
Abstract
Cytotoxic macrocyclic trichothecenes such as satratoxins are produced by chemotype S strains of Stachybotrys chartarum. Diseases such as stachybotryotoxicosis in animals and the sick building syndrome as a multifactorial disease complex in humans have been associated with this mold and its toxins. Less toxic non-chemotype S strains of S. chartarum are morphologically indistinguishable from chemotype S strains, which results in uncertainties in hazard characterization of isolates. To selectively identify macrocyclic trichothecene producing S. chartarum isolates, a set of sat14 gene-specific primers was designed and applied in a loop-mediated isothermal amplification (LAMP) assay using neutral red for visual signal detection. The assay was highly specific for S. chartarum strains of the macrocyclic trichothecene producing chemotype and showed no cross-reaction with non-macrocyclic trichothecene producing S. chartarum strains or 152 strains of 131 other fungal species. The assay’s detection limit was 0.635 pg/rxn (picogram per reaction) with a reaction time of 60 min. Its high specificity and sensitivity as well as the cost-saving properties make the new assay an interesting and powerful diagnostic tool for easy and rapid testing.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The dematiaceous hyphomycete fungus Stachybotrys (S.) chartarum (Ehrenb.) S. Hughes plays an important role as a potential health hazard for humans [1, 2] and animals [3] due to its production of highly toxic secondary metabolites, known as mycotoxins. Especially, macrocyclic trichothecenes such as satratoxins [4] produced by S. chartarum chemotype S strains have a high cytotoxic potential [5]. In horses and other animals, a disease known as stachybotryotoxicosis was described already in 1945 [6, 7]. The disease’s symptoms are catarrh with hemorrhage and ulceration of the mucosae of mouth, nose, and throat, followed by lymphadenopathy with fever, leucopenia, agranulocytic anemia, and death. They result from the ingestion of moldy fodder that had been contaminated with S. chartarum and its associated mycotoxins. Moreover, also inhalation of S. chartarum-contaminated farm dust and spores were demonstrated to result in clinical signs of stachybotryotoxicosis in animals and farm workers [8]. Skin contact with farm dust and moldy straw bedding resulted in a moist dermatitis with crusts of dried serous exudation in farm animals and workers.
Besides its role in stachybotryotoxicosis, S. chartarum was discussed as being involved in fatal cases of pulmonary hemorrhage in infants, which occurred in the 1990s after extensive flooding in Cleveland, Ohio [9, 10]. The toxic fungus was regularly isolated from water damaged cellulosic materials such as wallpapers, plasterboard, or wood [11,12,13,14]. Other studies confirmed this correlation [15, 16]. Moreover, S. chartarum has been associated with the sick building syndrome [17, 18], a difficult to define medical condition that is caused by multiple factors from the indoor environment of a building. Even though some of the implications that S. chartarum might have on humans are under discussion today [19, 20], its negative influence on animal health and the involvement of macrocyclic trichothecenes in its pathology are beyond doubt. The detection of airborne macrocyclic trichothecenes proved that exposure to these toxins is possible by inhalation [21,22,23]. Furthermore, also foodstuffs such as dried culinary herbs were recently reported to be a possible vector of Stachybotrys chartarum [24].
Since S. chartarum is a hazardous fungus, the distinction of strains that produce rather harmless atranones from strains that produce highly toxic macrocyclic trichothecenes is of high importance for the estimation of the toxicological potential of animal feed and food as well as for the hazard assessment of indoor environments. The distinction of strains according to chemotypes has been achieved in the past by using liquid chromatography with mass spectrometry analysis for identification of compounds (LC-MS/MS) [25, 26] or through cytotoxicity testing of culture extracts using the MTT assay [27, 28]. However, both methods are complex, expensive, and time-consuming because fungal isolates must be cultivated up to 3 weeks [27] for analytes to be produced. Indirect methods such as micro- and macro-morphology of cultures and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) failed to distinguish highly toxic strains from others [29, 30]. Distinguishing chemotypes also failed with polymerase chain reaction (PCR) assays that were based either on sequences of the 18S rRNA gene [31] or the ITS1-5.8S-ITS2 rRNA gene region [32]. With a C to T exchange in nucleotide position 279 of the tri5 gene, Andersen et al. distinguished between genotype S and genotype A in strains of S. chartarum [26, 33]. However, this distinction was only poorly correlated with the two chemotypes of the species. A recent study has found that the two chemotypes known in S. chartarum are represented by three different genotypes instead of two [34]. In contrast to genotypes A and H (both atranone-producing chemotype A), strains of genotype S produce satratoxins F, G, and H and other highly toxic macrocyclic trichothecenes like verrucarin J, roridin E, and L-2 (chemotype S) [35, 36]. It was demonstrated that this genotype exclusively harbors the complete set of 21 genes in the sat-cluster that is necessary for the production of macrocyclic trichothecenes. Chemotype A strains were either devoid of sat-genes (genotype A) or the gene cluster was incomplete (genotype H). In particular, genes sat11 through sat16 were exclusively present in strains that produced macrocyclic trichothecenes in culture [34].
