
Abstract
Ecological interactions, especially those that are beneficial (i.e. mutualism) or detrimental (i.e. parasitism), play important roles during the establishment and spread of alien species. This chapter explores the role of these interactions during biological invasions in South Africa, covering a wide range of taxonomic groups and interaction types. We first discuss the different ways in which interactions can be reassembled following the introduction of alien species, and how these depend on the eco-evolutionary experience of the alien species. We then discuss documented examples of parasitism and mutualism associated with invasions in South Africa and how these relate to various ecological and evolutionary hypotheses aimed at explaining species invasiveness. Selected examples of how invasive species impact on native species interactions are provided. A diverse array of biotic interactions (e.g. pollination, fish and mollusc parasitism, plant-soil mutualistic bacteria, seed dispersal) have been studied for various invasive species in South Africa. Surprisingly, only a few of these studies explicitly tested any of the major hypotheses that invoke biotic interactions and are commonly tested in invasion ecology. We argue that many invasions in South Africa are promising candidates for testing hypotheses related to species interactions and invasiveness.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

1 Introduction
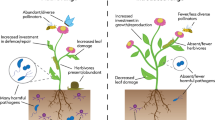
All organisms interact, directly or indirectly , with other organisms in the environments in which they find themselves. Direct interactions may benefit both interacting partners (i.e. mutualism ), benefit only one partner (i.e. commensalism), benefit one partner at the expense of the other (parasitism) , or may have no effect on one or both partners (Fig. 14.1). Symbiotic interactions imply that interacting organisms live in close physical association with each other for a significant portion of their lives, and brief interactions like predation, are therefore not viewed as symbiotic. Together with abiotic environmental conditions, biotic interactions shape the diversity, structure, and function that underlie biological communities (Post and Palkovacs 2009). Consequently, biological invasions present unique opportunities to explore the processes that govern the assembly of these interactions and their impact on population demography and community structure. Many hypotheses in invasion ecology invoke biotic interactions (Table 14.1), though ultimately, they come down to the same three processes (mutualism , commensalism or parasitism ): during the introduction process, some interactions might be lost if there is no co-introduction , but novel interactions might develop through ecological fitting or co-xenic associations .

Different types of ecological interactions based on the benefit, harm or neutral effects on interacting partners
1.1 Ecological Fitting, Co-xenic Associations, and Co-introductions
The act of moving a species across a biogeographical barrier often means that it will lose key biotic interactions that were present in its native range, but experience a whole suite of new interactions in its alien range. These effects might enhance performance in the new environment, or provide obstacles to establishment and subsequent success (Enders et al. 2018). For example, a reduction in, or more frequently, the total absence of, specialist enemies following introduction can allow individuals of a species in the alien range to realise greater reproductive output than individuals in their native source populations (the Enemy Release Hypotheses —ERH, Colautti et al. 2004; see Table 14.1 for a summary of the hypotheses mentioned throughout this chapter). In general, alien species can reassemble biotic interactions through: (1) novel associations with organisms native to the new environment (so-called ecological fitting; Le Roux et al. 2017); (2) associations between organisms that are both alien to the new environment, but that do not co-occur in their respective native ranges (so-called co-xenic associations; Nuñez and Dickie 2014); or (3) co-introduction of interacting partners (the so-called co-introduction pathway; Le Roux et al. 2017).
All biotic interactions span a continuum of specificity from the viewpoint of both interacting partners. At the one end of the spectrum, highly specialised interactions are characterised by those restricted to two species, or even biotypes. On the other hand, some organisms can interact effectively with a range of different partners, i.e. being generalists. For mutualistic interactions required for the successful completion of an organism’s life cycle, such as pollination , levels of specialisation will have significant impacts on the establishment success of alien species following introduction into new environments when not co-introduced. The loss of highly specialised mutualists may hamper establishment success (i.e. Missed Mutualisms Hypothesis ; Catford et al. 2009). This was the case for many Pinus species introduced to the southern Hemisphere in previous centuries, where invasions only occurred after pine-specific mycorrhizal fungi were introduced (e.g. Richardson et al. 1994). The loss of generalist interactions intuitively poses less pivotal constraints as these can potentially be replaced by novel interactions through ecological fitting (see Heleno et al. 2012 for seed dispersal example) .
As posited by the ERH , the liberation from highly specialised parasitic interactions (such as herbivores or pathogens ) will aid establishment success. More than a century of biological control of invasive plants in South Africa provides strong support for the role of enemy release in plant invasiveness (Zachariades et al. 2017; also see Hill et al. 2020, Chap. 19), although the high levels of control observed can equally be explained by the biological control agents having been released from their natural enemies. Levels of interaction specificity are also important when considering interactions between the alien species and resident species (e.g. symbionts) in the new range, i.e. ecological fitting. That is, establishment success and invasive performance are expected to be enhanced when resident antagonists or predators are highly specialised and/or resident mutualists are generalist (so called Specialist-Generalist Hypothesis, Catford et al. 2009).
