
Abstract
Proteins do not only serve as nutrients to fulfill the demand for food, but also are used as a source of bioactive proteins/polypeptides for regulating physical functions and promoting physical health. Female breast cancer has the highest incidence in the world and is a serious threat to women’s health. Bioactive proteins/polypeptides exert strong anti-tumor effects and exhibit inhibition of multiple breast cancer cells. This review discussed the suppressing effects of bioactive proteins/polypeptides on breast cancer in vitro and in vivo, and their mechanisms of migration and invasion inhibition, apoptosis induction, and cell cycle arrest. This may contribute to providing a basis for the development of bioactive proteins/polypeptides for the treatment of breast cancer.
Graphical abstract

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Proteins are one of the main nutrients for the requirement of humans and possess functions of promoting growth and development, strengthening immunity, and being involved in metabolism (Kitada et al., 2019). Some special proteins also have particular abilities, such as anti-tumor, immunomodulation, anti-oxidant, and anti-thrombosis, and are termed bioactive proteins (Auestad and Layman, 2021). Proteins are hydrolyzed to polypeptides. These polypeptides commonly contain 2–30 amino acids and have multiple functions determined by their amino acid sequence, being called bioactive polypeptides (Chakrabarti et al., 2018; Chelliah et al., 2021). They have good health effects such as lowering blood pressure, antibacterial activity, anti-tumor, and anti-inflammation. It has been found that bioactive proteins/polypeptides have remarkable anti-tumor effects. For example, fungal immunomodulatory proteins (FIPs) are a kind of bioactive proteins from fungi such as Ganoderma lucidum and Flammulina velutipes (Li et al., 2010a, 2011a), and have inhibitory effects against multiple cancer cells such as gastric, liver and breast cancer cells (Li et al., 2011b, 2019b).
Cancer is a major health problem in the world. Breast cancer is the most commonly diagnosed cancer with 2.26 million new cases worldwide in 2020 (Ferlay et al., 2021). Despite some clinical success, high incidence of recurrence and metastasis still lead to high mortality in breast cancer patients (Akram et al., 2017). Some chemotherapeutic drugs with high doses have serious side effects and drug resistance in the therapy of breast cancer (Al-Mahmood et al., 2018). Thus, it is of great importance to discover and identify more effective agents with fewer side effects (Hu et al., 2019). Natural products are a precious resource for developing and discovering antitumor drugs due to multi-targeting efficacy, no toxicity, and no drug resistance (Hashem et al., 2022; Ullah et al., 2022). Moreover, it has been demonstrated that naturally bioactive products possess abilities in the prevention and treatment of breast cancer (Bak et al., 2016). This review aims to discuss the potential of bioactive proteins/polypeptides in the prevention and treatment of breast cancer.
Acquirement of bioactive proteins/polypeptides
Proteins and polypeptides are the material basis of life. Bioactive proteins/polypeptides are special proteins and polypeptides with particular physiological functions. Bioactive proteins/polypeptides originate mainly from meat, milk, cereal, fish, seaweeds, vegetables, and fungi (Kaur et al., 2021). The most common methods to produce proteins/polypeptides include enzyme hydrolysis and microbial fermentation (Daliri et al., 2017). Besides, a PepSAVI-MS (statistically-guided bioactive peptides prioritized via mass spectrometry) pipeline is developed for bioactive peptide discovery (Kirkpatrick et al., 2017). Using this pipeline, some bioactive peptides are successfully identified from biological species such as Enterococcus faecalis (Kirkpatrick et al., 2018b), Ustilago maydis (Kirkpatrick et al., 2018a), Viola odorata (Parsley et al., 2018) and Amaranthus tricolor (Moyer et al., 2019).
Mechanism of breast cancer suppression by bioactive proteins/polypeptides
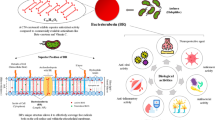
Bioactive proteins/polypeptides have potential medicinal values, especially anti-tumor effects (Li et al., 2011b; Wu et al., 2014). Some bioactive proteins/polypeptides from plants, animals, and microbes can inhibit breast cancer cell growth (Tables 1 and 2) based on mechanisms of migration and invasion inhibition, apoptosis induction, and cell cycle arrest (Fig. 1).

Possible anti-tumor mechanisms of bioactive proteins/polypeptides against breast cancer. Akt, PKB protein kinase B; Bad Bcl-xL/Bcl-2-associated death promoter; Bak Bcl-2 antagonist killer 1; Bax Bcl-2-associated X protein; Bcl-2 B cell lymphoma 2; Bcl-xL B-cell lymphoma extra-large; Bid BH3-interacting domain death agonist; CCR10 chemokine receptor type 10; CD147 cluster of differentiation 147; CD151 cluster of differentiation 151; CD31, PECAM-1 platelet endothelial cell adhesion molecule-1; CHOP C/EBP homologous protein; COX-2 cyclooxygenase 2; ECM extracellular matrix; EGFR epidermal growth factor receptor; ER endoplasmic reticulum; ERK extracellular signal-regulated kinases; FAK focal adhesion kinase; FoxO3a forkhead box O3a; GSK3β glycogen synthase kinase 3-β; IL-6 interleukin 6; JAK2 Janus protein tyrosine kinase 2; MAPK mitogen-activated protein kinase; MEK mitogen-activated protein kinase kinase; MMPs matrix metalloproteinases; PI3K phosphatidylinositol 3-kinase; PTEN phosphatase and tensin homolog; Ras rat sarcoma virus; ROS reactive oxygen species; Src proto-oncogene tyrosine-protein kinase Src; STAT3 signal transducer and activator of transcription 3; tBid truncated Bid
Inhibition of migration and invasion
Cancer cells are capable of migration and invasion to spread within tissues (Chambers et al., 2002). Mechanisms of the migration include extracellular matrix (ECM) degradation, which needs corresponding enzymes, such as matrix metalloproteinases (MMPs) (Alaseem et al., 2019). MMPs are zinc-dependent endopeptidases and contribute to the pathogenesis of various diseases (Bassiouni et al., 2021). Platelet endothelial cell adhesion molecule-1 (PECAM-1 or CD31) is a 130-kDa highly glycosylated transmembrane member of the Ig superfamily (Caligiuri, 2020) and mediates cancer metastasis by activating integrins (Zhang et al., 2018). Tetraspanins CD151, a transmembrane 4 superfamily protein, regulate the epidermal growth factor receptor (EGFR)/focal adhesion kinase (FAK) signaling pathway by affecting integrins (Zhu et al., 2021). Integrins comprise a family of 24 heterodimeric receptors formed by 18 α-subunits and eight β-subunits (Desgrosellier and Cheresh, 2010). Distinct integrin heterodimers are in specific cancer cells. For example, α6β4 and ανβ3 are expressed in breast cancer. Integrins bind to ECM proteins and activate the FAK and proto-oncogene tyrosine-protein kinase Src (Src) family kinase (SFK) signaling, then impinging on the rat sarcoma virus (Ras)/extracellular signal-regulated kinases (ERK) and phosphatidylinositol 3-kinase (PI3K)/protein kinase B (Akt) pathways (Cooper and Giancotti, 2019). The pathways contribute to the regulation of nuclear factor-kappaB (NF-κB) activity (Mali et al., 2018; Mao et al., 2019), promoting the expressions of MMP-2 (Tao et al., 2018), -7 (Liu et al., 2017a) and -9 (Tan et al., 2022). Iturin A is a lipopeptide consisting of a hydrophilic peptide moiety linked to a hydrophobic fatty acid chain and is purified from a marine bacterium Bacillus megaterium (Dey et al., 2015). Zhang et al. isolated water-soluble polypeptides from a traditional Chinese pharmaceutical and functional food Pilose antler, named PAWPs (Zheng et al., 2020). In xenograft models, both iturin A and PAWPs reduce CD31 expression, leading to metastasis inhibition (Dey et al., 2015; Zheng et al., 2020). An important characteristic of iturin A is amphiphilic. Zhao et al. found that the hydrophilic peptide (NYNQPNS) showed no effect on HepG2 cells proliferation, suggesting that the fatty acid chain plays an important role in its activity (Zhao et al., 2021). Using the FireDock web server and immunoprecipitation assay, it is found that a marine natural metabolite Cyclo (l-Leucyl-l-Prolyl) peptide (CLP) from Streptomyces mangrovisoli has a strong interaction with CD151 and reduces CD151 expression (Kgk et al., 2020). Melittin is the main component of bee venom (Raghuraman and Chattopadhyay, 2007), and inhibits the activation of FAK and MMP-9 by blocking the PI3K/Akt/mammalian target of rapamycin (mTOR) signaling pathway (Jeong et al., 2014). Melittin consists of 26 amino acids (GIGAVLKVLTTGLPALISWIKRKRQQ) with a hydrophobic N-terminal and a hydrophilic C-terminal (Raghuraman and Chattopadhyay, 2007). Melittin with the total + 6 charges at physiological pH binds to negatively charged membrane surface and then disturbs the membrane integrity, ultimately leading to cell lysis (Jamasbi et al., 2016), and is also considered as a promising candidate for cancer chemotherapy (Schweizer, 2009). Lunasin, a 43-amino acid peptide isolated from soybean seed (Galvez and de Lumen, 1999), is divided into four regions: N-terminus, central part, RGD motif and C-terminus (Jones and Srivastava, 2014). The full sequence of lunasin exerts cytotoxicity to MDA-MB-231 cells through inhibition of H3 and H4 acetylation (Hernandez-Ledesma et al., 2011), while the RGD motif involves in cancer metastasis (Nieberler et al., 2017). It can also inhibit the phosphorylation of FAK, Src, Akt, and ERK, suppress the nucleus translocation of NF-κB, and reduce the activity and expression of MMP-2 and -9, resulting in the inhibition of invasion of MCF-7 and MDA-MB-231 cells (Jiang et al., 2016). This mechanism might involve that lunasin interacts with integrins, leading to the suppression of the signaling axis (Vuyyuri et al., 2018). Besides, an earthworm fibrinolytic protein (EFP) isolated from Eisenia fetida inhibits FAK expression and the migration of MCF-7 cells (Chen, 2014).