Loop-mediated isothermal amplification (LAMP) is a DNA-based molecular technology that uses a set of four oligonucleotide primers, which need to hybridize to six different locations in the genome of a target organism before enzymatic autocycling in vitro biosynthesis of DNA occurs under isothermal conditions at 65 °C [37, 38]. Advantages of LAMP over PCR-based assays are their higher reaction speed, simplicity of application, and reduced proneness to inhibitors present in sample materials. Its characteristics make LAMP an ideal tool for point of analysis (POA) applications. Due to the application of color change reactions, a visual readout of results can be performed with the naked eye immediately after the reaction has been terminated. Among other organisms, various assays have been designed for the specific detection of filamentous fungi and yeasts [39]. Only recently, the group-specific detection of the fum1-gene in fumonisin-producing Fusarium spp. in maize [40] was successfully demonstrated. Another example for the application of LAMP to the detection of mycotoxin-producing fungi is the detection of patulin producers among Penicillium species and its application to the analysis of grapes and apples [41].
The goals of the current study were to set up and optimize a LAMP-based assay for the selective identification of macrocyclic trichothecene producing (genotype S) strains of S. chartarum and to determine its characteristics (sensitivity, selectivity).
Materials and methods
Chemicals
Tris-HCL and EDTA were purchased from Gerbu Biotechnik GmbH (Heidelberg, Germany). Sodium chloride, sodium acetate, isopropyl alcohol, and dimidium bromide were purchased from Karl Roth GmbH & Co. KG (Karlsruhe, Germany). SDS was purchased from SERVA Electrophoresis GmbH (Heidelberg, Germany). Ethanol analytical grade and acetic acid were purchased from VWR International (Radnor, PA, USA). DNA loading buffer and DNA Ladder GeneRuler 100bp were purchased from Thermo Fisher Scientific Inc. (Waltham, MA, USA). HPLC grade water was used in all experiments unless stated otherwise and was purchased from J.T. Baker (Center Valley, USA).
Fungal cultures and culture conditions
A complete list of 227 fungal isolates used during the current study is given in Table S1 (see Supplementary Information, ESM). Strains of S. chartarum (CBS 414.95, CBS 129.13, and CBS 324.65) were used as reference strains for the genotypes and used for validation and as controls during method development. Cultures were obtained upon direct request to the institutions given as sources. Fungal stock cultures were maintained in glycerol at −80 °C as described by Niessen et al. [42]. Working cultures of all strains were grown on 2.0% malt extract agar plates (MEA, 20 g/L malt extract, 2 g/L soy peptone, and 15 g/L agar (Difco, Heidelberg, Germany), adjust to pH 5.4). Prior to use, all media were sterilized by autoclaving at 121 °C for 15 min. All cultures were grown at ambient temperature (AT, 22 ± 1 °C) for 7 days. Agar cultures were inoculated with a small piece of mycelium from working culture plates and incubated for 5 days at AT. For DNA extraction, cultures were grown in 500 μL of malt extract broth in sterile 1.5 mL reaction vessels at AT on a horizontal shaker at 80 rpm.
LC-MS/MS measurement
Detection of macrocyclic trichothecenes (satratoxin G, H, and F; roridin E and L-2; verrucarin J) produced by the tested isolates in this study was performed in previous studies by our working group. The LC-MS/MS system consisted of a HPLC device (Shimadzu LC-20AB, SIL-20AC HT, CTO-20AC, CBM-20A, Duisburg, Germany) coupled to an API 4000 triple quadrupole mass spectrometer (Sciex, Darmstadt, Germany). In brief, strains were cultivated on MEA agar for 21 days at 25 °C and plates were extracted by treating in a bag mixer with 50 mL acetonitrile/water 84/16 (v/v). An aliquot (5 mL) of the extract was evaporated to dryness and the residues were reconstituted in 1 mL acetonitrile/water 30/70 (v/v). The limits of detection (LOD) were calculated using the signal-to-noise approach and ranged between 0.1 and 7.8 ng/g MEA agar. For detailed information regarding sample preparation and substance-specific and measurement parameters, refer to Ulrich et al. [34].
DNA isolation and amplification
DNA extraction from fungal cultures was performed using the protocol described in [43] with some modifications. We used 1.5-mL Eppendorf tubes as culture vessels, containing 500 μL malt extract broth (see above, no agar added). Tubes were inoculated with a loop-full of fungal conidia and incubated horizontally at AT on a rotary shaker at 150 rpm for 2 days. Following centrifugation at 17,900×g for 5 min and washing with 500 μL of sterile HPLC grade water, mycelial pellets were mixed with sterile glass beads (1.25–1.65 mm diameter, Carl Roth, Karlsruhe, Germany), sea sand (Merck, Darmstadt, Germany), 300 μL extraction buffer (200 mM Tris-HCl (Gerbu Biotechnik, Heidelberg, Germany), 250 mM sodium chloride (Carl Roth, Karlsruhe, Germany), 25 mM EDTA (Gerbu), 0.5% (w/v) SDS (SERVA Electrophoresis, Heidelberg, Germany), and treated in a TissueLyser (45 s; 5.5 m/s, FastPrep®-24, MP Biomedicals Germany, Eschwege, Germany) to release genomic DNA from cells. The remaining steps were performed as described in Cenis et al. [43]. Vacuum-dried genomic DNA was suspended in sterile HPLC grade water. DNA concentrations were monitored using a NanoDrop ND1000 spectrophotometer (Peqlab Biotechnologie, Erlangen, Germany) according to the manufacturer’s recommendations.
