
Abstract
Ants are able to modulate their behavior according to private and collective information. Collective information is coded in different concentrations of pheromone deposited in the environment, especially on ant trails, whereas private information is learned and memorized by individual ants. It has been shown that both kinds of information act synergistically on the collective trail-following behavior of ant colonies. Another important factor influencing ant behavior is their motivation to follow pheromone trails. Here, we show how private information and motivation modulate the response to collective information. We investigate these effects using a recently proposed approach that employs psychophysical methods to measure the response to varying pheromone concentrations. We studied the effect of private information (route memory) in the species Lasius niger, Euprenolepis procera, and Linepithema humile. Additionally, the effect of motivation was studied in the species E. procera and L. humile. Using psychophysical methods, we quantified these effects for important biological parameters like behavioral thresholds and error rates. The differential changes in these parameters between the three species imply specific adaptations to their environment.






Similar content being viewed by others

References
Aron S, Deneubourg JL, Pasteels JM (1988) Visual cues and trail-following idiosyncracy in Leptothorax unifasciatus: an orientation process during foraging. Insectes Sociaux 35:355–66
Aron S, Beckers R, Deneubourg JL, Pasteels JM (1993) Memory and chemical communication in the orientation of two mass-recruiting ant species. Insect Soc 40:369–80
Baird JC (1970) Psychophysical analysis of visual space, 1st edn. Pergamon, Oxford, New York
Beckers R, Deneubourg J, Goss S (1992) Trail laying behaviour during food recruitment in the ant Lasius niger (L.). Insect Soc 39:59–72
Beckers R, Deneubourg JL, Goss S (1993) Modulation of trail laying in the ant Lasius niger (Hymenoptera: Formicidae) and its role in the collective selection of a food source. J Insect Behav 6:751–9
Bestmann HJ, Kern F, Schafer D, Witschel MC (1992) 3,4-Dihydroisocoumarins, a new class of ant trail pheromones. Angew Chem Int Edit 6:795–6
Cavill GWK, Robertson PL, Davies NW (1979) An Argentine ant aggregation factor. Experientia (Basel) 35:989–90
Cavill GWK, Davies NW, McDonald JF (1980) Characterization of aggregation factors and associated compounds from the Argentine ant, Iridomyrmex humilis. J Chem Ecol 6:371–84
Czaczkes TJ, Grüter C, Jones SM, Ratnieks FLW (2011) Synergy between social and private information increases foraging efficiency in ants. Biol Lett 7:521–4
Czaczkes TJ, Gruter C, Ellis L, Wood E, Ratnieks FL (2013) Ant foraging on complex trails: route learning and the role of trail pheromones in Lasius niger. J Exp Biol 216:188–97
de Biseau JC, Pasteels JM (2000) Response thresholds to recruitment signals and the regulation of foraging intensity in the ant Myrmica sabuleti (Hymenoptera, Formicidae). Behav Processes 48:137–48
Dorai-Raj S (2009) Binomial confidence intervals for several parameterizations. In: R package version 10-6/r2
Evison SEF, Petchey OL, Beckerman AP, Ratnieks FLW (2008) Combined use of pheromone trails and visual landmarks by the common garden ant Lasius niger. Behav Ecol Sociobiol 63:261–7
Franks NR, Sendova-Franks AB (2013) Learning and Decision Making in a Social Context. In: Menzel R, Benjamin PR (eds) Invertebrate Learning and Memory (Handbook of Behavioral Neuroscience). Academic Press/Elsevier, Amsterdam, pp 530–545
Giraud T, Pedersen JS, Keller L (2002) Evolution of supercolonies: the Argentine ants of southern Europe. Proc Natl Acad Sci U S A 99:6075–9
Grüter C, Czaczkes TJ, Ratnieks FL (2011) Decision making in ant foragers (Lasius niger) facing conflicting private and social information. Behav Ecol Sociobiol 65:141–8
Hamilton WD (1964) The genetical evolution of social behaviour, I, II. J Theor Biol 7:1–52
