
Abstract
Activated small ubiquitin-like modifiers (SUMOs) have been implicated in neuropathological processes following ischemic stroke. However, the target proteins of SUMOylation and their contribution to neuronal injury remain to be elucidated. MLK3 (mixed-lineage kinase 3), a member of the mitogen-activated protein kinase kinase kinase (MAPKKK) family, is a critical regulator of neuronal lesions following cerebral ischemia. Here, we found that SUMOylation of MLK3 increases in both global and focal ischemic rodent models and primary neuronal models of oxygen and glucose deprivation (OGD). SUMO1 conjugation at the Lys401 site of MLK3 promoted its activation, stimulated its downstream p38/c-Jun N-terminal kinase (JNK) cascades, and led to cell apoptosis. The interaction of MLK3 with PIAS3, a SUMO ligase, was elevated following ischemia and reperfusion. The PINIT domain of PIAS3 was involved in direct interactions with MLK3. Overexpression of the PINIT domain of PIAS3 disrupted the MLK3-PIAS3 interaction, inhibited SUMOylation of MLK3, suppressed downstream signaling, and reduced cell apoptosis and neurite damage. In rodent ischemic models, the overexpression of the PINIT domain reduced brain lesions and alleviated deficits in learning, memory, and sensorimotor functions. Our findings demonstrate that brain ischemia-induced MLK3 SUMOylation by PIAS3 is a potential target against poststroke neuronal lesions and behavioral impairments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ischemic stroke causes neuronal damage and leads to high rates of disability and mortality [1]. The main clinical treatments are thrombolysis and thrombectomy to restore blood supply [2, 3]. However, blood reperfusion treatments does not directly intervene in the neuronal damage caused by stroke. Due to the limitations, thrombolysis and thrombectomy are applied to only a minor proportion of stroke patients. Therefore, exploring the molecular mechanisms underlying stroke-induced neuronal injury is essential to identify potential intervention targets.
Mixed lineage kinase 3 (MLK3) belongs to a superfamily of mitogen-activated protein kinase kinase kinase (MAPKKK) and functions as an upstream initiator of multiple signaling events, including the c-Jun N-terminal kinase (JNK) and P38 MAPK pathways, in response to stress, inflammation, and excitotoxic stimuli [4,5,6,7,8,9]. Upon elicitation, MLK3 is activated by phosphorylation at the Thr277/Ser281 residues [4]. Activated MLK3 then phosphorylates MAPK kinases (MKKs), such as MKK4/7 and MKK3/6, activating the JNK and p38 MAPK pathways. The MLK3–MKK–MAPK signaling cascades contribute to the pathogenesis of ischemic brain lesions [10,11,12]. MLK3 is a critical upstream regulator of cerebral ischemia, and blocking MLK3 activation exerts neuroprotective effects. Elucidating the regulatory mechanisms underlying MLK3 activation will provide potential therapeutic targets for cerebral ischemic injury.
Protein post-translational modification is critical for regulating protein structure and function. SUMOylation, a regulatory post-translational modification, involves the reversible covalent conjugation of small ubiquitin-like modifiers (SUMOs) to target proteins at lysine residues [13, 14]. Conjugation of SUMO1 with target proteins regulates neuronal functions such as neuronal excitability and synaptic plasticity [15,16,17,18,19,20,21,22]. Mounting evidence indicates that abnormal SUMOylation homeostasis contributes to various neurological disorders, including ischemic stroke, Alzheimer's disease, Huntington's disease, and Parkinson's disease [23,24,25,26,27,28,29,30]. In stroke patients, SUMO1 expression is upregulated in penumbra brain tissues [31]. Moreover, an imbalance in SUMOylation homeostasis has been detected in both global and focal cerebral ischemic animal models [32,33,34,35,36,37,38,39,40]. However, whether MLK3 is a target of SUMO1 modification and the role of MLK3 SUMOylation in ischemic stroke remains unknown.
SUMOylation is an enzymatic cascade analogous to the ubiquitin pathway, in which the SUMO protein is successively conjugated to target proteins by SUMO-activating enzymes, SUMO-specific conjugating enzymes, and substrate-selective SUMO ligases [14, 23, 24]. The SUMO ligase PIAS3 colocalizes with SUMO1 in the hippocampal neurons [15, 41]. PIAS3 participates in the SUMOylation of various neuronal proteins to regulate neuronal function [15, 18, 21, 22]. However, it is unknown whether PIAS3 is involved in the SUMOylation of MLK3.
In the study, we identified an additional mechanism for MLK3 regulation by SUMOylation, and PIAS3 facilitated MLK3 SUMOylation after cerebral ischemia. Disruption of the MLK3-PIAS3 interaction by the PINIT domain of PIAS3 inhibited MLK3 SUMOylation and its downstream apoptotic signaling and reduced neurological deficits in neuronal and rodent ischemia models.
Materials and methods
Antibodies and plasmids
Primary antibodies against PIAS3 (cat #4164S), SUMO1 (4930S), MLK3 (2817S), cleaved caspase-3 (9661S), p38 (9212S), and phospho-JNKs (Thr183 + Tyr185, 4668S) were from Cell Signaling Technology (Danvers, MA, USA). Flag antibody (F1804S) was purchased from Sigma (St. Louis, MO, USA). Myc antibody (05-419) was purchased from Millipore (St. Louis, MO, USA). PIAS3 (sc-14017), JNK (sc-7345), MLK3 (sc-166639), and SUMO1 (sc-5308) antibodies were purchased from Santa Cruz Biotechnology (Dallas, Texas, USA). MAP2 (17490-1-AP) and β-actin (66009-1-Ig) antibodies were purchased from Proteintech (Wuhan, Hubei, China). The phospho-MLK3 antibody (Thr277 + Ser281, AF8079) was purchased from Affinity (Jiangsu, China). Phospho-p38 antibody (ER2001-52) was purchased from HUABIO (Hangzhou, Zhejiang, China). Horseradish peroxidase-labeled goat anti-mouse and anti-rabbit IgG (H + L), Alexa Fluor 488 goat anti-mouse IgG (H + L), and Alexa Fluor 594 goat anti-rabbit IgG (H + L) antibodies were from Thermo Fisher Scientific (Waltham, MA, USA). IRDye®680RD goat anti-rabbit (926–68071) and IRDye®C800CW goat anti-mouse (926–32210) antibodies were purchased from LI-COR Biosciences (Lincoln, Nebraska, USA).
The vector pcDNA3.1( +) was used to express Myc-tagged MLK3, HA-tagged SUMO1, PIAS3, His-tagged wild-type PIAS3 (His-PIAS3 WT), Myc-tagged wild-type PIAS3 (Myc-PIAS3 WT) and its corresponding domain-deleted mutant constructs (AAs 1–500, 1–320, 1–132, and 133–320). The point mutation of MLK3 from lysine 401 to arginine (MLK3 K401R) was generated by QuickChange II XL site-directed mutagenesis kit from Agilent Technologies (Santa Clara, CA, USA).
Animals
Sprague‒Dawley rats and C57BL/6J mice were group-housed on a 12 h light–dark cycle with free access to food and water before surgery. All experimental procedures were implemented in accordance with the guidelines described in the revised 2011 Chinese Regulations for the Administration of Affairs Concerning Experimental Animals and approved by the Institutional Animal Care and Use Committee of Xuzhou Medical University. All efforts were applied to minimize the potential discomfort and pain of animals during experiments.
Primary cortical neuron culture and lentivirus infection
Cortical neurons were isolated from the cerebral cortices of E18 rats and plated at a density of 0.8–1.0 × 105 cells/cm2 onto poly-D-lysine-coated dishes or coverslips to culture at 37 °C/5% CO2 [21, 22]. The culture medium (Neurobasal serum-free medium with 0.5 mM GlutaMax™ and 2% B27) was replaced every 3 days. Lentivirus expressing the PINIT domain of PIAS3 (Lenti-PINIT), negative control, MLK3 WT or its K401R mutant was added to the cortical culture (MOI = 10) at DIV 9.