A glycoprotein cluster of differentiation 147 (CD147) is an MMP inducer and as well as promotes interleukin (IL)-6 production via NF-κB (Dana et al., 2021). IL-6 primarily activates Janus protein tyrosine kinase (JAK) 1 and JAK2 to drive signal transduction, including the activity of signal transducer and activator of transcription (STAT), mitogen-activated protein kinase (MAPK), and PI3K-Akt (Kang et al., 2020). The activated STAT3 increases the expression of MMP-2, -7, and -9 (Banerjee and Resat, 2016; Cao et al., 2022). Melittin downregulates CD147 and MMP-9, leading to inhibiting MCF-7 cell invasion (Wang et al., 2017). Ilamycins are a series of cyclic peptides and are isolated from the deep South China Sea-derived S. atratus SCSIO ZH16 and engineered mutant strains (Ma et al., 2017). Ilamycin C can inhibit the invasion and migration of MDA-MB-231 cells via decreasing MMP-2 and MMP-9 by suppressing IL-6-induced STAT3 phosphorylation (Xie et al., 2019).
Chemokine receptor type 10 (CCR10) is a member of the chemokine receptor subfamily and promotes cancer cell invasion and migration through the ERK and PI3K/Akt signaling pathway with the regulation of MMPs (Lin et al., 2017; Liu et al., 2021). A modified linear peptide brintonamide D from samples of intertidal cyanobacterial mats can be used as a CCR10 antagonist to reduce the proliferation and migration of MDA-MB-231 cells (Al-Awadhi et al., 2018). Brintonamide D forms hydrogen bonds with Arg247, Arg322 and Arg345 of CCR10 and the aromatic ring on the modified side of brintonamide D has π–π interaction with residue Tyr227 in the middle of the CCR10 β-sheet.
Inducing apoptosis
Apoptosis is a kind of regulated cell death process (Carneiro and El-Deiry, 2020). Cancer cells are capable of evading apoptosis (Brown and Attardi, 2005). Reactive oxygen species (ROS) increase the levels of phosphatase and tensin homolog (PTEN), as a tumor suppressor gene, negatively regulating PI3K/Akt pathway (Wang et al., 2020, 2022), thereby inducing caspases-dependent apoptosis through itself, forkhead box O3a (FoxO3a) (Yan et al., 2020), and glycogen synthase kinase 3-β (GSK3β) (Guo et al., 2020). Members of the caspase family participate in the initiation and execution of apoptosis (Boice and Bouchier-Hayes, 2020). In MCF-7 and MDA-MB-231 cells, roe protein hydrolysate prepared from defatted Alburnus tarichi roe powder causes the significant production of intracellular ROS, and significantly increases the expressions of caspase-3, -7, -8, and -9, inducing apoptosis (Berkoz et al., 2020). In MDA-MB-231 cells, lunasin does not exhibit the effect of apoptosis, but it promoted aspirin-induced apoptosis (Hsieh et al., 2010). While, in MCF-7 cells, lunasin upregulates PTEN promoter activity, increases PTEN transcript and protein levels, and enhances nuclear PTEN localization, leading to cellular apoptosis (Pabona et al., 2013). A small molecular oligopeptide, sea cucumber intestinal peptide (SCIP) with being rich in hydrophobic amino acids (Ala, Val, Leu, Gly, Phe, and Met) and branched-chain amino acids (Val, Ile, and Leu), is extracted from sea cucumber intestines and promotes the apoptosis of MCF-7 cells through the inactivation of PI3K/Akt signaling pathway with elevating the expression of cleaved caspase-9 and -3 (Wei et al., 2021). But the detailed mechanism remains unclear. In MDA-MB-231 and MCF-7 cells, the apoptotic effect of iturin A is attributed to the Akt-mediated FoxO3a and GSK3β (Dey et al., 2015). Although the activation of caspases is a feature of apoptosis, HN-1, a naturally occurring host defense peptide identified from Amolops hainanensis, induces caspase-independent apoptosis (Qiao et al., 2019).
Cyclooxygenase 2 (COX-2) is an inducible enzyme compared with constitutive COX-1 and is often overexpressed in breast cancer with poor survival (Ristimaki et al., 2002). COX-2 binds with and inactivates p53 (Feng et al., 2019), which regulates apoptotic genes directly and indirectly (Mihara et al., 2003; Hemann and Lowe, 2006). B cell lymphoma 2 (Bcl-2) gene family, including pro-apoptotic members such as Bcl-2-associated X protein (Bax), Bcl-2 antagonist killer 1 (Bak), Bcl-2-related ovarian killer (Bok), Bcl-2-interacting mediator of cell death (Bim), BH3-interacting domain death agonist (Bid), B-cell lymphoma extra-large (Bcl-xL)/Bcl-2-associated death promoter (Bad), and p53 upregulated modulator of apoptosis (Puma), and anti-apoptotic members such as Bcl-2, Bcl-xL, Bcl-2-like protein 2 (Bcl-w), and mantle cell lymphoma 1 (Mcl-1), has a significant role in regulating apoptosis (Ashkenazi et al., 2017). The activation of caspases is mainly regulated by the Bcl-2 family (Tzifi et al., 2012). It has been found that in MCF-7 cells, the roe protein hydrolysate downregulates COX-2 level (Berkoz et al., 2020). In MCF-7 and MDA-MB-231 cells, tuna cooking juice hydrolysate by protease XXIII (PA) with > 2.5 kDa ultrafiltration fraction (PAH2.5) (Hung et al., 2014), CPe-III derived from chickpea albumin hydrolysate (Xue et al., 2015), and AtMP1 and AtMP2 identified from Anabas testudineus antimicrobial peptides (Najm et al., 2021) significantly increase the expressions of p53 and Bax, decrease the level of Bcl-2 which could be overwhelmed by Bax (Yin et al., 1997), and upregulate caspases levels. HN-1 also activates p53 and induces a p53-dependent increase of Bax/Bcl-2 ratio in xenograft tumors (Qiao et al., 2019). Molecular docking analysis showed that Arg1, Gln2, Ala6, Ala8, and Gln9 of CPe-III combine the DNA binding domain of p53 protein (Thr102, Leu111, Asn131, Gln144, Asp228, and Asn268) by hydrogen bonds, resulting in the induction of p53 expression (Xue et al., 2015). Najm et al. found that hydrogen bonds formed between AtMP1/AtMP2 and p53, Bax, Bcl-2, or caspases (Najm et al., 2021). Balsamin is a type I ribosome-inactivating protein purified from Momordica balsamina (Kaur et al., 2012). It increases the expressions of Bax, Bid, and Bad, reduces the levels of Bcl-2 and Bcl-xL, and increases the activities of caspase-3 and -8 in MCF-7 and BT549 cells (Ajji et al., 2017). Halilectin-3 containing three subunits is isolated from Haliclona caerulea (Carneiro et al., 2013), and induces MCF-7 apoptosis with a decrease of Bcl-2 and an increase of caspase-9 (do Nascimento-Neto et al., 2018). Both the α-chain and β-chain of halilectin-3 have N-glycosylation sites with affinity to N-acetylgalactosamine (GalNAc) (Carneiro et al., 2013), thereby recognizing abnormal expressing GalNAc-containing antigen on MCF-7 cells (do Nascimento-Neto et al., 2018). α-Lactalbumin, β-lactoglobulin, and lactoferrin, nutritional components in milk and dairy products, downregulate Bcl-2 and upregulate Bax, leading to increasing caspase-3 in MDA-LM2 cells (Li et al., 2019a). Melittin increases the levels of Bax, caspase-3, and -8 in MDA-MB-231 cells (Daniluk et al., 2019; Mir Hassani et al., 2021). A cytotoxic protein fraction F5 is isolated and is mainly composed of 97.29% serine protease encoded by GME4347_g (Yap et al., 2018). The F5 induces the increases of Bax, Bid, and cleaved Bid, and the decrease of Bcl-2, leading to the upregulation of caspase-8 and -9 activities.
C/EBP homologous protein (CHOP) is a pro-apoptotic endoplasmic reticulum (ER) stress marker, which can be regulated by ROS-mediated ER stress (Zhu et al., 2022), subsequently regulating Bcl-2/Bax (Li et al., 2019c; Liu et al., 2020). WSPF is a novel protein fraction isolated from Withania somnifera roots and induces apoptosis of MDA-MB-231 cells through the production of extensive ROS, leading to reducing Bcl-2 expression, increasing Bax expression, and elevating cleaved caspase-3 expression (Dar et al., 2019). Ilamycin E is another cyclic peptide (Ma et al., 2017). It activates ER stress, increases CHOP, and downregulates Bcl-2, which promotes apoptosis in HCC1937 and MDA-MB-468 cells (Zhou et al., 2019).
Bcl-2 can also be regulated by STAT3 (Liu et al., 2017b). Ilamycin C promotes Bax/Bcl-2-related caspase-dependent apoptosis through IL-6/JAK2/STAT3 as well (Xie et al., 2019).
