
Abstract
Key message
A unique, global onion diversity set was assembled, genotyped and phenotyped for beneficial traits. Accessions with strong basal rot resistance and increased seedling vigour were identified along with associated markers.
Abstract
Conserving biodiversity is critical for safeguarding future crop production. Onion (Allium cepa L.) is a globally important crop with a very large (16 Gb per 1C) genome which has not been sequenced. While onions are self-fertile, they suffer from severe inbreeding depression and as such are highly heterozygous as a result of out-crossing. Bulb formation is driven by daylength, and accessions are adapted to the local photoperiod. Onion seed is often directly sown in the field, and hence seedling establishment is a critical trait for production. Furthermore, onion yield losses regularly occur worldwide due to Fusarium basal rot caused by Fusarium oxysporum f. sp. cepae. A globally relevant onion diversity set, consisting of 10 half-sib families for each of 95 accessions, was assembled and genotyping carried out using 892 SNP markers. A moderate level of heterozygosity (30–35%) was observed, reflecting the outbreeding nature of the crop. Using inferred phylogenies, population structure and principal component analyses, most accessions grouped according to local daylength. A high level of intra-accession diversity was observed, but this was less than inter-accession diversity. Accessions with strong basal rot resistance and increased seedling vigour were identified along with associated markers, confirming the utility of the diversity set for discovering beneficial traits. The onion diversity set and associated trait data therefore provide a valuable resource for future germplasm selection and onion breeding.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Onion (Allium cepa L.) is a major horticultural crop belonging to the order Asparagales which, of the monocot orders, is second only to the Poales for global economic value (Kuhl et al. 2004). Worldwide, 93 Mt of bulb onions are produced annually (FAOSTAT 2016) and onion consumption is associated with potential health benefits, mainly linked to alk(en)yl cysteine sulphoxides and flavonoid compounds (Griffiths et al. 2002). Although onions are self-fertile, they are considered outbreeding crops as they exhibit severe inbreeding depression (Jones and Davis 1944; Brewster 2008). In one study, 75% of seed derived from cross-pollination produced bulbs in a field trial, whereas less than 50% of seed from self-pollination produced bulbs (Currah and Ockendon 1983). This means that most onion cultivars are heterozygous with each plant being genetically unique. Onion is a diploid species (2n = 2x = 16) but possesses one of the largest genomes among eukaryotes (16.3 Gb per 1C nucleus; Arumuganathan and Earle 1991). Together, the large genome and outbreeding nature of onion have significantly hampered the development of advanced genomic resources compared to most other major crops. Although there is no publicly available nuclear DNA sequence, both the mitochondrial and chloroplast genomes have been sequenced (Shigyo et al. 2018). Molecular markers for genotyping have been developed through restriction fragment length polymorphisms (RFLPs), amplified fragment length polymorphisms (AFLPs), simple sequence repeats (SSRs) and single nucleotide polymorphisms (SNPs) (King et al. 1998; van Heusden et al. 2000; Martin et al. 2005; Baldwin et al. 2012a, b; Jo et al. 2017). More recently, Duangjit et al. (2013) reported the development of 930 validated Kompetitive Allele-Specific PCR (KASP™; developed by LGC Ltd.) SNP assays from transcriptome sequencing, together with a reference genetic map.
Daylength sensitivity is a key trait for onion production. In the UK and northern Europe, long-day (LD) cultivars requiring > 16 h day light for bulb initiation are grown, while in other parts of the world, intermediate day (ID; requiring 13–14 h daylength) or short-day (SD; requiring 12 h daylength) cultivars are grown (Brewster 2008). Therefore, breeding programmes are generally confined to germplasm adapted to each specific daylength, generating a major breeding bottleneck for the exchange of alleles between the different daylength-adapted genepools. There is also considerable variability between onion cultivars for important traits such as bulb colour/size, time to maturity and pungency suggesting that genetic diversity within A. cepa exists (Griffiths et al. 2002). Given the increasing speed of genetic erosion in A. cepa and related species (Keller et al. 2013), genebanks play a crucial role in the conservation of genetic diversity so that it can be further exploited to discover alleles for beneficial traits. A good example of the value of a closely related species is the introgression of downy mildew resistance into A. cepa from A. roylei (Scholten et al. 2007). However, the difficulty in introgressing this resistance without other associated unfavourable alleles meant that it took approximately 20 years to breed this trait into a commercial onion cultivar (Scholten et al. 2007). Therefore, to shorten the breeding process, it is more efficient to identify beneficial alleles within A. cepa germplasm.
For much of onion production, seed is directly sown in the field and hence seedling establishment is an important trait for a uniform, high yielding crop (Finch-Savage et al. 2010). Seedling emergence has a direct effect on crop yield and cannot be compensated for with subsequent inputs (Bleasdale 1967), while predictable and even emergence is crucial for crop uniformity and for effective timing of herbicide applications, required to maximise yield. Seedling vigour is an indicator of how well a seed lot will establish seedlings (Finch-Savage et al. 2010; Finch-Savage and Bassel 2016) and is determined by the interaction between genetics and environment (Whittington 1973; Hodgkin and Hegarty 1978). It is important to consider both germination and post-germination seedling growth. Three main traits have been established for measuring vigour: germination speed, speed of initial downward root growth and upward shoot growth in compacted soil (Finch-Savage et al. 2010). However, the standard test used to measure seed quality in the onion industry is a simple filter paper germination test (ISTA 2013). While this can be used to measure viability, it gives little information on vigour. Effective tests have been developed to measure all three of the main seedling vigour traits in Brassica oleracea under controlled conditions (Bettey et al. 2000; Finch-Savage et al. 2010).
One of the major constraints to onion production worldwide is Fusarium basal rot (FBR) caused by the soilborne fungus Fusarium oxysporum f. sp. cepae (FOC). The pathogen can cause symptoms at all stages of plant development leading to both pre- and post-harvest losses (Entwistle 1990; Taylor et al. 2013). FOC is a major problem for onion growers in the UK, and losses are predicted to increase further under current climate change models as infection is favoured by warmer temperatures (optimum 30 °C; Cramer 2000). As FOC produces chlamydospores that can survive in the soil for many years, disease management is challenging and has previously relied on soil sterilisation or fungicides (Brayford 1996; Cramer 2000). These approaches have largely been unsuccessful, have undesirable environmental effects and are threatened restrictions on pesticide use. The development of resistant cultivars is therefore a desirable option. Onion seedling assays have been employed for rapid initial screening for FBR resistance (Retig et al. 1970; Abawi and Lorbeer 1971b; Holz and Knox-Davies 1974; Krueger et al. 1989; Galván et al. 2008; Saxena and Cramer 2009; Taylor et al. 2013) and using this approach together with a subsequent mature plant assay, partial resistance has been identified (Taylor et al. 2013). This supports previous reports where only partial resistance to FBR has ever been identified in onion (Cramer 2000; Saxena and Cramer 2009; Gei et al. 2014). High-level resistance has been reported in closely related Allium species such as A. roylei, A. fistulosum and A. galanthum (Galván et al. 2008; Rout et al. 2016), but these species do not produce a bulb and would require a long breeding programme to achieve a commercial FBR resistant onion cultivar. FOC isolates can also vary in their aggressiveness (Özer et al. 2004; Galván et al. 2008; Taylor et al. 2013), although there does not always appear to be a FOC isolate × cultivar interaction (Taylor et al. 2013). The best strategy is therefore to identify and utilise a highly aggressive FOC isolate for FBR resistance screening.