Oxygen and glucose deprivation/reoxygenation (OGD/R) treatment
OGD treatment was performed on DIV 13 primary neurons. For OGD treatment, the cortical neurons were kept in DMEM without glucose at 1% O2 for 2 h. After reoxygenation for the indicated periods, the neurons were lysed in ice-cold homogenization buffer [20 mM N-ethylmaleimide (NEM), 50 mM MOPS (pH 7.4), 320 mM sucrose, 100 mM KCl, 0.5 mM MgCl2, phosphatase and protease inhibitors (20 mM β-glycerophosphate, 20 mM sodium pyrophosphate, 50 mM NaF, 1 mM each of EDTA, EGTA, sodium orthovanadate, p-nitrophenyl phosphate, PMSF and benzamidine, and 5 μg/ml each of aprotinin, leupeptin and pepstatin A)] and used for immunoblotting and immunoprecipitation.
Transient global ischemia model
Transient global brain ischemia was induced by the four-vessel occlusion method as described previously [37]. Briefly, male rats (220–300 g) were anesthetized by isoflurane. The vertebral arteries were electrocauterized. On the following day, the carotid arteries were occluded for 15 min to induce ischemia. The sham-operated rats were subjected to the same surgical procedures except for occluding carotid arteries. Rat rectal temperature was maintained at 36.5 °C–37.5 °C during brain ischemia and 2 h of reperfusion.
Middle cerebral artery occlusion (MCAO) model
The mouse MCAO/R method was used to establish a transient focal cerebral ischemic model [29, 34]. Briefly, male mice (25–30 g) were anesthetized with 2,2,2-tribromoethanol (0.02 mL/g). A 7–0 nylon monofilament with a silicone-embedded front end 200–250 mm in diameter was inserted into the right internal carotid artery (ICA) to block the origin of the right middle cerebral artery (MCA). After 60 min of MCAO, the nylon monofilament was removed to restore the blood (reperfusion). The sham-operated mice underwent the same surgical procedures except for occluding carotid arteries.
Adeno-associated virus (AAV) injection
The rats (170–180 g) were stereotaxically injected with 7.12 × 109 GC of AAVs expressing the PINIT domain of PIAS3 [AAV-PINIT] or the negative control (AAV-NC) into the bilateral hippocampal CA1 region with two points (relative to bregma: 3.3 mm anteroposterior, 2.0 mm lateral, and 2.8/3.1 mm depth; 3.9 mm anteroposterior, 3.0 mm lateral, and 2.6/3.1 mm depth) and then subjected to a transient global ischemia model after 5 weeks. The mice (20–22 g) were stereotaxically injected with 6.4 × 109 GC of AAV-PINIT or AAV-NC into the right cortex at two points (relative to bregma: 0.8 mm anteroposterior, 2.5 mm lateral, and 2 mm depth; −1.5 mm anteroposterior, 3.0 mm lateral, and 2 mm depth) and then subjected to the MCAO model after 5 weeks.
Tissue preparation
Rat hippocampal CA1 or mouse cerebral hemispheres were isolated and homogenized in ice-cold homogenization buffer. The homogenates were centrifuged at 800 × g for 10 min at 4 °C, and the supernatants were collected for immunoprecipitation and immunoblot analysis.
Immunoblotting
Equal amounts of protein samples were separated by SDS‒PAGE and then electrotransferred to nitrocellulose membranes (Amersham Biosciences). After the membranes were blocked with 3% bovine serum albumin, incubated with the indicated primary antibodies followed by horseradish peroxidase-conjugated secondary antibodies, and then, the membranes were developed with Immobilon® Western Chemiluminescent HRP Substrate (Millipore). The images were obtained and analyzed by the ChemiDoc-It imaging system Fluor Chem FC3 (Protein Sample).
Immunoprecipitation
Protein samples were diluted with immunoprecipitation (IP) buffer [20 mM NEM, 50 mM HEPES (pH 7.1), 10% glycerol (vol/vol), 150 mM NaCl, 1 mM ZnCl2, 1.5 mM MgCl2, 1% Triton X-100, 0.5% NP-40, and phosphatase and protease inhibitor as described above] and incubated with the indicated primary antibodies and Protein A + G magnetic beads (Beyotime) overnight at 4 °C. The beads were gathered by a magnetic stand and washed with IP buffer three times. Then, the bound proteins were eluted by boiling for 5 min in Laemmli sample buffer and used for immunoblot analysis.
Cell culture and plasmid transfection
HEK293T cells or HT22 cells were cultured in DMEM with 10% fetal bovine serum. When the cells reached approximately 90% confluence, plasmids were transiently transfected with polyethylenimine (PEI, Sigma) [21]. Briefly, DNA and PEI were dissolved individually in DMEM, and then, the PEI/DMEM mix was added to the DNA/DMEM mix. After 15 min of incubation, the PEI/DNA/DMEM mixtures were added to the culture medium. A medium change was performed after 2–3 h of incubation. The cells were collected for assays 24 h post-transfection.
In vitro SUMOylation assay
The SUMOylation kit (Enzo Life Sciences, NY, USA) was used for the in vitro SUMOylation assay. The reaction system [10 × SUMOylation buffer (2 μL), 20 × E1 activating enzyme (1 μL), 20 × E2 conjugating enzyme (1 μL), 20 × Mg-ATP (1 μL), 20 × SUMO1 (WT or mutant, 1 μL), synthesized Myc-MLK3 (390–413) (WT or K401R, 0.4 μg)] was incubated at 37 °C for 4 h [21]. Then, the reaction was boiled in Laemmli sample buffer for 5 min for analysis by immunoblotting.
His pulldown assay
His-PIAS3 WT or 133–320-Myc was purified with high-affinity Ni-Charge resin (GeneScript, Nanjing, China). Briefly, HEK293T cells overexpressing His-PIAS3 WT or 133–320-Myc were lysed in ice-cold lysis buffer (50 mM NaH2PO4, 300 mM NaCl, pH 8.0). Cleared lysates were incubated with Ni2+-NTA resin. Next, the resins loaded with His-tagged PIAS bait protein were mixed with cell lysate overexpressing MLK3 at 4 °C for 3 h. After 3 washes with wash buffer (50 mM NaH2PO4, 300 mM NaCl, 60 mM imidazole, pH 8.0), the proteins on the resin were eluted through boiling in Laemmli sample buffer.
Purification of Tat-tagged PIAS3 (133–320) (Tat-PINIT)
Tat-tagged PIAS3(133–320) cDNAs were first subcloned into the pET-28( +) vector, and then induced to express in the Escherichia coli strain BL21. The harvested BL21 cells were resuspended in lysis buffer and then sonicated by 180 cycles of 1-s bursts at high intensity followed by a 3-s cooling period. Then, the lysate was cleared by centrifugation and loaded onto Ni2+-NTA resin. The bound proteins were eluted with elution buffer (50 mM NaH2PO4, 300 mM NaCl, 250 mM imidazole, pH 8.0) and then desalted and concentrated in an Amicon Ultra 10 kD centrifuge tube (Millipore) [21].
Immunofluorescence
Neurons were first fixed with 4% paraformaldehyde for 20 min at room temperature (RT) and blocked with 10% normal goat serum (NGS) and 0.3% Triton X-100 in PBS for 1 h. Next, the neurons were successively incubated with primary antibodies, Alexa Fluor 594 goat anti-rabbit and Alexa Fluor 488 goat anti-mouse secondary antibodies, and DAPI counterstain. The stained neurons were mounted with ProLong™ Diamond Antifade Mountant (Invitrogen). Confocal images were acquired by a Zeiss LSM710 laser-scanning confocal microscope with 6–8 randomly chosen fields (200 ×) per well. The overall dendritic length and cell numbers were calculated by ImageJ software, and the average dendrite length of a single cell was used for statistical analysis. The data are from a single experiment (four wells) performed in triplicate.
