
Abstract
Somitogenesis is the process for the development of somites in vertebrate embryos. This process is timely regulated by synchronous oscillatory expression of the segmentation clock genes. Mathematical models expressed by delay equations or ODEs have been proposed to depict the kinetics of these genes in interacting cells. Through mathematical analysis, we investigate the parameter regimes for synchronous oscillations and oscillation-arrested in an ODE model and a model with transcriptional and translational delays, both with Michaelis–Menten type degradations. Comparisons between these regimes for the two models are made. The delay model has larger capacity to accommodate synchronous oscillations. Based on the analysis and numerical computations extended from the analysis, we explore how the periods and amplitudes of the oscillations vary with the degradation rates, synthesis rates, and coupling strength. For typical parameter values, the period and amplitude increase as some synthesis rate or the coupling strength increases in the ODE model. Such variational properties of oscillations depend also on the magnitudes of time delays in delay model. We also illustrate the difference between the dynamics in systems modeled with linear degradation and the ones in systems with Michaelis–Menten type reactions for the degradation. The chief concerns are the connections between the dynamics in these models and the mechanism for the segmentation clocks, and the pertinence of mathematical modeling on somitogenesis in zebrafish.















Similar content being viewed by others

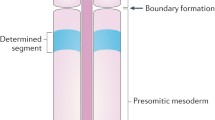
References
Asada T, Yoshida H (2003) Coefficient criterion for four-dimensional Hopf bifurcations: a complete mathematical characterization and applications to economic dynamics. Chaos Solitons Fractals 18:525–536
Ay A, Knierer S, Sperlea A, Holland J, Özbudak EM (2013) Short-lived Her proteins drive robust synchronized oscillations in the zebrafish segmentation clock. Development 140(15):3244–3253
Ay A, Holland J, Sperlea A, Devakanmalai GS, Knierer S, Sangervasi S, Stevenson A, Özbudak EM (2014) Spatial gradients of protein-level time delays set the pace of the traveling segmentation clock waves. Development 141(21):4158–4167
Baker RE, Schnell S, Maini PK (2006) A mathematical investigation of a clock and wavefornt model for somitogenesis. J Math Biol 52(4):458–482
Baker RE, Schnell S (2009) How can mathematics help us explore vertebrate segmentation? HFSP 3(1):1–5
Campanelli M, Gedeon T (2010) Somitogenesis clock-wave initiation requires differential decay and multiple binding sites for clock protein. PLoS Comput Biol 6(4):e1000728
Cinquin O (2007) Repressor dimerization in the zebrafish somitogenesis clock. PLoS Comput Biol 3(2):e32
Cooke J, Zeeman EC (1976) A clock and wavefront model for control of the number of repeated structures during animal morphogenesis. J Theor Biol 58(2):455–476
Dubrulle J, McGrew MJ, Pourquié O (2001) FGF signalling controls somite boundary position and regulates segmentation clock control of spatiotemporal Hox gene activation. Cell 106(2):219–232
Dubrulle J, Pourquié O (2002) From head to tail: links between the segmentation clock and antero-posterior patterning of the embryo. Curr Opin Genet Dev 12(5):519–523
Ermentrout GB, Terman DH (2010) Mathematical foundations of neuroscience. Springer, New York
Goldbeter A, Pourquié O (2008) Modeling the segmentation clock as a network of coupled oscillations in the Notch, Wnt and FGF signaling pathways. J Theor Biol 252(3):574–585
Görlich D, Kutay U (1999) Transport between the cell nucleus and the cytoplasm. Annu Rev Cell Dev Biol 15:607–660
Hanneman E, Westerfield M (1989) Early expression of acetyl-choline-sterase activity in functionally distinct neurons of the zebrafish. J Comp Neurol 284(3):350–361
Hassard BD, Kazarinoff ND, Wan YH (1981) Theory and applications of Hopf bifurcation. Cambridge University, Cambridge, New York
