05, Fig. 2a, PERMANOVA: F5,17 = 9.050, p = 0.001). Three sites formed a small subgroup within the exposed sites (light blue), characterized by relatively high cover of branching and tabular Acropora (variables contributing 23 and 14% to PC2, respectively). Three sites in the sheltered group (pink) were characterized by relatively high cover of Porites and soft corals, but less rubble or dead Acropora tables than at most other wave protected sites, and were therefore clustered with the exposed sites in the 5% SIMPROF analysis (Fig. 2a). The other two sites that were isolated in the cluster analysis are located at the southwest corner of the atoll and were characterized by either very high branching and table Acropora cover (light green) or high dead Acropora table cover (dark green) compared to other sheltered sites.

Reef community composition around Salomon atoll. a CLUSTER/SIMPROF Analysis indicating significant differences in community composition among sites at a significance level of p = 0.001 (coloured boxes) and p = 0.05 (coloured lines). Categories driving the differences were added post analysis. Photographs in the right panel show reef community and structure at exposed and sheltered sites indicated by coloured frames and site numbers, their locations around the atoll are indicated in Fig. 1; b Principal component analysis (PCA) showing similarities in community composition in a two-dimensional space with sites coloured according to SIMPROF Cluster Analysis at p = 0.05 and symbols and ellipses denoting the gradient in wave exposure (triangle/blue: exposed, circle/green: sheltered; empty symbols represent centre points of ellipses (ellipse level = 0.95)). All displayed vectors significantly drive the differences between sites (at p < 0.05; Online Resource 1, ESM 3)
In 2020, sheltered sites had significantly higher coral cover than exposed sites (t(19.27) = 2.55, p = 0.019). However, sites at the northeast shore showed much higher coral cover (mean: 20.3%, range: 13.2–26.2%) than sites along the southeast shore (mean: 8.6%, range: 5.2–17.6%), despite all being classified as ‘exposed’ by the wave exposure model. In fact, coral cover at the former sites was similar to those along the sheltered northwest (mean: 21.6%, range: 14.0–38.6%) and southwest shores (mean: 20.1%, range: 19.8–20.4%). This discrepancy may be explained by a reduction in wave exposure at northeast compared to southeast sites, as southeasterly along-shelf currents have to cross larger distances over the reef terrace. Further reasons may be weaker exposure to other local-scale physical forcing such as internal tides or patterns of lagoon outflow (Williams et al. 2013), which were not accounted for in this study. At remote Pacific reefs, horizontal gradients in wave energy explained benthic community patterns and variation in hard coral cover at Kingman Reef, but not at Palmyra (Williams et al. 2013). A more detailed nearshore hydrodynamic model around Palmyra captured additional physical parameters and consequently found wave forcing and geomorphology to be major drivers of benthic regimes (Gove et al. 2015). Detailed bathymetric and hydrodynamic data around Salomon atoll may therefore help to further explain small-scale differences in benthic communities.
Substrate rugosity around Salomon atoll also differed significantly with wave exposure (t(18.14) = 3.65, p = 0.002). Markedly low rugosity values were recorded along the exposed southeast side of the atoll (range: 1–2), which presently consists of a flat surface of probably pre-Holocene reef rock where all new coral growth gets episodically stripped off during high wave energy events (Grigg 1998). Notable exceptions were the two easternmost sites 5 and 6 (rugosity of 2.5 and 3, respectively) which were characterized by pronounced spur and groove formations, indicating highest exposure to the main direction of wind-driven swell (Storlazzi et al. 2003; Duce et al. 2016). Rugosity at the remaining sites ranged from 2–3.5.