Propidium iodide (PI) staining
Neurons were first incubated with Hoechst 33342 (0.005 mg/ml) for 10 min and then with propidium iodide (PI) (0.001 mg/ml) for 45 min at 37 °C. Next, the neurons were rinsed with culture medium and imaged with an inverted fluorescence microscope (Olympus) with 6–8 randomly chosen fields (200 ×) per well. The data are from a single experiment (three wells) performed in triplicate.
Morris water maze
The Morris water maze was used to examine hippocampus-dependent behaviors [29]. In brief, the rats were trained for five consecutive days (4 trials a day). On the sixth day, the hidden platform was removed to administer a probe test for 60 s. The training and test were video recorded and analyzed by ANY-maze software.
Contextual fear conditioning test
During training, the rats were allowed to freely explore a fear conditioning training box for 2 min. Then, five 2-s foot shocks (0.7 mA; constant current) with a 58-s interval were administered. After 24 h, the rats were placed in the same box for 8 min. The stiffness was recorded and reported as a percentage of the total observation time.
Modified neurological severity scores (mNSS)
Neurological dysfunction was evaluated by the modified Neurological Severity Score (mNSS) [42]. The mNSS assessment included motor tests (including forelimb flexion, hindlimb flexion, and head deviates, scored 0 to 3), reflexes absent and abnormal movements (scored 0 to 3), beam balance tests (scored 0 to 6), auditory reflex (scored 0 to 1), and corneal reflex (scored 0 to 1).
TTC staining
The 2-mm mouse brain slices were stained with 1% TTC (2,3,5-triphenyltetrazolium chloride) for 5 min and then fixed with 4% paraformaldehyde. The infarct area of each section was calculated by ImageJ software. The total infarct volume is the sum of the infarct volume of each layer (infarct area × thickness of each layer). The ischemic infarct volume was reported as (volume of contralateral hemisphere−volume of normal tissue of ipsilateral hemisphere)/volume of contralateral hemisphere × 100%.
Rotarod test
The rotarod test was used to evaluate sensorimotor functions [29]. Briefly, mice were trained on the three days prior to MCAO surgery with three trials per day (5 min training with 30 min internal). The program was set from 5 to 40 rpm/min with 7 rpm/min acceleration. Seven days after MCAO, mice were tested with the same program. The average falling time of three trials was reported.
Adhesive removal test
In habituation session, each mouse was allowed to explore for 15 min. The left and right paws of the mice were pasted with red circular patches (6-mm diameter) separately. The process of removing the patch was recorded for 2 min. The average time of three trials tearing off the left and right claw patches was used for statistical analysis.
Statistical analysis
All the data were statistically analyzed by GraphPad Prism 7.0 software. Immunoblot, immunostaining, and PI staining results are presented as the means ± standard deviations (SD), and data for behavioral studies, mNSS, TTC staining and Nissl staining are presented as the means ± standard errors (SE). Comparisons between two groups were performed by paired or unpaired Student’s t test. Comparisons among multiple groups in escape latency (Morris water maze test) and mNSS were carried out by two-way ANOVA, while other group comparisons were statistically made using one-way ANOVA followed by Dunnett’s t test. P < 0.05 was considered significant.
Results
MLK3 is modified by SUMO1, and cerebral ischemia increases MLK3 SUMOylation
To determine the changes in SUMOylation in cerebral ischemia models, SUMOylation of MLK3 (MLK3-SUMO1 conjugation) was first detected in the normal rat hippocampal CA1 subfield (Figure S1A–S1E). We then measured the conjugation of MLK3 with SUMO1 in OGD and transient global ischemia models (Fig. 1A). In the in vitro rat primary cortical neuron model, MLK3 SUMOylation increased significantly after OGD/R treatment, as shown by coimmunoprecipitation (Fig. 1B). In the in vivo rat global ischemia model (I/R), the level of MLK3-SUMO1 conjugation increased significantly six hours post-I/R in the hippocampal CA1 subfield (Fig. 1C), one of the areas most vulnerable to global ischemia. These data suggest that MLK3 is a novel target of SUMO1 and that brain ischemia promotes MLK3 SUMOylation.

Cerebral ischemia induces MLK3 SUMOylation, and K401 is the major SUMOylation site. A Schematic diagram of evaluating MLK3 SUMOylation in OGD and transient global ischemia models. B, C Time-dependent changes in MLK3 SUMOylation in the primary neuron OGD/R model (B) and in the CA1 region of the rat global I/R model (C) by co-IP and immunoblotting. Relative levels were normalized to the normoxia (B) or sham (C) group. Data are presented as the mean ± SD (n = 3–4). *P < 0.05, compared with the normoxia (B) or sham (C) group; one-way ANOVA with Dunnett’s t test. D Interaction of MLK3 with SUMO1 in HEK293T cells by co-IP and immunoblotting. E In vitro SUMOylation assay for the conjugation of MLK3(390–413) with SUMO1. F In vitro SUMOylation assay for SUMOylation of MLK3 at its K401 residue. G SUMOylation of full-length MLK3 WT or K401R mutant overexpressed in HEK293T cells
To delineate the conjugation of MLK3 with SUMO1, we first confirmed MLK3-SUMO1 conjugation in HEK293T cells coexpressing MLK3 with wild-type (WT) SUMO1 or an inactive SUMO1 mutant control. As shown in Fig. 1D and S1F–S1G, MLK3 interacted with WT SUMO1 but not with the SUMO1 mutant control. Next, we explored SUMOylation sites in MLK3. Sequence analysis using SUMOplot (http://www.abgent.com/sumoplot), JASSA [43], and GPS-SUMO [44] software predicted that a "WKRE" sequence (amino acids 400–403) of MLK3 was a conserved SUMOylation site, with sequence similarity with the consensus SUMOylation motif of ψKX(E/D) [13, 14] and surface localization in the MLK3 protein (Fig. S2A, B). To validate the predicted SUMOylation site, a Myc-tagged MLK3 peptide representing AAs 390–413 (corresponding to the "WKRE"-containing α-helix) [Myc-MLK3(390–413)] was synthesized chemically for an in vitro SUMOylation assay (Figs. 1E, S2C). A single SUMOylation band at ~ 28 kDa was detected in the presence of SUMO1(WT) but not in the presence of the SUMO1 mutant, indicating covalent conjugation of the MLK3(390–413) peptide with SUMO1. To verify whether K401 in the predicted "WKRE" sequence was the SUMOylation site, a mutated peptide, Myc-MLK3(390–413) K401R, was synthesized chemically for the in vitro SUMOylation assay. As shown in Fig. 1F, SUMOylation of the Myc-MLK3(390–413) peptide was abolished in the K401R mutation group. Moreover, the SUMOylation of full-length MLK3 in cells was significantly diminished by the K401R mutation (Fig. 1G). These findings reveal that the K401 residue was the primary site for MLK3-SUMO1 conjugation, although other SUMOylation sites for MLK3 could not be excluded.
The SUMO ligase PIAS3 is involved in brain ischemia-induced MLK3 SUMOylation
PIAS3 is a SUMO ligase that participates in the SUMOylation of target proteins to regulate neurological functions [15, 18, 21, 22]. To examine the role of PIAS3 in the SUMOylation of MLK3, the interaction between MLK3 and PIAS3 was first evaluated in a rat primary neuron OGD/R model and transient global ischemia model (Fig. 2A). As shown in Fig. 2B, OGD/R treatment promoted the interaction of MLK3 with PIAS3 in primary cortical neurons, with a time course corresponding to that of MLK3 SUMOylation. Similarly, the interaction between MLK3 and PIAS3 was significantly enhanced in the rat hippocampal CA1 region following I/R (Fig. 2C). To determine whether PIAS3 is involved in the SUMOylation of MLK3, coexpression experiments were performed using HEK293T cells. SUMOylation of MLK3 dramatically increased in the presence of PIAS3 (Fig. 2D).These data suggest that PIAS3 facilitates MLK3 SUMOylation following brain ischemia.