Hammer M, Menzel R (1995) Learning and memory in the honeybee. J neurosci : off j Soc Neurosci 15:1617–30
Hemelrijk CK, Hildenbrandt H (2011) Some causes of the variable shape of flocks of birds. PLoS One 6:e22479
Hildenbrandt H, Carere C, Hemelrijk CK (2010) Self-organized aerial displays of thousands of starlings: a model. Behav Ecol 21:1349–59
Hölldobler B (1995) The chemistry of social regulation: multicomponent signals in ant societies. Proc Natl Acad Sci U S A 92:19–22
Hölldobler B (1999) Multimodal signals in ant communication. J Comp Physiol 184:129–41
Hölldobler B, Wilson EO (1990) The ants. Harvard University Press, Cambridge, Mass
Holway DA, Case TJ (2000) Mechanisms of dispersed central-place foraging in polydomous colonies of the Argentine ant. Anim Behav 59:433–41
Johnson RA (1991) Learning, memory, and foraging efficiency in two species of desert seed-harvester ants. Ecology 72:1408–19
Kingdom FAA, Prins N (2010) Psychophysics, 1st edn. Elsevier, London
Laland KN (2004) Social learning strategies. Learn Behav 32:4–14
Mailleux AC, Deneubourg JL, Detrain C (2000) How do ants assess food volume? Anim Behav 59:1061–9
Mailleux A-C, Buffin A, Detrain C, Deneubourg JL (2011) Recruitment in starved nests: the role of direct and indirect interactions between scouts and nestmates in the ant Lasius niger. Insect Soc 58:559–67
Mallis A (1942) Half a century with the successful Argentine ant. Sci Mon 55:536–45
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Reynolds CW (1987) Flocks, herds, and schools: a distributed behavioral model. Comput Graph 21:25–34
Robertson PL, Dudzinski ML, Orton CJ (1980) Excocrine gland involvement in trailing behaviour in the argentine ant (Formicidae: Dolichoderinae). Anim Behav 28:1255–73
Rodrigues D, Weiss MR (2012) Reward tracking and memory decay in the monarch butterfly, Danaus plexippus L. (Lepidoptera: Nymphalidae). Ethology 118:1122–31
Sarris V (2006) Relational psychophysics in humans and animals. Psychology, New York
Seifert B (2007) Die Ameisen Mittel- und Nordeuropas. Lutra Verlags- und Vertriebsgesellschaft, Tauer
Suarez AV, Bolger DT, Case TJ (1998) Effects of fragmentation and invasion on native ant communities in coastal southern California. Ecology 79:2041–56
Van Vorhis Key SE, Baker TC (1982) Trail-following responses of the Argentine ant, Iridomyrmex humilis (Mayr) to a synthetic trail pheromone component and analogs. J Chem Ecol 8:3–14
von Beeren C, Lizon à l’Allemand S, Hashim R, Witte V (2014) Collective exploitation of a temporally unpredictable food source: mushroom harvesting by the ant Euprenolepis procera. Anim Behav 89:71–7
von Thienen W, Metzler D, Choe D-H, Witte V (2014) Pheromone communication in ants: a detailed analysis of concentration dependent decisions in three species. Behav Ecol Sociobiol 68:1611–27
von Thienen W, Metzler D, Witte V (2015) Modelling shortest path selection of the ant Linepithema humile using psychophysical theory and realistic parameter values. Journal of Theoretical Biology 372:168–178
Wetterer JK, Wild AL, Suarez AV, Roura-Pascual N, Espadaler X (2009) Worldwide spread of the Argentine ant, Linepithema humile (Hymenoptera: Formicidae). Myrmecol News 12:187–94
Wilson EB (1927) Probable inference, the law of succession, and statistical inference. J Am Stat Assoc 22:209–12
Wilson EO (1958) A chemical releaser of alarm and digging behavior in the ant Pogonomyrmex badius (Latreille). Psyche 65:41–51
Wilson EO, Pavan M (1959) Glandular sources and specificity of some chemical releasers of social behavior in dolichoderine ants. Psyche 66:70–6
Witte V, Maschwitz U (2008) Mushroom harvesting ants in the tropical rainforest. Naturwissenschaften 95:1049–54
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical standards and conflict of interest
The authors declare that the experiments comply with the current law of the country in which they had been performed and that they have no conflict of interest.