While some studies reported that high wave forcing favours wave-tolerant morphologies, such as encrusting and massive corals (Storlazzi et al. 2005; Madin et al. 2006; Gove et al. 2015), we found these taxa had higher coverage at our wave protected sites. Specifically, sheltered sites had significantly higher cover of massive Porites (t(10.96) = 4.96, p = 0.004) and encrusting corals (t(9.72) = 5.63, p = 0.003), while cover of branching Acropora, tabular Acropora and other branching corals was not significantly different between exposure groups. It is important to remember, however, that Salomon’s reefs in 2020 represent assemblages at four yrs post-disturbance and will inherently differ from mature communities (especially in cover of Acropora spp.). Our exposed sites were characterized by higher cover of pavement (t(20.992) = − 4.64, p = 0.002), which is consistent with reports from other remote areas, where the reefs are dominated by low-lying algal species (turf algae and CCA) at high exposure sites (Williams et al. 2013; Gove et al. 2015). Fleshy macroalgal cover in our study was generally very low (< 2% at all sites), but slightly higher at exposed than at sheltered sites. This pattern differs from observed macroalgae dominance in regions of low wave forcing where vulnerability to physical dislodgement is lowest (Gove et al. 2015), but supports studies finding lower richness, biomass and bite rates of herbivores at highly exposed sites (Karkarey et al. 2020). Dead table corals (t(9.07) = 4.77, p = 0.009) and rubble (t(11.09) = 5.70, p = 0.002) were much less prevalent at exposed sites, because they tend to be rapidly removed by periodic high energy monsoonal waves at wind exposed reefs (Yadav et al. 2016).
To summarize, in 2020 the reef structure along the exposed eastern margin of Salomon atoll consisted of a flat surface, in some parts heavily infested by Cliona spp. sponges, with relatively small branching and table coral colonies growing on top of it. In contrast, the reef structure along the sheltered western side of the atoll consisted of massive Porites colonies, recently dead coral rock which still retained a high structural complexity, and large dead Acropora tables, themselves often colonized with juvenile branching coral. The observed pattern suggests synergistic effects between average wave exposure and periodic high energy wave events from storms in structuring coral communities, similar to patterns observed around the Hawaiian Islands (Dollar 1982; Grigg 1983; Franklin et al. 2013). Despite the relatively clear impact of wave exposure on community composition, a more detailed nearshore hydrodynamic model and the inclusion of additional drivers such as temperature and nutrient concentrations would presumably capture additional physical forcings and may serve as an enhanced tool for exploring biophysical coupling in more detail (Williams et al. 2013; Gove et al. 2015).
Pre-bleaching status and recovery trajectories
A key question arising from the above observations relates to the extent to which reef communities around Salomon atoll differed before the 2015/2016 bleaching event. Our analysis of photographs from 2006 shows distinct spatial community patterns (PERMANOVA: F1,20 = 4.793, p = 0.002), with higher cover of total live coral (t(19.05) = 2.49, p = 0.022) and table Acropora at sheltered sites, and higher cover of branching Acropora, soft corals and sponges at exposed sites (photographs in Online Resource 1, ESM 2).
Ordination of community composition over time displays a shift from live coral categories in 2006 and 2010 towards high cover of pavement, dead Acropora tables and rubble following the 2015/2016 bleaching event (Fig. 3). From 2019 onwards, coral cover and community composition show recovery trends towards pre-bleaching levels at all sites. Interestingly, the trajectories of reef communities at sheltered and exposed sites are proceeding in a surprisingly parallel way, with reefs in both exposure regimes retaining their distinct communities throughout. In contrast, we expected that the different communities would be more alike after the large-scale disturbance event or that they would show contrasting rates of recovery (e.g. Smith et al. 2008, Johns et al. 2014).