Cell cycle arrest
The cell cycle is a tightly regulated process including Gap 1 (G1), DNA-synthesis (S), Gap 2 (G2), and mitosis (M) phases. The core cell-cycle proteins are frequently dysregulated in human cancers, and targeting these proteins seems to represent an effective strategy for inhibiting tumors (Suski et al., 2021). Mitogenic signals upregulate cyclin D, which binds and activate cyclin‐dependent kinase (CDK) 4 or CDK6 to drive the progression from the G0 or G1 into the S phase. Degradation of cyclin D is a promising targeted therapy for the cancer cell cycle (Caudron-Herger and Diederichs, 2021; Chaikovsky et al., 2021; Maiani et al., 2021; Simoneschi et al., 2021). p21 and p27 are capable of inhibiting cyclin/CDK complexes comprising CDK1 or 2 and cyclin A; CDK1 and CDK2 can be activated by cyclin A which has a critical role in the S and G2-M phase (Suski et al., 2021; Garcia-Osta et al., 2022). Ilamycin E decreases Cyclin D1, and increases p21 and p27 levels, thereby inducing G1/S cell cycle arrest in HCC1937 and MDA-MB-468 cells (Zhou et al., 2019). PAH2.5 increases p21 and p27 protein expression and decreases cyclin A expression, which induces cell cycle arrest in the S phase in MCF-7 cells (Hung et al., 2014).
Conclusions and perspectives
Breast cancer is the leading cause of cancer death among women (Ghoncheh et al., 2016). It is a hormone-dependent tumor (Russnes et al., 2017), and its targeted therapies include human epidermal growth factor receptor 2-targeted agents and endocrine therapy (Turashvili and Brogi, 2017). However, drug resistance is still a major challenge in the treatment of breast cancer (Karami Fath et al., 2022). Bioactive proteins/polypeptides exhibit a great potential against cancers, including breast cancer. Identification of novel bioactive proteins/polypeptides and development of novel functions of the existing bioactive proteins/polypeptides are necessary. Generally, protein is digested and absorbed in the form of amino acids from diet, although proteins with medicinal value still have bioactivity (Lee et al., 2018). Treatments of iturins (Zhao et al., 2018), FIP-gmi (Hsin et al., 2020) and PAWPs (Zheng et al., 2020) by gavage still exhibit bioactivity without side effects. On the other hand, unfortunately, some proteins/polypeptides such as melittin exhibits extensive hemolysis and cytotoxicity, which may limit its application in clinical practice (Askari et al., 2021). Thus, the pharmacokinetics and pharmacodynamics of bioactive proteins/polypeptides have been well studied further. At the same time, it is critical to modify the proteins/polypeptides to enhance their effects (Berdan et al., 2021). Several analogues of galaxamide which is isolated from Galaxaura filamentosa are synthesized (Xu et al., 2008; Lunagariya et al., 2017). These analogues exhibit greater excellent toxicity toward breast cancer cells. Whey protein isolates modified with rosmarinic acid at alkaline conditions exhibits enhanced antioxidative capacity (Ali et al., 2018). The covalent complex of soy protein isolates and epigallocatechin gallate has higher thermal stability and oxidation resistance and a polyphenol-protective effect (Zhou et al., 2020a). A peptide drug conjugate named TAMpepK, consisting of melittin and a pro-apoptotic peptide, targets M2-like tumor-associated macrophages, thereby inhibiting breast cancer metastasis in the mouse model (Lee et al., 2022). In the previous study, we found that N-glycosylation significantly improves the functional properties of FIP-glu (Li et al., 2021a). Besides, combination regimen-based therapies are promising strategies (Li et al., 2021b). For example, Co-treatment of melittin and hormone therapeutic drugs (Yen et al., 2022), drugs with anti-tumor potential (Duarte et al., 2022), agents (Shaw et al., 2019), and miRNAs (Motiei et al., 2021) reveals synergistic effects in breast cancer cells. Bioactive proteins/polypeptides are suitable for gene therapy, a promising strategy for cancer treatment (Zhou et al., 2020b). The anti-tumor research of bioactive proteins/polypeptides is mainly performed in cell and animal models. However, relevant clinical studies are rare (Table 3). Although the anti-tumor mechanisms of bioactive proteins/polypeptides have been actively studied, there are still problems in the production, administration and regulation of bioactive proteins/polypeptides (Chakrabarti et al., 2018). Therefore, further studies are needed to evaluate the physiological efficacy of these bioactive proteins/polypeptides in human clinical studies. To summarize, bioactive proteins/polypeptides have impressive anti-tumor effects against breast cancer. It is suggested that bioactive proteins/polypeptides with great potential are promising agents for the treatment of breast cancer.
References
Ajji PK, Walder K, Puri M. Functional analysis of a type-I ribosome inactivating protein balsamin from Momordica balsamina with anti-microbial and Dnase activity. Plant Foods for Human Nutrition. 71: 265-271 (2016)
Ajji PK, Binder MJ, Walder K, Puri M. Balsamin induces apoptosis in breast cancer cells via DNA fragmentation and cell cycle arrest. Molecular and Cellular Biochemistry. 432: 189-198 (2017)
Akram M, Iqbal M, Daniyal M, Khan AU. Awareness and current knowledge of breast cancer. Biological Research. 50: 33 (2017)
Al Kashgry NAT, Abulreesh HH, El-Sheikh IA, Almaroai YA, Salem R, Mohamed I, Waly FR, Osman G, Mohamed MSM. Utilization of a recombinant defensin from Maize (Zea mays L.) as a potential antimicrobial peptide. AMB Express. 10: 208 (2020)
Alaseem A, Alhazzani K, Dondapati P, Alobid S, Bishayee A, Rathinavelu A. Matrix Metalloproteinases: A challenging paradigm of cancer management. Seminars in Cancer Biology. 56: 100-115 (2019)
Al-Awadhi FH, Gao B, Rezaei MA, Kwan JC, Li C, Ye T, Paul VJ, Luesch H. Discovery, synthesis, pharmacological profiling, and biological characterization of brintonamides A-E, novel dual protease and GPCR modulators from a marine cyanobacterium. Journal of Medicinal Chemistry. 61: 6364-6378 (2018)
Alfaleh K. Bovine lactoferrin prevents invasive fungal infections in very low birth weight infants: a randomized controlled trial. Journal of Clinical Neonatology. 1: 68-69 (2012)
Ali M, Keppler JK, Coenye T, Schwarz K. Covalent whey protein-rosmarinic acid interactions: A comparison of alkaline and enzymatic modifications on physicochemical, antioxidative, and antibacterial properties. Journal of Food Science. 83: 2092-2100 (2018)
Al-Mahmood S, Sapiezynski J, Garbuzenko OB, Minko T. Metastatic and triple-negative breast cancer: Challenges and treatment options. Drug Delivery and Translational Research. 8: 1483-1507 (2018)
Ashkenazi A, Fairbrother WJ, Leverson JD, Souers AJ. From basic apoptosis discoveries to advanced selective BCL-2 family inhibitors. Nature Reviews: Drug Discovery. 16: 273-284 (2017)
Askari P, Namaei MH, Ghazvini K, Hosseini M. In vitro and in vivo toxicity and antibacterial efficacy of melittin against clinical extensively drug-resistant bacteria. BMC Pharmacol Toxicol. 22: 42 (2021)
Auestad N, Layman DK. Dairy bioactive proteins and peptides: A narrative review. Nutrition Reviews. 79: 36-47 (2021)
Bak MJ, Das Gupta S, Wahler J, Suh N. Role of dietary bioactive natural products in estrogen receptor-positive breast cancer. Seminars in Cancer Biology. 40-41: 170-191 (2016)
Banerjee K, Resat H. Constitutive activation of STAT3 in breast cancer cells: A review. International Journal of Cancer. 138: 2570-2578 (2016)
Bassiouni W, Ali MAM, Schulz R. Multifunctional intracellular matrix metalloproteinases: implications in disease. FEBS Journal. 288: 7162-7182 (2021)
Berdan VY, Klauser PC, Wang L. Covalent peptides and proteins for therapeutics. Bioorganic and Medicinal Chemistry. 29: 115896 (2021)
Berkoz M, Ozkan-Yilmaz F, Ozluer-Hunt A, Krosniak M, Turkmen O, Yunusoglu O. Roe protein hydrolysate of Alburnus tarichi induces apoptosis in breast cancer MCF-7 and MDA-MB-231 cells through a caspase-dependent pathway. General Physiology and Biophysics. 39: 449-460 (2020)
Boice A, Bouchier-Hayes L. Targeting apoptotic caspases in cancer. Biochimica et Biophysica Acta - Molecular Cell Research. 1867: 118688 (2020)