Here we report the development of a unique onion diversity set consisting of up to 10 half-sib families (Cramer 2006) for each of 95 accessions to enable pre-breeding (trait screening) research. Genotyping using 892 SNP markers was utilised to determine the underlying population structure and relatedness. The utility of the diversity set was also assessed by screening one half-sib family per accession for response to FOC and improved seedling vigour leading to the identification of accessions with high levels of basal rot resistance and increased vigour. These phenotype data were then used to explore marker-trait associations.
Materials and methods
Development of an onion diversity set
Ninety-six onion accessions were selected from the UK Vegetable Genebank, University of Warwick (Table 1) considering geographic origin (including LD, ID and SD types) and including a mixture of advanced cultivars, traditional cultivars and landraces. A. cepa var. ascalonicum (shallot), A. fistulosum (bunching onion) A. cepa × A. fistulosum cross and the two wild species A. roylei and A. vavilovii were also included. Seed was sown in beds in Wellesbourne, UK in April 2006. Bulbs were harvested at maturity, dried and in late 2006–early 2007, ten healthy bulbs per accession planted in pots in separate insect proof cages in a glasshouse under natural conditions. Upon flowering, blow flies were used to randomly cross-pollinate or self-pollinate the ten plants and seed was collected from each plant individually. The seed from each plant therefore represents a half-sib (HS) family for which the female parent is known, but the male parent could be any one of the ten plants from the same accession. Seed was stored under Genebank long-term storage conditions (− 20 C, 5% moisture content; FAO 2013) in hermetically sealed foil-laminate pouches.
Onion SNP genotyping and analysis
To examine genetic diversity across founder onion accessions and between and within half-sib families, genotyping was carried out using DNA extracted from leaf tissue of (1) one of the 10 HS mother plants from 91 founder onion accessions and three cultivars used as resistant (SE, Saxena and Cramer 2009; ACP, Taylor et al. 2013), or susceptible (HY, Taylor et al. 2013) controls in FOC resistance screens, (2) the nine remaining HS mother plants for four selected onion accessions (CA, DM, GR and HO) and (3) ten individual plants from the same single HS family (same mother plant) for each of the four selected accessions in (2). Leaf tissue was flash frozen in liquid nitrogen, lyophilised and 20 mg disrupted in a lysing matrix A tube (MPBio) by a FastPrep-24™ machine (MPBio) set at 6 ms−1 for 40 s. DNA was extracted using a DNeasy plant mini kit (Qiagen), quality checked using a Nanodrop spectrophotometer (Thermo Scientific) and genotyping carried out by LGC Ltd., UK using 892 published KASP™ SNP markers (Duangjit et al. 2013).
Markers were removed from further analysis if they were monomorphic, had a minor allele frequency below 5%, had more than five missing data points or repeatedly gave illegitimate genotypes in the male parent analysis described below. The heterozygosity of each of the 94 HS mother plants was determined using the remaining 765 markers. To examine the genotypic diversity across the 94 representative mother plants from each of the founder accessions, genotype scores were converted to standard IUPAC single letter nucleotide ambiguity codes and used to construct a neighbour-joining (NJ) tree using MEGA 7 (Kumar et al. 2016) with 1000 bootstrap replications. Default parameters were used for non-protein coding sequence.
To examine the genotypic diversity both within and between accessions, the data from (2) and (3) described above were again converted to single letter codes and an NJ tree constructed as described. For the four selected half-sib families, the male parent of each of the 10 half-sib individuals was identified by testing whether the combination of the female genotype and the candidate male genotype was compatible with the genotype of each progeny HS individual for every marker. For the deduced male parent, SNP markers were deemed to give illegitimate genotypes if the HS genotype could not be derived from the two parent genotypes.
Population structure
Estimates of the underlying population sub-structure were calculated for the 765 marker loci across the 94 representative mother plants from each of the founder accessions using STRUCTURE v2.3.4 (Pritchard et al. 2000). The analyses were performed using a burn-in period of 500,000 Markov Chain Monte Carlo iterations, with a 500,000 run-length using an admixture model and correlated allele frequencies for K sub-populations between 1–6, and 4 independent replications. The output was summarised using the Python script structureharvester.py v0.6.92 (Earl and von Holdt 2012), and the most probable underlying value for K was estimated from the ΔK values using the Evanno method (Evanno et al. 2005). The.*indfile for K = 4 was used as input for CLUMPP_OSX.1.1.2 (Jakobsson and Rosenberg 2007) using the Fullsearch algorithm, with weighted H and the G similarity statistic. Summarised cluster membership matrices (q values) were generated for both individuals and populations. The KASP SNP data were also analysed using a principal component analysis (PCA). Eigen values were estimated using the singular value decomposition method implemented in the R package (R Core Team 2011) FactoMineR v1.31.4 (Lê et al. 2008). Individuals within the PCA plots were identified based on the proportion of q-value cluster membership in the STRUCTURE analyses, where membership is assigned when q ≤ 0.25.
Screening the onion diversity set for FBR resistance
One HS family from each of 83 onion diversity set accessions (Table 1) was screened for FBR resistance using a seedling assay (Taylor et al. 2013). Briefly, this involved soaking onion seeds in a spore suspension (1 × 106 spores mL−1) produced from the highly aggressive FOC isolate FUS2 (Lincolnshire, UK) and sowing in modular trays. The highly susceptible cultivar Hystar F1 (HY), as well as the partially resistant Ailsa Craig Prizewinner (ACP), was included in the test (Taylor et al. 2013). Non-inoculated controls (seed soaked in water only) were included for each HS. In total, there were four independent replicates of 28 seeds per HS and trays were positioned in a randomised block design in a glasshouse maintained at 25 °C day and 18 °C night with a 16 h photoperiod. Percentage onion seedling survival four weeks post-inoculation was calculated relative to the non-inoculated control treatment for each HS. Significant differences in seedling survival between the HS families were determined using a residual (or restricted) maximum likelihood (REML) analysis following angular transformation of the data (Welham and Thompson 1997) using GENSTAT v12 (VSN International).