The different pathways for interaction reassembly (ecological fitting , co-introduction vs. co-xenic) can have distinct impacts on the establishment success of aliens, and many of these have been formally described as hypotheses in invasion ecology. For example, ecological fitting may either enhance or impede the performance of introduced species (so-called New Associations Hypothesis; Catford et al. 2009) , while co-xenic associations may lead to invasional meltdown , whereby positive interactions among different invasive species initiate feedbacks that intensify their ecosystem impacts and/or promote secondary invasions by other species (Simberloff and Von Holle 1999). Co-introduction of mutualists almost always benefits invaders. In some instances, co-introduced enemies may be less effective, or may even have an opposite effect, in the new environment (i.e. Enemy Inversion Hypothesis ; Catford et al. 2009). The Enemy of my Enemy Hypothesis can operate through apparent competition , whereby the enemy ends up causing more damage to maladapted native species than the alien species, potentially reducing inter-specific competition between invasive and resident species (Catford et al. 2009). The outcomes of an introduction (i.e. invasiveness) will therefore to a large degree depend on the structure of ecological interaction networks in both native and non-native communities (Fig. 14.2).
1.2 The Structure of Ecological Interaction Networks and Their Infiltration by Invasive Species
Ecological networks with interactions varying in their specificity can show high levels of nestedness, e.g. if specialist plants in a community only interact with a subset of the pollinators with which generalist plants interact (Bascompte 2009). Specialisation also means that species pairs may not have the same chances for interacting. For example, networks will become modular when host plants only interact with mutualists that they share a co-evolutionary history with , and vice versa (Bascompte 2009). Therefore, modularity and/or nestedness of networks are dependent on the prevalence of interaction specialisation within communities.
Following introduction, the reassembly of beneficial mutualistic interactions are clearly more important for successful establishment and subsequent invasion than the formation of harmful antagonistic interactions. So how do aliens typically infiltrate existing host-mutualist ecological networks in their new ranges? Empirical evidence suggests that they are often generalist species and this allows them to utilise existing mutualists found in their new ranges (i.e. web infiltration through ecological fitting , e.g. Aizen et al. 2012; Fig. 14.2). On the other hand, highly specialised hosts, accompanied by their mutualists through co-introduction , can integrate into native community networks as novel modules, consisting of interacting (co-introduced) taxa that are not present in native interaction webs (e.g. Le Roux et al. 2016; Fig. 14.2). This complexity might seem to preclude prediction, but there are often some general phylogenetic patterns. For example, in South Africa an interaction network between legumes and rhizobia found invasive acacias to form strong interaction modules, resulting from interactions between acacias and Bradyrhizobium strains. Native South African legumes (outside acacia-rhizobium modules) rarely associated with bradyrhizobia, but largely with phylogenetically distinct rhizobia (Le Roux et al. 2016).

Predictions of how mutualist co-introduction versus ecological fitting , in conjunction with interaction specialisation, may allow alien species interaction web infiltration (adapted from Le Roux et al. 2017). (I) Communities in both native and invasive ranges (pre-introduction) will have interaction webs containing both specialist and generalist taxa. (II) Strong interaction modules may emerge following the co-introduction of a highly specialised host and its mutualist, (III) whereas co-introduced generalists are expected to form novel associations (red lines) to replace those lost during introduction (dashed lines). (IV) Ecological fitting by generalist hosts will only involve novel associations while, (V) ecological fitting of a highly specialist host may lead to no interactions and possibly establishment failure of the introduced species
1.3 Eco-evolutionary Experience and Biological Invasions
Other than specialisation , the phylogenetic composition of recipient communities can impact on the rate and nature of interaction reassembly, alien species establishment success, and ultimately, invasion. That is, when organisms are introduced into environments with phylogenetically closely-related congeners, the accumulation of interactions (especially more specialised associations) is expected to occur more rapidly compared to environments lacking closely-related taxa (Darwin 1859; Daehler 2001). Related to this, the successful ecological integration of aliens into novel community contexts will also rely on the eco-evolutionary experience of both the introduced species and the recipient community. That is, historical evolution that has shaped a species’ adaptations to biotic interactions (ecology) in its native range will be the basis for ease of integration into novel ecological contexts, such as those underlying species invasions (Saul et al. 2013). The same applies to native species’ responses and eco-evolutionary experience with the newly arriving species. Therefore, attributes of eco-evolutionary experience on either side, alien versus native, can be interpreted in terms of the alien species’ invasiveness and the native community’s invasibility , respectively (Saul et al. 2013).