Cerebral ischemia increases the interaction of MLK3 with the SUMO ligase PIAS3, and the PINIT domain of PIAS3 directly binds MLK3. A Schematic diagram of assessing the MLK3-PIAS3 interaction in OGD and transient global ischemia models. B, C Time-dependent changes in the MLK3-PIAS3 interaction in the primary neuron OGD/R model (B) and in the CA1 region of the rat global I/R model (C) by co-IP and immunoblot analysis. Relative levels were normalized to the normoxia (B) or sham (C) group. Data are presented as the mean ± SD (n = 3). *P < 0.05, compared with the normoxia (B) or sham (C) group; one-way ANOVA with Dunnett’s t test. D The role of PIAS3 in MLK3 SUMOylation in HEK293T cells. E The structural pattern of Myc-tagged full-length rat PIAS3 (Myc-PIAS3 WT) and its domain-deleted mutants. F, G The interaction of MLK3 with Myc-PIAS3 WT or its domain-deleted mutants in HEK293T cells. H His pulldown assay for the interaction of MLK3 with His-PIAS3 WT or 133–320-Myc-His. I Disruption of the MLK3-PIAS3 interaction by 133–320 in HEK293T cells
The PINIT domain of PIAS3 directly binds MLK3
To identify the structural basis for the MLK3-PIAS3 interaction, recombinant constructs coding for Myc-tagged full-length rat PIAS3 (Myc-PIAS3 WT) and its truncated mutants representing AAs 1–500, 1–320, and 1–132 were generated individually (Fig. 2E) and then coexpressed with MLK3 in HEK293T cells. Co-IP experiments showed that interaction with MLK3 was detected in the full-length PIAS3 and its truncated 1–500 and 1–320 groups but not in the 1–132 group (Fig. 2F), suggesting that PIAS3 AAs 133–320, which correspond to the PINIT domain, interact with MLK3. The interaction between MLK3 and the PINIT domain was confirmed using both co-IP (Fig. 2G) and His pulldown assays (Fig. 2H). Moreover, overexpression of the PINIT domain of PIAS3 blocked the MLK3-PIAS3 interaction in a dose-dependent manner (Fig. 2I). These data indicate that MLK3 directly interacts with the PINIT domain of PIAS3 and that overexpression of the PINIT domain disrupts the interaction of MLK3 with PIAS3.
PIAS3 regulates the transcription activity of signal transducer and activator of transcription 3 (STAT3), which has been implicated in cerebral ischemia [45, 46]. In this study, OGD/R-induced STAT3-PIAS3 interaction was not blocked by Tat-PINIT (Fig. S3), which further proves the specific inhibitory effect of Tat-PINIT on MLK3–PIAS3 interaction.
Disrupting MLK3 SUMOylation with the PINIT domain of PIAS3 reduces neuronal injury in the OGD/R model
Reportedly, MLK3 phosphorylation (activation) peaks at six hours post-I/R, which is consistent with elevated SUMOylation [11, 12]. To elucidate the effects of SUMOylation on the activation of MLK3 and its downstream apoptotic cascades, wild-type MLK3 or its K401R mutant were coexpressed with SUMO1 in HEK293T cells. As shown in Fig. S4, compared to the MLK3 single transfection group, coexpression of MLK3 with SUMO1 facilitated the activation of MLK3 (phosphorylation at T277/S281 sites), increased cleaved caspase-3 levels, increased cell apoptosis, and promoted the phosphorylation (activation) of p38 and JNKs. Moreover, the SUMOylation-deficient K401R mutant had no effect. These results indicate that MLK3 SUMOylation at K401 is vital in regulating its activation and downstream apoptotic signaling.
Next, we explored whether the PINIT domain of PIAS3 interrupts SUMOylation of endogenous MLK3 by PIAS3 in neurons and its effects on neuronal injury in the OGD/R model (Fig. 3A). Primary cortical neurons at DIV 9 were infected with either lenti-PINIT or lenti-NC. At DIV 13, approximately 80% of the neurons expressed EGFP (Fig. 3B–D). As shown in Fig. 3E, overexpression of the PINIT domain blocked the OGD/R-induced MLK3-PIAS3 interaction and SUMOylation of endogenous MLK3 in primary cortical neurons, whereas lenti-NC had no effect. The overexpressed PINIT domain inhibited MLK3, p38, and JNKs phosphorylation (activation) and cleaved caspase-3 expression, which was unaffected by lenti-NC (Fig. 3F–G). These results indicate that overexpression of the PINIT domain intervened in the MLK3–PIAS3 interaction, inhibited MLK3 SUMOylation and enzymatic activity, and ultimately restrained the downstream apoptotic p38/JNKs pathway after OGD/R.

Overexpressing the PINIT domain of PIAS3 blocks the MLK3-PIAS3 interaction and inhibits MLK3 SUMOylation and downstream MLK3-JNK/p38 apoptotic cascade activation after OGD/R. A Schematic diagram of blocking PIAS3-mediated MLK3 SUMOylation by the PINIT domain in the OGD model. B Diagram of lentiviral plasmids expressing the PINIT domain and negative control. C Lentivirus infection efficiency (green) in primary cortical neurons. Bar = 50 μm. D Schematic diagram of the overexpression of the PINIT domain in the primary neuron OGD model. E–G The effects of overexpressing the PINIT domain on the MLK3-PIAS3 interaction, MLK3 SUMOylation (E), MLK3 activation/phosphorylation (F), and downstream JNK/p38 apoptotic signaling (G) after OGD/R. − , without lentivirus infection. Relative levels were normalized to the normoxia group. Data are presented as the mean ± SD (n = 3–4). *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001; ns nonsignificant; one-way ANOVA with Dunnett’s t test
To further examine the protective role of the PINIT domain in OGD/R-induced injury, cortical neurons were pretreated with the penetrating peptide Tat-tagged PINIT (Tat-PINIT) or the negative control (Tat-EGFP) for three hours and then subjected to OGD/R treatment (Fig. 4A). As shown in Fig. 4B, the average dendrite length was shortened after OGD/R, and this change was rescued by preincubation with Tat-PINIT but not with Tat-EGFP. Additionally, Tat-PINIT reduced OGD/R-induced apoptosis (Fig. 4C). Moreover, lower level of OGD/R-induced dendrite injury and neuronal apoptosis were observed in the Lenti-MLK3 K401R mutant group compared with the WT group (Fig. S5). Together, these results indicated that Tat-PINIT has neuroprotective effects against OGD/R-induced neuronal damage.

Overexpressing the PINIT domain of PIAS3 alleviates neuronal injury after OGD/R. A Schematic diagram of the effect of the penetrating peptide Tat-tagged PINIT (Tat-PINIT) or Tat-EGFP (negative control) on neuronal injury in the primary neuron OGD model. B, C The effects of overexpressing the PINIT domain (5 μmol/L) on OGD/R-induced neuronal dendrite injury (B) and cell apoptosis (C). Tat-PINIT was checked by IF with an anti-Myc antibody. The average length of dendrites is presented as the total dendritic length (MAP2) divided by the number of cells. − , without Tat treatment. Bar = 50 μm. Data are presented as the mean ± SD (n = 3–4). **P < 0.01; ****P < 0.0001; ns, nonsignificant; one-way ANOVA with Dunnett’s t test
Interrupting MLK3 SUMOylation with the PINIT domain of PIAS3 reduces neuronal injury and relieves learning and memory defects after I/R
Next, we evaluated the neuroprotective effect of the PINIT domain of PIAS3 against transient global ischemia-induced neuronal injury. The recombinant AAV encoding the PINIT domain of PIAS3 (AAV-PINIT) and negative control empty vector AAV (AAV-NC) were injected into the bilateral rat hippocampal CA1 region, an area vulnerable to global ischemia, with two points in each hemisphere (Fig. 5A–C). After five weeks, broad expression of AAV-PINIT was observed in the hippocampal CA1 region (Fig. 5D). The rats were subjected to 15 min of ischemia followed by six hours of reperfusion, and the hippocampal CA1 subfield was extracted to assess the MLK3-PIAS3 interaction, MLK3 SUMOylation, and downstream apoptosis signaling (Fig. 5E). As shown in Fig. 5F–H, overexpressing PINIT interrupted the interaction between MLK3 and PIAS3, inhibited MLK3 SUMOylation, reduced the phosphorylation of MLK3, p-38 and JNKs, and decreased cleaved caspase-3 levels. These data reveal that the PINIT domain interferes with the MLK3-PIAS3 interaction, blocks MLK3 SUMOylation and catalytic activity, and decreases downstream p38/JNK apoptotic cascades after I/R.