A subset of resistant and susceptible HS families was further tested using a mature onion plant assay (Taylor et al. 2013). Briefly, four-week-old onion seedlings were transplanted into 7-cm square pots containing compost infested with FOC isolate FUS2 (1 × 105 cfu g−1). Control seedlings were transplanted into compost only. Pots were arranged in a randomised block design in a glasshouse (25 °C day, 18 °C night) with three replicate blocks of 20 plants per HS family. Plants were grown to the mature bulb stage (nine weeks after transplanting), left for two weeks without watering, then weighed and scored for severity of FBR symptoms (Taylor et al. 2013). As the HS families had different daylength responses (Brewster 2008; Taylor et al. 2010), two independent assays were carried out. Firstly, a short-day (12 h photoperiod) assay was set up using two resistant (HO, PO) and two susceptible (EX, NU) HS families along with a partially resistant cultivar (cv. Serrana, Saxena and Cramer 2009). Secondly, a long-day (16 h photoperiod) assay was set up using nine resistant (HYS, CA, DM, GR, ACP, MA, SY, PL, JC) and three susceptible (GRE, HY, ST) HS families. Significant differences between symptom scores were determined using REML and results of seedling and mature plant assays compared using Pearson correlation coefficients (Genstat).
Screening the onion diversity set for improved seedling vigour
Seedling vigour for 89 HS families was assessed using two different assays, following published protocols with some modifications. The first assay assessed onion germination and initial seedling growth (Finch-Savage et al. 2010). Seed was surface sterilised by immersing in sodium dichlorocyanuric acid (6.2 g, Sigma, UK) in distilled water (100 ml) with two drops of Nonidet P-40 (Sigma, UK) for 10 min on an orbital shaker (200 rpm) followed by three rinses in sterile distilled water. Seeds were then placed on moist sloping filter papers (190 mm wide, Gray and Steckel, 1983) in trays of water and supported at an angle of 30° to the vertical and extending to 100 mm above the water level. Ten seeds from each HS family were placed 55 mm above the surface of the water and slopes arranged in a randomised block design at 15 °C in the dark and observed twice daily for 21 days. Germination and the time for roots to reach 3 cm and shoots to reach 2 cm were recorded. Four replicate experiments were carried out, each with 10 seeds per HS family. The time taken for 50% of viable seeds to complete germination (T50) was calculated. Time to germination for each seedling was subtracted from time for roots to reach 3 cm and shoots to reach 2 cm so that growth times were not confounded by differences in germination times. All data were analysed using REML (Genstat).
The second assay assessed upward growth in strong soil (Finch-Savage et al. 2010). Soil (sandy loam) was dried (80 °C for 3 days) and sieved (2 mm mesh) before adding 60 g of distilled water per kg of soil. Soil was mixed, sealed in bags incubated at room temperature for 2 days. Onion seeds were sterilised as described and then placed on damp chromatography paper in clear plastic germination boxes. Boxes were incubated at 15 °C until a 2 mm radicle was visible (3–8 days depending on the HS). Three layers of soil were placed in 8 × 8 cm pots (250 ml, 150 ml, 150 ml) with each layer compacted using a 7.3 kg weight before adding the next in order to achieve a uniform bulk density. Pre-germinated onion seeds from each HS family (10 per pot) were then placed on the soil surface after which a final layer of soil (150 ml) was added and either compacted or left uncompacted. Pots were sealed in bags in the dark and placed in a randomised block design at 15 °C. Seedling emergence was recorded at 7, 11, 15, 20 and 25 days, and the experiment replicated three times. Angular transformed data for percentage seedling emergence (relative to treatments with a non-compacted top layer) were analysed using REML. The weight of 100 seeds from each HS family was also recorded. Pearson correlation coefficients (Genstat) were calculated to assess whether any of the seedling vigour traits were correlated.
Association analysis
A preliminary association analysis was carried out to explore if the phenotypic variation for FOC resistance and seedling vigour could be associated with any of the 765 KASP™ markers. Marker data were converted to A (X), B (Y) or H (heterozygous) and markers sorted into linkage map order (Duangjit et al. 2013) with unmapped markers assigned to a pseudo-group. Kruskal–Wallis rank sum tests were carried out using the mean phenotype data to produce the test statistic (K) with an associated significance value using MapQTL® version 6 (Van Ooijen 2009). Markers with a P value ≤ 0.005 were considered as being putatively associated with the trait of interest.
Results
Development of an onion diversity set
An onion diversity set, consisting of up to 10 half-sib families derived from each of 95 founder genebank accessions, was successfully produced (Table 1), and samples can be sourced from the University of Warwick (https://warwick.ac.uk/fac/sci/lifesci/research/vegin/onion/diversity/).
SNP genotyping and analysis
Of the 892 SNPs (Table S1) used to genotype mother plants of the HS families from 91 accessions and HY, SE and ACP, 33 were monomorphic, 53 had > 5 missing data points, and 15 had a minor allele frequency < 5%. A further 26 markers had more than one illegitimate genotype in the kinship test described below. The remaining 765 polymorphic markers were used for subsequent genetic analysis.
The majority of the onion accessions were moderately heterozygous, with a modal value of 30–35% (Table 1, Fig. 1), consistent with onion being an outbreeding species. The least heterozygous accessions were A. roylei (WSR) and A. fistulosum (HLW) with 0.31% and 0.55% heterozygosity, respectively. However, these had 128 and 215 missing SNPs, respectively, suggesting that the KASP assays did not detect these loci. Three other accessions had high numbers of missing SNPs, GU (A. cepa × A. fistulosum), HY and SE with 122, 205, and 219, respectively. By comparison, the mean number of missing SNPs for the remaining onion accessions was only 11. The HS family for GU (A. cepa × A. fistulosum) was moderately heterozygous (33.3%) likely due to SNPs from homeologs in the amphidiploid, while the most heterozygous A. cepa HS was WE (41.6%). As expected, the commercial hybrid cultivar Hystar (HY) was highly heterozygous (47.3%). The least heterozygous A. cepa HS was BE (4.9%). Overall, F1 hybrid accessions ranged in heterozygosity from 30–40% and were no more heterozygous than many of the open pollinated lines (5–42%, mean 31%). Heterozygous alleles were not restricted to certain SNP markers or regions of the genome, as all but one marker detected heterozygous alleles and 310 markers possessed 30–40% heterozygous alleles. Percentage heterozygosity was not correlated with any of the phenotyping traits (data not shown) and accessions with low heterozygosity still resulted in HS families with good seedling vigour. For instance, accession DM (5.8% heterozygosity) performed well for all vigour measurements.

Frequency distribution of recorded heterozygous alleles for 94 accessions in the onion diversity set. A total of 765 KASP markers with minor allele frequency < 5%, with 5 or fewer missing data points, and with zero or one inconsistent alleles in the half-sib female parent analysis (Table S4) were used to determine the proportion of heterozygous alleles
The NJ tree for genotypes for the single mother plants from each of the 94 onion accessions showed long branch lengths, indicating a high degree of genetic divergence between accessions (Fig. 2). The wild species A. roylei and the bunching onion A. fistulosum formed an outgroup which also contained the A. cepa × A. fistulosum amphidiploid (GU). As expected, shallot was grouped with other onion accessions (Bark and Havey 1995). The wild species A. vavilovii grouped with A. cepa lines, suggesting that this accession may be mislabeled or is a feral A. cepa. The A. cepa accessions did not group by country of origin, most likely because SD, ID and LD cultivars can be grown in the same region depending on autumn or spring plantings. However, there was some grouping according to daylength requirements and production area. The largest grouping was predominantly composed of accessions that are spring-seeded in LD regions such as Northern Europe, Japan, and North America while another group generally comprised autumn-seeded SD cultivars grown in the Mediterranean, Middle East and India which were subsequently introduced to the southern USA (Fig. 2). A third group predominantly comprised both LD and SD spring-seeded onions from the Mediterranean and the Middle East.