Herrgen L, Ares S, Morelli LG, Schröter C, Jülicher F, Oates AC (2010) Intercellular coupling regulates the period of the segmentation clock. Curr Biol 20(14):1244–1253
Holley SA (2007) The genetics and embryology of zebrafish metamerism. Dev Dyn 236(6):1422–1449
Horikawa K, Ishimatsu K, Yoshimoto E, Kondo S, Takeda H (2006) Noise-resistant and synchronized oscillation of the segmentation clock. Nature 441(7094):719–723
Jenkins RP, Hanisch A, Soza-Ried C, Sahai E, Lewis J (2015) Stochastic regulation of her1/7 gene expression is the source of noise in the zebrafish somite clock counteracted by Notch signalling. PLoS Comput Biol 11(11):e1004459
Jiang YJ, Aerne BL, Smithers L, Haddon C, Ish-Horowicz D, Lewis J (2000) Notch signaling and the synchronization of the somite segmentation clock. Nature 408(6811):475–479
Kawamura A, Koshida S, Hijikata H, Sakaguchi T, Kondoh H, Takada S (2005) Zebrafish hairy/enhancer of split protein links FGF signaling to cyclic gene expression in the periodic segmentation of somites. Genes Dev 19(10):1156–1161
Kuramoto Y (1984) Chemical oscillations, waves, and turbulence. Springer, Berlin
Lewis J (2003) Autoinhibition with transcriptional delay: a simple mechanism for the zebrafish somitogenesis oscillator. Curr Biol 13(16):1398–1408
Liao KL (2012) Analysis on mathematical models of somitogenesis in zebrafish. Dissertation, National Chiao Tung University, Taiwan
Liao KL, Shih CW (2012) A lattice model on somitogenesis of zebrafish. Discrete Contin Dyn Syst B 17(8):2789–2814
Liao KL, Shih CW, Tseng JP (2012) Synchronized oscillations in mathematical model of segmentation in zebrafish. Nonlinearity 25:869–904
Makarov DE (2009) Computer simulations and theory of protein translocation. Acc Chem Res 42(2):281–289
Mara A, Schroeder J, Chalouni C, Holley SA (2007) Priming, initiation and synchronization of the segmentation clock by deltaD and deltaC. Nat Cell Biol 9(5):523–530
Morelli LG, Ares S, Herrgen L, Schröter C, Jölicher F, Oates AC (2009) Delayed coupling theory of vertebrate segmentation. HFSP 3(1):55–66
Murray JD (2002) Mathematical biology. Springer, New York
Özbudak EM, Lewis J (2008) Notch signalling synchronizes the zebrafish segmentation clock but is not needed to create somite boundaries. PLoS Genet 4(2):e15
Pourquié O (2004) The chick embryo: a leading model in somitogenesis studies. Mech Dev 121(9):1069–1079
Riedel-Kruse IH, Müller C, Oates AC (2007) Synchrony dynamics during initiation, failure, and rescue of the segmentation clock. Science 317(5846):1911–1915
Schröter C, Ares S, Morelli LG, Isakova A, Hens K, Soroldoni D, Gajewski M, Jülicher F, Maerkl SJ, Deplancke B, Oates AC (2012) Topology and dynamics of the zebrafish segmentation clock core circuit. PLoS Biol 10(7):e1001364
Schwendinger-Schreck J, Kang Y, Holley SA (2014) Modeling the zebrafish segmentation clock’s gene regulatory network constrained by expression data suggests evolutionary transitions between oscillating and nonoscillating transcription. Genetics 197(2):725–738
Simon SM, Peskin CS, Oster GF (1992) What drives the translocation of proteins? PNAS 89(9):3770–3774
Shih CW, Tseng JP (2008) Convergent dynamics for multistable delayed neural networks. Nonlinearity 21:2361–2389
Shih CW, Tseng JP (2011) Global synchronization and asymptotic phases for a ring of identical cells with delayed coupling. SIAM J Math Anal 43(4):1667–1697
Sieger D, Ackermann B, Winkler C, Tautz D, Gajewski M (2006) her1 and her13.2 are jointly required for somitic border specification along the entire axis of the fish embryo. Dev Biol 293:242–251
Uriu K, Morishita Y, Iwasa Y (2009) Traveling wave formation in vertebrate segmentation. J Theor Biol 257(3):385–396
Uriu K, Morishita Y, Iwasa Y (2010) Synchronized oscillation of the segmentation clock gene in vertebrate development. J Math Biol 61:207–229
