
Abstract
We analyze the long-term behavior of interacting populations which can be controlled through harvesting. The dynamics is assumed to be discrete in time and stochastic due to the effect of environmental fluctuations. We present powerful extinction and coexistence criteria when there are one or two interacting species. We then use these tools in order to see when harvesting leads to extinction or persistence of species, as well as what the optimal harvesting strategies, which maximize the expected long-term yield, look like. For single species systems, we show under certain conditions that the optimal harvesting strategy is of bang-bang type: there is a threshold under which there is no harvesting, while everything above this threshold gets harvested. We are also able to show that stochastic environmental fluctuations will, in most cases, force the expected harvesting yield to be lower than the deterministic maximal sustainable yield. The second part of the paper is concerned with the analysis of ecosystems that have two interacting species which can be harvested. In particular, we carefully study predator–prey and competitive Ricker models. We are able to analytically identify the regions in parameter space where the species coexist, one species persists and the other one goes extinct, as well as when there is bistability. We look at how one can find the optimal proportional harvesting strategy. If the system is of predator–prey type, the optimal proportional harvesting strategy is, depending on the interaction parameters and the price of predators relative to prey, either to harvest the predator to extinction and maximize the asymptotic yield of the prey or to not harvest the prey and to maximize the asymptotic harvesting yield of the predators. If the system is competitive, in certain instances it is optimal to drive one species extinct and to harvest the other one. In other cases, it is best to let the two species coexist and harvest both species while maintaining coexistence. In the setting of the competitive Ricker model, we show that if one competitor is dominant and pushes the other species to extinction, the harvesting of the dominant species can lead to coexistence.









Similar content being viewed by others

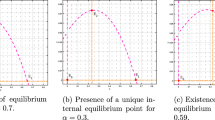
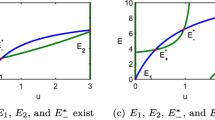
References
Alvarez, L.H.R., Hening, A.: Optimal sustainable harvesting of populations in random environments. Stoch. Process. Appl. https://doi.org/10.1016/j.spa.2019.02.008
Alvarez, L.H.R., Lungu, E., Øksendal, B.: Optimal multi-dimensional stochastic harvesting with density-dependent prices. Afr. Mat. 27(3–4), 427–442 (2016)
Alvarez, L.H.R., Shepp, L.A.: Optimal harvesting of stochastically fluctuating populations. J. Math. Biol 37, 155–177 (1998)
Anderson, C.N., Hsieh, C.-H., Sandin, S.A., Hewitt, R., Hollowed, A., Beddington, J., May, R.M., Sugihara, G.: Why fishing magnifies fluctuations in fish abundance. Nature 452(7189), 835–839 (2008)
Bayliss, P.: Population dynamics of magpie geese in relation to rainfall and density: implications for harvest models in a fluctuating environment. J. Appl. Ecol. 26, 913–924 (1989)
Benaïm, M., Schreiber, S.J.: Persistence and extinction for stochastic ecological models with internal and external variables. J. Math. Biol. 79(1), 393–431 (2019)
Cameron, T.C., O’Sullivan, D., Reynolds, A., Hicks, J.P., Piertney, S.B., Benton, T.G.: Harvested populations are more variable only in more variable environments. Ecol. Evol. 6(12), 4179–4191 (2016)
Caswell, H.: Predator-mediated coexistence: a nonequilibrium model. Am. Nat. 112(983), 127–154 (1978)
Chesson, P.L.: The stabilizing effect of a random environment. J. Math. Biol. 15(1), 1–36 (1982)
Chesson, P.L., Ellner, S.: Invasibility and stochastic boundedness in monotonic competition models. J. Math. Biol. 27(2), 117–138 (1989)