Neighbour-joining phylogenetic tree showing the relationship between 94 accessions from the onion diversity set. 765 KASP™ markers with minor allele frequency < 5%, with 5 or fewer missing data points, and with zero or one inconsistent alleles in kinship analysis (Tables S2 & S4) were used to calculate the tree. Accessions marked by diamonds are wild species. Accessions marked by circles are known to have a long-day requirement for bulbing, those marked with a triangle have a short-day requirement and those with a square have an intermediate day length requirement. Filled symbols indicate that the daylength requirement has been clearly identified for that accession. Bootstrap values are from 1000 replicates and only those > 30% are displayed. Accession names are indicated by the 2- and 3-letter codes as listed in Table 1
The NJ tree for ten mother plants from each of the four selected onion founder accessions (CA, DM, GR and HO) as well as the ten individuals from a single half-sib family from one of these mother plants indicated extensive diversity both between and within HS families from the same parent accession (Fig. 3, Table S2). This shows that the genetic variability within a single onion accession can be high, owing to its out-crossing nature. However, greater genetic variation was observed between accessions than within an accession and all mother plants and half-sib individuals from the same founder onion accession clustered in the same clade indicating that an onion accession is still a recognisably discrete genetic entity (Fig. 3). Comparable levels of heterozygosity for each individual plant within a HS family were also observed (Table S3). Based on the lowest number of conflicting alleles, it was possible to deduce the male parent for each of the individuals genotyped from a single HS family (Table S4). It was found that 40–50% of the half-sib individuals were the result of self-pollination of the mother plant (5/10 for CA, 5/10 for DM, 4/10 for GR and 4/10 for HO). In addition, HS individuals were seen to cluster with their deduced male parent in the phylogenetic tree (Fig. 3).

Neighbour-joining tree of half-sib parent lines and half-sib individuals for four onion accessions. Up to ten half-sib parent lines and half-sib individuals were genotyped and a subset of 568 of the 765 KASP™ markers used to calculate the tree. P1_1 indicates parent 1 of HS family 1 while HS1_1 indicates half-sib individual 1 of accession 1 etc. The percentage bootstrap values for 1000 bootstrap repetitions are indicated on the primary branches marking each of the four accessions. HS1 = Candy F1 (CA), HS family 2 = Downy Mildew Resistant Selection (DM), HS family 3 = Giant Rocca Brown (GR) and HS family 4 = Hojem (HO). *Indicates the female parent of the ten HS individuals and colours indicate the deduced male parent (Table S4)
Population structure
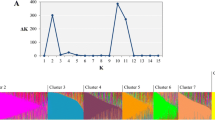
STRUCTURE analysis indicated that the best estimation of sub-population membership was K = 4. Clear clusters were observed, with q2, q3, and q4 explaining the majority of the variation (Fig. 4), corresponding to the groupings observed in the NJ tree (Fig. 2). Accessions from A. fistulosum (HLW) and A. roylei (WSR) had q1 as the dominant membership, with the A. cepa × A. fistulosum hybrid GU also having > 25% q1 membership, reflecting its pedigree. Cluster q2 contained spring-seeded LD accessions originating from northern Europe and introduced into LD production regions such as New Zealand (PL) or USA (SW). Cluster q3 contained LD, SD and ID accessions that are generally autumn-seeded and grown in the Mediterranean region. Cluster q4 was dominated by accessions from the USA, many of which are autumn-seeded SD onions (e.g. NU, GN, BE, BJ, and YB) originating from Spanish populations such as Babosa or Valenciana (Havey and Ghavami 2018). The broad groupings identified from the STRUCTURE analysis were also present in the complementary principal component analysis of the SNP data (Fig. 5). Individuals that have dominant q2, q3, or q4 cluster membership were clearly separated in dimension 1, with varying combinations of cluster memberships interspersed between these groupings. The two A. fistulosum (HLW) and A. roylei (WSR) accessions in cluster q1 were clearly resolved in dimension 2, once again highlighting how different these are from the rest of the diversity set. Interestingly, Candy F1 (CA) is derived from an LD × SD cross and was placed between q3 and q4 on the PCA plot.

Population structure for 94 accessions from the onion diversity set following analysis of KASP™ SNP data using STRUCTURE. Data plot is of estimated sub-population membership when K = 4. Data are sorted by q value membership (q1 = red, q2 = green, q3 = yellow, q4 = blue). Accessions are grouped by q value membership using a minimum threshold of ≥ 0.25. Accession names are indicated by the 2- and 3-letter codes as listed in Table 1; for reference, three letter country codes are presented on the secondary x-axis

Principal component analysis of KASP™ SNP data from the onion diversity set. Individuals factor map is estimated using HS individuals from 94 accessions genotyped at 765 loci. Individuals are coloured based on their q value cluster membership (q ≥ 0.25) following the STRUCTURE analysis (Fig. 4). Admixed clusters, are ordered in descending cluster membership, i.e. the highest q value first. Accession names are indicated by the 2- and 3-letter codes as listed in Table 1. The accession DM (Downy Mildew Resistant Selection) was placed close to the amphidiploid
Screening the onion diversity set for FBR resistance
In the seedling assay, significant variation (P < 0.001) in relative plant survival compared to the uninoculated controls (Fig. 6), ranging from 25.2% (WSV) to 88.6% (HO), was observed. Six accessions (HO, PL, DM, HYS, JC and GR) showed a significantly greater level of FOC resistance (76.4–88.6%) than the partially resistant ACP (57.6%). HS families from 78 accessions were significantly more resistant than the susceptible control cultivar Hystar (18.2%).

Percentage seedling survival (relative to the untreated control, angular transformed data) for 85 onion diversity set accessions following screening for resistance to Fusarium oxysporum f.sp cepae (isolate FUS2). Error bar indicates maximum LSD (5% level). Green bars, resistant LD/ID accessions used in the mature plant assay; purple bars, resistant SD accessions used in the mature plant assay; red bars, susceptible LD/ID accessions used in mature plant assay; orange bars, susceptible SD accessions used in mature plant assay. Accession names are indicated by the 2- and 3-letter codes as listed in Table 1
FBR resistance was confirmed in the mature plant assays. Within the SD material, HO which was the most resistant accession in the seedling assay had a significantly lower FBR (mean score 0.23) than the other three accessions tested (NU, EX, PO; Fig. 7a; P < 0.001) as well as the cv. Serrana (mean score 0.87). The most susceptible accession was PO (mean score 1.85) which was partially resistant in the seedling assay. Significant reductions in bulb weight compared to the uninoculated control (P < 0.001) were observed for Serrana, NU and PO (Fig. 7b) but not for HO. Although no significant reduction in bulb weight was observed for EX, this data do not include five plants which died due to FOC infection.