Yeung HKS, Strogatz SH (1999) Time delay in the Kuramoto model of coupled oscillators. Phys Rev Lett 82:648–651
Yu W, Cao J (2006) Stability and Hopf bifurcation analysis on a four-neuron BAM neural network with time delays. Phys Lett A 351:64–78
Zeiser S, Müller J, Liebscher V (2007) Modeling the Hes1 oscillator. J Comput Biolo 14(7):984–1000
Acknowledgements
The authors are grateful to Chia-Chieh Jay Chu for his advice on computing the bifurcation value in ODEs. The authors were supported, in part, by the Ministry of Science and Technology, Taiwan.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1: Proof of Theorem 2
Under the conditions of the theorem, the characteristic equation (30) has a pair of complex roots \(R(\mu )\pm iI(\mu )\) for \(\mu \) near \(\mu ^*\). At \(\mu = \mu ^*\), this pair becomes purely imaginary, and the other roots have negative real parts, i.e.,
according to Proposition 6. Substituting \(\lambda (\mu )=R(\mu )+iI(\mu )\) into \(\varDelta _{+}\), and collecting the real and imaginary parts, we have
where \(R=R(\mu )\), \(I=I(\mu )\). If (54) has a solution with \(I(\mu )\ne 0\), then
Thus,
Substituting (55) into (53), using \(\alpha _1(\mu ^{*})\alpha _2(\mu ^{*})\alpha _3(\mu ^{*})-\alpha _3^2(\mu ^{*})-\alpha _1^2(\mu ^{*})\alpha _4^{+}(\mu ^{*})=0\), differentiating with respect to \(\mu \), and utilizing \(R(\mu ^{*})=0\) and \(I(\mu ^{*})\ne 0\), we obtain
by the assumption of the theorem. Thus the transversality condition is met. We conclude that system (8) undergoes a Hopf bifurcation at \(\bar{\mathbb {X}}\) and \(\mu =\mu ^{*}\). \(\square \)
Appendix 2: Stability of bifurcating periodic solution, ODE case
We express system (26) into the form:
where \(\mathbb {X}=(x_1,\ldots ,x_4,y_1,\ldots ,y_4)\), \(A(\mu )\) is the linear part, and \(\mathbf{f}=(f_1,\ldots ,f_8)\) is the nonlinear term. At \(\mu =\mu ^{*}\), \(A(\mu ^*)\) has a pair of purely imaginary eigenvalues \(\lambda (\mu ^{*})=\pm i\omega _0\). Let us make a transformation so that the linear part is in normal form. We consider the change of variables \(\mathbb {X}=P\mathbb {Z}\), where
and \(\mathbf{u}\in \mathbb {R}^8\) is the eigenvector of \(A(\mu ^{*})\) corresponding to the eigenvalue \(i\omega _0\) and \(\mathbf{v}_3,\ldots ,\mathbf{v}_8\) are the generalized eigenvectors for the remaining eigenvalues. Thus
where D is an \(6\times 6\) matrix. Then the transformed system becomes
where \(\mathbb {Z}=(z_1,\ldots , z_8)\) and \(\mathbf{F}(\mathbb {Z},\mu )=P^{-1}{} \mathbf{f}(P\mathbb {Z},\mu )\) with \(\mathbf{F}=(F_1,\ldots ,F_8)\). At \(\mu =\mu ^{*}\), \(\mathbb {Z}=\mathbf{0}\), we define
Next, for \(k=3,\ldots ,8\), we set
and let \(w^{k-2}_{11}\), \(w^{k-2}_{20} \in {\mathbb C}^{6}\) be the solutions of
where D is defined in (56). For \(k=3,\ldots , 8\), let
Then we define
Appendix 3: Sketch of steps (I)–(III) in Sect. 4.2
Step (I): For any fixed \(\tau _4 \ge 0\), we substitute \(\lambda =i\omega \) with \(\omega >0\) into the characteristic equation (42), i.e., \( \triangle _{\pm }(i\omega ,r,\tau _4)=\tilde{R}_{\pm }(i\omega ,r,\tau _4)+i \tilde{I}_{\pm }(i\omega ,r,\tau _4)=0,\) where the real and imaginary parts are respectively
By the properties of trigonometric functions, (57) can be written as
where \(L_{\pm }(\omega ):=[\nu _{3}\nu _{5}(d_{4}\gamma _{1}\pm \gamma _{2}\gamma _{3} \cos {(\tau _4 \omega )})]^2+[\nu _{3}\nu _{5}(\gamma _{1} \omega \mp \gamma _{2}\gamma _{3} \sin {(\tau _4 \omega )})]^{2} >0 \), if \(\omega \) is a solution of (58), and \(\phi _{\pm }\in [0, 2\pi )\) with
In order to find solution \(\omega \) to (58), we sum up the square of equations (58) and obtain
where
Since \(Q_{\pm }(0)=\beta _{4}^{2}\mp 2 d_{4}\nu _{3}^{2}\nu _{5}^{2}\gamma _{1}\gamma _{2}\gamma _{3}\) and both \(Q_{+}(\omega )\) and \(Q_{-}(\omega )\) are strictly increasing eventually and blow up as \(\omega \rightarrow \infty \), we thus establish Proposition 7.