Bosseboeuf A, Baron A, Duval E, Gautier A, Sourdaine P, Auvray P. K092A and K092B, two peptides isolated from the dogfish (Scyliorhinus canicula L.), with potential antineoplastic activity against human prostate and breast cancer cells. Marine Drugs. 17: 672 (2019)
Bovi M, Carrizo ME, Capaldi S, Perduca M, Chiarelli LR, Galliano M, Monaco HL. Structure of a lectin with antitumoral properties in king bolete (Boletus edulis) mushrooms. Glycobiology. 21: 1000-1009 (2011)
Brown JM, Attardi LD. The role of apoptosis in cancer development and treatment response. Nature Reviews: Cancer. 5: 231-237 (2005)
Caligiuri G. CD31 as a therapeutic target in atherosclerosis. Circulation Research. 126: 1178-1189 (2020)
Cam A, de Mejia EG. RGD-peptide lunasin inhibits Akt-mediated NF-kappaB activation in human macrophages through interaction with the alphaVbeta3 integrin. Molecular Nutrition & Food Research. 56: 1569-1581 (2012)
Cao L, Zhang Y, Mi J, Shi Z, Fang Z, Jia D, Pan Z, Peng P. alpha-Hederin inhibits the platelet activating factor-induced metastasis of HCC cells through disruption of PAF/PTAFR axis cascaded STAT3/MMP-2 expression. Pharmacological Research. 178: 106180 (2022)
Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nature Reviews: Clinical Oncology. 17: 395-417 (2020)
Carneiro RF, de Melo AA, de Almeida AS, Moura Rda M, Chaves RP, de Sousa BL, do Nascimento KS, Sampaio SS, Lima JP, Cavada BS, Nagano CS, Sampaio AH. H-3, a new lectin from the marine sponge Haliclona caerulea: purification and mass spectrometric characterization. International Journal of Biochemistry & Cell Biology. 45: 2864-2873 (2013)
Caudron-Herger M, Diederichs S. Insights from the degradation mechanism of cyclin D into targeted therapy of the cancer cell cycle. Signal Transduction and Targeted Therapy. 6: 311 (2021)
Chaikovsky AC, Li C, Jeng EE, Loebell S, Lee MC, Murray CW, Cheng R, Demeter J, Swaney DL, Chen SH, Newton BW, Johnson JR, Drainas AP, Shue YT, Seoane JA, Srinivasan P, He A, Yoshida A, Hipkins SQ, McCrea E, Poltorack CD, Krogan NJ, Diehl JA, Kong C, Jackson PK, Curtis C, Petrov DA, Bassik MC, Winslow MM, Sage J. The AMBRA1 E3 ligase adaptor regulates the stability of cyclin D. Nature. 592: 794-798 (2021)
Chakrabarti S, Guha S, Majumder K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients. 10: 1738 (2018)
Chambers AF, Groom AC, MacDonald IC. Dissemination and growth of cancer cells in metastatic sites. Nature Reviews: Cancer. 2: 563-572 (2002)
Chelliah R, Wei S, Daliri EB, Elahi F, Yeon SJ, Tyagi A, Liu S, Madar IH, Sultan G, Oh DH. The role of bioactive peptides in diabetes and obesity. Foods. 10: 2220 (2021)
Chen XT. The preliminary studies on the separation and purification of earthworm fibrinolytic protein and its effects on MCF-7 cell growth and adhesion. Master Master thesis, Shandong University. (2014) (in Chinese)
Cooper J, Giancotti FG. Integrin signaling in cancer: Mechanotransduction, stemness, epithelial plasticity, and therapeutic resistance. Cancer Cell. 35: 347-367 (2019)
Crusca E, Jr., Basso LGM, Altei WF, Marchetto R. Biophysical characterization and antitumor activity of synthetic Pantinin peptides from scorpion's venom. Biochimica et Biophysica Acta - Biomembranes. 1860: 2155-2165 (2018)
Daliri EB, Oh DH, Lee BH. Bioactive peptides. Foods. 6: 32 (2017)
Dana P, Kariya R, Lert-Itthiporn W, Seubwai W, Saisomboon S, Wongkham C, Okada S, Wongkham S, Vaeteewoottacharn K. Homophilic interaction of CD147 promotes IL-6-mediated cholangiocarcinoma invasion via the NF-kappaB-dependent pathway. International Journal of Molecular Sciences. 22: 13496 (2021)
Daniluk K, Kutwin M, Grodzik M, Wierzbicki M, Strojny B, Szczepaniak J, Balaban J, Sosnowska M, Chwalibog A, Sawosz E, Jaworski S. Use of selected carbon nanoparticles as melittin carriers for MCF-7 and MDA-MB-231 human breast cancer cells. Materials (Basel). 13: 90 (2019)
D'Anna R, Corrado F, Loddo S, Gullo G, Giunta L, Di Benedetto A. Myoinositol plus alpha-lactalbumin supplementation, insulin resistance and birth outcomes in women with gestational diabetes mellitus: a randomized, controlled study. Scientific Reports. 11: 8866 (2021)
Dar PA, Mir SA, Bhat JA, Hamid A, Singh LR, Malik F, Dar TA. An anti-cancerous protein fraction from Withania somnifera induces ROS-dependent mitochondria-mediated apoptosis in human MDA-MB-231 breast cancer cells. International Journal of Biological Macromolecules. 135: 77-87 (2019)
Desgrosellier JS, Cheresh DA. Integrins in cancer: Biological implications and therapeutic opportunities. Nature Reviews: Cancer. 10: 9-22 (2010)
Dey G, Bharti R, Dhanarajan G, Das S, Dey KK, Kumar BN, Sen R, Mandal M. Marine lipopeptide Iturin A inhibits Akt mediated GSK3beta and FoxO3a signaling and triggers apoptosis in breast cancer. Scientific Reports. 5: 10316 (2015)
Dey G, Bharti R, Ojha PK, Pal I, Rajesh Y, Banerjee I, Banik P, Parida S, Parekh A, Sen R, Mandal M. Therapeutic implication of 'Iturin A' for targeting MD-2/TLR4 complex to overcome angiogenesis and invasion. Cellular Signalling. 35: 24-36 (2017)
Dia VP, Gonzalez de Mejia E. Lunasin potentiates the effect of oxaliplatin preventing outgrowth of colon cancer metastasis, binds to alpha5beta1 integrin and suppresses FAK/ERK/NF-kappaB signaling. Cancer Letters. 313: 167-180 (2011)
do Nascimento-Neto LG, Cabral MG, Carneiro RF, Silva Z, Arruda FVS, Nagano CS, Fernandes AR, Sampaio AH, Teixeira EH, Videira PA. Halilectin-3, a lectin from the marine sponge Haliclona caerulea, induces apoptosis and autophagy in human breast cancer MCF7 cells through caspase-9 pathway and LC3-II protein expression. Anti-Cancer Agents in Medicinal Chemistry. 18: 521-528 (2018)
Duarte D, Falcao SI, El Mehdi I, Vilas-Boas M, Vale N. Honeybee venom synergistically enhances the cytotoxic effect of CNS drugs in HT-29 colon and MCF-7 breast cancer cell lines. Pharmaceutics. 14: 511 (2022)
Duffy C, Sorolla A, Wang E, Golden E, Woodward E, Davern K, Ho D, Johnstone E, Pfleger K, Redfern A, Iyer KS, Baer B, Blancafort P. Honeybee venom and melittin suppress growth factor receptor activation in HER2-enriched and triple-negative breast cancer. npj Precision Oncology. 4: 24 (2020)
El Mehdi I, Falcao SI, Harandou M, Boujraf S, Calhelha RC, Ferreira I, Anjos O, Campos MG, Vilas-Boas M. Chemical, cytotoxic, and anti-inflammatory assessment of honey bee venom from Apis mellifera intermissa. Antibiotics (Basel). 10: 1514 (2021)
Feng F, Wang Z, Li R, Wu Q, Gu C, Xu Y, Peng W, Han D, Zhou X, Wu J, He H. Citrus alkaline extracts prevent fibroblast senescence to ameliorate pulmonary fibrosis via activation of COX-2. Biomedicine and Pharmacotherapy. 112: 108669 (2019)
Ferlay J, Colombet M, Soerjomataram I, Parkin DM, Pineros M, Znaor A, Bray F. Cancer statistics for the year 2020: An overview. International Journal of Cancer. 149: 778-789 (2021)
Galvez AF, de Lumen BO. A soybean cDNA encoding a chromatin-binding peptide inhibits mitosis of mammalian cells. Nature Biotechnology. 17: 495-500 (1999)
Garcia-Osta A, Dong J, Moreno-Aliaga MJ, Ramirez MJ. p27, the cell cycle and Alzheimer´s disease. International Journal of Molecular Sciences. 23: 1211 (2022)
Gasymov OK, Celik S, Agaeva G, Akyuz S, Kecel-Gunduz S, Qocayev NM, Ozel AE, Agaeva U, Bakhishova M, Aliyev JA. Evaluation of anti-cancer and anti-covid-19 properties of cationic pentapeptide Glu-Gln-Arg-Pro-Arg, from rice bran protein and its d-isomer analogs through molecular docking simulations. Journal of Molecular Graphics and Modelling. 108: 107999 (2021)
Ghoncheh M, Pournamdar Z, Salehiniya H. Incidence and mortality and epidemiology of breast cancer in the world. Asian Pacific Journal of Cancer Prevention. 17: 43-46 (2016)
Guha S, Ferrie RP, Ghimire J, Ventura CR, Wu E, Sun L, Kim SY, Wiedman GR, Hristova K, Wimley WC. Applications and evolution of melittin, the quintessential membrane active peptide. Biochemical Pharmacology. 193: 114769 (2021)
Guo X, Zhu K, Zhang H, Yao H. Anti-tumor activity of a novel protein obtained from tartary buckwheat. International Journal of Molecular Sciences. 11: 5201-5211 (2010)