Severity of Fusarium basal rot following screening of 17 accessions from the onion diversity set for resistance to Fusarium oxysporum f.sp cepae (isolate FUS2) using a mature plant assay; a mean symptom score for SD assay; b mean bulb fresh weight for SD assay; c mean symptom score for LD assay. d Mean bulb fresh weight for LD assay. Error bars indicate LSDs (5% level) for comparing lines (a, c) or comparing inoculated and control (b, d). *Indicates a significant difference compared to the control. Accession names are indicated by the 2- and 3-letter codes as listed in Table 1
In the LD assay, HS families from seven (HYS, CA, DM, GR, ACP, MA and SY) of the nine onion accessions selected as being resistant in the seedling assay had significantly (P < 0.001) lower mean FBR disease scores (0.7–1.6) than the susceptible HY control (mean disease score 3.6, Fig. 7c). The remaining two accessions, PL and JC, had mean disease scores of 2.4 and 3.6, respectively. The highest level of FBR resistance was shown by HYS, CA and DM (mean disease scores of 0.72, 0.75 and 0.88, respectively), and for HYS there was also no significant reduction in bulb weight (Fig. 7d). HS families from the two accessions selected as being susceptible in the seedling assay (ST, GRE) were also susceptible in the mature plant assay with mean disease scores of 4.0 and 2.9, respectively. Results across all LD and SD data indicated that the seedling test correlated well with the mature plant test with a strong negative correlation (n = 16, r = − 0.57, P = 0.020) between percentage seedling survival in the seedling assay and disease score in the mature plant assay (Fig. S1). HS families from the seven accessions with the highest level of FOC resistance were in different clades of the phylogenetic tree (Fig. 2), suggesting multiple independent loci linked to a resistance phenotype.
Screening the onion diversity set for improved seedling vigour
Significant differences were observed between 83 HS families tested for root growth rate, shoot growth rate, T50, percentage germination and emergence from compacted soil (P < 0.001 for all traits). T50 values ranged from 50 h (MA) to 148 h (WSV, Fig. 8). Percentage germination was generally high across accessions indicating that seeds were of high quality, but this was not correlated with T50 (n = 87, r = − 0.12, P = 0.25, Table 2) showing that the two traits are independent. The time for roots to reach 3 cm ranged from 94 h (HYS) to 195 h (WSV, Fig. S2), while the time taken for shoots to reach 2 cm ranged from 179 h (KU) to 279 h (WSV, Fig. S2). HS families from accessions HYS, DP and WE showed very high levels of vigour for both root and shoot growth rate. There was a strong positive correlation between root and shoot growth (n = 87, r = 0.59, P < 0.0001; Table 2). A wide range of responses were also observed for seedling emergence in compacted soil across accessions, varying from 20.9% (GRE) to 72.3% (MA) relative to the uncompacted control (Fig. 9). This trait was strongly positively correlated with seed weight (n = 82, r = 0.46, P < 0.0001) and moderately positively correlated with percentage germination (n = 82, r = 0.36, P < 0.001).

Percentage germination (angular transformed) and time to 50% viability (T50) for 87 accessions from the onion diversity set. Error bars indicate LSD (5% level). Accession names are indicated by the 2- and 3-letter codes as listed in Table 1

Percentage emergence in compact soil (angular transformed data, relative to a non-compacted top layer control) for 82 accessions from the onion diversity set. Error bar indicates LSD (5% level). Accession names are indicated by the 2- and 3-letter codes as listed in Table 1
Correlation coefficients were also calculated for all other combinations of traits measured across the accessions in the diversity set (Table 2). A moderate correlation between survival in the FBR seedling assay and time for root to reach 3 cm was observed (n = 83, r = − 0.37, P < 0.001), suggesting a possible general trend between fast root growth and FOC resistance. There was also a weak correlation between FOC resistance and percentage seed germination. However, the accession LJ performed extremely well for all vigour parameters tested yet was susceptible to FOC, suggesting that FBR resistance is a discrete independent trait and not a pleiotropic effect of increased seedling vigour.
Preliminary association analysis
The Kruskal–Wallis tests identified SNP markers that were significantly associated with each of the tested traits (Table 3). For FOC resistance, five markers were identified, three of which are mapped (Duangjit et al. 2013), indicating possible quantitative trait loci (QTL) on chromosomes 1, 6B and 8. A large number of SNP markers were found to be associated with root or shoot growth with two unmapped markers associated with both. Two SNP markers (both unmapped) were significantly associated with growth in compact soil and were also associated with shoot growth. Since this method is a single marker test and does not use genomic control, a stringent P value of ≤ 0.005 was used to minimise the risk of false positives.
Discussion
Conserving biodiversity is critical for the future sustainability of crop production by providing diverse alleles for exploitation in breeding programmes (Keller et al. 2013). Central Asia is the centre of diversity for many Alliums and has been a source of wild species exhibiting potentially useful traits (Simon 2005; Pandey et al. 2008; Baldwin et al. 2012a; Havey and Ghavami 2018). However, the difficulty in introgressing traits means that it can take as long as 20 years to breed these traits into a commercial cultivar (Scholten et al. 2007; Chuda and Adamus 2008). For this reason, the diversity set reported here was predominantly created using A. cepa accessions. Many collections contain large numbers of Allium accessions (Khosa et al. 2016), too many to feasibly phenotype for traits of interest. To address this, diversity core collections are often developed which aim to represent the diversity in a crop in a manageable number of accessions (Walley and Moore 2015). A recent study examined genetic diversity in a set of 73 onion accessions consisting of commercial cultivars (3 countries) and Italian landraces (Vilano et al. 2019). Crops such as barley and lettuce are naturally inbreeding and as such diversity sets derived from genebank accessions are more amenable to association mapping (Houston et al. 2014; Walley et al. 2017). In other crops, such as vegetable brassicas, value has been added to diversity sets by generating homozygous lines (Walley et al. 2012; Hatzig et al. 2015; Havlickova et al. 2018).