Clark, C.W.: Mathematical Bioeconomics: The Mathematics of Conservation, vol. 91. John Wiley & Sons, New Jersey (2010)
Crowley, P.H.: Predator-mediated coexistence: an equilibrium interpretation. J. Theor. Biol. 80(1), 129–144 (1979)
Dai, G., Tang, M.: Coexistence region and global dynamics of a harvested predator-prey system. SIAM J. Appl. Math. 58(1), 193–210 (1998)
Ellner, S.: Asymptotic behavior of some stochastic difference equation population models. J. Math. Biol. 19(2), 169–200 (1984)
Ellner, S.: Convergence to stationary distributions in two-species stochastic competition models. J. Math. Biol. 27(4), 451–462 (1989)
Fagerholm, H., Högnäs, G.: Stability classification of a ricker model with two random parameters. Adv. Appl. Probab. 34(1), 112–127 (2002)
Gamelon, M., Sandercock, B.K., Sæther, B.-E.: Does harvesting amplify environmentally induced population fluctuations over time in marine and terrestrial species? J. Appl. Ecol. 56(9), 2186–2194 (2019)
Getz, W.M., Haight, R.G.: Population Harvesting: Demographic Models of Fish, Forest, and Animal Resources, vol. 27. Princeton University Press, Princeton (1989)
Hening, A., Nguyen, D.: Coexistence and extinction for stochastic Kolmogorov systems. Ann. Appl. Probab. 28(3), 1893–1942 (2018)
Hening, A., Nguyen, D., Chesson, P.: A general theory of coexistence and extinction for stochastic ecological communities (2020). arXiv preprint arXiv:2007.09025
Hening, A., Nguyen, D.H.: The competitive exclusion principle in stochastic environments. J. Math. Biol. 80(5), 1323–1351 (2020)
Hening, A., Nguyen, D.H., Ungureanu, S.C., Wong, T.K.: Asymptotic harvesting of populations in random environments. J. Math. Biol. 78(1–2), 293–329 (2019)
Hening, A., Strickler, E.: On a predator-prey system with random switching that never converges to its equilibrium. SIAM J. Math. Anal. 51(5), 3625–3640 (2019)
Hening, A., Tran, K.: Harvesting and seeding of stochastic populations: analysis and numerical approximation. J. Math. Biol. 81, 65–112 (2020)
Hening, A., Tran, K., Phan, T., Yin, G.: Harvesting of interacting stochastic populations. J. Math. Biol. 79(2), 533–570 (2019)
Hilborn, R., Walters, C.J., et al.: Quantitative fisheries stock assessment: choice, dynamics and uncertainty. Rev. Fish Biol. Fish. 2(2), 177–178 (1992)
Hilker, F.M., Liz, E.: Proportional threshold harvesting in discrete-time population models. J. Math. Biol. 79(5), 1927–1951 (2019)
Hofbauer, J., Hutson, V., Jansen, W.: Coexistence for systems governed by difference equations of Lotka–Volterra type. J. Math. Biol. 25(5), 553–570 (1987)
Hofmann, E.E., Powell, T.: Environmental variability effects on marine fisheries: four case histories. Ecol. Appl. 8(sp1), S23–S32 (1998)
Hsu, S.: Predator-mediated coexistence and extinction. Math. Biosci. 54(3–4), 231–248 (1981)
Hulme, P.E.: Adapting to climate change: is there scope for ecological management in the face of a global threat? J. Appl. Ecol. 42(5), 784–794 (2005)
Larkin, P.: Some observations on models of stock and recruitment relationships for fishes. Rapp. et Proces Verbaux des Réun. Cons. Perm. Int. pour l’Explor. de la Mer 164, 316–324 (1973)
Lungu, E.M., Øksendal, B.: Optimal harvesting from a population in a stochastic crowded environment. Math. Biosci. 145(1), 47–75 (1997)
Martin, A., Ruan, S.: Predator-prey models with delay and prey harvesting. J. Math. Biol. 43(3), 247–267 (2001)
May, R.M., Beddington, J., Horwood, J., Shepherd, J.: Exploiting natural populations in an uncertain world. Math. Biosci. 42(3–4), 219–252 (1978)
Meyn, S.P., Tweedie, R.L.: Stability of Markovian processes. I. criteria for discrete-time chains. Adv. Appl. Probab. 24(3), 542–574 (1992)
Myerscough, M., Gray, B., Hogarth, W., Norbury, J.: An analysis of an ordinary differential equation model for a two-species predator-prey system with harvesting and stocking. J. Math. Biol. 30(4), 389–411 (1992)