Guo Y, Wei L, Zhou Y, Lu N, Tang X, Li Z, Wang X. Flavonoid GL-V9 induces apoptosis and inhibits glycolysis of breast cancer via disrupting GSK-3beta-modulated mitochondrial binding of HKII. Free Radical Biology and Medicine. 146: 119-129 (2020)
Haddad Tabrizi S, Haddad E, Rajaram S, Oda K, Kaur A, Sabate J. The effect of soybean lunasin on cardiometabolic risk factors: A randomized clinical trial. Journal of Dietary Supplements. 17: 286-299 (2020)
Hashem S, Ali TA, Akhtar S, Nisar S, Sageena G, Ali S, Al-Mannai S, Therachiyil L, Mir R, Elfaki I, Mir MM, Jamal F, Masoodi T, Uddin S, Singh M, Haris M, Macha M, Bhat AA. Targeting cancer signaling pathways by natural products: Exploring promising anti-cancer agents. Biomedicine and Pharmacotherapy. 150: 113054 (2022)
Hemann MT, Lowe SW. The p53-Bcl-2 connection. Cell Death & Differentiation. 13: 1256-1259 (2006)
Hernandez Marin I, Picconi O, Lagana AS, Costabile L, Unfer V. A multicenter clinical study with myo-inositol and alpha-lactalbumin in Mexican and Italian PCOS patients. European Review for Medical and Pharmacological Sciences. 25: 3316-3324 (2021)
Hernandez-Ledesma B, Hsieh CC, de Lumen BO. Antioxidant and anti-inflammatory properties of cancer preventive peptide lunasin in RAW 264.7 macrophages. Biochemical and Biophysical Research Communications. 390: 803-808 (2009)
Hernandez-Ledesma B, Hsieh CC, de Lumen BO. Relationship between lunasin's sequence and its inhibitory activity of histones H3 and H4 acetylation. Molecular Nutrition & Food Research. 55: 989-998 (2011)
Hew CS, Khoo BY, Gam LH. The anti-cancer property of proteins extracted from Gynura procumbens (Lour.) Merr. PloS One. 8: e68524 (2013)
Hsieh CC, Hernandez-Ledesma B, de Lumen BO. Lunasin, a novel seed peptide, sensitizes human breast cancer MDA-MB-231 cells to aspirin-arrested cell cycle and induced apoptosis. Chemico-Biological Interactions. 186: 127-134 (2010)
Hsin IL, Chiu LY, Ou CC, Wu WJ, Sheu GT, Ko JL. CD133 inhibition via autophagic degradation in pemetrexed-resistant lung cancer cells by GMI, a fungal immunomodulatory protein from Ganoderma microsporum. British Journal of Cancer. 123: 449-458 (2020)
Hsu KC, Li-Chan ECY, Jao CL. Antiproliferative activity of peptides prepared from enzymatic hydrolysates of tuna dark muscle on human breast cancer cell line MCF-7. Food Chemistry. 126: 617-622 (2011)
Hu DD, Zhang RY, Zhang GQ, Wang HX, Ng TB. A laccase with antiproliferative activity against tumor cells from an edible mushroom, white common Agrocybe cylindracea. Phytomedicine. 18: 374-379 (2011)
Hu MH, Wu TY, Huang Q, Jin G. New substituted quinoxalines inhibit triple-negative breast cancer by specifically downregulating the c-MYC transcription. Nucleic Acids Research. 47: 10529-10542 (2019)
Hung CC, Yang YH, Kuo PF, Hsu KC. Protein hydrolysates from tuna cooking juice inhibit cell growth and induce apoptosis of human breast cancer cell line MCF-7. Journal of Functional Foods. 11: 563-570 (2014)
Jahanbani R, Ghaffari SM, Salami M, Vahdati K, Sepehri H, Sarvestani NN, Sheibani N, Moosavi-Movahedi AA. Antioxidant and anticancer activities of walnut (Juglans regia L.) protein hydrolysates using different proteases. Plant Foods for Human Nutrition. 71: 402-409 (2016)
Jamasbi E, Mularski A, Separovic F. Model Membrane and Cell Studies of Antimicrobial Activity of Melittin Analogues. Current Topics in Medicinal Chemistry. 16: 40-45 (2016)
Jeong YJ, Choi Y, Shin JM, Cho HJ, Kang JH, Park KK, Choe JY, Bae YS, Han SM, Kim CH, Chang HW, Chang YC. Melittin suppresses EGF-induced cell motility and invasion by inhibiting PI3K/Akt/mTOR signaling pathway in breast cancer cells. Food and Chemical Toxicology. 68: 218-225 (2014)
Jiang Q, Pan Y, Cheng Y, Li H, Liu D, Li H. Lunasin suppresses the migration and invasion of breast cancer cells by inhibiting matrix metalloproteinase-2/-9 via the FAK/Akt/ERK and NF-kappaB signaling pathways. Oncology Reports. 36: 253-262 (2016)
Jones G, Srivastava A. Understanding Lunasin's biology and potential as a cancer therapeutic by utilizing Drosophila genetics. Experimental Biology and Medicine (Maywood, N.J.). 239: 519-528 (2014)
Kang S, Narazaki M, Metwally H, Kishimoto T. Historical overview of the interleukin-6 family cytokine. Journal of Experimental Medicine. 217: e20190347 (2020)
Kannan A, Hettiarachchy NS, Lay JO, Liyanage R. Human cancer cell proliferation inhibition by a pentapeptide isolated and characterized from rice bran. Peptides. 31: 1629-1634 (2010)
Karami Fath M, Azargoonjahromi A, Kiani A, Jalalifar F, Osati P, Akbari Oryani M, Shakeri F, Nasirzadeh F, Khalesi B, Nabi-Afjadi M, Zalpoor H, Mard-Soltani M, Payandeh Z. The role of epigenetic modifications in drug resistance and treatment of breast cancer. Cellular & Molecular Biology Letters. 27: 52 (2022)
Kaur I, Yadav SK, Hariprasad G, Gupta RC, Srinivasan A, Batra JK, Puri M. Balsamin, a novel ribosome-inactivating protein from the seeds of Balsam apple Momordica balsamina. Amino Acids. 43: 973-981 (2012)
Kaur I, Puri M, Ahmed Z, Blanchet FP, Mangeat B, Piguet V. Inhibition of HIV-1 replication by balsamin, a ribosome inactivating protein of Momordica balsamina. PloS One. 8: e73780 (2013)
Kaur A, Kehinde BA, Sharma P, Sharma D, Kaur S. Recently isolated food-derived antihypertensive hydrolysates and peptides: A review. Food Chemistry. 346: 128719 (2021)
Kgk D, Kumari S, G S, Malla RR. Marine natural compound cyclo(L-leucyl-L-prolyl) peptide inhibits migration of triple negative breast cancer cells by disrupting interaction of CD151 and EGFR signaling. Chemico-Biological Interactions. 315: 108872 (2020)
Kirkpatrick CL, Broberg CA, McCool EN, Lee WJ, Chao A, McConnell EW, Pritchard DA, Hebert M, Fleeman R, Adams J, Jamil A, Madera L, Stromstedt AA, Goransson U, Liu Y, Hoskin DW, Shaw LN, Hicks LM. The "PepSAVI-MS" pipeline for natural product bioactive peptide discovery. Analytical Chemistry. 89: 1194-1201 (2017)
Kirkpatrick CL, Parsley NC, Bartges TE, Cooke ME, Evans WS, Heil LR, Smith TJ, Hicks LM. Fungal secretome analysis via PepSAVI-MS: Identification of the bioactive peptide KP4 from Ustilago maydis. Journal of the American Society for Mass Spectrometry. 29: 859-865 (2018a)
Kirkpatrick CL, Parsley NC, Bartges TE, Wing CE, Kommineni S, Kristich CJ, Salzman NH, Patrie SM, Hicks LM. Exploring bioactive peptides from bacterial secretomes using PepSAVI-MS: Identification and characterization of Bac-21 from Enterococcus faecalis pPD1. Microbial Biotechnology. 11: 943-951 (2018b)
Kitada M, Ogura Y, Monno I, Koya D. The impact of dietary protein intake on longevity and metabolic health. EBioMedicine. 43: 632-640 (2019)
Kong Y. Nebrodeolysin, a novel hemolytic protein from the mushroom Pleurotus nebrodensis with cytostatic activity against human cancer cell lines and anti-HIV-1 activity. Master Master thesis, Sichuan University. (2007) (in Chinese)
Lam SK, Ng TB. A dimeric high-molecular-weight chymotrypsin inhibitor with antitumor and HIV-1 reverse transcriptase inhibitory activities from seeds of Acacia confusa. Phytomedicine. 17: 621-625 (2010)
Lam YW, Ng TB, Wang HX. Antiproliferative and antimitogenic activities in a peptide from puffball mushroom Calvatia caelata. Biochemical and Biophysical Research Communications. 289: 744-749 (2001)
Lee YT, Wu CT, Sun HL, Ko JL, Lue KH. Fungal immunomodulatory protein-fve could modulate airway remodel through by affect IL17 cytokine. J Microbiol Immunol Infect. 51: 598-607 (2018)
Lee C, Kim S, Jeong C, Cho I, Jo J, Han IH, Bae H. TAMpepK suppresses metastasis through the elimination of M2-like tumor-associated macrophages in triple-negative breast cancer. International Journal of Molecular Sciences. 23: 2157 (2022)
Li QZ, Wang XF, Bao TW, Ran L, Lin J, Zhou XW. In vitro synthesis of a recombinant fungal immunomodulatory protein from Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (W.Curt.:Fr.) P.Karst. (Aphyllophoromycetideae) and analysis of its immunomodulatory activity. International Journal of Medicinal Mushrooms. 12: 347-358 (2010a)
Li Y, Zhang G, Ng TB, Wang H. A novel lectin with antiproliferative and HIV-1 reverse transcriptase inhibitory activities from dried fruiting bodies of the monkey head mushroom Hericium erinaceum. Journal of Biomedicine and Biotechnology. 2010: 716515 (2010b)
