
Abstract
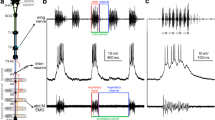
The electrosensory primary afferents in elasmobranchs are responsive to electric potentials created by the animal's own ventilation, while the second-order neurons (AENs) which receive this afferent input in the medulla suppress responses to ventilatory potentials but retain their extreme sensitivity to electric signals in the environment. Ventilatory potentials are common mode signals in elasmobranchs and a common mode rejection mechanism is one way the AENs suppress ventilatory noise. By pressure injecting the GABA-A receptor antagonist SR95531 while extracellularly recording from AENs, we tested the hypothesis that the subtractive circuitry that selectively reduces common mode signals in AENs utilizes GABA, and that a GAB-Aergic component of the dorsal nucleus commissural pathway mediates crossed inhibition of AENs. Local application of SR95531 increased the spontaneous activity and the responsiveness of AENs to electrosensory stimuli. AEN responses to a common mode stimulus were selectively increased compared to responses to a localized stimulus due to SR95531 application. Contralateral inhibition of AENs was blocked by SR95531, indicating that GABAergic commissural cells may inhibit AENs when the contralateral side of the body is stimulated, as with common mode stimulation. We conclude that GABAergic inhibition contributes significantly to the shaping of AEN responses including common mode rejection.
Similar content being viewed by others

Abbreviations
- AENs :
-
ascending efferent neurons
- GABA:
-
gamma-aminobutyric acid
References
Agardh E, Bruun B, Ehinger P, Ekstrom van Veen T, Wu J-Y (1987) Gamma-aminobutyric acid- and glutamic acid decarboxylase-immunoreactive neurons in the retina of different vertebrates. J Comp Neurol 258: 622–630
Bodznick D, Schmidt AW (1982) Functional connections of the medullary electroreceptor nucleus in the little skate. Soc Neurosci Abstr 8: 763
Bodznick D, Montgomery JC (1992) Suppression of ventilatory reafference in the elasmobranch electrosensory system: medullary neuron receptive fields support a common mode rejection mechanism. J Exp Biol 171: 127–137
Bodznick D, Montgomery JC, Bradley DJ (1992) Suppression of common mode signals within the electrosensory system of the little skate Raja erinacea. J Exp Biol 171: 107–125
Boord RL, Northcutt RG (1982) Ascending lateral line pathways to the midbrain of the clearnose skate, Raja eglanteria. J Comp Neurol 207: 274–282
Brunken WJ, Witkovsky P, Karten HJ (1986) Retinal neurochemistry of three elasmobranch species: an immunohistochemical approach. J Comp Neurol 243: 1–12
Caspary DM (1986) Cochlear nuclei: functional neuropharmacology of the principal cell types. In: Altschuler R et al. (eds) Neurobiology of hearing: the cochlea. Raven Press, New York, pp 303–332
Cohen JL (1985) Effects of glycine and GABA on the ganglion cells of the retina of the skate Raja erinacea. Brain Res 332: 169–173
Conley RA (1995) Descending input from the vestibulolateral cerebellum suppresses electrosensory responses in the dorsal octavolateralis nucleus of the elasmobranch, Raja erinacea. J Comp Physiol A 176: 325–335
Duman CH (1995) Interneurons in the dorsal octavolateralis nucleus of the little skate, Raja erinacea: possible roles in the rejection of electrosensory noise. Ph.D. dissertation, Wesleyan University
Duman CH, Bodznick D (1996) Distinct but overlapping populations of commissural and GABAergic neurons in the dorsal nucleus of the little skate, Raja erinacea. Brain Behav Evol (in press)
Evans EF, Zhao W (1993) Neuropharmacological and neurophysiological dissection of inhibition in the mammalian dorsal cochlear nucleus. In: Merchan MA et al. (eds) The mammalian cochlear nuclei: organization and function. Plenum Press, New York, pp 253–266
Heaulme M, Chambon JP, Leyris R, Molimard J-C, Wermuth CG, Biziere K (1986) Biochemical characterization of the interaction of three pyridazinyl-GABA derivatives with the GABAA receptor site. Brain Res 384: 224–223