Reed, W.J.: The steady state of a stochastic harvesting model. Math. Biosci. 41(3–4), 273–307 (1978)
Reed, W.J.: Optimal escapement levels in stochastic and deterministic harvesting models. J. Environ. Econ. Manag. 6(4), 350–363 (1979)
Rouyer, T., Sadykov, A., Ohlberger, J., Stenseth, N.C.: Does increasing mortality change the response of fish populations to environmental fluctuations? Ecol. Lett. 15(7), 658–665 (2012)
Schreiber, S.J.: Persistence for stochastic difference equations: a mini-review. J. Differ. Equ. Appl. 18(8), 1381–1403 (2012)
Schreiber, S.J., Benaïm, M., Atchadé, K.A.S.: Persistence in fluctuating environments. J. Math. Biol. 62(5), 655–683 (2011)
Shelton, A.O., Mangel, M.: Fluctuations of fish populations and the magnifying effects of fishing. Proc. Nat. Acad. Sci. 108(17), 7075–7080 (2011)
Sih, A., Crowley, P., McPeek, M., Petranka, J., Strohmeier, K.: Predation, competition, and prey communities: a review of field experiments. Ann. Rev. Ecol. Syst. 16(1), 269–311 (1985)
Slobodkin, L.: The Growth and Regulation of Animal Numbers. Holt, Reinhardt and Winston, New York (1961)
Tran, K., Yin, G.: Optimal harvesting strategies for stochastic competitive Lotka–Volterra ecosystems. Automatica 55, 236–246 (2015)
Tran, K., Yin, G.: Numerical methods for optimal harvesting strategies in random environments under partial observations. Automatica 70, 74–85 (2016)
Turelli, M.: Does environmental variability limit niche overlap? Proc. Nat. Acad. Sci. 75(10), 5085–5089 (1978)
Vellekoop, M., Högnäs, G.: A unifying framework for chaos and stochastic stability in discrete population models. J. Math. Biol. 35(5), 557–588 (1997)
Xia, J., Liu, Z., Yuan, R., Ruan, S.: The effects of harvesting and time delay on predator-prey systems with holling type ii functional response. SIAM J. Appl. Math. 70(4), 1178–1200 (2009)
Yodzis, P.: The effects of harvesting on competitive systems. Bull. Math. Biol. 38(2), 97–109 (1976)
Yodzis, P.: Harvesting and limiting similarity. Am. Nat. 111(981), 833–843 (1977)
Acknowledgements
The author thanks Dang Nguyen and Sergiu Ungureanu for helpful discussions related to the paper.
Funding
The funding was provided by Division of Mathematical Sciences (Grant No. 1853463).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Charles R. Doering.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The author is being supported by the NSF through the Grant DMS-1853463.
Appendices
Appendix A. Criteria for Persistence and Extinction
1.1 Single Species System
Suppose we have one species whose dynamics is given by
We present a few known results which give the existence of a unique invariant probability measure. These results appear in work by Ellner (1984, 1989), Vellekoop and Högnäs (1997), Fagerholm and Högnäs (2002), Schreiber (2012).
Theorem A.1
Assume that \(F(x,\xi )=xf(x,\xi )\) is continuously differentiable and strictly increasing in x, and \(f(x,\xi )\) is strictly decreasing in x. If \({\mathbb {E}}[\ln f(0,\xi _1)]>0\) and \(\lim _{x\rightarrow \infty } {\mathbb {E}}[\ln f(x,\xi _1)]<0\), then there exists a positive invariant probability measure \(\mu \) and the distribution of \(X_t\) converges weakly to \(\mu \) whenever \(X_0=x>0\).
Sometimes, if monotonicity fails, one can make use of the following result (Vellekoop and Högnäs 1997).
Theorem A.2
Assume that
where g is a positive differentiable function such that \(x\mapsto xh'(x)/h(x)\) is strictly increasing on \([0,\infty )\). Assume \({\mathbb {E}}\xi _1, {\mathbb {E}}\xi _1^2 <\infty \) and \(\xi _1\) has a positive density on (0, L) for some \(0<L<\infty \). Then, there is a positive invariant probability measure \(\mu \) and the distribution of \(X_t\) converges to \(\mu \) whenever \(X_0=x>0\).