PCR amplification of target DNA
Chemotype-specific triplex PCR with DNA of S. chartarum isolates was performed using the Taq Core Kit 10 (MP Biomedicals) with the primers and cycling protocol described in [34]. The PCR products were separated on 1.3% agarose gel (Biozym Scientific, Hessisch Oldendorf, Germany) at 120 V and 200 mAmp for 1 h and subsequently stained in a dimidium bromide bath for 10 min. Gels were subsequently washed with deionized water for 10 min before visual analysis on a UV transilluminator (UVT-28 M, Herolab, Wiesloch, Germany). PCR products were purified directly from amplification reactions using the QIAquick PCR Purification Kit according to the manufacturer’s instructions (QIAGEN, Hilden, Germany). PCR and LAMP products that were excised from agarose gels with a scalpel were purified using the MinElute Gel Extraction Kit (QIAGEN, Hilden, Germany) according to manufacturer’s instructions.
LAMP amplification of target DNA
Primer design was done, using the Primer Explorer V5 online software (Eiken Chemical Co., Ltd., Tokyo, Japan). The master mix for one LAMP reaction contained 2.5 μL 10× ammonium sulfate buffer (100 mM ammonium sulfate (Gerbu), 100 mM potassium chloride (Carl Roth), pH 8.7), 1 μL magnesium chloride (200 mM, Carl Roth), 3.5 μL dNTPs mix (10 mM each GATC, MP Biomedicals GmbH, Eschwege, Germany), 2.6 μL primer mix (1.6 μM each FIP and BIP, 0.8 μM each LF and LB, 0.2 μM each F3 and B3 final concentration in master mix, see Table 1 for primer sequences), 0.75 μL formamide (Sigma-Aldrich, Taufkirchen, Germany), 1 μL Bst polymerase (8 U/μL, New England BioLabs, Frankfurt am Main, Germany), 1 μL neutral red (2.5 mM, SERVA Electrophoresis), 7.65 μL sterile deionized UV-treated water, and 5 μL of template DNA per 25 μL of reaction volume. The assay was incubated at 65.5 °C for 60 min in a Mastercycler Gradient Thermal Cycler (Eppendorf, Hamburg, Germany). Genomic DNA of strain S. chartarum CBS 414.95 was used as positive control and sterile HPLC grade water was used as negative control throughout the study. Assays for validation were performed in triplicates.
A few reactions were performed with the ESEQuant TS real-time fluorimeter (QIAGEN Lake Constance GmbH, Stockach, Germany) to determine the effect of our loop primers on the reaction. V13 (V13-01184, Dyomics GmbH, Jena, Germany) was used as a fluorescent indicator dye instead of neutral red using all other reaction components as previously described.
Assay validation
Cross-reactions in the assay were eliminated by increasing formamide concentration (see Fig. S2 in ESM) and temperature (see Fig. S1 in ESM). The presence of the correct amplified LAMP product in positive LAMP reactions was confirmed by sequence analysis of the smallest DNA fragment visible when LAMP reactions were separated by agarose gel electrophoresis. The smallest amplified product was cut out from the gel and a PCR with the primers F2 and B2 (part of FIP and BIP, see Table 1) was done with the purified DNA fragment as template. Subsequently, the PCR product was isolated as described previously and Sanger sequenced using primers F2 and B2. Sequences were pairwise aligned and a consensus sequence was generated.
The effect of loop primers LF and LB (see Table 1) on the reaction speed is a helpful parameter for the fine-tuning of a LAMP assay. We measured this effect in a real-time ESE Quant TS tube scanner fluorimeter (QIAGEN Lake Constance, Germany) using the V 13-01184 fluorescent dye (Dyomics, Jena, Germany) as indicator.
The sensitivity of the assay was analyzed by testing a serial dilution of S. chartarum CBS 414.95 gDNA as template.
Results
Development and optimization of the LAMP assay
A set of LAMP primers, including loop primers, were designed using the Primer Explorer V5 software. Since S. chartarum genotypes A and H, as well as S. chlorohalonata, lack the presence of the satratoxin core cluster 2 (SC2) [34, 44], we designed our primer set (see Table 1) to hybridize with the sat14 gene exclusively present in SC2 of genotype S strains of S. chartarum (= chemotype S). If a strain belongs to chemotype S, the primers hybridize with the sat14 gene and DNA is amplified during the LAMP reaction. Positive results are visualized by a color change from yellow to pink and can be read with the naked eye due to the utilization of neutral red as pH indicator. Cross-reactions in our LAMP were eliminated step by step by adding formamide to the master mix and increasing the reaction temperature, thus making the reaction more specific. At low temperatures and without the addition of formamide to the reaction, the binding of the primers became unspecific which led to cross-reactions with several of the tested fungal species. Highly specific reactions occurred however, when 0.75 μL of formamide was added per reaction. Optimization experiments using the temperature gradient function of the thermal cycler revealed 65.5 °C with a reaction time of 60 min as optimum reaction conditions for the rapid, sensitive, and specific amplification of target DNA.
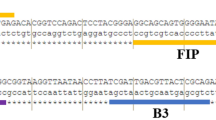
The alignment of the obtained sequence with the sat14 gene sequence is shown in Fig. 1. Results showed a 100% homology between both sequences confirming the correct binding and specificity of the designed primers.

Comparison of the sat14 gene partial sequence with a consensus sequence of the PCR product obtained with primers F2/B2 on the smallest DNA fragment produced during reactions with the sat14 gene specific LAMP assay
Figure 2 shows the influence of both loop primers on the reaction speed. Using purified gDNA of S. chartarum CBS 414.95 as template, the LAMP reaction showed no measurable signal within 90 min runtime when none of the loop primers LF or LB was added. Addition of either of primers LF or LB resulted in a LAMP signal starting at about 50 min runtime. A fluorescent signal that emerged from the background after only 20 min was detected in the LAMP reaction when both primers LF and LB were added to the master mix.