Non-metric multidimensional scaling (nMDS) of benthic communities between 2006 and 2020 at sheltered (< 1000 J m−3; green) and exposed (> 1000 J m−3; blue) fore reefs around Salomon atoll. Vectors connecting years display directional change in coral community composition. Benthic groups driving differences are displayed in grey (dark grey p < 0.05, light grey p > 0.05). Scaled points indicate mean per cent hard coral cover per year
It must be noted that reefs in 2006 do not necessarily represent pristine communities, as they reflect conditions eight years after the 1997/1998 bleaching event, when recovery was possibly still ongoing. However, the comparison of reef communities in 2006 (8 yrs post-bleaching) and 2020 (4 yrs post-bleaching) reveals significant differences. At sheltered sites, total live coral cover decreased from 48.3 ± 6.3% (mean ± SE) in 2006 to 6.8 ± 0.8% after the 2015/16 bleaching event (Lange and Perry 2019), but had recovered to 20.5 ± 2.4% by 2020 (42% of 2006 levels; t(14.08) = 4.12, p = 0.001). Remaining differences are mainly due to very low cover of tabular Acropora (30.8 ± 6.6% in 2006 and 4.1 ± 1.7% in 2020) (Fig. 4). At exposed sites, total coral cover dropped from 29 ± 4.5% in 2006 to 9.9 ± 3.5% in 2016 (Head et al. 2019) and recovered slightly to 12.5 ± 2.1% in 2020 (43% of 2006 levels; t(12.74) = 3.34, p = 0.006). As Acropora cover was comparatively low even pre-bleaching (branching: 8.8 ± 2.2%, table: 6.4 ± 3.0%), the difference is mainly due to loss of massive Porites cover (8.6 ± 2.8% in 2006 to 0.8 ± 0.4% in 2020) (Fig. 4). Dead Acropora tables were much less prevalent in 2006 (0.8 ± 0.7%) than in 2020 (10.0 ± 2.0%) at sheltered sites, and generally absent at exposed sites, where degradation of dead reef structure may be faster due to continuously high prevalence of boring sponges (14.6 ± 4.6% in 2006 and 10.4 ± 2.1% in 2020) and physical substrate stripping. However, differences in rubble and sand cover between 2006 and 2020 were small at all sites (Fig. 4), indicating that the breakdown of reef substrate after the bleaching event is still ongoing (sheltered sites) or that rubble was rapidly transported off-reef (exposed sites).

Per cent cover of main benthic categories at sheltered and exposed sites in 2006 and 2020. Note that ‘all massive corals’ includes Porites massive and other massive and that ‘pavement’ was not plotted (2006: 36 ± 5% (mean ± SE) at sheltered and 45 ± 4% at exposed sites; 2020: 53 ± 8% at sheltered and 71 ± 10% at exposed sites). Boxes depict 25th and 75th percentiles with median line. Results of Welch’s t-tests with Holm’s correction of p-values are stated if significant (* < 0.05)
Future community trajectories
The current status of Salomon’s reefs in combination with data from previous years indicates that both sheltered and exposed sites are on a trajectory of recovery to their distinct pre-bleaching communities. The process may take longer at wave exposed sites, as the recovery of massive corals is slower than that of fast growing tabular Acropora and because the sites experience a constant turnover associated with breakage, scour and abrasion (Grigg 1998; Madin et al. 2006). However, as the mechanical stability of settlement structures is critical in determining post-settlement coral survival (Yadav et al. 2016), the high prevalence of dead Acropora tables may slow recovery at sheltered sites as juveniles preferentially settle on this unstable substrate (Arthur et al. 2006; Sheppard et al. 2017).
There is no indication of coral species dominance changes compared to pre-bleaching compositions as reported for some reefs in the central Indian Ocean after the 1997/1998 bleaching event (Arthur et al. 2006; Morri et al. 2015). Consistently low macroalgal cover further suggests that reefs are unlikely to shift to algal-dominated states as observed for several reefs in the more anthropogenically impacted Seychelles after 1998 (Graham et al. 2015). Generally, most fore reefs in the Central Indian Ocean have low cover of fleshy macroalgae, probably due to high abundance of herbivorous fishes (Arthur et al. 2005, 2006; Graham et al. 2015; Morri et al. 2015). Substrata made available by the death of corals are instead colonized by fine turfing and coralline algae, which promote coral settlement. Successful coral recruitment is of especially high importance for remote reefs, as without an external supply of recruits, it is assumed that reefs will be slow to recover from severe disturbance (Roberts 1997; Graham et al. 2006; McClanahan et al. 2012). Recruit numbers across the Chagos Archipelago were indeed very low in 2017 (Sheppard et al. 2017). However, during our surveys we observed high numbers of juvenile Acropora colonies around Salomon atoll at both sheltered and exposed sites, indicating high survival rates of locally produced coral larvae. This gives reason to hope that reproductive output, recruitment, coral cover and community structure will recover to pre-disturbance levels within a decade as observed for several remote Indian Ocean reefs after the 1997/1998 bleaching event (Gilmour et al. 2013; Sheppard et al. 2017). Ultimately however, the recovery of reefs in this region will depend on the recurrence intervals and magnitudes of heat stress events in the near future (Van Hooidonk et al. 2016).