We developed a unique onion diversity set, designed to capture diversity of this crucially important vegetable crop, comprising 90 A. cepa accessions from 23 different countries, together with four close relatives and an A. cepa × A. fistulosum hybrid. As onion exhibits severe inbreeding depression (Brewster 2008), the generation of a homozygous diversity set was not possible; therefore, value was added by creating seed stocks of multiple HS families for each accession. Seed is stored under Genebank conditions meaning it will remain viable for decades (FAO 2013). Using KASP™ markers (Duangjit et al. 2013), we determined that genetic diversity was captured both across the diversity set and within accessions, thus providing an invaluable resource for future research and breeding. Our analysis revealed that 765 of the published SNPs were informative for this diversity set, whereas a recent publication on 73 onion accessions (Vilano et al. 2019) tested only 400 of these SNPs, finding 375 to be informative. The observation that germination rate was high indicates that vigour was maintained due to the approach of generating HS families and the KASP™ marker data indicated that this is due to heterozygosity being maintained. Despite this, the analysis of individuals within HS families and between mother plants (Fig. 3) showed that the genetics of the original accessions were still maintained as individuals and parents from a HS family clustered together. The KASP™ markers revealed a moderate level of heterozygosity (mode 30–35%) in the HS families with the exception of A. roylei (WSR) and A. fistulosum (HLW). This is likely due to the fact that the KASP™ markers were designed to SNPs between two A. cepa populations (Duangjit et al. 2013) and are not polymorphic in other Allium species. A previous study using 1226 SNP markers reported slightly lower heterozygosity (23.5%) in 14 OP onion populations (Havey and Ghavami 2018) while another reported 22% heterozygosity using 166 SSR markers to assess 24 populations (Baldwin et al. 2012a). Conversely, an earlier study using 56 SSR markers showed that the median level of heterozygosity in 72 OP onion populations was 70% (McCallum et al. 2008). A moderate to high level of heterozygosity is consistent with onion being an outbreeding species, and relatively high levels (up to 20%) of heterozygosity have even been observed in inbred onion lines (Bradeen and Havey 1995; Duangjit et al. 2013). The present study is the first to examine heterozygosity across a large panel of diverse onion accessions. We observed a small number of onion accessions which were not highly heterozygous (e.g. BE and DM) which might have been expected to lack vigour due to inbreeding depression (Jones and Davis 1944; Brewster 2008). However, while BE has slow root and shoot growth rates, DM did not suffer any loss of vigour suggesting that either this low level of heterozygosity (5.8%) is sufficient to retain vigour or this accession is heterozygous at the loci needed to maintain vigour.
Genotyping of 91 diversity set accessions showed that they were related to each other predominantly through long phylogenetic branches indicating a low degree of genetic redundancy. There was also considerable variation within the four HS families analysed, indicating that an onion cultivar is a population of non-identical but genetically related individuals. This has implications for introgressing traits into a new cultivar as attention will need to be paid to maximising the frequency of beneficial alleles into as many individuals as possible. The results from the phylogenetic and population structure analyses supported the historic breeding of onion for adaption to daylengths at different latitudes (Brewster 2008; Taylor et al. 2010), highlighting the importance of conserving a range of SD, LD and ID onion lines in Genebank collections. Furthermore, both analyses grouped a shallot line (AF, A. cepa var. ascalonicum) with A. cepa supporting the previous assertion that this species should be classified as A. cepa (Brewster 2008; McCallum et al. 2008).
The onion diversity set was successfully screened for resistance to FBR, a global problem for onion growers (Cramer 2000; Taylor et al. 2016). Previous studies have identified partial resistance or ‘tolerance’ to FBR in A. cepa, but high-level resistance is lacking (Saxena and Cramer 2009; Taylor et al. 2013; Gei et al. 2014). While high levels of FBR resistance have been identified in other Allium species (Galván et al. 2008; Rout et al. 2016), the problems associated with introgression of this trait into A. cepa have prevented this resistance being exploited. We identified high levels of FBR resistance in a number of A. cepa accessions from different genetic backgrounds which in the future may be particularly valuable for ‘stacking’ of resistance genes for improved durability. Furthermore, FBR resistance was identified in both SD and LD material thus simplifying breeding for different latitudes. Our diversity set also included an A. fistulosum accession (HLW), a species which has previously been shown to have higher resistance to FOC than A. cepa (Abawi and Lorbeer 1971a; Holz and Knox-Davies 1974; Galván et al. 2008). However, in the seedling assay, HS families from several A. cepa accessions were as resistant as HLW, while HO was significantly more resistant. This confirms the utility of the onion diversity set in identifying high levels of resistance to FBR in A. cepa. A significant correlation was observed between seedling and mature plant assays, suggesting that the former can be used as a rapid phenotyping approach to screen large populations for resistance. This confirms our previous work (Taylor et al. 2013) and that of others (Retig et al. 1970; Özer et al. 2004) who confirmed that there is a strong correlation between the results of glasshouse and field resistance screening. However, we suggest that seedling resistance should be confirmed using a mature plant assay as this tests resistance at the bulb stage. While resistance was only confirmed using a single FOC isolate, this isolate was shown to be highly virulent and previous studies suggested that there is no cultivar × isolate interaction (Taylor et al. 2013; Gei et al. 2014). A partial correlation was observed between FBR resistance and root growth rate, suggesting that these two traits may be linked. However, this was not always the case as HS family LJ scored highly in all vigour assays but did not exhibit FOC resistance. In previous research, Allium accessions with denser root systems had higher resistance to FOC (Galván et al. 2008). Developing SNP markers linked to FBR resistance in onion will allow rapid breeding using marker-assisted selection, something which would be greatly beneficial due to the biennial life cycle of onion. Association analyses identified three SNPs significantly correlated with FBR resistance. While this result suggests an excellent chance of identifying QTL in the future, linkage of these markers to FBR resistance loci needs to be confirmed using biparental haploid mapping populations derived from resistant × susceptible crosses.
Seedling vigour is a critical trait for sustainable crop production (Finch-Savage et al. 2010; Finch-Savage and Bassel 2016), and this is the first time that a diverse range of onion accessions have been examined for this trait. Highly vigorous accessions were identified and rapid and high-throughput phenotyping assays developed, something which is often the ‘bottleneck’ for genetic analyses. The protocol used here aligns with the ISTA seed testing protocols for onion (ISTA 2013). It was apparent that many of the individual components that constitute ‘vigour’ are correlated, and potentially under common genetic control with the notable exception of T50 which was a distinct trait. This is an important finding as fewer parameters could be selected to assess vigour of different seed lots. Of particular note is the strong correlation between seed weight and emergence in compact soil. A similar observation was made for other crops (Finch-Savage et al. 2010; Finch-Savage and Bassel 2016) meaning that seed weight can be used as a predictor of emergence in compact soil, which is relevant to production in soils liable to capping. All seedling vigour parameters measured here will potentially influence establishment in the field and the HS family LJ performed well across all of them, providing an excellent source of pre-breeding material for this trait.
Kruskal–Wallis tests identified SNP markers that may be linked to both FBR resistance and seedling vigour. In the absence of estimates of linkage disequilibrium, these markers represent preliminary associations. Since this method is a single marker test without a genomic control, a stringent P value of ≤ 0.005 was used to reduce false positives. When the genome sequence of onion becomes available, we will be able to align the markers used in this research, and generate additional markers to gain a representative sampling of the genome, and estimate the underlying linkage disequilibrium. Until then, the putative marker-trait associations identified in this work are a useful starting point for future selection in breeding programmes, in combination with the phenotypic variation for FBR resistance and seedling vigour that we have discovered.