Additional information
Communicated by M. Giurfa
Significance statement
Collective organization in ants is significant to environmental adaptation. Worker foraging may be guided by private (memory) and social (pheromonal) information, and the relative reliance on these information sources should vary interspecifically. Memory and motivation were found to vary in their influence on response thresholds to trail pheromones and error rates in three ant species. This variation among species underscores specific ecological adaptations.
Appendix
Appendix
Introduction into psychophysical methods used by von Thienen et al. (2014)
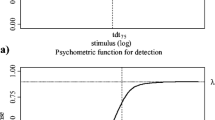
In psychophysics, a psychometric function (PF) describes the relationship between the probability of a positive response p to a stimulus and the stimulus strength x
x—stimulus strength, λ—guess rate, γ—lapse rate, F(x)—function describing the probability to detect a stimulus by the underlying sensory mechanism (Kingdom and Prins 2010 p. 74). For F(x) a probability distribution like the Weibull distribution is applied. For mathematical details, please refer to von Thienen et al. (2014).
von Thienen et al. (2014) used the PF to provide a mathematical description for the response of ants to trail pheromones of varying concentrations. They performed two types of psychophysical experiments: Detection experiments and discrimination experiments. The detection experiments measured the ability to detect a trail pheromone of varying intensity against a null stimulus or noise. Discrimination experiments measured the ants’ ability to discriminate a trail pheromone of varying concentration from a reference trail pheromone of constant concentration. Both types are of specific biological importance for understanding the collective behavior of ants. Detection is the ants’ ability to detect a pheromone trail and become attracted to it. It is important, for instance, to describe how ants at the nest entrance respond to a trail that leads out of the nest to a food source. Discrimination is the ability to differentiate pheromone concentrations at a trail bifurcation. This is especially important in situations where ants have to collectively find the better of two alternate solutions, for instance food of different quality. PFs are S-shaped curves providing a probability of response, theoretically ranging from 0.5 to 1 in detection experiments and from 0 to 1 in discrimination experiments. The stimuli were presented as trail pheromone of varying concentrations on one of the two branches of a y-shaped bridge. Parameters of the PF are the lapse rate (the distance between upper asymptote and 1) and the 75 %-detection/discrimination threshold. von Thienen et al. (2014) interpreted the lapse rate biologically in two ways: as a measure for the errors that ants make and as a measure for the independence from pheromone guided behavior. It might be important for ants not to get stuck in suboptimal solutions and explore alternate solutions. The thresholds are standardized measures of the ants’ ability to detect a stimulus or to discriminate between two stimuli, respectively. von Thienen et al. (2014) had shown that differences between the PFs of the three species (L. humile, E. procera, and L. niger) can be attributed to specific adaptations to their environment. L. humile ants coordinate the work of extremely large and interconnected colonies (Holway and Case 2000; Giraud et al. 2002). They depend on persistent as well as short-lived food supplies (Mallis 1942; Suarez et al. 1998). Thus, they depend highly on collective information and must be able to communicate even the smallest changes in their environment. This reflects in a low lapse rate and a low detection threshold. L. niger, on the other hand, has comparably small colony sizes and uses persistent as well as short-lived food sources (Seifert 2007). Thus, collective information is less important compared to L. humile while individual information plays an important part. This reflects in a higher lapse rate and a high detection threshold. E. procera is positioned between these two extremes. Their colonies are also much smaller than those of L. humile. Furthermore, the ants are nocturnal so that visual cues are rare (Witte and Maschwitz 2008; von Beeren et al. 2014). Consequently, they depend more on collective information and show a high trail fidelity, which reflects in the lowest lapse rate of all three species. Since their food sources are much more stable compared to the other two species, they do not have to share as much information and consequently have a comparatively high detection threshold.
Rights and permissions
About this article

Cite this article
von Thienen, W., Metzler, D. & Witte, V. How memory and motivation modulate the responses to trail pheromones in three ant species. Behav Ecol Sociobiol 70, 393–407 (2016). https://doi.org/10.1007/s00265-016-2059-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00265-016-2059-5