Due to its remote environment and near absence of direct human impact, Salomon atoll provided a unique opportunity to study the effects of wave exposure on reef benthic community patterns and recovery potential. The distinct communities at sheltered and exposed sites both before and after a major disturbance event confirm the dominant role of wave forcing in shaping reef benthic composition. Importantly, our results highlight that communities remained distinct during widespread coral loss in 2015/2016 and the following recovery trajectories, and that communities at all sites are on their way to pre-bleaching levels. These patterns emphasize the importance of managing local pressures on reefs to promote natural biophysical coupling and resilience to climate change in the future.
Data availability
Site information and benthic cover data from picture analysis and referenced data sets can be downloaded from University of Exeter Repository at https://doi.org/10.24378/exe.3523.
References
Andréfouët S, Muller-Karger FE, Robinson JA, Kranenburg CJ, Torres-Pulliza D, Spraggins SA, Murch B (2006) Global assessment of modern coral reef extent and diversity for regional science and management applications: a view from space. In: Proceedings of the 10th International Coral Reef Symposium 2006 Jun 28 Vol 2, pp 1732–1745. Japanese Coral Reef Society Okinawa, Japan
Arthur R, Done TJ, Marsh H (2005) Benthic recovery four years after an El Niño-induced coral mass mortality in the Lakshadweep atolls. Curr Sci 89:694–699
Arthur R, Done TJ, Marsh H, Harriott V (2006) Local processes strongly influence post-bleaching benthic recovery in the Lakshadweep Islands. Coral Reefs 25:427–440
Beenaerts N, Berghe EV (2005) Comparative study of three transect methods to assess coral cover, richness and diversity. West Indian Ocean J Mar Sci 4(1):29–38
Beijbom O, Edmunds PJ, Roelfsema C, Smith J, Kline DI, Neal BP, Dunlap MJ, Moriarty V, Fan T-Y, Tan C-J (2015) Towards automated annotation of benthic survey images: variability of human experts and operational modes of automation. PloS One 10:e0130312
Ceccarelli DM, Evans RD, Logan M, Mantel P, Puotinen M, Petus C, Russ GR, Williamson DH (2020) Long-term dynamics and drivers of coral and macroalgal cover on inshore reefs of the Great Barrier Reef Marine Park. Ecol Appl 30:e02008
Chollett I, Mumby PJ (2012) Predicting the distribution of Montastraea reefs using wave exposure. Coral Reefs 31:493–503
Clarke KR, Somerfield PJ, Gorley RN (2008) Testing of null hypotheses in exploratory community analyses: similarity profiles and biota-environment linkage. J Exp Mar Biol Ecol 366:56–69
Dollar S (1982) Wave stress and coral community structure in Hawaii. Coral Reefs 1:71–81
Duce S, Vila-Concejo A, Hamylton S, Webster JM, Bruce E, Beaman RJ (2016) A morphometric assessment and classification of coral reef spur and groove morphology. Geomorphology 265:68–83
Ekebom J, Laihonen P, Suominen T (2003) A GIS-based step-wise procedure for assessing physical exposure in fragmented archipelagos. Estuar Coast Shelf Sci 57:887–898
Enochs IC, Manzello DP, Donham EM, Kolodziej G, Okano R, Johnston L, Young C, Iguel J, Edwards CB, Fox MD, Valentino L (2015) Shift from coral to macroalgae dominance on a volcanically acidified reef. Nat Clim Chang 5(12):1083–1088
Fabricius KE (2005) Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Mar Pollut Bull 50:125–146
Ford AK, Jouffray J-B, Norström AV, Moore BR, Nugues MM, Williams GJ, Bejarano S, Magron F, Wild C, Ferse SC (2020) Local human impacts disrupt relationships between benthic reef assemblages and environmental predictors. Front Mar Sci 7:571115