Li QZ, Huang L, Wang XF, Li XS, Wu SQ, Zhou XW. Fungal immunomodulatory protein from Flammulina velutipes induces cytokine gene expression in mouse spleen cells. Current Topics in Nutraceutical Research. 9: 111-118 (2011a)
Li QZ, Wang XF, Zhou XW. Recent status and prospects of the fungal immunomodulatory protein family. Critical Reviews in Biotechnology. 31: 365-375 (2011b)
Li S, Hao L, Bao W, Zhang P, Su D, Cheng Y, Nie L, Wang G, Hou F, Yang Y. A novel short anionic antibacterial peptide isolated from the skin of Xenopus laevis with broad antibacterial activity and inhibitory activity against breast cancer cell. Archives of Microbiology. 198: 473-482 (2016a)
Li S, Zhao L, Xu W, Jiang Z, Kang J, Wang F, Xin F. Identification and characterisation of a novel protein FIP-sch3 from Stachybotrys chartarum. PloS One. 11: e0168436 (2016b)
Li HY, Li P, Yang HG, Wang YZ, Huang GX, Wang JQ, Zheng N. Investigation and comparison of the anti-tumor activities of lactoferrin, alpha-lactalbumin, and beta-lactoglobulin in A549, HT29, HepG2, and MDA231-LM2 tumor models. Journal of Dairy Science. 102: 9586-9597 (2019a)
Li QZ, Zheng YZ, Zhou XW. Fungal immunomodulatory proteins: Characteristic, potential antitumor activities and their molecular mechanisms. Drug Discovery Today. 24: 307-314 (2019b)
Li Y, Jiang W, Niu Q, Sun Y, Meng C, Tan L, Song C, Qiu X, Liao Y, Ding C. eIF2alpha-CHOP-BCl-2/JNK and IRE1alpha-XBP1/JNK signaling promote apoptosis and inflammation and support the proliferation of Newcastle disease virus. Cell Death & Disease. 10: 891 (2019c)
Li QZ, Chen X, Mao PW, Jin MY, Wu Q, Zhou XW. N-Glycosylated Ganoderma lucidum immunomodulatory protein improved anti-inflammatory activity via inhibition of the p38 MAPK pathway. Food & Function. 12: 3393-3404 (2021a)
Li QZ, Zuo ZW, Zhou ZR, Ji Y. Polyamine homeostasis-based strategies for cancer: The role of combination regimens. European Journal of Pharmacology. 910: 174456 (2021b)
Lin P, Ng TB. Brassiparin, an antifungal peptide from Brassica parachinensis seeds. Journal of Applied Microbiology. 106: 554-563 (2009)
Lin HY, Sun SM, Lu XF, Chen PY, Chen CF, Liang WQ, Peng CY. CCR10 activation stimulates the invasion and migration of breast cancer cells through the ERK1/2/MMP-7 signaling pathway. International Immunopharmacology. 51: 124-130 (2017)
Liu JY, Dai YB, Li X, Cao K, Xie D, Tong ZT, Long Z, Xiao H, Chen MK, Ye YL, Liu B, Tan J, Tang J, Xu ZZ, Gan Y, Zhou YH, Deng F, He LY. Solute carrier family 12 member 5 promotes tumor invasion/metastasis of bladder urothelial carcinoma by enhancing NF-kappaB/MMP-7 signaling pathway. Cell Death & Disease. 8: e2691 (2017a)
Liu K, Ren T, Huang Y, Sun K, Bao X, Wang S, Zheng B, Guo W. Apatinib promotes autophagy and apoptosis through VEGFR2/STAT3/BCL-2 signaling in osteosarcoma. Cell Death & Disease. 8: e3015 (2017b)
Liu TH, Tu WQ, Tao WC, Liang QE, Xiao Y, Chen LG. Verification of resveratrol inhibits intestinal aging by downregulating ATF4/Chop/Bcl-2/Bax signaling pathway: Based on network pharmacology and animal experiment. Frontiers in Pharmacology. 11: 1064 (2020)
Liu Y, Xiao A, Zhang B. CCR10/CCL27 crosstalk regulates cell metastasis via PI3K-Akt signaling axis in non-small-cell lung cancer. American Journal of Translational Research. 13: 13135-13146 (2021)
Lu J, Chen ZW. Isolation, characterization and anti-cancer activity of SK84, a novel glycine-rich antimicrobial peptide from Drosophila virilis. Peptides. 31: 44-50 (2010)
Lunagariya J, Liao X, Long W, Zhong S, Bhadja P, Li H, Zhao B, Xu S. Cytotoxicity study of cyclopentapeptide analogues of marine natural product galaxamide towards human breast cancer cells. Oxidative Medicine and Cellular Longevity. 2017: 8392035 (2017)
Ma S, Huang D, Zhai M, Yang L, Peng S, Chen C, Feng X, Weng Q, Zhang B, Xu M. Isolation of a novel bio-peptide from walnut residual protein inducing apoptosis and autophagy on cancer cells. BMC Complementary and Alternative Medicine. 15: 413 (2015)
Ma J, Huang H, Xie Y, Liu Z, Zhao J, Zhang C, Jia Y, Zhang Y, Zhang H, Zhang T, Ju J. Biosynthesis of ilamycins featuring unusual building blocks and engineered production of enhanced anti-tuberculosis agents. Nature Communications. 8: 391 (2017)
Maiani E, Milletti G, Nazio F, Holdgaard SG, Bartkova J, Rizza S, Cianfanelli V, Lorente M, Simoneschi D, Di Marco M, D'Acunzo P, Di Leo L, Rasmussen R, Montagna C, Raciti M, De Stefanis C, Gabicagogeascoa E, Rona G, Salvador N, Pupo E, Merchut-Maya JM, Daniel CJ, Carinci M, Cesarini V, O'Sullivan A, Jeong YT, Bordi M, Russo F, Campello S, Gallo A, Filomeni G, Lanzetti L, Sears RC, Hamerlik P, Bartolazzi A, Hynds RE, Pearce DR, Swanton C, Pagano M, Velasco G, Papaleo E, De Zio D, Maya-Mendoza A, Locatelli F, Bartek J, Cecconi F. AMBRA1 regulates cyclin D to guard S-phase entry and genomic integrity. Nature. 592: 799-803 (2021)
Mali AV, Joshi AA, Hegde MV, Kadam SS. Enterolactone modulates the ERK/NF-kappaB/Snail signaling pathway in triple-negative breast cancer cell line MDA-MB-231 to revert the TGF-beta-induced epithelial-mesenchymal transition. Cancer Biology & Medicine. 15: 137-156 (2018)
Manzoni P, Rinaldi M, Cattani S, Pugni L, Romeo MG, Messner H, Stolfi I, Decembrino L, Laforgia N, Vagnarelli F, Memo L, Bordignon L, Saia OS, Maule M, Gallo E, Mostert M, Magnani C, Quercia M, Bollani L, Pedicino R, Renzullo L, Betta P, Mosca F, Ferrari F, Magaldi R, Stronati M, Farina D. Bovine lactoferrin supplementation for prevention of late-onset sepsis in very low-birth-weight neonates: a randomized trial. Journal of the American Medical Association. 302: 1421-1428 (2009)
Manzoni P, Meyer M, Stolfi I, Rinaldi M, Cattani S, Pugni L, Romeo MG, Messner H, Decembrino L, Laforgia N, Vagnarelli F, Memo L, Bordignon L, Maule M, Gallo E, Mostert M, Quercia M, Bollani L, Pedicino R, Renzullo L, Betta P, Ferrari F, Alexander T, Magaldi R, Farina D, Mosca F, Stronati M. Bovine lactoferrin supplementation for prevention of necrotizing enterocolitis in very-low-birth-weight neonates: a randomized clinical trial. Early Human Development. 90 Suppl 1: S60-65 (2014)
Mao X, He S, Zhang T, Guo X, Ge Y, Ma C, Zhang X. Isolation and characterization of antiproliferative peptides from Chinese three-striped box turtle (Cuora trifasciata). Biotechnology and Applied Biochemistry. 64: 827-835 (2017)
Mao M, Chen Y, Jia Y, Yang J, Wei Q, Li Z, Chen L, Chen C, Wang L. PLCA8 suppresses breast cancer apoptosis by activating the PI3k/AKT/NF-kappaB pathway. Journal of Cellular and Molecular Medicine. 23: 6930-6941 (2019)
Marcela GM, Eva RG, Del Carmen RM, Rosalva ME. Evaluation of the antioxidant and antiproliferative effects of three peptide fractions of germinated soybeans on breast and cervical cancer cell lines. Plant Foods for Human Nutrition. 71: 368-374 (2016)
Memariani H, Memariani M. Anti-fungal properties and mechanisms of melittin. Applied Microbiology and Biotechnology. 104: 6513-6526 (2020)
Mihara M, Erster S, Zaika A, Petrenko O, Chittenden T, Pancoska P, Moll UM. p53 has a direct apoptogenic role at the mitochondria. Molecular Cell. 11: 577-590 (2003)
Mir Hassani Z, Nabiuni M, Parivar K, Abdirad S, Karimzadeh L. Melittin inhibits the expression of key genes involved in tumor microenvironment formation by suppressing HIF-1alpha signaling in breast cancer cells. Medical Oncology. 38: 77 (2021)
Monastra G, Sambuy Y, Ferruzza S, Ferrari D, Ranaldi G. Alpha-lactalbumin Effect on Myo-inositol Intestinal Absorption: In vivo and In vitro. Curr Drug Deliv. 15: 1305-1311 (2018)
Mose M, Moller N, Jessen N, Mikkelsen UR, Christensen B, Rakvaag E, Hartmann B, Holst JJ, Jorgensen JOL, Rittig N. beta-Lactoglobulin Is Insulinotropic Compared with Casein and Whey Protein Ingestion during Catabolic Conditions in Men in a Double-Blinded Randomized Crossover Trial. Journal of Nutrition. 151: 1462-1472 (2021)
Motiei M, Aboutalebi F, Forouzanfar M, Dormiani K, Nasr-Esfahani MH, Mirahmadi-Zare SZ. Smart co-delivery of miR-34a and cytotoxic peptides (LTX-315 and melittin) by chitosan based polyelectrolyte nanocarriers for specific cancer cell death induction. Materials Science & Engineering. C: Materials for Biological Applications. 128: 112258 (2021)