Influence of loop primers on reaction speed of the sat14 gene specific LAMP reaction using V13 as fluorescent indicator. Graph-pair 1: Both LF + LB loop primers added to the reaction. Graph-pair 2: only LB primer added to the reaction. Graph-pair 3: only LF primer added to the reaction. Graph-pair 4: no loop primers added to the reaction
The sensitivity of the optimized assay was analyzed by the addition of purified gDNA from S. chartarum CBS 414.95 as template from a 10-fold serial dilution. Figure 3 shows that the addition of template DNA from a 10−5 dilution still resulted in a visible color change when neutral red was used as an indicator that changes color from yellow to pink when the pH drops during positive LAMP reactions. This dilution was equivalent to a concentration of 6.35 pg template DNA per reaction. S. chartarum has a haploid genome size of 36.5 Mbp. Therefore, the limit of detection of the LAMP assay translates into a genomic copy number of 15.8 cp/reaction. Since the sat14 gene is a single-copy gene, the assay can detect a limit of 15.8 theoretical cells per reaction [44].

Sat14 gene specific LAMP reaction with a serial dilution of gDNA of reference strain CBS 414.95 using neutral red as pH sensitive indicator (positive reaction indicated by color change from yellow to purple). Cap with 10 -1 holds 1.27 ng/μl (equivalent to 6.35 ng per reaction) as initial template DNA concentration, NTC = No template control
During the current study, a total of 227 fungal isolates were analyzed using the newly developed LAMP assay (see Table S1 in ESM). The tested isolates included 75 strains of S. chartarum (see Table 2). Among the tested S. chartarum strains, 30 genotype S, 29 genotype A, and 14 genotype H were identified by triplex PCR according to [34]. Three strains (CBS 414.95, CBS 129.13, and CBS 324.65) were used as reference strains for the genotypes. Two strains (S1344, IBT 8935) could not be assigned to any of the genotypes using triplex PCR but reacted positive in the LAMP assay. LAMP with all S. chartarum genotype S strains resulted in a positive signal, whereas all strains of genotypes A and H showed negative results in the LAMP assay. Also included in the total of the tested strains were 29 strains of other Stachybotrys species including 21 strains of the closely related S. chlorohalonata as well as one strain each of eight other Stachybotrys species and two strains of Memnoniella echinata as a more remotely related species (for all S. chartarum see ESM Table S1). None of the tested strains reacted positive in the LAMP assay. In order to test unrelated species, 121 isolates of fungal species were tested that potentially occur in environments in which S. chartarum can be found (plant material, indoor environments, straw bedding). All isolates tested negative with the gene-specific LAMP assay.
Every isolate of the 30 S. chartarum Chemotype S which all had a positive result in the sat14-specific LAMP reaction showed the production of macrocyclic trichothecenes in culture as measured by LC-MS/MS in previous studies [24, 34, 45, 46]. Table 3 shows the isolates with underlying data for LC-MS/MS compared to LAMP results. Genotypes A and H, as well as closely related Stachybotrys species, neither showed a positive reaction in the new LAMP assay nor did they produce any of the analyzed macrocyclic trichothecenes. As an exception, the isolate S. dichroa ATCC 18913 was reported to produce roridin E [36] and verrucarin J [46], but lacks the ability to produce satratoxins. However, it correctly proved negative in the LAMP assay.
Specificity testing performed during the current study demonstrated clearly that the designed set of LAMP primers selectively bind to the DNA of genotype S strains of S. chartarum. No cross-reactions were detected with non-satratoxin-producing strains (genotypes A and H) or with closely related Stachybotrys species. Also, the various other tested fungal species did not result in positive LAMP reactions. In order to reach these results, the assay conditions needed to be optimized regarding incubation temperature and the addition of formamide to the master mix to reduce cross-reactions. Moreover, adding an additional pair of loop primers resulted in a considerable reduction of the reaction time as compared to the reaction time without loop primers.
Discussion
According to Semeiks et al. [44], 21 genes are necessary in S. chartarum to produce satratoxins. Previous studies showed that only those strains of the fungus are able to produce satratoxins that harbor the full set of genes in their genomes, including genes sat11 through sat16 (the satratoxin core cluster 2 (SC2) genes) [34]. Moreover, it was demonstrated that non-satratoxin-producing strains of S. chartarum could have two different genotypes, which either have none of the sat-genes or miss sat11 through sat16 (the SC2). In some genotype H strains, also some of the genes in SC1 and SC3 seemed to be missing or truncated [34]. In order to differentiate satratoxin-producing strains from non-satratoxin-producing strains of S. chartarum, we based our assay on the detection of sequences from one of the genes in SC2. Comparing sat-gene sequences of two strains of S. chartarum for which annotated genomic sequences are available, we found that the sat14 gene showed the lowest number of mismatches and SNPs. Moreover, the gene had the lowest number of introns among the six genes in SC2. Both features present in high numbers would bear a potential of designing primers that will not properly hybridize to the DNA of all strains of the target species.
As we pointed out earlier, the LAMP reaction is characterized by a high level of sensitivity so that even small amounts of DNA lead to a positive result. On the other hand, too high DNA concentrations can inhibit the amplification and may lead to false negative results. Thus, measuring concentrations and diluting samples correctly are important. Particular care should be taken with false negative samples when testing on site, as no purified DNA sample is tested and therefore a reliable measurement of DNA concentration as well as occurring reaction inhibitors in the sample are problems to be dealt with. With an adequate adjustment of reaction temperature and formamide concentration, false positive reactions were completely excluded and did not occur among the tested isolates in this study.