Overexpressing the PINIT domain of PIAS3 inhibits global cerebral ischemia-induced MLK3 SUMOylation and downstream apoptotic cascade stimulation in the hippocampal CA1 subfield. A Schematic diagram of the PINIT domain blocking PIAS3-mediated MLK3 SUMOylation in a rat global ischemia model. B Diagram of AAVs expressing the PINIT domain and negative control. C Schematic representation of AAV injection into the rat bilateral hippocampal CA1 subfield. D AAV infection (green) in the hippocampal CA1 region. Bar = 50 μm. E Schematic diagram of AAV recombinant overexpression in a rat global ischemia model. F–H The effects of PINIT overexpression on the MLK3-PIAS3 interaction, MLK3 SUMOylation (F), MLK3 activation/phosphorylation (G) and downstream apoptotic cascade activation (H) after I/R. − , without AAV injection. Relative levels were normalized to those of the sham group. Data are presented as the mean ± SD (n = 3–4). *P < 0.05; **P < 0.01; ****P < 0.0001; one-way ANOVA with Dunnett’s t test
The hippocampus CA1 subfield is one of the regions most vulnerable to global ischemia, and is involved in spatial learning and memory. Therefore, we next examined the effects of PINIT overexpression on spatial learning and memory impairments post-brain ischemia. After injection of AAV-PINIT or AAV-NC for five weeks, the rats were subjected to the Morris water maze test and contextual fear conditioning 7 and 13 days post-I/R, respectively (Fig. 6A). During the five days of training in the Morris water maze, the navigation paths were recorded and analyzed (Fig. 6B). I/R rats presented a longer escape latency, which was shortened by treatment with AAV-PINIT but not with AAV-NC (Fig. 6C). On the last day of training, there was no significant difference in the mean swimming speed between the groups (Fig. 6D). Treatment with AAV-PINIT, but not AAV-NC, rescued the I/R-induced reduction in the percentage of time spent in the target quadrant and the number of platform crossovers (Fig. 6E, F). In the contextual fear conditioning test, freezing time was significantly reduced in the I/R-treated rats. AAV-PINIT increased the I/R-induced freezing time, whereas AAV-NC had no effect (Fig. 6G). These results demonstrate that overexpressing the PINIT domain of PIAS3 rescues hippocampus-dependent spatial learning and memory defects in global ischemia.

Overexpressing the PINIT domain of PIAS3 relieves learning and memory defects in rats after global cerebral ischemia. A Schematic diagram of AAV recombinant overexpression in a rat global ischemia model. B–F Effects of overexpressing the PINIT domain on spatial memory (Morris water maze) in a rat I/R model. B Representative navigation traces on Day 7 in the Morris water maze test. C Escape latency over the five-day training session. D Mean swimming speed. E Percentage of time spent in the target quadrant. F Number of platform crossovers. G Effects of overexpressing the PINIT domain on contextual fear conditioning in a rat I/R model. Data are presented as the mean ± SEM (n = 10). *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001; ns, nonsignificant. Two-way ANOVA with Dunnett’s t test (C), one-way ANOVA with Dunnett’s t test (D–G). − , without AAV injection
Overexpressing the PINIT domain of PIAS3 reduces brain infarction and ameliorates sensorimotor function defects after MCAO/R
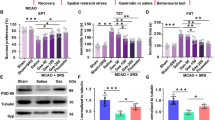
To determine whether the PINIT domain also has protective effects in focal ischemia, a mouse MCAO model was examined. Similar to the global ischemia model, MLK3-SUMO1 conjugation and MLK3-PIAS3 interaction in the ischemic hemisphere (ipsilateral or Ipsi) increased significantly after MCAO/R compared to those in the nonischemic hemisphere (contralateral or Contra) in the MCAO model (Fig. 7A–C). To determine the effects of the PINIT domain on brain infarction and neurological damage, AAV-PINIT or AAV-NC was stereotactically injected into the cortical regions susceptible to brain injury following MCAO (Fig. 7D–F). Widespread expression was observed five weeks after the injection (Fig. 7G). AAV-PINIT decreased the modified neurological severity score (mNSS) in the MCAO-treated mice after 1, 3, and 7 days of reperfusion compared to that of the AAV-NC group (Fig. 7H). The infarct volume in the ipsilateral cerebral regions was reduced by AAV-PINIT treatment after MCAO/R at 1 (Fig. 7I) and 7 days (Figure S6). Behavioral tests were performed to evaluate the effects of AAV-PINIT on sensorimotor functions in the MCAO-treated mice. In the rotarod test, PINIT overexpression increased the time spent on the rod after MCAO/R, which was not affected before MCAO (Fig. 7J). In the adhesive removal test, compared to the AAV-NC-treated group, the AAV-PINIT group showed a reduced time to remove adhesive tape from the left paw (affected side) and presented no difference in removing tape from the right paw (unaffected side) after MCAO/R (Fig. 7K). These data indicate that the overexpression of the PINIT domain of PIAS3 alleviates neuronal lesions and relieves sensorimotor function deficits in MCAO-treated mice.

Overexpressing the PINIT domain of PIAS3 reduces brain infarction and rescues motion function defects in mice after focal ischemia. A Schematic diagram of the mouse MCAO focal ischemia model. B, C Changes in MLK3 SUMOylation (B) and MLK3-PIAS3 interaction (C) in ipsilateral ischemic tissue (Ipsi) and contralateral normal tissue (Contra) in the focal ischemia model. Relative levels were normalized to those of the Contra groups. Data are presented as the mean ± SD (n = 3). *P < 0.05; **P < 0.01; paired t test. D–E Schematic diagram of the effect of overexpressing the PINIT domain on PIAS3-mediated MLK3 SUMOylation and reducing neurological defects in a mouse MCAO model. F Schematic representation of AAV injection into the mouse right cortex. G AAV expression (green) in the mouse cortex five weeks after injection. Bar = 500 μm. H–K Effects of overexpressing the PINIT domain on mNSS (H), brain infarction (I), and sensorimotor behaviors assessed by the rotarod test (J) and adhesive removal test (K). Data are presented as the mean ± SEM (n = 10). *P < 0.05; ***P < 0.001; ****P < 0.0001; ns nonsignificant. Two-way ANOVA with the Sidak method (H) and unpaired t test (I–K)
Discussion
Here, the role of MLK3 SUMOylation in ischemic stroke was assessed. Our data identified MLK3 as a target of SUMO1 and provided an additional modification mechanism for regulating MLK3 enzymatic activity. The results showed that MLK3 SUMOylation plays a pivotal role in ischemic neuronal lesions. Blocking the MLK3-PIAS3 interaction inhibits MLK3 SUMOylation and stimulation, alleviates neuronal injury and cognitive impairment, and plays a neuroprotective role after cerebral ischemia.
Disrupted SUMOylation homeostasis was observed in the penumbra tissue of stroke patients, global and focal cerebral ischemic animal models, and in vitro OGD models [32,33,34,35]. Some studies suggest that SUMOylation is an endogenous neuroprotective response after cerebral ischemia [38,39,40, 47]. However, other findings show the opposite. For instance, focal cerebral ischemia increases the expression of SENP1 (sentrin/SUMO-specific protease 1), which then decreases global SUMOylation levels, relieves neural damage, and exerts a protective effect [36]. In addition, SUMOylation of the kainate receptor subunit GluK2 activates the MLK3-JNK3 apoptotic signaling pathway and participates in ischemic neuronal lesions [37]. Based on the above findings, we speculate that cerebral ischemia induces the conjugation of SUMOs with different targets, in which some SUMOylated substrates exert protective effects and others exert damaging effects. Here, we found that MLK3 is a new SUMO1 target and that MLK3 SUMOylation contributes to ischemic cognitive impairment. Clarifying the mechanism for MLK3 SUMOylation will help identify intervention targets and strategies for the treatment of ischemic stroke.