Heaulme M, Chambon JP, Leyris R, Wermuth CG, Biziere K (1987) Characterization of the binding of (3H)SR 95531, a GABAA antagonist, to rat brain membranes. J Neurochem 48: 1677–1686
Hebebrand J, Friedl W, Reichelt R, Schmitz E, Moller P, Propping P (1988) The shark GABA-benzodiazepine receptor: further evidence for a not so late phylogenetic appearance of the benzodiazepine receptor. Brain Res 446: 251–261
Ito Y, Koshiba T, Doi M, Asami S, Fukuda H, Murakoshi Y (1992) Characterization of antagonistic activity and binding properties of SR 95531, a pyridazinyl-GABA derivative, in rat brain and cultured cerebellar neuronal cells (1992) Synapse 10: 326–333
Kalmijn AJ (1971) The electric sense of sharks and rays. J Exp Biol 55: 371–383
Kalmijn AJ (1978) Electric and magnetic sensory world of sharks, skates and rays. In: Hodgson ES, Mathewson RF (eds) Sensory biology of sharks, skates and rays. Office of Naval Research, Arlington, VA, pp 507–528
Krupp J, Feltz P (1993) Synaptic- and agonist-induced chloride currents in neonatal rat sympathetic preganglionic neurones in vitro. J Physiol 471: 729–748
Lukasiewicz PD, Werblin FS (1994) A novel GABA receptor modulates synaptic transmission from bipolar to ganglion and amacrine cells in the tiger salamander retina. J Neurosci 14: 1213–1223
Lukasiewicz PD, Maple BR, Werblin FS (1994) A novel GABA receptor on bipolar cell terminals in the tiger salamander retina. J Neurosci 14: 1202–1212
Mann E, Enna SJ (1980) Phylogenetic distribution of bicucullinesensitive gamma-aminobutyric acid (GABA) receptor binding. Brain Res 184: 367–373
Michaud JC, Mienville JM, Chambon JP, Biziere K (1986) Interactions between three pyridazinyl-GABA derivatives and central GABA and glycine receptors in the rat, an in vivo microiontophoretic study. Neuropharmacology 25: 1197–1203
Montgomery JC (1984) Noise cancellation in the electrosensory system of the thornback ray; common mode rejection of input produced by the animals own ventilatory movement. J Comp Physiol A 155: 103–111
Montgomery JC, Bodznick D (1993) Hindbrain circuitry mediating common mode suppression of ventilatory reafference in the electrosensory system of the little skate Raja erinacea. J Exp Biol 183: 203–215
Murray RW (1965) Electroreceptor mechanism: the relation of impulse frequency to stimulus strength and responses to pulsed stimuli in the ampullae of Lorenzini of elasmobranchs. In: Cahn P (ed) Lateral line detectors. University of Indiana Press, Bloomington, pp 277–293
New JG (1990) Medullary electrosensory processing in the little skate. I. Response characteristics of neurons in the dorsal octavolateralis nucleus. J Comp Physiol A 167: 285–294
New JG, Bodznick D (1990) Medullary processing in the little skate. II. Suppression of self-generated electrosensory interference during respiration. J Comp Physiol A 167: 295–307
Nicholson LFB, Montgomery JC, Faull LM (1994) GABA, muscarinic cholinergic, excitatory amino acid, neurotensin and opiate binding sites in the octavolateralis column and cerebellum of the skate Raja nasuta (Pices: Rajidae). Brain Res 652: 40–48
Olsen RW, Ban M, Miller T, Johnston GAR (1975) Chemical instability of the GABA antagonist bicuculline under physiological conditions. Brain Res 98: 383
Paul DH, Roberts BL (1977) Studies on a primitive cerebellar cortex. I. The anatomy of the lateral-line lobes of the dogfish, Scyliorhinus canicula. Proc R Soc London Ser B 195: 453–466
Roberts E (1974) Gamma-aminobutyric acid and nervous system function- a perspective. Biochem Pharmacol 23: 2637–2649
Spencer RF, Baker R (1992) GABA and glycine as inhibitory neurotransmitters in the vestibuloocular reflex. Ann NY Acad Sci 656: 602–611
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Duman, C.H., Bodznick, D. A role for GABAergic inhibition in electrosensory processing and common mode rejection in the dorsal nucleus of the little skate, Raja erinacea . J Comp Physiol A 179, 797–807 (1996). https://doi.org/10.1007/BF00207358
Accepted:
Issue Date:
DOI: https://doi.org/10.1007/BF00207358