Franklin EC, Jokiel PL, Donahue MJ (2013) Predictive modeling of coral distribution and abundance in the Hawaiian Islands. Mar Ecol Prog Ser 481:121–132
Gilmour JP, Smith LD, Heyward AJ, Baird AH, Pratchett MS (2013) Recovery of an isolated coral reef system following severe disturbance. Science 340:69–71
Gove JM, Williams GJ, McManus MA, Clark SJ, Ehses JS, Wedding LM (2015) Coral reef benthic regimes exhibit non-linear threshold responses to natural physical drivers. Mar Ecol Prog Ser 522:33–48
Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux JP, Robinson J (2006) Dynamic fragility of oceanic coral reef ecosystems. Proc Natl Acad Sci 103:8425–8429
Graham NAJ, Pratchett MS, McClanahan TR, Wilson SK (2013) The status of coral reef fish assemblages in the Chagos Archipelago, with implications for protected area management and climate change. In: Sheppard CRC (ed) Coral reefs of the United Kingdom overseas territories. Springer, Dordrecht, pp 253–270
Graham NAJ, Jennings S, MacNeil MA, Mouillot D, Wilson SK (2015) Predicting climate-driven regime shifts versus rebound potential in coral reefs. Nature 518:94–97
Grigg RW (1983) Community structure, succession and development of coral reefs in Hawaii. Mar Ecol Prog Ser 11:1–14
Grigg R (1998) Holocene coral reef accretion in Hawaii: a function of wave exposure and sea level history. Coral Reefs 17:263–272
Hays GC, Koldewey HJ, Andrzejaczek S, Attrill MJ, Barley S, Bayley DT, Benkwitt CE, Block B, Schallert RJ, Carlisle AB (2020) A review of a decade of lessons from one of the world’s largest MPAs: conservation gains and key challenges. Mar Biol 167:159
Head CE, Bayley DT, Rowlands G, Roche RC, Tickler DM, Rogers AD, Koldewey H, Turner JR, Andradi-Brown DA (2019) Coral bleaching impacts from back-to-back 2015–2016 thermal anomalies in the remote central Indian Ocean. Coral Reefs 38:605–618
Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of the world’s coral reefs. Mar Freshw Res 50:839–866
Hughes TP (1994) Catastrophes, phase shifts, and large-scale degradation of a Caribbean coral reef. Science 265:1547–1551
Hughes TP, Graham NA, Jackson JB, Mumby PJ, Steneck RS (2010) Rising to the challenge of sustaining coral reef resilience. Trends Ecol Evol 25:633–642
Hughes TP, Anderson KD, Connolly SR, Heron SF, Kerry JT, Lough JM, Baird AH, Baum JK, Berumen ML, Bridge TC, Claar DC, Eakin CM, Gilmour JP, Graham NAJ, Harrison H, Hobbs JPA, Hoey AS, Hoogenboom M, Lowe RJ, McCulloch MT, Pandolfi JM, Pratchett M, Schoepf V, Torda G, Wilson SK (2018) Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 359:80–83
Husson F, Lê S, Pagès J (2010) Exploratory multivariate analysis by example using R. CRC Press, Boca Raton
Johns KA, Osborne KO, Logan M (2014) Contrasting rates of coral recovery and reassembly in coral communities on the Great Barrier Reef. Coral Reefs 33:553–563
Johnson GB, Taylor BM, Robbins WD, Franklin EC, Toonen R, Bowen B, Choat JH (2019) Diversity and structure of parrotfish assemblages across the northern Great Barrier Reef. Diversity 11:14
Jokiel PL, Rodgers KS, Brown EK, Kenyon JC, Aeby G, Smith WR, Farrell F (2015) Comparison of methods used to estimate coral cover in the Hawaiian Islands. PeerJ 3:e954
Jouffray J-B, Wedding LM, Norström AV, Donovan MK, Williams GJ, Crowder LB, Erickson AL, Friedlander AM, Graham NA, Gove JM (2019) Parsing human and biophysical drivers of coral reef regimes. Proc R Soc B 286:20182544