We note that the above theorem provides a classification of the stochastic Ricker model if the random variable \(\xi _1\) has a density and is supported on (0, L) for some \(L>0\). One can also fully classify (Fagerholm and Högnäs 2002) the stochastic Ricker model if the random coefficients do not have compact support.
Theorem A.3
Consider the stochastic Ricker model \(X_{t+1} = X_t \exp (r_{t+1}-a_{t+1}X_t)\) where
-
\(r_1,\ldots \) is a sequence of i.i.d. random variables such that \({\mathbb {E}}[r_1]<\infty \) and \(r_1\) has positive density on \((-\infty ,+\infty )\),
-
\(a_1,\ldots \) is a sequence of positive i.i.d. random variables independent of \(r_t\) such that \({\mathbb {E}}[a_1]<\infty \) and
-
there exists \(x_c\) such that \({\mathbb {E}}[\exp (r_1x)]<\infty \) for all \(x\in [0, x_c]\).
Then, if \({\mathbb {E}}[r_1]<0\), \(X_t\rightarrow 0\) with probability 1, while if \({\mathbb {E}}[r_1]>0\), there is a positive invariant measure \(\mu \) such that \(X_t\) converges weakly to \(\mu \).
1.2 Two Species Systems
Suppose we have a two species system. The following result appeared in work by Ellner (1989).
Theorem A.4
Assume the following assumptions are satisfied
-
For each \(i=1,2\), there exists a positive invariant measure \(\mu _i\) such that the distribution \(\mu _i\) such that the distribution of \(X_t^i\) converges to \(\mu _i\) weakly whenever \(X_0^i>0\) and \(X_0^j=0\).
-
The mean per capita growth rates \(r_i({\mathbf {x}})\) are continuous functions.
-
The process \({\mathbf {X}}\) is irreducible on \((0,\infty )\times (0,\infty )\),
-
For any Borel measurable \(A\subset {\mathbb {R}}_+^2\) we have \({\mathbb {P}}({\mathbf {X}}_1\in A|{\mathbf {X}}_0={\mathbf {x}}_n) \rightarrow {\mathbb {P}}({\mathbf {X}}_1\in A|{\mathbf {X}}_0={\mathbf {x}})\) whenever \({\mathbf {x}}_n\rightarrow {\mathbf {x}}\).
-
For any \({\mathbf {x}}\in {\mathbb {R}}_+^2\), \(\sup _{t>0}{\mathbb {E}}[\ln ^+ X_t^i~|~{\mathbf {X}}_0={\mathbf {x}}]<\infty \) for \(i=1,2\).
If \(r_1(\mu _2)>0\) and \(r_2(\mu _1)>0\), then there exists a unique positive invariant measure \(\mu \) and the distribution of \({\mathbf {X}}_t\) converges to \(\mu \) weakly whenever \(X_0^1, X_0^2>0\).
1.3 General Criteria for Coexistence
Assume we have a general n species system modeled by
The subset \({\mathcal {S}}\subset {\mathbb {R}}_+^n\) will denote the state space of the dynamics. It will either be a compact subset of \({\mathbb {R}}_+^n\) or all of \({\mathbb {R}}_+^n\). The coexistence set is the subset \({\mathcal {S}}_+=\{{\mathbf {x}}\in {\mathcal {S}}~|~x_i>0, i=1,\ldots n\}\) of the state space where no species is extinct. We will make the following assumptions:
- (A1):
-
\(\xi _1,\ldots ,\xi _n,\ldots \) is a sequence of i.i.d. random variables taking values in a Polish space E.
- (A2):
-
For each i the fitness function \(f_i({\mathbf {x}},\xi )\) is continuous in \({\mathbf {x}}\), measurable in \(({\mathbf {x}},\xi )\) and strictly positive.