The isolate S. dichroa ATCC 18913 revealed a negative result in the sat14-specific LAMP reaction. However, it was previously shown that this isolate produced the macrocyclic trichothecenes roridin E [36] and verrucarin J [46], but no satratoxins. Our LAMP reaction amplifies the sat14 gene which the fungus needs to synthesize this specific group of toxins and that in turn corresponds to an absence of sat14. This observation highlights ambiguities in the metabolite profile of Stachybotrys species, which is still part of ongoing research [47, 48].
During the current study, we used neutral red as a pH-sensitive indicator for in-tube indirect visual signal detection under daylight conditions. The advantage of this indicator compared to turbidimetric or fluorescence-based detection of LAMP signals [49] is that results can be read visually without the need for a device such as a turbidimeter, fluorimeter, or even a UV lamp. Moreover, neutral red can be added to the master mix before the reaction starts; it is not toxic for those handling it [50] and does not interfere with the LAMP reaction. This indicator was used successfully in several previous studies [40, 51, 52]. Furthermore, a big advantage is the minimization of possible lab space contamination with product DNA. This is because a color change indicates a positive reaction, and no further handling of the DNA, like gel electrophoresis, is needed. Optimization and specificity testing were performed using genomic DNA after isolation from pure culture mycelia. As was reported in literature, LAMP assays can also be successfully performed using direct addition of fungal spores as template [42, 53,54,55]. However, during the current study, it turned out that direct addition of spore material of S. chartarum genotype S strains into the master mix did not lead to reliable results (results not shown). Mechanical disruption of the spores by vortexing with and without glass beads, as well as thermal treatment, did not result in the release of sufficiently reproducible gDNA amounts that are needed for a reproducible output of the LAMP assay. We assume that factors such as melanin [29] and polysaccharides from the cell wall and proteinases from the cytoplasm might inhibit the polymerase used in the LAMP reaction. Moreover, DNAses might lead to the rapid digestion of the genomic DNA, hampering its detection by the assay. A working on-site application could contain a portable DNA extraction kit and a portable battery-operated heating device like the ESEQuant TS real-time fluorimeter (QIAGEN Lake Constance GmbH, Stockach, Germany), which was used in this study. As already mentioned, experiments with direct measurement of spores and also with a strongly simplified DNA extraction were not successful by our working group, but we suggest experimental approaches using glass beads/sea sand or similar disruption media combined with vortexing and/or thermal disruption to get usable amounts of DNA for LAMP. Thermal treatment was used successfully by Jayanath et al. in detecting hepatitis B virus in human serum by LAMP [56]. However, according to our experience, it takes higher efforts to disrupt the rigid cell walls of fungal spores. Another study showed that human serum has an inhibitory effect on LAMP reactions [57]. Also, this observation coincides with our experience of inhibitory fungal cell components such as DNAses. An approach to exclude such components from extracts could be a syringe filter-based DNA extraction like the one described by Lee et al. [58]. The most simple way of LAMP analysis of fungi is the direct addition of spores or mycelia into the LAMP master mix as demonstrated by several studies [42, 53, 55, 59]. All chemicals that we used in the current LAMP assay can be transported on-site, e.g., in an ice box. For long time storage of LAMP reagents without cooling, freeze drying of the master mix might be an alternative. However, further research is needed here.
Macrocyclic trichothecenes such as satratoxins are hazardous to the health of farm animals and humans living or working in moldy environments. To detect these toxins in feed, food, or environmental samples, contaminated materials can be analyzed by LC-MS/MS, ELISA, and MTT tests [21,22,23, 27, 28, 60]. Such tests are more or less expensive and time-consuming because samples have to be sent to a testing facility with special instrumentation, while LAMP is a format that involves little costs, time, and equipment, and could potentially be applied even in the field [61, 62]. Therefore, the assay developed during the current study may pave the way for the development of rapid test kits for animal feed and bedding materials and for testing of indoor environments in human dwellings and workplaces. For the implication of these tests to field conditions, further research is needed to set up sample preparation protocols for the rapid direct analysis of sample materials.
Data availability
All methods and materials have been described in full. All used cultures are available from public strain collections as indicated in the text or are available from the authors’ in house collection on request.
Code availability
Not applicable.
References
Etzel RA. Stachybotrys. Curr Opin Pediatr. 2003;15(1):103–6.
Johanning E, Yang C. Health problems related to fungal exposure—the example of toxigenic Stachybotrys chartarum. Eastern New York Occupational Health Program—CHP. 1994. pp. 169-82.
Forgacs J, Carll WT, Herring AS, Hinshaw WR. Toxicity of Stachybotrys atra for animals. Trans N Y Acad Sci. 1958;20(8):787–808.
Jarvis BB, Salemme J, Morais A. Stachybotrys toxins. 1. Nat Toxins. 1995;3(1):10–6. https://doi.org/10.1002/nt.2620030104.
Nielsen C, Casteel M, Didier A, Dietrich R, Märtlbauer E. Trichothecene-induced cytotoxicity on human cell lines. Mycotox Res. 2009;25(2):77–84. https://doi.org/10.1007/s12550-009-0011-5.
Drobotko VG. Stachybotryotoxicosis. A new disease of horses and humans. Am Rev Soviet Med. 1945;2:238–42.