Moyer TB, Heil LR, Kirkpatrick CL, Goldfarb D, Lefever WA, Parsley NC, Wommack AJ, Hicks LM. PepSAVI-MS reveals a proline-rich antimicrobial peptide in Amaranthus tricolor. Journal of Natural Products. 82: 2744-2753 (2019)
Najm AAK, Azfaralariff A, Dyari HRE, Othman BA, Shahid M, Khalili N, Law D, Syed Alwi SS, Fazry S. Anti-breast cancer synthetic peptides derived from the Anabas testudineus skin mucus fractions. Scientific Reports. 11: 23182 (2021)
Ng TB, Cheung RC, Wong JH, Wang Y, Ip DT, Wan DC, Xia J. Antiviral activities of whey proteins. Applied Microbiology and Biotechnology. 99: 6997-7008 (2015)
Nieberler M, Reuning U, Reichart F, Notni J, Wester HJ, Schwaiger M, Weinmuller M, Rader A, Steiger K, Kessler H. Exploring the Role of RGD-Recognizing Integrins in Cancer. Cancers. 9 (2017)
Ochoa TJ, Chea-Woo E, Baiocchi N, Pecho I, Campos M, Prada A, Valdiviezo G, Lluque A, Lai D, Cleary TG. Randomized double-blind controlled trial of bovine lactoferrin for prevention of diarrhea in children. Journal of Pediatrics. 162: 349-356 (2013)
Ochoa TJ, Zegarra J, Cam L, Llanos R, Pezo A, Cruz K, Zea-Vera A, Carcamo C, Campos M, Bellomo S, Group NR. Randomized controlled trial of lactoferrin for prevention of sepsis in peruvian neonates less than 2500 g. Pediatric Infectious Disease Journal. 34: 571-576 (2015)
Pabona JM, Dave B, Su Y, Montales MT, de Lumen BO, de Mejia EG, Rahal OM, Simmen RC. The soybean peptide lunasin promotes apoptosis of mammary epithelial cells via induction of tumor suppressor PTEN: similarities and distinct actions from soy isoflavone genistein. Genes & Nutrition. 8: 79-90 (2013)
Park BT, Na KH, Jung EC, Park JW, Kim HH. Antifungal and anticancer activities of a protein from the mushroom Cordyceps militaris. Korean Journal of Physiology & Pharmacology. 13: 49-54 (2009)
Parsley NC, Kirkpatrick CL, Crittenden CM, Rad JG, Hoskin DW, Brodbelt JS, Hicks LM. PepSAVI-MS reveals anticancer and antifungal cycloviolacins in Viola odorata. Phytochemistry. 152: 61-70 (2018)
Permyakov EA. alpha-lactalbumin, amazing calcium-binding protein. Biomolecules. 10: 1210 (2020)
Pushparajah V, Fatima A, Chong CH, Gambule TZ, Chan CJ, Ng ST, Tan CS, Fung SY, Lee SS, Tan NH, Lim RL. Characterisation of a new fungal immunomodulatory protein from tiger milk mushroom, Lignosus rhinocerotis. Scientific Reports. 6: 30010 (2016)
Qiao X. The antitumor activity and mechanism of HN-1 from Amolops hainanensis on breast cancer. Docter Docter Thesis, Dalian University of Technology. (2019) (in Chinese)
Qiao X, Yang H, Gao J, Cai S, Shi N, Wang M, Wang Y, Yu H. A small cytotoxic peptide from frog elicits potent antitumor immunity to prevent local tumor growth and metastases. Future Medicinal Chemistry. 11: 2505-2525 (2019)
Raghuraman H, Chattopadhyay A. Melittin: A membrane-active peptide with diverse functions. Bioscience Reports. 27: 189-223 (2007)
Rayaprolu SJ, Hettiarachchy NS, Horax R, Phillips GK, Mahendran M, Chen P. Soybean peptide fractions inhibit human blood, breast and prostate cancer cell proliferation. Journal of Food Science and Technology. 54: 38-44 (2017)
Ristimaki A, Sivula A, Lundin J, Lundin M, Salminen T, Haglund C, Joensuu H, Isola J. Prognostic significance of elevated cyclooxygenase-2 expression in breast cancer. Cancer Research. 62: 632-635 (2002)
Russnes HG, Lingjaerde OC, Borresen-Dale AL, Caldas C. Breast cancer molecular stratification: From intrinsic subtypes to integrative clusters. American Journal of Pathology. 187: 2152-2162 (2017)
Russo R, Karadja E, De Seta F. Evidence-based mixture containing Lactobacillus strains and lactoferrin to prevent recurrent bacterial vaginosis: a double blind, placebo controlled, randomised clinical trial. Benef Microbes. 10: 19-26 (2019)
Schweizer F. Cationic amphiphilic peptides with cancer-selective toxicity. European Journal of Pharmacology. 625: 190-194 (2009)
Ser HL, Palanisamy UD, Yin WF, Abd Malek SN, Chan KG, Goh BH, Lee LH. Presence of antioxidative agent, Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro- in newly isolated Streptomyces mangrovisoli sp. nov. Frontiers in Microbiology. 6: 854 (2015)
Shang D, Yu F, Li J, Zheng J, Zhang L, Li Y. Molecular cloning of cDNAs encoding antimicrobial peptide precursors from the skin of the Chinese brown frog, Rana chensinensis. Zoological Science. 26: 220-226 (2009)
Shaw P, Kumar N, Hammerschmid D, Privat-Maldonado A, Dewilde S, Bogaerts A. Synergistic effects of melittin and plasma treatment: A promising approach for cancer therapy. Cancers. 11: 1109 (2019)
Shi Y, Zhang Y, Lai N, Lin L, Jiang S, Lu J. Research on purification and identification of anti-tumor bioactive peptides obtained from Chinemys reevesii. Modern Food Science and Technology. 34: 1-8 (2018) (in Chinese)
Simoneschi D, Rona G, Zhou N, Jeong YT, Jiang S, Milletti G, Arbini AA, O'Sullivan A, Wang AA, Nithikasem S, Keegan S, Siu Y, Cianfanelli V, Maiani E, Nazio F, Cecconi F, Boccalatte F, Fenyo D, Jones DR, Busino L, Pagano M. CRL4(AMBRA1) is a master regulator of D-type cyclins. Nature. 592: 789-793 (2021)
Smedegaard SB, Mose M, Hulman A, Mikkelsen UR, Moller N, Wegener G, Jessen N, Rittig N. beta-Lactoglobulin Elevates Insulin and Glucagon Concentrations Compared with Whey Protein-A Randomized Double-Blinded Crossover Trial in Patients with Type Two Diabetes Mellitus. Nutrients. 13 (2021)
Sun M. Study of purification, antitumor and safty of ussurin from Gloydius ussuriensis. Master Master Thesis, Jilin University. (2013) (in Chinese)
Sun J, Chen QJ, Cao QQ, Wu YY, Xu LJ, Zhu MJ, Ng TB, Wang HX, Zhang GQ. A laccase with antiproliferative and HIV-I reverse transcriptase inhibitory activities from the mycorrhizal fungus Agaricus placomyces. Journal of Biomedicine and Biotechnology. 2012: 736472 (2012)
Suski JM, Braun M, Strmiska V, Sicinski P. Targeting cell-cycle machinery in cancer. Cancer Cell. 39: 759-778 (2021)
Tan H, Zhang M, Xu L, Zhang X, Zhao Y. Gypensapogenin H suppresses tumor growth and cell migration in triple-negative breast cancer by regulating PI3K/AKT/NF-kappaB/MMP-9 signaling pathway. Bioorganic Chemistry. 126: 105913 (2022)
Tao J, Tan Z, Diao L, Ji Z, Zhu J, Chen W, Hu Y. Co-delivery of dihydroartemisinin and docetaxel in pH-sensitive nanoparticles for treating metastatic breast cancer via the NF-kappaB/MMP-2 signal pathway. RSC Advances. 8: 21735-21744 (2018)
Tran TV, Siniavin AE, Hoang AN, Le MTT, Pham CD, Phung TV, Nguyen KC, Ziganshin RH, Tsetlin VI, Weng CF, Utkin YN. Phospholipase A2 from krait Bungarus fasciatus venom induces human cancer cell death in vitro. PeerJ. 7: e8055 (2019)
Turashvili G, Brogi E. Tumor heterogeneity in breast cancer. Frontiers in Medicine. 4: 227 (2017)
Tzifi F, Economopoulou C, Gourgiotis D, Ardavanis A, Papageorgiou S, Scorilas A. The role of BCL2 family of apoptosis regulator proteins in acute and chronic leukemias. Advances in Hematology. 2012: 524308 (2012)
Ullah R, Rehman NU, Jamshidi-Adegani F, Bari A. Editorial: Medicinal plants and marine-derived natural products as cancer chemopreventive agents. Frontiers in Pharmacology. 13: 900275 (2022)
Vuyyuri SB, Shidal C, Davis KR. Development of the plant-derived peptide lunasin as an anticancer agent. Current Opinion in Pharmacology. 41: 27-33 (2018)
Wang JJ. Study of effect of recombinant human lactoferrin on breast cancer and its mechanism. Doctor Doctor Thesis, Yanshan University. (2011) (in Chinese)
Wang C, Li HB, Li S, Tian LL, Shang DJ. Antitumor effects and cell selectivity of temporin-1CEa, an antimicrobial peptide from the skin secretions of the Chinese brown frog (Rana chensinensis). Biochimie. 94: 434-441 (2012)
Wang J, Li F, Tan J, Peng X, Sun L, Wang P, Jia S, Yu Q, Huo H, Zhao H. Melittin inhibits the invasion of MCF-7 cells by downregulating CD147 and MMP-9 expression. Oncology Letters. 13: 599-604 (2017)
Wang B, Timilsena YP, Blanch E, Adhikari B. Lactoferrin: Structure, function, denaturation and digestion. Critical Reviews in Food Science and Nutrition. 59: 580-596 (2019)