- (A3):
-
If the dynamics is unbounded: There exists a function \(V:{\mathcal {S}}_+\rightarrow {\mathbb {R}}_+\) and constants \(\gamma _1,\gamma _3, C>0\) and \(\rho \in (0,1)\) such that for all \({\mathbf {x}}\in {\mathcal {S}}_+\) we have
- (i):
-
\(V({\mathbf {x}})\ge |{\mathbf {x}}|^{\gamma _1}+1\),
- (ii):
-
\({\mathbb {E}}\left[ V({\mathbf {x}}^\top f({\mathbf {x}},\xi _1))\ell ({\mathbf {x}},\xi _1)\right] \le \rho V({\mathbf {x}})+C\), where
$$\begin{aligned} \ell ({\mathbf {x}},\xi ):=\left( \max _{i=1}^n \left\{ \max \left\{ f_i({\mathbf {x}},\xi ), \frac{1}{f_i({\mathbf {x}},\xi )}\right\} \right\} \right) ^{\gamma _3}. \end{aligned}$$
- (A4):
-
If the dynamics is bounded: There exists a constant \(\gamma _3>0\) such that for all \({\mathbf {x}}\in {\mathcal {S}}_+\) we have
$$\begin{aligned} {\mathbb {E}}\left[ \ell ({\mathbf {x}},\xi _1)\right] <\infty . \end{aligned}$$
Remark A.1
In particular, if one supposes the conditions
-
1)
There is a compact subset \(K\subset {\mathbb {R}}_+^n\times {\mathbb {R}}^{\kappa _0}\) such that all solutions \({\mathbf {X}}_t\) satisfy \({\mathbf {X}}_t\in K\) for \(t\in {\mathbb {Z}}_+\) sufficiently large;
-
2)
For all \(i=1,2,\ldots ,n \),
$$\begin{aligned} \sup _{{\mathbf {x}},\xi }|\ln f_i({\mathbf {x}},\xi )|<\infty ; \end{aligned}$$
then assumption (A4) is satisfied.
Assumptions (A1) and (A2) ensure that \({\mathbf {X}}_t\) is a Feller process that lives on \({\mathcal {S}}_{+}\), i.e., \({\mathbf {X}}_t\in {\mathcal {S}}_+, t\in {\mathbb {Z}}_+\) whenever \({\mathbf {X}}_0\in {\mathcal {S}}_{+}\). One has to make the extra assumptions (A3) or (A4) in order to ensure the process does not blow up or fluctuate too abruptly between 0 and \(\infty \). We note that most ecological models will satisfy these assumptions. For more details see the work by Benaïm and Schreiber (2019), Hening et al. (2020).
We will follow the notation, methods and results developed by Meyn and Tweedie (1992). A point \({\mathbf {y}}\in {\mathbb {R}}_+^n\) is said to be accessible from \({\mathbf {x}}\in {\mathcal {S}}_+\) if for every neighborhood U of y, there exists \(t\ge 0\) such that \(P_t({\mathbf {x}},U)>0\). Define
and for \(A\subset {\mathbb {R}}_+^n\)
Note that \(\Gamma _A\) is the set of points which are accessible from every point of A. We say a set A is accessible if for all \({\mathbf {x}}\in {\mathbb {R}}_+^{n,\circ }\)
Suppose there exist \({\mathbf {x}}^*\in \Gamma _{{\mathcal {S}}_+}\), a neighborhood U of \({\mathbf {x}}^*\), and a nonzero measure \(\phi \) on \({\mathcal {S}}_+\), such that for any \({\mathbf {x}}\in U\) there is \(m^*\in {\mathbb {Z}}_+\) such that
We will assume that such conditions are satisfied in our models. In many cases, it is not hard to check that these conditions hold—see Ellner (1989), Hening et al. (2020). Suppose the dynamics happens in either a compact subset of \({\mathbb {R}}_+^n\) or in \({\mathbb {R}}_+^n\). We denote the state space of the dynamics by \({\mathcal {S}}\). We define the extinction set, where at least one species is extinct, by
For any \(\eta >0\) let
be the subset of \({\mathcal {S}}\) where at least one species is within \(\eta \) of extinction. Denote by \({\mathcal {M}}\) the set of all ergodic invariant probability measures supported on \({\mathcal {S}}_0\) and by \({{\,\mathrm{Conv}\,}}({\mathcal {M}})\) the set of all invariant probability measures supported on \({\mathcal {S}}_0\). We say (A.2) is stochastically persistent in probability (Chesson 1982) if for all \(\varepsilon >0\) there exists \(\eta (\varepsilon )=\eta >0\) such that for all \({\mathbf {x}}\in {\mathcal {S}}_+\)
For any \(t\in {\mathbb {N}}\) define the normalized occupation measure
where \(\delta _{{\mathbf {X}}(s)}\) is the Dirac measure at \({\mathbf {X}}(s)\) and B is any Borel subset of \({\mathcal {S}}\). Note that \(\Pi _t\) is a random probability measure and \(\Pi _t(B)\) tells us the proportion of time the system spends in B up to time t. Denote the (random) set of \(\hbox {weak}^*\)-limit points of \((\Pi _t)_{t\in {\mathbb {N}}}\) by \({\mathcal {U}}={\mathcal {U}}(\omega )\). We say (A.2) is almost surely stochastically persistent (Schreiber 2012; Benaïm and Schreiber 2019) if for all \(\varepsilon >0\) there exists \(\eta (\varepsilon )=\eta >0\) such that for all \({\mathbf {x}}\in {\mathcal {S}}_+\)
The following general theorem gives us persistence for a general n species system.