Scudamore KA, Livesey CT. Occurrence and significance of mycotoxins in forage crops and silage: a review. J Sci Food Agric. 1998;77((1):1–17. https://doi.org/10.1002/(SICI)1097-0010(199805)77:1<1::AID-JSFA9>3.0.CO;2-4.
Hintikka E-L, Nikulin M. Airborne mycotoxins in agricultural and indoor environments. Indoor Air. 1998;8(S4):66–70. https://doi.org/10.1111/j.1600-0668.1998.tb00011.x.
Etzel R, Montana E, Sorenson WG, Kullman G, Miller JD, Jarvis B, et al. Pulmonary hemosiderosis associated with exposure to Stachybotrys Atra: T 78. Epidemiology. 1996;7(4):S38.
Etzel RA, Montana E, Sorenson WG, Kullman GJ, Allan TM, Dearborn DG. Acute pulmonary hemorrhage in infants associated with exposure to Stachybotrys atra and other fungi. Arch Pediatr Adolesc Med. 1998;152(8):757–62. https://doi.org/10.1001/archpedi.152.8.757.
Gravesen S, Nielsen PA, Iversen R, Nielsen KF. Microfungal contamination of damp buildings - examples of risk constructions and risk materials. Environ Health Perspect. 1999;107:505–8.
Nielsen KF, Thrane U, Larsen TO, Nielsen PA, Gravesen S. Production of mycotoxins on artificially inoculated building materials. Int Biodeterior Biodegrad. 1998;42(1):9–16. https://doi.org/10.1016/S0964-8305(98)00038-9.
Andersson MA, Nikulin M, Koljalg U, Andersson MC, Rainey F, Reijula K, et al. Bacteria, molds, and toxins in water-damaged building materials. Appl Environ Microbiol. 1997;63(2):387–93. https://doi.org/10.1128/AEM.63.2.387-393.1997.
Andersen B, Frisvad JC, Sondergaard I, Rasmussen IS, Larsen LS. Associations between fungal species and water-damaged building materials. Appl Environ Microbiol. 2011;77(12):4180–8. https://doi.org/10.1128/AEM.02513-10.
Dearborn DG, Smith PG, Dahms BB, Allan TM, Sorenson WG, Montana E, et al. Clinical profile of 30 infants with acute pulmonary hemorrhage in Cleveland. Pediatrics. 2002;110(3):627–37. https://doi.org/10.1542/peds.110.3.627.
Dearborn DG, Yike I, Sorenson WG, Miller MJ, Etzel RA. Overview of investigations into pulmonary hemorrhage among infants in Cleveland, Ohio. Environ Health Pers. 1999;107:495–9. https://doi.org/10.1289/ehp.99107s3495.
Johanning E, Biagini R, Hull D, Morey P, Jarvis B, Landsbergis P. Health and immunology study following exposure to toxigenic fungi (Stachybotrys chartarum) in a water-damaged office environment. Int Arch Occup Environ Health. 1996;68(4):207–18. https://doi.org/10.1007/BF00381430.
Cooley JD, Wong WC, Jumper CA, Straus DC. Correlation between the prevalence of certain fungi and sick building syndrome. Occup Environ Med. 1998;55(9):579–84. https://doi.org/10.1136/oem.55.9.579.
(CDC) CfDCaP. Update: Pulmonary hemorrhage/hemosiderosis among infants--Cleveland, Ohio, 1993-1996. Morbid Mortal Week Rep. 2000;49(9):180–4.
Terr AI. Sick building syndrome: is mould the cause? Med Mycol. 2009;47(Supplement_1):S217–S22. https://doi.org/10.1080/13693780802510216.
Gottschalk C, Bauer J, Meyer K. Detection of satratoxin g and h in indoor air from a water-damaged building. Mycopathologia. 2008;166(2):103–7. https://doi.org/10.1007/s11046-008-9126-z.
Bloom E, Bal K, Nyman E, Must A, Larsson L. Mass spectrometry-based strategy for direct detection and quantification of some mycotoxins produced by Stachybotrys and Aspergillus spp. in indoor environments. Appl Environ Microbiol. 2007;73(13):4211–7. https://doi.org/10.1128/AEM.00343-07.
Brasel TL, Martin JM, Carriker CG, Wilson SC, Straus DC. Detection of airborne Stachybotrys chartarum macrocyclic trichothecene mycotoxins in the indoor environment. Appl Environ Microbiol. 2005;71(11):7376–88. https://doi.org/10.1128/AEM.71.11.7376-7388.2005.
Biermaier B, Gottschalk C, Schwaiger K, Gareis M. Occurrence of Stachybotrys chartarum chemotype S in dried culinary herbs. Mycotoxin Res. 2015;31(1):23–32. https://doi.org/10.1007/s12550-014-0213-3.
Gottschalk C, Bauer J, Meyer K. Determination of macrocyclic trichothecenes in mouldy indoor materials by LC-MS/MS. Mycotoxin Res. 2006;22(3):189–92. https://doi.org/10.1007/BF02959275.
Andersen B, Nielsen KF, Thrane U, Szaro T, Taylor JW, Jarvis BB. Molecular and phenotypic descriptions of Stachybotrys chlorohalonata sp. nov. and two chemotypes of Stachybotrys chartarum found in water-damaged buildings. Mycologia. 2003;95(6):1227–58. https://doi.org/10.1080/15572536.2004.11833031.
Gareis M, Gottschalk C. Stachybotrys spp. and the guttation phenomenon. Mycotoxin Res. 2014;30(3):151–9. https://doi.org/10.1007/s12550-014-0193-3.