Post-translational modification is recognized as an important way to regulate protein structure and function. Reportedly, MLK3 activity is positively regulated by both autophosphorylation and upstream kinase-mediated phosphorylation at threonine 277 and serine 281 residues within the kinase domain [4], which plays an important role in cell death, contributing to the pathogenesis of ischemic brain damage, Alzheimer’s disease, and oxidative stress injury [10, 37, 48,49,50]. Here, we found that SUMOylation deficiency at K401 led to decreased phosphorylation of MLK3(T277/S281), revealing crosstalk between SUMOylation and phosphorylation. We deduced that SUMO1 conjugation at K401 increases MLK3(T277/S281) phosphorylation, which in turn potentiates MLK3 catalytic activity. Moreover, we found that SUMOylation stimulates MLK3-p38/JNK apoptotic cascades and ultimately leads to cell apoptosis. Therefore, MLK3 SUMOylation not only participates in ischemic cerebral damage but may also be involved in other pathogenesis mediated by MLK3 activation, such as Alzheimer’s disease and oxidative stress injury.
In SUMOylation cascades, SUMO-activating enzymes (SAE1/2) and SUMO-specific conjugating enzymes (Ubc9) are conserved and unique, whereas SUMO ligases are substrate selective. SUMO ligases include the family of protein inhibitors of activated STAT proteins (PIASs), nuclear pore proteins RanBP2 and Pc2 [23, 24]. The PIASs family is localized in neurons and participates in synaptic plasticity and cognitive function regulation [15, 18, 21, 22, 41, 51,52,53]. Our results reveal that cerebral ischemia induces MLK3 SUMOylation in the hippocampus and cortex by promoting the interaction of the SUMO ligase PIAS3 and that AAs 133–320 in PIAS3 directly bind MLK3. AAs 133–320 correspond to the PINIT domain of PIAS3, which has 75% homology with PIAS1, 68% homology with PIAS2, and 50% homology with PIAS4. Therefore, whether PIAS1/2/4 plays a role in MLK3 SUMOylation still needs to be elucidated. Moreover, overexpressing the PINIT domain of PIAS3 does not completely block the MLK3-PIAS3 interaction in vivo and in vitro. This finding suggests that other binding regions in PIAS3 cannot be ruled out.
Here, we show that MLK3 SUMOylation leads to MLK3-p38/JNK apoptotic cascade stimulation, neuronal lesions, and cognitive impairment. Overexpression of PIAS3 (133–320) by lentivirus or AAV recombinants interrupted the MLK3-PIAS3 interaction in vivo and in vitro, inhibited MLK3 and downstream apoptotic cascade activation, effectively rescued neuronal lesions, and ameliorated cognitive defects. The PINIT domain (AAs 133–320) of PIAS3 is highly conserved in rats, mice, and humans. Therefore, MLK3-PIAS3 is a potential interruption target for ischemic stroke, and the PINIT peptide is a promising therapeutic biopharmaceutical. Moreover, Tat-PINIT relieves OGD/R-induced dendrite injury and cell apoptosis and presents lower cytotoxicity than Lenti-PINIT. This finding reveals that purified protein may be more suitable for disease treatment. The PINIT domain of PIAS3 contains 188 amino acids, which is a relatively long fragment. Shorter peptides and small molecular compounds need to be identified to be more applicable neuroprotective drugs for ischemic stroke therapy.
In conclusion, our data provide the first evidence that reversible MLK3 SUMOylation is involved in neuronal lesions and cognitive impairment after cerebral ischemia. MLK3-PIAS3 is a potential intervention target for ischemic stroke therapy, and PINIT intervention is a promising neuroprotective strategy.
Data availability
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Abbreviations
- AAV-NC:
-
AAVs negative control
- AAV-PINIT:
-
AAVs expressing the PINIT domain of PIAS3
- Co-IP:
-
Co-immunoprecipitation
- Contra:
-
Contralateral
- His-PIAS3 WT:
-
His-tagged wild-type PIAS3
- IB:
-
Immunoblot
- IP:
-
Immunoprecipitation
- Ipsi:
-
Ipsilateral
- I/R:
-
Reperfusion after global ischemia
- JNK:
-
C-Jun N-terminal kinase
- Lenti-NC:
-
Lentivirus recombinants negative control
- Lenti-PINIT:
-
Lentivirus recombinants containing the PINIT domain of PIAS3
- MAPKKK:
-
Mitogen-activated protein kinase kinase kinase
- MCAO:
-
Middle cerebral artery occlusion
- Myc-PIAS3 WT:
-
Myc-tagged wild-type PIAS3
- MKKs:
-
MAPK kinases
- MLK3 K401R:
-
MLK3 lysine 401 to arginine
- MLK3:
-
Mixed-lineage kinase 3
- mNSS:
-
Modified Neurological Severity Score
- OGD:
-
Oxygen and glucose deprivation
- PI:
-
Propidium iodide
- SUMOs:
-
Small ubiquitin-like modifiers
- Tat-EGFP:
-
Tat-tagged EGFP
- Tat-PINIT:
-
Tat-tagged PINIT domain of PIAS3
References
Feske SK (2021) Ischemic stroke. Am J Med 134:1457–1464. https://doi.org/10.1016/j.amjmed.2021.07.027
Rabinstein AA (2020) Update on treatment of acute ischemic stroke. Continuum (Minneap Minn) 26:268–286. https://doi.org/10.1212/CON.0000000000000840
Herpich F, Rincon F (2020) Management of acute ischemic stroke. Crit Care Med 48:1654–1663. https://doi.org/10.1097/CCM.0000000000004597
Gallo KA, Johnson GL (2002) Mixed-lineage kinase control of JNK and p38 MAPK pathways. Nat Rev Mol Cell Biol 3:663–672. https://doi.org/10.1038/nrm906
Handley ME, Rasaiyaah J, Chain BM, Katz DR (2007) Mixed lineage kinases (MLKs): a role in dendritic cells, inflammation and immunity? Int J Exp Pathol 88:111–126. https://doi.org/10.1111/j.1365-2613.2007.00531.x
Yu H, Song L, Cao X, Li W, Zhao Y, Chen J, Li J, Chen Y, Yu W, Xu Y (2020) Hederagenin attenuates cerebral ischaemia/reperfusion injury by regulating MLK3 signalling. Front Pharmacol 11:1173. https://doi.org/10.3389/fphar.2020.01173
Ha AT, Cho JY, Kim D (2022) MLK3 regulates inflammatory response via activation of AP-1 pathway in HEK293 and RAW264.7 cells. Int J Mol Sci 23:10874. https://doi.org/10.3390/ijms231810874
Wang J, Deng B, Liu Q, Huang Y, Chen W, Li J, Zhou Z, Zhang L, Liang B, He J, Chen Z, Yan C, Yang Z, Xian S, Wang L (2020) Pyroptosis and ferroptosis induced by mixed lineage kinase 3 (MLK3) signaling in cardiomyocytes are essential for myocardial fibrosis in response to pressure overload. Cell Death Dis 11:574. https://doi.org/10.1038/s41419-020-02777-3
Schroyer AL, Stimes NW, Abi Saab WF, Chadee DN (2018) MLK3 phosphorylation by ERK1/2 is required for oxidative stress-induced invasion of colorectal cancer cells. Oncogene 37:1031–1040. https://doi.org/10.1038/onc.2017.396