Theorem A.5
Suppose that for all \(\mu \in {{\,\mathrm{Conv}\,}}({\mathcal {M}})\) we have
Then, the system is almost surely stochastically persistent and stochastically persistent in probability. Under additional irreducibility conditions, there exists a unique invariant probability measure \(\pi \) on \({\mathcal {S}}_+\) and as \(t\rightarrow \infty \) the distribution of \({\mathbf {X}}_t\) converges in total variation to \(\pi \) whenever \({\mathbf {X}}(0)={\mathbf {x}}\in {\mathcal {S}}_+\). Furthermore, if \(w:{\mathcal {S}}_+\rightarrow {\mathbb {R}}\) is continuous and either bounded or satisfies
then
Sketch of proof
First, using the Markov property and Assumption A3) one can show that for all \(t\in {\mathbb {Z}}_+\) and \({\mathbf {x}}\in {\mathcal {S}}\)
and
As a next step, one can show that if a continuous function w satisfies \(\lim _{{\mathbf {x}}\rightarrow \infty } \frac{w({\mathbf {x}})}{{\mathbb {E}}[V({\mathbf {x}}^T {\mathbf {f}}({\mathbf {x}},\xi _t))\ell ({\mathbf {x}},\xi _t)]}=0\), then w is \(\mu \)-integrable for any invariant probability measure \(\mu \) of \({\mathbf {X}}\). Moreover, the strong law of large numbers for martingales will show that
where P is the transition operator of \({\mathbf {X}}_t\). This combined with arguments by Benaïm and Schreiber (2019) implies that if \(\mu ({\mathcal {S}}_+)=1\) then \(r_i(\mu )=0\) for any \(i\in I\).
The next step is to show that there exist \(M, C_2, \gamma _4>0,\rho _2\in (0,1)\) such that
for any \({\mathbf {p}}=(p_1,\ldots ,p_n)\in {\mathbb {R}}^n\) satisfying
It is shown in Schreiber et al. (2011) by the min-max principle that Assumption (A.3) is equivalent to the existence of \({\mathbf {p}}>0\) such that
One can then prove, using arguments by Hening and Nguyen (2018), that there exists an integer \(T^*>0\) such that, for any \(T>T^*\), \({\mathbf {x}}\in {\mathcal {S}}_0, |{\mathbf {x}}|<M\) one has
Define \(U:{\mathcal {S}}_+\rightarrow {\mathbb {R}}_+\) by
with \({\mathbf {p}}\) and \(r^*\) satisfying (A.6). Let \(n^*\in {\mathbb {N}}\) be such that
Using the previous results, as well as the analysis developed by Hening and Nguyen (2018) one can prove the following: There exist numbers \(\theta \in \left( 0,\frac{\gamma _4}{2}\right) \), \(K_\theta >0\), such that for any \(T\in [T^*,n^*T^*]\cap {\mathbb {Z}}\) and \({\mathbf {x}}\in {\mathcal {S}}_+, \Vert {\mathbf {x}}\Vert \le M\),
One can show that the process \((\rho ^{-t}_2 U({\mathbf {X}}(t)))_{t\ge 0}\) is a supermartingale and use this in conjunction with the Markov property to show that there exist numbers \(\kappa =\kappa (\theta ,T^*)\in (0,1)\) and \({{\tilde{K}}}={{\tilde{K}}}(\theta ,T^*)>0\) such that
If the Markov chain \({\mathbf {X}}_t\) is irreducible and aperiodic on \({\mathcal {S}}_+\), and a compact set is petite, then one can use the well-known results by Meyn and Tweedie (1992) in conjunction with the Lyapunov condition (A.9) to conclude that there is \(c_4>1\) such that for all \({\mathbf {x}}\in {\mathcal {S}}_+\)