Gareis M. Diagnostischer Zellkulturtest (MTT-Test) für den Nachweis von zytotoxischen Kontaminanten und Rückständen. J Verbr Lebensm. 2006;1(4):354–63. https://doi.org/10.1007/s00003-006-0058-6.
Ulrich S, Biermaier B, Bader O, Wolf G, Straubinger RK, Didier A, et al. Identification of Stachybotrys spp. by MALDI-TOF mass spectrometry. Anal Bioanal Chem. 2016;408(27):7565–81. https://doi.org/10.1007/s00216-016-9800-9.
Gruenwald M, Rabenstein A, Remesch M, Kuever J. MALDI-TOF mass spectrometry fingerprinting: a diagnostic tool to differentiate dematiaceous fungi Stachybotrys chartarum and Stachybotrys chlorohalonata. J Microbiol Methods. 2015;115:83–8. https://doi.org/10.1016/j.mimet.2015.05.025.
Cruz-Perez P, Buttner MP, Stetzenbach LD. Specific detection of Stachybotrys chartarum in pure culture using quantitative polymerase chain reaction. Mol Cell Probes. 2001;15(3):129–38. https://doi.org/10.1006/mcpr.2001.0347.
Haugland RA, Vesper SJ, Wymer LJ. Quantitative measurement of Stachybotrys chartarum conidia using real time detection of PCR products with the TaqMan(TM)fluorogenic probe system. Mol Cell Probes. 1999;13(5):329–40. https://doi.org/10.1006/mcpr.1999.0258.
Cruse M, Telerant R, Gallagher T, Lee T, Taylor JW. Cryptic species in Stachybotrys chartarum. Mycologia. 2002;94(5):814–22. https://doi.org/10.2307/3761696.
Ulrich S, Niessen L, Ekruth J, Schäfer C, Kaltner F, Gottschalk C. Truncated satratoxin gene clusters in selected isolates of the atranone chemotype of Stachybotrys chartarum (Ehrenb.) S. Hughes. Mycotoxin Res. 2019. https://doi.org/10.1007/s12550-019-00371-x.
Jarvis BB, Eppley RM, Mazzola EP. Chemistry of trichothecenes - chemistry and bioproduction of macrocyclic trichothecenes. In: Ueno Y, editor. Trichothecenes - chemical, biological and toxicological aspects. Amsterdam: Elsevier Science Publishers B.V; 1983. p. 20–38.
Andersen B, Nielsen KF, Jarvis BB. Characterization of Stachybotrys from water-damaged buildings based on morphology, growth, and metabolite production. Mycologia. 2002;94(3):392–403. https://doi.org/10.1080/15572536.2003.11833204.
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino N, et al. Loop-mediated isothermal amplification of DNA. Nucl Acids Res. 2000;28(12):e63-e. https://doi.org/10.1093/nar/28.12.e63.
Notomi T, Mori Y, Tomita N, Kanda H. Loop-mediated isothermal amplification (LAMP): principle, features, and future prospects. J Microbiol. 2015;53(1):1–5. https://doi.org/10.1007/s12275-015-4656-9.
Niessen L. Current state and future perspectives of loop-mediated isothermal amplification (LAMP)-based diagnosis of filamentous fungi and yeasts. Appl Microbiol Biotechnol. 2015;99(2):553–74. https://doi.org/10.1007/s00253-014-6196-3.
Wigmann ÉF, Meyer K, Cendoya E, Maul R, Vogel RF, Niessen L. A loop-mediated isothermal amplification (LAMP) based assay for the rapid and sensitive group-specific detection of fumonisin producing Fusarium spp. Int J Food Microbiol. 2020;325:108627. https://doi.org/10.1016/j.ijfoodmicro.2020.108627.
Frisch LM, Niessen L. Development and optimization of a group-specific loop-mediated isothermal amplification (LAMP) assay for the detection of patulin-producing Penicillium species. Int J Food Microbiol. 2019;298:20–30. https://doi.org/10.1016/j.ijfoodmicro.2019.03.010.
Niessen L, Vogel RF. Detection of Fusarium graminearum DNA using a loop-mediated isothermal amplification (LAMP) assay. Int J Food Microbiol. 2010;140(2):183–91. https://doi.org/10.1016/j.ijfoodmicro.2010.03.036.
Cenis JL. Rapid extraction of fungal DNA for PCR amplification. Nucl Acids Res. 1992;20(9):2380. https://doi.org/10.1093/nar/20.9.2380.
Semeiks J, Borek D, Otwinowski Z, Grishin NV. Comparative genome sequencing reveals chemotype-specific gene clusters in the toxigenic black mold Stachybotrys. BMC Genomics. 2014;15(1):590. https://doi.org/10.1186/1471-2164-15-590.
Ekruth J, Gottschalk C, Ulrich S, Gareis M, Schwaiger K. Differentiation of S. chartarum (Ehrenb.) S. Hughes chemotypes A and S via FT-IR spectroscopy. Mycopathologia. 2020;185(6):993–1004. https://doi.org/10.1007/s11046-020-00495-0.
Ulrich S-U. Charakterisierung ausgewählter Isolate von Stachybotrys spp. anhand kulturmorphologischer, molekularbiologischer und massenspektrometischer Verfahren: lmu; 2016.
Ding Z-G, Ding J-H, Zhao J-Y, Chunyu W-X, Li M-G, Gu S-J, et al. A new phenylspirodrimane dimer from the fungus Stachybotrys chartarum. Fitoterapia. 2018;125:94–7. https://doi.org/10.1016/j.fitote.2017.12.022.