Song YJ, Dai CX, Li M, Cui MM, Ding X, Zhao XF, Wang CL, Li ZL, Guo MY, Fu YY, Wen XR, Qi DS, Wang YL (2019) The potential role of HO-1 in regulating the MLK3-MKK7-JNK3 module scaffolded by JIP1 during cerebral ischemia/reperfusion in rats. Behav Brain Res 359:528–535. https://doi.org/10.1016/j.bbr.2018.11.003
Hu SQ, Ye JS, Zong YY, Sun CC, Liu DH, Wu YP, Song T, Zhang GY (2012) S-nitrosylation of mixed lineage kinase 3 contributes to its activation after cerebral ischemia. J Biol Chem 287:2364–2377. https://doi.org/10.1074/jbc.M111.227124
Chen J, Li C, Pei DS, Han D, Liu XM, Jiang HX, Wang XT, Guan QH, Wen XR, Hou XY, Zhang GY (2009) GluR6-containing KA receptor mediates the activation of p38 MAP kinase in rat hippocampal CA1 region during brain ischemia injury. Hippocampus 19:79–89. https://doi.org/10.1002/hipo.20479
Creton S, Jentsch S (2010) SnapShot: the SUMO system. Cell 143:848-848.e1. https://doi.org/10.1016/j.cell.2010.11.026
Wilkinson KA, Nakamura Y, Henley JM (2010) Targets and consequences of protein SUMOylation in neurons. Brain Res Rev 64:195–212. https://doi.org/10.1016/j.brainresrev.2010.04.002
Martin S, Nishimune A, Mellor JR, Henley JM (2007) SUMOylation regulates kainate-receptor-mediated synaptic transmission. Nature 447:321–325. https://doi.org/10.1038/nature05736
Chamberlain SE, González-González IM, Wilkinson KA, Konopacki FA, Kantamneni S, Henley JM, Mellor JR (2012) SUMOylation and phosphorylation of GluK2 regulate kainate receptor trafficking and synaptic plasticity. Nat Neurosci 15:845–852. https://doi.org/10.1038/nn.3089
Plant LD, Dowdell EJ, Dementieva IS, Marks JD, Goldstein SA (2011) SUMO modification of cell surface Kv2.1 potassium channels regulates the activity of rat hippocampal neurons. J Gen Physiol 137:441–454. https://doi.org/10.1085/jgp.201110604
Ghosh H, Auguadri L, Battaglia S, Simone Thirouin Z, Zemoura K, Messner S, Acuña MA, Wildner H, Yévenes GE, Dieter A, Kawasaki H, Hottiger OM, Zeilhofer HU, Fritschy JM, Tyagarajan SK, (2016) Several posttranslational modifications act in concert to regulate gephyrin scaffolding and GABAergic transmission. Nat Commun 7:13365. https://doi.org/10.1038/ncomms13365
Craig TJ, Anderson D, Evans AJ, Girach F, Henley JM (2015) SUMOylation of syntaxin1A regulates presynaptic endocytosis. Sci Rep 5:17669. https://doi.org/10.1038/srep17669
Nair RR, Patil S, Tiron A, Kanhema T, Panja D, Schiro L, Parobczak K, Wilczynski G, Bramham CR (2017) Dynamic arc SUMOylation and selective interaction with F-Actin-binding protein Drebrin A in LTP consolidation in Vivo. Front Synaptic Neurosci 9:8. https://doi.org/10.3389/fnsyn.2017.00008
Du CP, Wang M, Geng C, Hu B, Meng L, Xu Y, Cheng B, Wang N, Zhu QJ, Hou XY (2020) Activity-induced SUMOylation of neuronal nitric oxide synthase is associated with plasticity of synaptic transmission and extracellular signal-regulated kinase 1/2 signaling. Antioxid Redox Signal 32:18–34. https://doi.org/10.1089/ars.2018.7669
Meng L, Du CP, Lu CY, Zhang K, Li L, Yan JZ, Hou XY (2021) Neuronal activity-induced SUMOylation of Akt1 by PIAS3 is required for long-term potentiation of synaptic transmission. FASEB J 35:e21769. https://doi.org/10.1096/fj.202002728R
Henley JM, Craig TJ, Wilkinson KA (2014) Neuronal SUMOylation: mechanisms, physiology, and roles in neuronal dysfunction. Physiol Rev 94:1249–1285. https://doi.org/10.1152/physrev.00008.2014
Krumova P, Weishaupt JH (2013) Sumoylation in neurodegenerative diseases. Cell Mol Life Sci 70:2123–2138. https://doi.org/10.1007/s00018-012-1158-3
Eckermann K (2013) SUMO and Parkinson’s disease. Neuromol Med 15:737–759. https://doi.org/10.1007/s12017-013-8259-5
Morozko EL, Smith-Geater C, Monteys AM, Pradhan S, Lim RG, Langfelder P, Kachemov M, Kulkarni JA, Zaifman J, Hill A, Stocksdale JT, Cullis PR, Wu J, Ochaba J, Miramontes R, Chakraborty A, Hazra TK, Lau A, St-Cyr S, Orellana I, Kopan L, Wang KQ, Yeung S, Leavitt BR, Reidling JC, Yang XW, Steffan JS, Davidson BL, Sarkar PS, Thompson LM (2021) PIAS1 modulates striatal transcription, DNA damage repair, and SUMOylation with relevance to Huntington’s disease. Proc Natl Acad Sci U S A 118:e2021836118. https://doi.org/10.1073/pnas.2021836118
O’Rourke JG, Gareau JR, Ochaba J, Song W, Raskó T, Reverter D, Lee J, Monteys AM, Pallos J, Mee L, Vashishtha M, Apostol BL, Nicholson TP, Illes K, Zhu YZ, Dasso M, Bates GP, Difiglia M, Davidson B, Wanker EE, Marsh JL, Lima CD, Steffan JS, Thompson LM (2013) SUMO-2 and PIAS1 modulate insoluble mutant huntingtin protein accumulation. Cell Rep 4:362–375. https://doi.org/10.1016/j.celrep.2013.06.034
Küçükali CI, Salman B, Yüceer H, Ulusoy C, Abacı N, Ekmekci SS, Tüzün E, Bilgiç B, Hanağası HA (2020) Small ubiquitin-related modifier (SUMO) 3 and SUMO4 gene polymorphisms in Parkinson’s disease. Neurol Res 42:451–457. https://doi.org/10.1080/01616412.2020.1724464
Xia Q, Mao M, Zeng Z, Luo Z, Zhao Y, Shi J, Li X (2021) Inhibition of SENP6 restrains cerebral ischemia-reperfusion injury by regulating Annexin-A1 nuclear translocation-associated neuronal apoptosis. Theranostics 11:7450–7470. https://doi.org/10.7150/thno.60277
Lee L, Sakurai M, Matsuzaki S, Arancio O, Fraser P (2013) SUMO and Alzheimer’s disease. Neuromol Med 15:720–736. https://doi.org/10.1007/s12017-013-8257-7
Bernstock JD, Ye DG, Griffin A, Lee YJ, Lynch J, Latour LL, Friedman GK, Maric D, Hallenbeck JM (2018) Cerebral ischemia increases small ubiquitin-like modifier conjugation within human penumbral tissue: radiological-pathological correlation. Front Neurol 8:738. https://doi.org/10.3389/fneur.2017.00738
Anderson DB, Zanella CA, Henley JM, Cimarosti H (2017) Sumoylation: implications for neurodegenerative diseases. Adv Exp Med Biol 963:261–281. https://doi.org/10.1007/978-3-319-50044-7_16
Yang W, Sheng H, Warner DS, Paschen W (2008) Transient global cerebral ischemia induces a massive increase in protein sumoylation. J Cereb Blood Flow Metabol 28:269–279. https://doi.org/10.1038/sj.jcbfm.9600523
Yang W, Sheng H, Warner DS, Paschen W (2008) Transient focal cerebral ischemia induces a dramatic activation of small ubiquitin-like modifier conjugation. J Cereb Blood Flow Metabol 28:892–896. https://doi.org/10.1038/sj.jcbfm.9600601
Silveirinha V, Stephens GJ, Cimarosti H (2013) Molecular targets underlying SUMO-mediated neuroprotection in brain ischemia. J Neurochem 127:580–591. https://doi.org/10.1111/jnc.12347
Zhang H, Wang Y, Zhu A, Huang D, Deng S, Cheng J, Zhu MX, Li Y (2016) SUMO-specific protease 1 protects neurons from apoptotic death during transient brain ischemia/reperfusion. Cell Death Dis 7:e2484. https://doi.org/10.1038/cddis.2016.290