References
Abawi GS, Lorbeer JW (1971a) Pathological histology of four onion cultivars infected by Fusarium oxysporum f. sp. cepae. Phytopathology 61:1164–1169
Abawi GS, Lorbeer JW (1971b) Reaction of selected onion varieties to infection by Fusarium oxysporum f. sp. cepae. Plant Dis Report 55:1000–1004
Arumuganathan K, Earle ED (1991) Nuclear DNA content of some important plant species. Plant Mol Biol Report 9:208–218
Baldwin S, Pither-Joyce M, Wright K, Chen L, Mccallum J (2012a) Development of robust genomic simple sequence repeat markers for estimation of genetic diversity within and among bulb onion (Allium cepa L.) populations. Mol Breed 30:1401–1411
Baldwin S, Revanna R, Thomson S, Pither-Joyce M, Wright K, Crowhurst R, Fiers M, Chen L, Macknight R, Mccallum JA (2012b) A toolkit for bulk PCR-based marker design from next-generation sequence data: application for development of a framework linkage map in bulb onion (Allium cepa L.). BMC Genom 13:637
Bark OH, Havey MJ (1995) Similarities and relationships among open-pollinated populations of the bulb onion as estimated by nuclear RFLPs. Theor Appl Genet 90:607–614
Bettey M, Finch-Savage WE, King GJ, Lynn JR (2000) Quantitative genetic analysis of seed vigour and pre-emergence seedling growth traits in Brassica oleracea. New Phytol 148:277–286
Bleasdale J (1967) The relationship between the weight of a plant part and total weight as affected by plant density. J Hortic Sci 42:51–58
Bradeen JM, Havey MJ (1995) Randomly amplified polymorphic DNA in bulb onion and its use to assess inbred integrity. J Am Soc Hortic Sci 120:752–758
Brayford D (1996) IMI descriptions of fungi and bacteria set 127. Mycopathologia 133:35–63
Brewster JL (2008) Onions and other vegetable alliums, 2nd edn. CAB International, Wallingford
Chuda A, Adamus A (2008) Aspects of interspecific hybridization within edible Alliaceae. Acta Physiol Plant 31:223
Cramer C (2000) Breeding and genetics of Fusarium basal rot resistance in onion. Euphytica 115:159–166
Cramer CS (2006) Onion trait heritability and response from selection. J Am Soc Hortic Sci 131:646–650
Currah L, Ockendon DJ (1983) Onion pollination by blowflies and honeybees in large cages. Ann Appl Biol 103:497–506
R Core Team (2011) R: a language and environment for statistical computing. The R foundation for statistical computing, Vienna, Austria. https://www.R-project.org
Duangjit J, Bohanec B, Chan AP, Town CD, Havey MJ (2013) Transcriptome sequencing to produce SNP-based genetic maps of onion. Theor Appl Genet 126:2093–2101
Earl DA, Von Holdt BM (2012) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 4:359–361
Entwistle AR (1990) Root diseases. In: Brewster JL (ed) Onions and allied crops. CRC Press, Boca Raton, pp 103–154
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620
FAO (2013) Genebank standards for plant genetic resources for food and agriculture. Rome
Faostat (2016) Food and agricultural organization of the united nations-production statistics. https://www.fao.org/faostat/en/#data. Accessed Nov 2018
Finch-Savage WE, Bassel GW (2016) Seed vigour and crop establishment: extending performance beyond adaptation. J Exp Bot 67:567–591
Finch-Savage WE, Clay HA, Lynn JR, Morris K (2010) Towards a genetic understanding of seed vigour in small-seeded crops using natural variation in Brassica oleracea. Plant Sci 179:582–589
Galván G, Koning-Boucoiran C, Koopman W, Burger-Meijer K, González P, Waalwijk C, Kik C, Scholten O (2008) Genetic variation among Fusarium isolates from onion, and resistance to Fusarium basal rot in related Allium species. Eur J Plant Pathol 121:499–512
Gei PFC, Valdez JG, Piccolo RJ, Galmarini CR (2014) Influence of Fusarium spp. isolate and inoculum density on resistance screening tests in onion. Trop Plant Pathol 39:19–27
Gray D, Steckel JRA (1983) A comparison of methods for evaluating seed quality in carrots (Daucus carota). Ann Appl Biol 103:327–334
Griffiths G, Trueman L, Crowther T, Thomas B, Smith B (2002) Onions—a global benefit to health. Phytother Res 16:603–615
Hatzig SV, Frisch M, Breuer F, Nesi N, Ducournau S, Wagner MH, Leckband G, Abbadi A, Snowdon RJ (2015) Genome-wide association mapping unravels the genetic control of seed germination and vigor in Brassica napus. Front Plant Sci 6:221
Havey MJ, Ghavami F (2018) Informativeness of Single Nucleotide Polymorphisms and relationships among onion populations from important world production regions. J Am Soc Hortic Sci 143:34–44
Hodgkin T, Hegarty TW (1978) Genetically determined variation in seed germination and field emergence of Brassica oleracea. Ann Appl Biol 88:407–413
Holz G, Knox-Davies PS (1974) Resistance of onion selections to Fusarium oxysporum f. sp.cepae. Phytophylactica 6:153–156
Houston K, Russell J, Schreiber M, Halpin C, Oakey H, Washington JM, Booth A, Shirley N, Burton RA, Fincher GB, Waugh R (2014) A genome wide association scan for (1,3;1,4)-β-glucan content in the grain of contemporary 2-row Spring and Winter barleys. BMC Genom 15:907
ISTA (2013) Handbook on seedling evaluation. The International Seed Testing Association, Bassersdorf
Jakobsson M, Rosenberg NA (2007) CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 23:1801–1806
Jo J, Purushotham PM, Han K, Lee H-R, Nah G, Kang B-C (2017) Development of a genetic map for onion (Allium cepa L.) using reference-free genotyping-by-sequencing and SNP assays. Front Plant Sci 8:1606
Jones HA, Davis D (1944) Inbreeding and heterosis and their relation to the development of new varieties of onion. U.S. Deptartment of Agriculture Technical Bulletin 874
Keller ERJ, Zanke C, Senula A, Breuing A, Hardeweg B, Winkelmann T (2013) Comparing costs for different conservation strategies of garlic (Allium sativum L.) germplasm in genebanks. Genet Resour Crop Evol 60:913–926
Khosa JS, Mccallum J, Dhatt AS, Macknight RC (2016) Enhancing onion breeding using molecular tools. Plant Breed 135:9–20
King JJ, Bradeen JM, Bark Mccallum JA, Havey MJ (1998) A low-density genetic map of onion reveals a role for tandem duplication in the evolution of an extremely large diploid genome. Theor Appl Genet 96:52–62
Krueger SK, Weinman AA, Gabelman WH (1989) Combining ability among inbred onions for resistance to Fusarium basal rot. HortScience 24:1021–1023
Kuhl JC, Cheung F, Yuan Q, Martin W, Zewdie Y, McCallum J, Catanach A, Rutherford P, Sink KC, Jenderek M, Prince JP, Town CD, Havey MJ (2004) A unique set of 11,008 onion expressed sequence tags reveals expressed sequence and genomic differences between the monocot orders Asparagales and Poales. Plant Cell 16:114–125
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874