Proposition 7 guarantees the existence of the minimal bifurcation value \(r_c\) and its corresponding purely imaginary eigenvalue \(i \omega _c\). In the sequel, we denote by \(\omega _+\) (resp., \(\omega _-\)) a positive solution to \(Q_{+}(\cdot )=\varGamma \) (resp., \(Q_{-}(\cdot )=\varGamma \)). By using (58), we have
Let \(\sigma =+\) or −. For each solution \(\omega _{\sigma }\) to \(Q_{\sigma }\), there exists a sequence \(\{r_{\sigma }^{(k)}(\omega _{\sigma })\}_{k \in \mathbb {Z}}\):
such that \(\triangle _{\sigma }( i \omega _{\sigma },r_{\sigma }^{(k)}(\omega _{\sigma }),\tau _4)=0\). To simplify the notation, we denote \(r_{\sigma }^{(k)}:= r_{\sigma }^{(k)}(\omega _{\sigma })\) as the bifurcation value. In particular, we shall take into account the case that \(r_{\sigma }^{(k)}>0\) in the following discussions. Accordingly, a positive solution \(\omega _{+}\) (resp., \(\omega _{-}\)) of \(Q_{+}(\cdot )=\varGamma \) (resp., \(Q_{-}(\cdot )=\varGamma \)) corresponds to a pair of purely imaginary roots \(\pm i \omega _{+}\) (resp., \(\pm i \omega _{-}\)) of \(\triangle _{+}(\cdot ,r_{+}^{(k)},\tau _4)=0\) (resp., \(\triangle _{-}(\cdot ,r_-^{(k)},\tau _4)=0\)). This completes the first step.
Step (II): Since every solution of (58) is also a solution to \(Q_+\) or \(Q_-\), if \(Q^{'}_{\sigma }(\omega _{\sigma })\ne 0\), and all other positive solutions to \(Q_{+}(\cdot )=\varGamma \) and \(Q_{-}(\cdot )=\varGamma \) are not integer multiples of \(\omega _{\sigma }\), then \( i \omega _{\sigma }\) is a simple purely imaginary eigenvalue, with \(\sigma =+\) or −.
Step (III): The following conditions guarantee the transversality property:
where
Appendix 4: Sketch of steps (IV) and (V) in Sect. 4.2
Step (IV): Let us consider the characteristic equation (41) with \(r=0\):
Our goal is to derive the condition for parameters under which all roots of (60) have negative real parts for all \(\tau _4\ge 0\). If so, then the stability switch will not occur for any \(\tau _4 \ge 0\) and \(r=0\). Moreover, for any fixed \(\tau _4 \ge 0\), there is also no stability switch when \(r<r_{c}\). Combining these properties, we can then confirm the asymptotical stability of the origin for system (17) for any \(\tau _4 \ge 0\) and \(r <r_{c}\).
We have seen that all characteristic values of (60) have negative real parts when \(\tau _4=0\), under condition (33) or (34). Therefore, it suffices to show that all characteristic values of (60) remain to have negative real parts for any \(\tau _4 \ge 0\) and \(r=0\).