where \(\Vert \cdot \Vert _{TV}\) is the total variation distance. In particular, this implies that the distribution of \({\mathbf {X}}_t\) converges weakly to \(\pi \) as \(t\rightarrow \infty \). \(\square \)
Appendix B. Two Species Systems
In general, one needs stronger assumptions for extinction. We will assume for simplicity \(n\le 2\), so that we have one or two species. We need one more condition for extinction. This condition makes sure that the martingale part of \({\mathbf {X}}_t\) is bounded and that the family of occupation measures \((\Pi _t)_{t\in {\mathbb {Z}}_+}\) is tight.
A5) There exists a function \(\phi :{\mathcal {S}}\rightarrow (0,\infty )\) and constants \(C, \delta _\phi >0\) such that for all \({\mathbf {x}}\in {\mathcal {S}}\)
and
Define \({\mathcal {S}}^j:=\{{\mathbf {x}}\in {\mathcal {S}}~|~x_i=0, i\ne j\}\) to be the subspace supported by the species j. If we restrict the process to \({\mathcal {S}}^j\), then the extinction set is given by \({\mathcal {S}}_0:=\{0\}\) and the persistence set by \({\mathcal {S}}_+^j:={\mathcal {S}}^j\setminus \{0\}\). Let \({\mathcal {M}}^j:=\{\mu \in {\mathcal {M}}~|~\mu ({\mathcal {S}}^j)=1\}, {\mathcal {M}}^{j,+}:=\{\mu \in {\mathcal {M}}~|~\mu ({\mathcal {S}}_+^j)=1\}\) be the sets of ergodic probability measures on \({\mathcal {S}}^j\) and \({\mathcal {S}}^{j}_+\). We also assume that the subspaces \({\mathcal {S}}_0^1, {\mathcal {S}}_0^2, {\mathcal {S}}_+\) are accessible, i.e., we can get close to them from any starting point \({\mathbf {x}}\in {\mathcal {S}}_+\) with positive probability, and each subspace supports at most one ergodic probability measure. Consider two species interacting via the general system
The results by Chesson and Ellner (1989), Ellner (1989) assumed some type of monotonicity and only looked at competitive behavior. They can be generalized as follows (see Hening et al. 2020 for proofs). We first look at the Dirac delta measure \(\delta _0\) at the origin (0, 0)
If \(r_i(\delta _0)>0\), then species i survives on its own and converges to a unique invariant probability measure \(\mu _i\) supported on \({\mathcal {S}}_+^i := \{{\mathbf {x}}\in {\mathcal {S}}~|~x_i\ne 0, x_j=0, i\ne j\}\). Remember that the (random) set of \(\hbox {weak}^*\)-limit points of the family of occupation measures \((\Pi _t)_{t\in {\mathbb {N}}}\) is denoted by \({\mathcal {U}}={\mathcal {U}}(\omega )\). Thus, if we say that \({\mathcal {U}}(\omega )=\{\mu _1\}\), this means that for the realization \(\omega \) we have \(\Pi _t \rightarrow \mu _1\) weakly.
-
(i)
Suppose \(r_1(\delta _0)>0, r_2(\delta _0)>0\). The realized per-capita growth rates can be computed via
$$\begin{aligned} r_i(\mu _j)=\int _{(0,\infty )}{\mathbb {E}}[\ln f_i(x,\xi _1)]\mu _j(\mathrm{d}x). \end{aligned}$$-
If \(r_1(\mu _2)>0\) and \(r_2(\mu _1)>0\), we have coexistence and convergence of the distribution of \({\mathbf {X}}_t\) to the unique invariant probability measure \(\pi \) on \({\mathcal {S}}_+\).