Karkarey R, Rathod P, Arthur R, Yadav S, Theo A, Alcoverro T (2020) Wave exposure reduces herbivory in post-disturbed reefs by filtering species composition, abundance and behaviour of key fish herbivores. Sci Rep 10:9854
Lange ID, Perry CT (2019) Bleaching impacts on carbonate production in the Chagos Archipelago: influence of functional coral groups on carbonate budget trajectories. Coral Reefs 38:619–624
Madin JS, Connolly SR (2006) Ecological consequences of major hydrodynamic disturbances on coral reefs. Nature 444:477–480
Madin JS, Black KP, Connolly SR (2006) Scaling water motion on coral reefs: from regional to organismal scales. Coral Reefs 25:635–644
Madin JS, Baird AH, Dornelas M, Connolly SR (2014) Mechanical vulnerability explains size-dependent mortality of reef corals. Ecol Lett 17:1008–1015
Madin JS, Anderson KD, Andreasen MH, Bridge TCL, Cairns SD, Connolly SR, Darling ES, Diaz M, Falster DS, Franklin EC, Gates RD, Harmer AMT, Hoogenboom MO, Huang DW, Keith SA, Kosnik MA, Kuo CY, Lough JM, Lovelock CE, Luiz O, Martinelli J, Mizerek T, Pandolfi JM, Pochon X, Pratchett MS, Putnam HM, Roberts TE, Stat M, Wallace CC, Widman E, Baird AH (2016) The Coral Trait Database, a curated database of trait information for coral species from the global oceans. Sci Data 3:160017
McClanahan TR, Donner SD, Maynard JA, MacNeil MA, Graham NA, Maina J, Baker AC, Beger M, Campbell SJ, Darling ES (2012) Prioritizing key resilience indicators to support coral reef management in a changing climate. PloS One 7:e42884
McCook LJ (1999) Macroalgae, nutrients and phase shifts on coral reefs: scientific issues and management consequences for the Great Barrier Reef. Coral Reefs 18:357–367
McManus JW, Menez LA, Kesner-Reyes KN, Vergara SG, Ablan M (2000) Coral reef fishing and coral-algal phase shifts: implications for global reef status. ICES J Mar Sci 57:572–578
Morri C, Montefalcone M, Lasagna R, Gatti G, Rovere A, Parravicini V, Baldelli G, Colantoni P, Bianchi CN (2015) Through bleaching and tsunami: coral reef recovery in the Maldives. Mar Pollut Bull 98:188–200
Norström AV, Nyström M, Lokrantz J, Folke C (2009) Alternative states on coral reefs: beyond coral–macroalgal phase shifts. Mar Ecol Prog Ser 376:295–306
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O'Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) Vegan: community ecology package. R Package Version 2.5–7
Page CE, Leggat W, Heron SF, Choukroun SM, Lloyd J, Ainsworth TD (2019) Seeking resistance in coral reef ecosystems: the interplay of biophysical factors and bleaching resistance under a changing climate: the interplay of a reef’s biophysical factors can mitigate the coral bleaching response. BioEssays 41:1800226
Pandolfi JM, Bradbury RH, Sala E, Hughes TP, Bjorndal KA, Cooke RG, McArdle D, McClenachan L, Newman MJ, Paredes G (2003) Global trajectories of the long-term decline of coral reef ecosystems. Science 301:955–958
Pandolfi JM, Connolly SR, Marshall DJ, Cohen AL (2011) Projecting coral reef futures under global warming and ocean acidification. Science 333:418–422
Perry CT, Murphy GN, Graham NA, Wilson SK, Januchowski-Hartley FA, East HK (2015) Remote coral reefs can sustain high growth potential and may match future sea-level trends. Sci Rep 5:18289
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Roberts CM (1997) Connectivity and management of Caribbean coral reefs. Science 278:1454–1457
Robinson JP, Williams ID, Yeager LA, McPherson JM, Clark J, Oliver TA, Baum JK (2018) Environmental conditions and herbivore biomass determine coral reef benthic community composition: implications for quantitative baselines. Coral Reefs 37:1157–1168