Proposition 8
For any \(\tau _4 \ge 0\), all characteristic values of (60) have negative real parts if
where
Proof
We substitute \(\lambda =i\nu \) with \(\nu \ge 0\) into \(\triangle _{\pm }(\lambda ,0,\tau _4)=0\) and collect the real and imaginary parts to obtain
Next, we sum up the square of equations (64) to get
where
Then \(E'(\nu )=8\nu ^7+E_5 \nu ^5+E_3 \nu ^3+E_1 \nu \), where \(E_1,~E_3,~E_5\) are defined in (63). Thus \(E(\nu )\) is strictly increasing on \([0, \infty )\), under condition (61). Moreover, it is straightforward to verify that \(E(0)=(d_{4}\nu _{3}\nu _{5}\gamma _{1}+\beta _{4})^{2}> (\nu _{3}\nu _{5}\gamma _{2}\gamma _{3})^{2}\) if (62) holds. Consequently, \(E(\nu )>(\nu _{3}\nu _{5}\gamma _{2}\gamma _{3})^{2}\), for all \(\nu \ge 0\), and hence for all \(\nu \in {\mathbb R}\), as E is an even function. Therefore, there does not exist any real solution to (65) under conditions (61) and (62). The assertion thus follows from continuity of characteristic values. This completes the proof.
Step (V): Setting \(\sigma =+\) (resp., \(\sigma =-\)) if \(Q_+(\omega _c)=\varGamma \) (resp., \(Q_-(\omega _c)=\varGamma )\), the following conditions ensure that the Hopf bifurcation occurs at \(r=r_c\) and \(\lambda =i \omega _c\):
-
Condition (B1): \(Q^{'}_\sigma (\omega _c)\ne 0\), and all other positive solutions to \(Q_{+}(\cdot )=\varGamma \) and \(Q_{-}(\cdot )=\varGamma \) satisfy \(\omega \ne m \omega _c\) for any integer m;
-
Condition (B2): \([R_\sigma (\omega _c,r_c)]^{2}+[I_\sigma (\omega _c,r_c)]^{2}\ne 0\), \(W^{2}_{1,\sigma }(\omega _c,r_c)+W^{2}_{2,\sigma }(\omega _c,r_c)\ne 0\), and \(Q_{1,\sigma }(\omega _c,r_c)W_{1,\sigma }(\omega _c,r_c)+Q_{2,\sigma }(\omega _c,r_c)W_{2,\sigma }(\omega _c,r_c) \ne 0\).
Appendix 5: Stability of bifurcating periodic solution, delay case
Step (VI): The stability of the bifurcating periodic solution is determined by the nonlinear terms of the equations. The condition for the stability can be expressed after the equations are put in a suitable form, under the theory of center manifold and normal form. Let us outline the main process for this step, following Hassard et al. (1981), see also Liao et al. (2012) and Yu and Cao (2006).
We write system (17) into the form:
where \({\mathbb X}(t)= (x_{1}(t), \ldots , x_{4}(t),y_{1}(t), \ldots , y_{4}(t))^T\), \({\mathbb X}_{t}(\theta )={\mathbb X}(t+\theta ), \theta \in [-\tau _M, 0]\), \(L_{\mu }:\mathscr {C}\rightarrow \mathbb {R}^{8}\) is a linear operator, and \(G_{\mu }:\mathscr {C}\rightarrow \mathbb {R}^{8}\) is a nonlinear operator, and the phase space \(\mathscr {C}:=\mathscr {C}([-\tau _{M},0],\mathbb {R}_+^{8})\) is a Banach space under the supremum norm \(\Vert \phi \Vert =\sup \{|\phi (\theta )|: {-\tau _{M}\le \theta \le 0}\}\). We label these operators by \(\mu :=r-r_{c}=\tau _{1}+\tau _{2}-r_{c}\), for a fixed minimal bifurcation value \(r_c\). Herein, \(L_{\mu }\) is defined by
where
as expressed in (40), with \(d_{1},d_{2},d_{3},d_{4}\) and \(\gamma _1, \gamma _2, \gamma _3\) defined in (28) and (29), respectively. In fact, according to the Riesz representation theorem, by choosing
we see that
where \(\delta (\cdot )\) is the Dirac delta function. For operator \(G_{\mu }:\mathscr {C}\rightarrow \mathbb {R}^{8}\), its eight components are
successively, where \(\phi =(\phi _1, \ldots , \phi _8)\), and
To put (66) in a suitable form, we define two operators on \(\mathscr {C}^1 := \mathscr {C}^1([-\tau _{M},0],\mathbb {R}^{8})\):
Then (66) can be recast into
The adjoint operator \(A_\mu ^{*}\) of \(A_\mu \) can be computed as
where \(\psi \in \mathscr {C}^1([0,\tau _{M}],\mathbb {R}^{8})\). In the following computation, for convenience, we allow functions to take values in \(\mathbb {C}^{8}\). We use the bilinear form
for \(\phi \in \mathscr {C}([-\tau _{M},0],\mathbb {C}^{8})\), \(\psi \in \mathscr {C}([0,\tau _{M}],\mathbb {C}^{8})\), to determine the coordinates of the center manifold near the origin of (66), where \(\eta (\theta ):=\eta (\theta ,0)\).