Jagels A, Lindemann V, Ulrich S, Gottschalk C, Cramer B, Hübner F, et al. Exploring secondary metabolite profiles of Stachybotrys spp. by LC-MS/MS. Toxins. 2019;11(3):133. https://doi.org/10.3390/toxins11030133.
Denschlag C, Vogel RF, Niessen L. Hyd5 gene based analysis of cereals and malt for gushing-inducing Fusarium spp. by real-time LAMP using fluorescence and turbidity measurements. Int J Food Microbiol. 2013;162(3):245–51. https://doi.org/10.1016/j.ijfoodmicro.2013.01.019.
Borenfreund E, Puerner JA. (1985). Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol Lett. 1985;24(2-3):119–24.
Tanner NA, Zhang Y Jr. TCE. Visual detection of isothermal nucleic acid amplification using pH-sensitive dyes. Biotechniques. 2015;58(2):59–68. https://doi.org/10.2144/000114253.
Niessen L, Bechtner J, Fodil S, Taniwaki MH, Vogel RF. LAMP-based group specific detection of aflatoxin producers within Aspergillus section Flavi in food raw materials, spices, and dried fruit using neutral red for visible-light signal detection. Int J Food Microbiol. 2018;266:241–50. https://doi.org/10.1016/j.ijfoodmicro.2017.12.013.
Luo J, Taniwaki MH, Iamanaka BT, Vogel RF, Niessen L. Application of loop-mediated isothermal amplification assays for direct identification of pure cultures of Aspergillus flavus, A. nomius, and A. caelatus and for their rapid detection in shelled Brazil nuts. Int J Food Microbiol. 2014;172:5–12. https://doi.org/10.1016/j.ijfoodmicro.2013.12.001.
Niessen L, Luo J, Denschlag C, Vogel RF. The application of loop-mediated isothermal amplification (LAMP) in food testing for bacterial pathogens and fungal contaminants. Food Microbiol. 2013;36(2):191–206. https://doi.org/10.1016/j.fm.2013.04.017.
Denschlag C, Vogel RF, Niessen L. Hyd5 gene-based detection of the major gushing-inducing Fusarium spp. in a loop-mediated isothermal amplification (LAMP) assay. Int J Food Microbiol. 2012;156(3):189–96. https://doi.org/10.1016/j.ijfoodmicro.2012.03.009.
Jayanath NY, Nguyen LT, Vu TT, Tran LD. Development of a portable electrochemical loop mediated isothermal amplification (LAMP) device for detection of hepatitis B virus. RSC Adv. 2018;8(61):34954–9. https://doi.org/10.1039/C8RA07235C.
Ihira M, Akimoto S, Miyake F, Fujita A, Sugata K, Suga S, et al. Direct detection of human herpesvirus 6 DNA in serum by the loop-mediated isothermal amplification method. J Clin Virol. 2007;39(1):22–6. https://doi.org/10.1016/j.jcv.2007.02.001.
Lee S, Khoo VSL, Medriano CAD, Lee T, Park S-Y, Bae S. Rapid and in-situ detection of fecal indicator bacteria in water using simple DNA extraction and portable loop-mediated isothermal amplification (LAMP) PCR methods. Water Res. 2019;160:371–9. https://doi.org/10.1016/j.watres.2019.05.049.
Niessen L, Gräfenhan T, Vogel RF. ATP citrate lyase 1 (acl1) gene-based loop-mediated amplification assay for the detection of the Fusarium tricinctum species complex in pure cultures and in cereal samples. Int J Food Microbiol. 2012;158(3):171–85. https://doi.org/10.1016/j.ijfoodmicro.2012.06.021.
Ulrich S, Schäfer C. Toxin production by Stachybotrys chartarum genotype S on different culture media. J Fungi. 2020;6(3):159. https://doi.org/10.3390/jof6030159.
Fakruddin M. Loop mediated isothermal amplification (LAMP)–an alternative to polymerase chain reaction (PCR). Bangladesh Res Pub J. 2011;5(4):425–439. http://www.bdresearchpublications.com/admin/journal/upload/09235/09235.pdf.
Parida M. Rapid and real-time detection technologies for emerging viruses of biomedical importance. J Biosci. 2008;33(4):617–28.
Acknowledgements
Thanks to Andrea Pape, Barbara Fritz, Verena Hohenester, and Erika Altgenug for their technical support. Thanks to Prof. Dr. Rudi F. Vogel for the friendly admission at his institute in Freising-Weihenstephan.
Funding
Open Access funding enabled and organized by Projekt DEAL. This research received funding from Brigitte and Wolfram Gedek Foundation, Ismaning, Germany.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by Johannes Köck and Ludwig Niessen. The first draft of the manuscript was written by Johannes Köck and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript. Conceptualization: Johannes Köck; project administration: Ludwig Niessen; writing—original draft: Johannes Köck; writing—review and editing; Christoph Gottschalk; validation: Karin Schwaiger and Christoph Gottschalk; methodology: Sebastian Ulrich; supervision: Manfred Gareis.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(PDF 485 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Köck, J., Gottschalk, C., Ulrich, S. et al. Rapid and selective detection of macrocyclic trichothecene producing Stachybotrys chartarum strains by loop-mediated isothermal amplification (LAMP). Anal Bioanal Chem 413, 4801–4813 (2021). https://doi.org/10.1007/s00216-021-03436-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00216-021-03436-y