Lê S, Josse J, Husson F (2008) FactoMineR: an R package for multivariate analysis. J Stat Softw 25:18
Havlickova L, He Z, Wang L, Langer S, Harper AL, Kaur H, Broadley MR, Gegas V, Bancroft I (2018) Validation of an updated Associative Transcriptomics platform for the polyploid crop species Brassica napus by dissection of the genetic architecture of erucic acid and tocopherol isoform variation in seeds. Plant J 93:181–192
Martin W, Mccallum J, Shigyo M, Jakse J, Kuhl J, Yamane N, Pither-Joyce M, Gokce A, Sink K, Town C, Havey M (2005) Genetic mapping of expressed sequences in onion and in silico comparisons with rice show scant colinearity. Mol Genet Genom 274:197–204
Mccallum J, Thomson S, Pither-Joyce M, Kenel F, Clarke A, Havey MJ (2008) Genetic diversity analysis and single-nucleotide polymorphism marker development in cultivated bulb onion based on expressed sequence tag–simple sequence repeat markers. J Am Soc Hortic Sci 133:810–818
Özer N, Köycü N, Chilosi G, Magro P (2004) Resistance to fusarium basal rot of onion in greenhouse and field and associated expression of antifungal compounds. Phytoparasitica 32:388–394
Pandey A, Pandey R, Negi KS, Radhamani J (2008) Realizing value of genetic resources of Allium in India. Genet Resour Crop Evol 55:985–994
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
Retig N, Kust AF, Gabelman WH (1970) Greenhouse and field tests for determining the resistance of onion lines to Fusarium basal rot. J Am Soc Hortic Sci 95:422–424
Rout E, Tripathy P, Nanda S, Nayak S, Joshi RK (2016) Evaluation of cultivated and wild allium accessions for resistance to Fusarium oxysporum f. sp. cepae. Proc Natl Acad Sci India Sect B Biol Sci 86:643–649
Saxena A, Cramer CS (2009) Screening of onion seedlings for resistance against New Mexico isolates of Fusarium oxysporum f. sp. cepae. J Plant Pathol 91:199–202
Scholten OE, Heusden AW, Khrustaleva LI, Burger-Meijer K, Mank RA, Antonise RGC, Harrewijn JL, Haecke W, Oost EH, Peters RJ, Kik C (2007) The long and winding road leading to the successful introgression of downy mildew resistance into onion. Euphytica 156:345–353
Shigyo M, Khar A, Abdelrahman M (2018) The allium genomes. Springer, Cham
Simon PW (2005) Realizing value from central Asian Allium germplasm collections. HortScience 40:309–310
Taylor A, Massiah AJ, Thomas B (2010) Conservation of Arabidopsis thaliana photoperiodic flowering time genes in onion (Allium cepa L.). Plant Cell Physiol 51:1638–1647
Taylor A, Vagany V, Barbara DJ, Thomas B, Pink DAC, Jones JE, Clarkson JP (2013) Identification of differential resistance to six Fusarium oxysporum f. sp. cepae isolates in commercial onion cultivars through the development of a rapid seedling assay. Plant Pathol 62:103–111
Taylor A, Vágány V, Jackson AC, Harrison RJ, Rainoni A, Clarkson JP (2016) Identification of pathogenicity-related genes in Fusarium oxysporum f. sp. cepae. Mol Plant Pathol 17:1032–1047
Van Heusden AW, Van Ooijen JW, Vrielink-Van Ginkel R, Verbeek WHJ, Wietsma WA, Kik C (2000) A genetic map of an interspecific cross in Allium based on amplified fragment length polymorphism (AFLPTM) markers. Theor Appl Genet 100:118–126
Van Ooijen JW (2009) MapQTL 6, Software for mapping the quantitative trait loci in experimental populations of diploid species. Kyazma BV (Wageningen, The Netherlands)
Vilano C, Esposito S, Carucci F, Iorizzo M, Frusciante L, Carputo D, Aversano R (2019) High-throughput genotyping in onion reveals structure of genetic diversity and informative SNPs useful for molecular breeding. Mol Breed 39:5
Walley PG, Hough G, Moore JD, Carder J, Elliott M, Mead A, Jones J, Teakle G, Barker G, Buchanan-Wollaston V, Hand P, Pink D, Collier R (2017) Towards new sources of resistance to the currant-lettuce aphid (Nasonovia ribisnigri). Mol Breed 37:4
Walley PG, Teakle GR, Moore JD, Allender CJ, Pink DAC, Buchanan-Wollaston V, Barker GC (2012) Developing genetic resources for pre-breeding in Brassica oleracea L.: an overview of the UK perspective. J Plant Biotechnol 39:62–68
Walley PG, Moore JD (2015) Biotechnology and genomics helping to understand and uncover the hidden potential of Crop Wild Relatives. In: Yadav S, Redden B, Fowler C, Maxted N, Smith P, Guarino L, Snook L, Dullo E (eds) Crop Wild relatives and climate change. Wiley, Hoboken
Welham SJ, Thompson R (1997) Likelihood ratio tests for fixed model terms using residual maximum likelihood. J R Stat Soc Ser B (Stat Methodol) 59:701–714
Whittington W (1973) Genetic regulation of germination. In: Heydecter W (ed) Seed Ecology. Butterworths, London, pp 5–30
Acknowledgements
We thank the Warwick Crop Centre GRU for providing onion seed and Horticultural Services staff for maintaining plants. We also acknowledge the support of Hazera Seeds (Reinout de Heer, Wessel van Leeuwen and Ningwen Zhang). This work was funded by the department for Food and Rural Affairs (Defra) as part of the Vegetable Genetic Improvement Network (VeGIN project IF01) and by the Biotechnology and Biological Sciences Research Council (BBSRC) as part of the Horticulture and Potato Initiative (HAPI; BB/K020870/1).
Author information
Authors and Affiliations
Contributions
AT planned and carried out the experiments, analysed data, created figures and drafted/edited the manuscript; GT analysed the genotyping data, created figures and wrote/edited the manuscript; PW carried out the population structure, PCA, and association analyses, created figures and wrote/edited the manuscript; WFS aided experimental design and wrote/edited the manuscript; AJ carried out laboratory/glasshouse experiments with AT and edited the manuscript; JJ designed experiments and was responsible for statistical analyses; PH obtained funding, was involved in planning experiments and edited the manuscript; BT managed the project, was involved in planning experiments and edited the manuscript; MH was involved in the planning and analysis of genotyping experiments and edited the manuscript; DP was responsible for the original project ideas, obtained funding, planned experiments, selected accessions and supervised development of the A. cepa diversity set, managed the research and edited the manuscript; JC obtained funding, planned experiments and wrote/edited the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Communicated by Heiko C. Becker.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Taylor, A., Teakle, G.R., Walley, P.G. et al. Assembly and characterisation of a unique onion diversity set identifies resistance to Fusarium basal rot and improved seedling vigour. Theor Appl Genet 132, 3245–3264 (2019). https://doi.org/10.1007/s00122-019-03422-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00122-019-03422-0