Next, denote by \(q(\theta )\) the eigenvector of \(A:=A_{0}\), and \(q^{*}(\theta ^{*})\) of \(A^{*}:=A^{*}_{0}\) corresponding to purely imaginary eigenvalues \(i \omega _{c}\) and \(-i\omega _{c} \), respectively, namely,
We also impose the normalized condition \(\langle q^{*},q \rangle =1\) and \(\langle q^{*},\bar{q} \rangle =0\). To this end, we assume that
for \(\theta \in [- \tau _{M},0)\), \(\theta ^{*}\in (0, \tau _{M}]\), and \(q(0)=(q_{1}, \ldots , q_{8})^{T},~ q^{*}(0)=\frac{1}{\rho }(q_{1}^{*}, \ldots , q^{*}_{8})^{T}\), where \(\rho \), \(q_{i}\) and \(q^{*}_{i}\), \(i=1,\ldots ,8\), are to be determined. Substituting (69) into (68) and evaluating at \(\theta =0\), we obtain
where \(U_{j}=e^{-i\omega _{c}\tau _{j}}\), \(j=1, 2, 4\), and \(V_{j}=i\omega _{c}+d_{j}\), for \(j=1,\ldots , 4\). Notice that \(\langle q^{*},q\rangle =1\) and \(\langle q^{*},\bar{q}\rangle =0\), if we set
where \(J_{1}:=-\gamma _1(q_{3}\overline{q}_{1}^{*}+q_{7}\overline{q}_{5}^{*}) +\gamma _2(q_{8}\overline{q}_{1}^{*}+q_{4}\overline{q}_{5}^{*}) \), \(J_{2}:= \nu _{3}(q_{1}\overline{q}_{2}^{*}+q_{5}\overline{q}_{6}^{*})\), and \(J_{4}:=-\gamma _3(q_{3}\overline{q}_{4}^{*}+q_{7}\overline{q}_{8}^{*})\).
Now, we use q and \(q^{*}\) to construct a coordinate on the center manifold \(\mathscr {M}_{0}\) at \(\mu =0\). For each \(\phi \in \mathscr {C}([-\tau _{M},0],\mathbb {C}^{8})\), we associate a pair (z, w), with \(z=\langle q^{*}, \phi \rangle \), \(w=\phi -z q -\overline{z} \overline{q}= \phi -2 \mathrm{Re}(zq)\). Let \({\mathbb X}_{t}={\mathbb X}_{t}(\theta )=(x_{1,t}(\theta ), \ldots ,x_{4,t}(\theta ),y_{1,t}(\theta ), \ldots , y_{4,t}(\theta ))^T\) be a solution of (67), and let
On the center manifold \(\mathscr {M}_{0}\), \(W(t,\theta )= w(z(t),\bar{z}(t),\theta )\), where
Herein, z and \(\bar{z}\) are the local coordinates of the center manifold \(\mathscr {M}_{0}\) in directions \(q^{*}\) and \(\overline{{q}^{*}}\), respectively.
Hence, the solution \({\mathbb X}_{t}\in \mathscr {M}_{0}\) of (67), at \(\mu =0\), satisfies
where
and
where \(w_{20}^{(k)}(\theta )\) and \(w_{11}^{(k)}(\theta )\) are the kth components of \(w_{20}(\theta )\) and \(w_{11}(\theta )\) respectively. And for \(-\tau _{M}\le \theta <0\), it can be computed that
where
With \(g_{20},~g_{11},~g_{02}\), and \(g_{21}\), we can compute \(C_{1}(r_c),~p_{2},~\zeta _{2}\), and \(T_{2}\) for the stability and the other properties for the bifurcating periodic solution.
Rights and permissions
About this article

Cite this article
Chen, KW., Liao, KL. & Shih, CW. The kinetics in mathematical models on segmentation clock genes in zebrafish. J. Math. Biol. 76, 97–150 (2018). https://doi.org/10.1007/s00285-017-1138-1
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-017-1138-1