Wang L, Wang L, Shi X, Xu S. Chlorpyrifos induces the apoptosis and necroptosis of L8824 cells through the ROS/PTEN/PI3K/AKT axis. Journal of Hazardous Materials. 398: 122905 (2020)
Wang K, Liu J, Li YL, Li JP, Zhang R. Ubiquitination/de-ubiquitination: A promising therapeutic target for PTEN reactivation in cancer. Biochimica et Biophysica Acta - Reviews on Cancer. 1877: 188723 (2022)
Wei W, Fan XM, Jia SH, Zhang XP, Zhang Z, Zhang XJ, Zhang JX, Zhang YW. Sea cucumber intestinal peptide induces the apoptosis of MCF-7 cells by inhibiting PI3K/AKT pathway. Frontiers in Nutrition. 8: 763692 (2021)
Wong JH, Ng TB, Wang H, Sze SC, Zhang KY, Li Q, Lu X. Cordymin, an antifungal peptide from the medicinal fungus Cordyceps militaris. Phytomedicine. 18: 387-392 (2011)
Wu Y, Xu H, Zhang X, Feng J. Purification and characterization of anti-tumor peptides from venom of Zhejiang Agkistrodon acutus. Chinese Journal of Pharmaceuticals. 43: 428-431 (2012) (in Chinese)
Wu D, Gao Y, Qi Y, Chen L, Ma Y, Li Y. Peptide-based cancer therapy: Opportunity and challenge. Cancer Letters. 351: 13-22 (2014)
Xie Q, Yang Z, Huang X, Zhang Z, Li J, Ju J, Zhang H, Ma J. Ilamycin C induces apoptosis and inhibits migration and invasion in triple-negative breast cancer by suppressing IL-6/STAT3 pathway. Journal of Hematology & Oncology. 12: 60 (2019)
Xu D. The physiological activity of walnut polypeptde fermented by Aspergllus niger. Master Master Thesis, Beijing Forestry University. (2014) (in Chinese)
Xu WJ, Liao XJ, Xu SH, Diao JZ, Du B, Zhou XL, Pan SS. Isolation, structure determination, and synthesis of galaxamide, a rare cytotoxic cyclic pentapeptide from a marine algae Galaxaura filamentosa. Organic Letters. 10: 4569-4572 (2008)
Xu H, Kong YY, Chen X, Guo MY, Bai XH, Lu YJ, Li W, Zhou XW. Recombinant FIP-gat, a fungal immunomodulatory protein from Ganoderma atrum, induces growth inhibition and cell death in breast cancer cells. Journal of Agricultural and Food Chemistry. 64: 2690-2698 (2016)
Xue ZH, Wen HC, Zhai LJ, Yu YQ, Li YN, Yu WC, Cheng AQ, Wang C, Kou XH. Antioxidant activity and anti-proliferative effect of a bioactive peptide from chickpea (Cicer arietinum L.). Food Research International. 77: 75-81 (2015)
Xue Z, Hou X, Yu W, Wen H, Zhang Q, Li D, Kou X. Lipid metabolism potential and mechanism of CPe-III from chickpea (Cicer arietinum L.). Food Research International. 104: 126-133 (2018)
Yan J, Yang S, Tian H, Zhang Y, Zhao H. Copanlisib promotes growth inhibition and apoptosis by modulating the AKT/FoxO3a/PUMA axis in colorectal cancer. Cell Death & Disease. 11: 943 (2020)
Yap HYY, Tan NH, Ng ST, Tan CS, Fung SY. Molecular attributes and apoptosis-inducing activities of a putative serine protease isolated from tiger milk mushroom (Lignosus rhinocerus) sclerotium against breast cancer cells in vitro. PeerJ. 6: e4940 (2018)
Yen C, Zhao F, Yu Z, Zhu X, Li CG. Interactions between natural products and tamoxifen in breast cancer: A comprehensive literature review. Frontiers in Pharmacology. 13: 847113 (2022)
Yin C, Knudson CM, Korsmeyer SJ, Van Dyke T. Bax suppresses tumorigenesis and stimulates apoptosis in vivo. Nature. 385: 637-640 (1997)
You L, Zhao M, Liu RH, Regenstein JM. Antioxidant and antiproliferative activities of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Journal of Agricultural and Food Chemistry. 59: 7948-7953 (2011)
Zeng XC, Zhou L, Shi W, Luo X, Zhang L, Nie Y, Wang J, Wu S, Cao B, Cao H. Three new antimicrobial peptides from the scorpion Pandinus imperator. Peptides. 45: 28-34 (2013)
Zeng X, Zheng P, Cao W, Gong L, Liu Y. Analysis of the chemical constituents of Agkistrodon acutus veom and their antitumor activity screening. China Pharmacist. 25: 439-444 (2022) (in Chinese)
Zhang GQ, Sun J, Wang HX, Ng TB. A novel lectin with antiproliferative activity from the medicinal mushroom Pholiota adiposa. Acta Biochimica Polonica. 56: 415-421 (2009)
Zhang G, Sun J, Wang H, Ng TB. First isolation and characterization of a novel lectin with potent antitumor activity from a Russula mushroom. Phytomedicine. 17: 775-781 (2010)
Zhang YY, Kong LQ, Zhu XD, Cai H, Wang CH, Shi WK, Cao MQ, Li XL, Li KS, Zhang SZ, Chai ZT, Ao JY, Ye BG, Sun HC. CD31 regulates metastasis by inducing epithelial-mesenchymal transition in hepatocellular carcinoma via the ITGB1-FAK-Akt signaling pathway. Cancer Letters. 429: 29-40 (2018)
Zhao S, Zhao Y, Li S, Zhang G, Wang H, Ng TB. An antiproliferative ribonuclease from fruiting bodies of the wild mushroom Russula delica. Journal of Microbiology and Biotechnology. 20: 693-699 (2010)
Zhao H, Li J, Zhang Y, Lei S, Zhao X, Shao D, Jiang C, Shi J, Sun H. Potential of iturins as functional agents: safe, probiotic, and cytotoxic to cancer cells. Food & Function. 9: 5580-5587 (2018)
Zhao H, Yan L, Guo L, Sun H, Huang Q, Shao D, Jiang C, Shi J. Effects of Bacillus subtilis iturin A on HepG2 cells in vitro and vivo. AMB Express. 11: 67 (2021)
Zheng Q, Qiu D, Liu X, Zhang L, Cai S, Zhang X. Antiproliferative effect of Dendrobium catenatum Lindley polypeptides against human liver, gastric and breast cancer cell lines. Food & Function. 6: 1489-1495 (2015)
Zheng K, Li Q, Lin D, Zong X, Luo X, Yang M, Yue X, Ma S. Peptidomic analysis of pilose antler and its inhibitory effect on triple-negative breast cancer at multiple sites. Food & Function. 11: 7481-7494 (2020)
Zhou H, Zhu K, Guo X, Zhou H. Purification and anti-tumor activity of wheat germ anti-tumor activity protein. China Oils and Fats. 38: 23-27 (2013) (in Chinese)
Zhou W, Fang H, Wu Q, Wang X, Liu R, Li F, Xiao J, Yuan L, Zhou Z, Ma J, Wang L, Zhao W, You H, Ju J, Feng J, Chen C. Ilamycin E, a natural product of marine actinomycete, inhibits triple-negative breast cancer partially through ER stress-CHOP-Bcl-2. International Journal of Biological Sciences. 15: 1723-1732 (2019)
Zhou SD, Lin YF, Xu X, Meng L, Dong MS. Effect of non-covalent and covalent complexation of (-)-epigallocatechin gallate with soybean protein isolate on protein structure and in vitro digestion characteristics. Food Chemistry. 309: 125718 (2020a)
Zhou Y, Zhang S, Chen Z, Bao Y, Chen AT, Sheu WC, Liu F, Jiang Z, Zhou J. Targeted delivery of secretory promelittin via novel poly(lactone-co-beta-amino ester) nanoparticles for treatment of breast cancer brain metastases. Advanced Science. 7: 1901866 (2020b)
Zhu J, Cai T, Zhou J, Du W, Zeng Y, Liu T, Fu Y, Li Y, Qian Q, Yang XH, Li Q, Huang JA, Liu Z. CD151 drives cancer progression depending on integrin alpha3beta1 through EGFR signaling in non-small cell lung cancer. Journal of Experimental and Clinical Cancer Research. 40: 192 (2021)
Zhu S, Li X, Dang B, Wu F, Gou K, Wang C, Lin C. Hydrogen sulfide protects retina from blue light-induced photodamage and degeneration via inhibiting ROS-mediated ER stress-CHOP apoptosis signal. Redox Report. 27: 100-110 (2022)
Acknowledgements
This work was supported by the Collaborative Grant-in-Aid of the HBUT National “111” Center for Cellular Regulation and Molecular Pharmaceutics [Grant Number XBTK-2021006]; the Open Project Funding of the Key Laboratory of Fermentation Engineering (Ministry of Education) [Grant Number 202105FE02]; the Hubei University of Technology [Grant Number BSQD2020038]; and the Shanghai Science and Technology Innovation Action Plan [Grant Number 19431901700].
Author information
Authors and Affiliations
Contributions
Q-ZL: conceptualization; methodology; formal analysis; investigation; writing—original draft; writing—review & editing; funding acquisition. Z-RZ: formal analysis; investigation; writing—original draft; visualization. C-YH: writing—original draft. X-BL: writing—original draft. Y-ZC: writing—original draft. YL: visualization; funding acquisition. Y-LW: methodology; formal analysis; writing—review & editing. X-WZ: methodology; formal analysis; writing—review & editing. All authors read and approved the final version of the work to be published.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Li, QZ., Zhou, ZR., Hu, CY. et al. Recent advances of bioactive proteins/polypeptides in the treatment of breast cancer. Food Sci Biotechnol 32, 265–282 (2023). https://doi.org/10.1007/s10068-022-01233-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10068-022-01233-6