Rohweder J, Rogala JT, Johnson BL, Anderson D, Clark S, Chamberlin F, Potter D, Runyon K (2012) Application of wind fetch and wave models for habitat rehabilitation and enhancement projects—2012 update. http://www.umesc.usgs.gov/management/dss/wind_fetch_wave_models_2012update.html
Sheppard CRC, Sheppard A, Mogg A, Bayley D, Dempsey AC, Roche R, Turner J, Purkins S (2017) Coral bleaching and mortality in the Chagos Archipelago. Atoll Res Bull 613:1–26
Smith LD, Gilmour JP, Heyward AJ (2008) Resilience of coral communities on an isolated system of reefs following catastrophic mass-bleaching. Coral Reefs 27:197–205
Storlazzi C, Logan J, Field M (2003) Quantitative morphology of a fringing reef tract from high-resolution laser bathymetry: Southern Molokai, Hawaii. Geo Soc Am Bull 115:1344–1355
Storlazzi C, Brown E, Field M, Rodgers K, Jokiel P (2005) A model for wave control on coral breakage and species distribution in the Hawaiian Islands. Coral Reefs 24:43–55
van Hooidonk R, Maynard J, Tamelander J, Gove J, Ahmadia G, Raymundo L, Williams G, Heron SF, Planes S (2016) Local-scale projections of coral reef futures and implications of the Paris Agreement. Sci Rep 6:39666
Wedding LM, Lecky J, Gove JM, Walecka HR, Donovan MK, Williams GJ, Jouffray J-B, Crowder LB, Erickson A, Falinski K, Friedlander AM, Kappel CV, Kittinger JN, McCoy K, Norström A, Nyström M, Oleson KLL, Stamoulis KA, White C, Selkoe KA (2018) Advancing the integration of spatial data to map human and natural drivers on coral reefs. PLoS One 13:e0189792
Williams GJ, Smith JE, Conklin EJ, Gove JM, Sala E, Sandin SA (2013) Benthic communities at two remote Pacific coral reefs: effects of reef habitat, depth, and wave energy gradients on spatial patterns. PeerJ 1:e81
Williams GJ, Gove JM, Eynaud Y, Zgliczynski BJ, Sandin SA (2015) Local human impacts decouple natural biophysical relationships on Pacific coral reefs. Ecography 38:751–761
Yadav S, Rathod P, Alcoverro T, Arthur R (2016) “Choice” and destiny: the substrate composition and mechanical stability of settlement structures can mediate coral recruit fate in post-bleached reefs. Coral Reefs 35:211–222
Acknowledgements
This research was funded by the Bertarelli Foundation as part of the Bertarelli Programme in Marine Science and was conducted under permit number 0003SE20. We thank Rachel Jones, Heather Koldewey and Emma Levy as well as the captain and crew of the support vessel for logistical help and for bringing us home during a global pandemic.
Author information
Authors and Affiliations
Contributions
CTP and IDL conceived the idea with CEB, JM-I, KLT, BT, MC, RLG, MP, MS and BW contributing to discussions; IDL, KLT, BT and CTP conducted the fieldwork in 2020 and HKE, NAJG, JT, CEB and IDL provided additional data; data analysis and manuscript writing were led by IDL with contributions to data analysis by CEB, JM-I and KLT and contributions to manuscript writing by CTP and NAJG. All authors have read and agreed to the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Topic Editor Morgan S. Pratchett
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lange, I.D., Benkwitt, C.E., McDevitt-Irwin, J.M. et al. Wave exposure shapes reef community composition and recovery trajectories at a remote coral atoll. Coral Reefs 40, 1819–1829 (2021). https://doi.org/10.1007/s00338-021-02184-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00338-021-02184-w