Zhu QJ, Xu Y, Du CP, Hou XY (2012) SUMOylation of the kainate receptor subunit GluK2 contributes to the activation of the MLK3-JNK3 pathway following kainate stimulation. FEBS Lett 586:1259–1264. https://doi.org/10.1016/j.febslet.2012.03.048
Cuomo O, Pignataro G, Sirabella R, Molinaro P, Anzilotti S, Scorziello A, Sisalli MJ, Di Renzo G, Annunziato L (2016) Sumoylation of LYS590 of NCX3 f-loop by SUMO1 participates in brain neuroprotection induced by ischemic preconditioning. Stroke 47:1085–1093. https://doi.org/10.1161/STROKEAHA.115.012514
Lee YJ, Hallenbeck JM (2013) SUMO and ischemic tolerance. Neuromol Med 15:771–781. https://doi.org/10.1007/s12017-013-8239-9
Ma R, Ma L, Weng W, Wang Y, Liu H, Guo R, Gao Y, Tu J, Xu TL, Cheng J, Zhu MX, Zhou A, Li Y (2020) DUSP6 SUMOylation protects cells from oxidative damage via direct regulation of Drp1 dephosphorylation. Sci Adv 6:eaaz0361. https://doi.org/10.1126/sciadv.aaz0361
Thirouin ZS, Figueiredo M, Hleihil M, Gill R, Bosshard G, McKinney RA, Tyagarajan SK (2022) Trophic factor BDNF inhibits GABAergic signaling by facilitating dendritic enrichment of SUMO E3 ligase PIAS3 and altering gephyrin scaffold. J Biol Chem 298:101840. https://doi.org/10.1016/j.jbc.2022.101840
Chen J, Li Y, Wang L, Zhang Z, Lu D, Lu M, Chopp M (2001) Therapeutic benefit of intravenous administration of bone marrow stromal cells after cerebral ischemia in rats. Stroke 32:1005–1011. https://doi.org/10.1161/01.str.32.4.1005
Beauclair G, Bridier-Nahmias A, Zagury JF, Saïb A, Zamborlini A (2015) JASSA: a comprehensive tool for prediction of SUMOylation sites and SIMs. Bioinformatics 31:3483–3491. https://doi.org/10.1093/bioinformatics/btv403
Zhao Q, Xie Y, Zheng Y, Jiang S, Liu W, Mu W, Liu Z, Zhao Y, Xue Y, Ren J (2014) GPS-SUMO: a tool for the prediction of sumoylation sites and SUMO–interaction motifs. Nucleic Acids Res 42:W325-330. https://doi.org/10.1093/nar/gku383
Chung CD, Liao J, Liu B, Rao X, Jay P, Berta P, Shuai K (1997) Specific inhibition of Stat3 signal transduction by PIAS3. Science 278:1803–1805. https://doi.org/10.1126/science.278.5344.1803
Suzuki S, Tanaka K, Nogawa S, Dembo T, Kosakai A, Fukuuchi Y (2001) Phosphorylation of signal transducer and activator of transcription-3 (Stat3) after focal cerebral ischemia in rats. Exp Neurol 170:63–71. https://doi.org/10.1006/exnr.2001.770
Li X, Xia Q, Mao M, Zhou H, Zheng L, Wang Y, Zeng Z, Yan L, Zhao Y, Shi J (2021) Annexin-A1 SUMOylation regulates microglial polarization after cerebral ischemia by modulating IKKα stability via selective autophagy. Sci Adv 7:eabc5539. https://doi.org/10.1126/sciadv.abc5539
Wang S, Zhang T, Yang Z, Lin J, Cai B, Ke Q, Lan W, Shi J, Wu S, Lin W (2017) Heme oxygenase-1 protects spinal cord neurons from hydrogen peroxide-induced apoptosis via suppression of Cdc42/MLK3/MKK7/JNK3 signaling. Apoptosis 22:449–462. https://doi.org/10.1007/s10495-016-1329-z
Zhang QG, Tian H, Li HC, Zhang GY (2006) Antioxidant N-acetylcysteine inhibits the activation of JNK3 mediated by the GluR6-PSD95-MLK3 signaling module during cerebral ischemia in rat hippocampus. Neurosci Lett 408:159–164. https://doi.org/10.1016/j.neulet.2006.07.007
Li H, Luo XB, Xu Y, Hou XY (2019) A brief ischemic postconditioning protects against amyloid-β peptide neurotoxicity by downregulating MLK3-MKK3/6-P38MAPK signal in rat hippocampus. J Alzheimers Dis 71:671–684. https://doi.org/10.3233/JAD-190207
Liu SY, Ma YL, Hsu WL, Chiou HY, Lee EHY (2019) Protein inhibitor of activated STAT1 Ser503 phosphorylation-mediated Elk-1 SUMOylation promotes neuronal survival in APP/PS1 mice. Br J Pharmacol 176:1793–1810. https://doi.org/10.1111/bph.14656
Chen YC, Hsu WL, Ma YL, Tai DJ, Lee EH (2014) CREB SUMOylation by the E3 ligase PIAS1 enhances spatial memory. J Neurosci 34:9574–9589. https://doi.org/10.1523/JNEUROSCI.4302-13.2014
Magalhaes J, Tresse E, Ejlerskov P, Hu E, Liu Y, Marin A, Montalant A, Satriano L, Rundsten CF, Carlsen EMM, Rydbirk R, Sharifi-Zarchi A, Andersen JB, Aznar S, Brudek T, Khodosevich K, Prinz M, Perrier JM, Sharma M, Gasser T, Issazadeh-Navikas S (2021) PIAS2-mediated blockade of IFN-β signaling: a basis for sporadic Parkinson disease dementia. Mol Psychiatry 26:6083–6099. https://doi.org/10.1038/s41380-021-01207-w
Acknowledgements
The authors would like to acknowledge Xiaoyu Hou, Xuecheng Qiu, Jingzhen Li, Jing Liu and Jianfeng Wei for their suggestions and comments on experimental design and data presentation, and Xinwang Wang and Haisu Yan for their preliminary research exploration. The authors also would like to acknowledge Xiaohui Yin for kindly sharing reagent for Nissl staining.
Funding
This work was supported by grants from the National Natural Science Foundation of China (32100769 and 82371401 to M.L. and 81100852 to C.-P.D.), the Natural Science Foundation of the Jiangsu Higher Education Institutions (20KJA310010 to C.-P.D.), and Xuzhou Medical University (D2020054 and JBGS202202 to M.L.)
Author information
Authors and Affiliations
Contributions
Yu Jiang, Lulu Hu, and Baixue Wang performed the experiments and analyzed data. Bingge Zhang assisted with mice MCAO model preparation. Mengwen Shao assisted with Nissl staining. Li Meng performed cells apoptosis experiments; Yan Xu assisted with primary neuron culture; Rourou Chen assisted with protein purification; Meng Li and Caiping Du designed research, wrote the paper, and provided supervision and funding.
Corresponding authors
Ethics declarations
Conflict of interests
The authors declare no competing interests. The authors have no relevant financial or non-financial interests to disclose.
Ethics approval
All experiments involving animals were complied with the guidelines described in the revised 2011 Chinese Regulations for the Administration of Affairs Concerning Experimental Animals and approved by the Institutional Animal Care and Use Committee of Xuzhou Medical University.
Consent for publication
The authors consent to publishing this work.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jiang, Y., Hu, L., Wang, B. et al. Disrupting PIAS3-mediated SUMOylation of MLK3 ameliorates poststroke neuronal damage and deficits in cognitive and sensorimotor behaviors. Cell. Mol. Life Sci. 81, 119 (2024). https://doi.org/10.1007/s00018-024-05166-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00018-024-05166-7