-
If \(r_1(\mu _2)>0\) and \(r_2(\mu _1)<0\), we have the persistence of \(X^1\) and extinction of \(X^2\). In other words, for any \({\mathbf {x}}\in {\mathcal {S}}_+\)
$$\begin{aligned} {\mathbb {P}}_{\mathbf {x}}\left\{ {\mathcal {U}}(\omega )=\{\mu _1\} ~\text {and}~\lim _{t\rightarrow \infty }\frac{\ln X^2_t}{t}=r_2(\mu _1)<0, \right\} =1. \end{aligned}$$ -
If \(r_1(\mu _2)<0\) and \(r_2(\mu _1)>0\), we have the persistence of \(X^2\) and extinction of \(X^1\). In other words, for any \({\mathbf {x}}\in {\mathcal {S}}_+\)
$$\begin{aligned} {\mathbb {P}}_{\mathbf {x}}\left\{ {\mathcal {U}}(\omega )=\{\mu _2\} ~\text {and}~\lim _{t\rightarrow \infty }\frac{\ln X^1_t}{t}=r_1(\mu _2)<0, \right\} =1. \end{aligned}$$ -
If \(r_1(\mu _2)<0\) and \(r_2(\mu _1)<0\), we have that for any \({\mathbf {x}}\in {\mathcal {S}}_+\)
$$\begin{aligned} p_{{\mathbf {x}},j}:={\mathbb {P}}_{\mathbf {x}}\left\{ {\mathcal {U}}(\omega )=\{\mu _j\} ~\text {and}~\lim _{t\rightarrow \infty }\frac{\ln X^i_t}{t}=r_i(\mu _j)<0, i\ne j \right\} \end{aligned}$$and
$$\begin{aligned} p_{{\mathbf {x}},1}+ p_{{\mathbf {x}},2}=1. \end{aligned}$$
-
-
(ii)
Suppose \(r_1(\delta _0)>0, r_2(\delta _0)<0\). Then, species 1 survives on its own and converges to its unique invariant probability measure \(\mu _1\) on \({\mathcal {S}}^1_+\).
-
If \(r_2(\mu _1)>0\), we have the persistence of both species and convergence of the distribution of \({\mathbf {X}}_t\) to the unique invariant probability measure \(\pi \) on \({\mathcal {S}}_+\).
-
If \(r_2(\mu _1)<0\), we have the persistence of \(X^1\) and the extinction of \(X^2\). In other words, for any \({\mathbf {x}}\in {\mathcal {S}}_+\)
$$\begin{aligned} {\mathbb {P}}_{\mathbf {x}}\left\{ {\mathcal {U}}(\omega )=\{\mu _1\} ~\text {and}~\lim _{t\rightarrow \infty }\frac{\ln X^2_t}{t}=r_2(\mu _1)<0, \right\} =1. \end{aligned}$$
-
-
(iii)
Suppose \(r_1(\delta _0)<0, r_2(\delta _0)<0\). Then, both species go extinct with probability one. In other words, for any \({\mathbf {x}}\in {\mathcal {S}}_+\)
$$\begin{aligned} {\mathbb {P}}_{\mathbf {x}}\left\{ \lim _{t\rightarrow \infty }\frac{\ln X^i_t}{t}=r_i(\delta _0)<0 \right\} , i=1,2. \end{aligned}$$
We note that our results are significantly more general than those from Ellner (1989). In Ellner (1989), the author only gives conditions for coexistence and does not treat the possibility of the extinction of one or both species.
Rights and permissions
About this article

Cite this article
Hening, A. Coexistence, Extinction, and Optimal Harvesting in Discrete-Time Stochastic Population Models. J Nonlinear Sci 31, 1 (2021). https://doi.org/10.1007/s00332-020-09667-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00332-020-09667-0
Keywords
- Harvesting
- Ricker model
- Random environmental fluctuations
- Ecosystems
- Conservation
- Optimal harvesting strategies
- Threshold harvesting