Integration of alien species into novel community contexts is then itself an ongoing process that will change over time and likely operate over both short and long evolutionary timescales, and will be influenced by residence time , i.e. time since introduction (e.g. Heleno et al. 2012). It is therefore expected that different insights might emerge when assessing the role of biotic interactions in allowing alien species to transition along different stages of the introduction-naturalisation-invasion continuum. While ecological integration is key to becoming invasive, interactions that are lost following species introductions are obviously important, such as release from specialist enemies. However, these losses may only be temporary. For widely established invasive plants, for example, interactions with resident and native herbivores and pathogens are expected to accumulate over time (e.g. Crous et al. 2017; Stricker et al. 2016), and the evolutionary component of such interactions has now been demonstrated in many cases (e.g. Strauss et al. 2006). The incidence and extent of such ‘catch-ups’ are expected to increase with residence time , as host abundance, and thus a possible unexploited resource and its discovery, increases (Carroll et al. 2005).
In this chapter, we review evidence for the role biotic interactions play in biological invasions in South Africa. Focusing on parasitism and mutualism , we aim to summarise evidence from South Africa in support of various ecological and evolutionary hypotheses put forward to explain species invasiveness and that invoke biotic interactions. We do not treat commensalism in depth here, as this interaction type is generally not clearly linked to any of the major ecological and evolutionary hypotheses related to invasibility/invasiveness or hardly studied, and neither do we focus on invasions in urban ecosystems (see Box 14.1 for a discussion of what has been termed human commensals). We do, however, discuss how native species interactions are impacted as alien species are integrated into the communities they invade. Lastly, we discuss the future directions for biotic interaction research on biological invasions in South Africa.
Box 14.1 Human Commensals
Humans are clearly a fundamental driver and mediator of invasions. Alien species, by definition, owe their presence in an area to human-mediated introduction (Richardson et al. 2000c). But humans play crucial, often dominant, roles at all stages of the introduction-naturalisation -invasion continuum (sensu Richardson and Pyšek 2012) by, among other things: mediating potential abiotic barriers; reshuffling the biotas of ecosystems (thereby potentially mediating biotic barriers posed through competition and other factors); affecting within-region dispersal in many ways (through accidental dispersal during human activities, cultivation and propagation, e.g. for aquaculture, mariculture, ornamental horticulture, forestry, and the pet trade); and by attempts to manage these species. The roles of humans in disseminating alien species in different taxonomic groups, and the diverse effects of people in mediating the abundance, distribution and impacts of these species are detailed in other chapters of this book (see Potgieter et al. 2020, Chap. 11; Faulkner et al. 2020, Chap. 12). One aspect that requires attention here, however, relates to urban environments and the confinement of many alien species to such environments. This implies that many alien species directly or indirectly interact with humans to ensure their existence in urban environments. Urban ecosystems in South Africa are, like those elsewhere in the world, hotspots for the arrival of alien species. These ecosystems also have the highest species richness of alien taxa of all habitats. Reasons for this include the demand for alien species for many purposes, high levels of propagule pressure , concentrated opportunities and conduits for dispersal, high levels of disturbance , and the diversity of habitats and niches provided by human activities. Many widespread invasive alien species in mainland South Africa are virtually confined to urban ecosystems. For example, bird invasions in South Africa are unusual in that all seven alien species with viable populations are strongly commensal with humans and none of the 48 alien bird species in South Africa has established viable populations in natural ecosystems (Richardson et al. 2011; Measey et al. 2020, Chap. 5). This is an ongoing natural experiment, but evidence suggests that while human-built environments provide many opportunities for alien birds, combined biotic and abiotic pressures present a strong barrier to the invasion of natural ecosystems in South Africa. Most invasive mammals in South Africa are also strongly commensal with humans; examples include the Grey Squirrel (Sciurus carolinensis), the House Mouse (Mus musculus), the Norway Rat (Rattus norvegicus), the Black Rat (Rattus rattus), and the Asian House Rat (Rattus tanezumi) (Richardson et al. 2011; Measey et al. 2020, Chap. 5). Many widespread alien plants also seem to be confined to human-dominated ecosystems, for example Tree of Heaven (Ailanthus altissima; Walker et al. 2017) and Red Valerian (Centranthus ruber; Geerts et al. 2017; Holmes et al. 2018). Urban areas provide important habitats for many other alien plants, and may act as beachheads for invasion into natural systems; they provide opportunities for species to accumulate high propagule pressure to drive invasions beyond the urban-wildland interface (e.g. Alston and Richardson 2006; Foxcroft et al. 2008; Donaldson et al. 2014). Urban invasion ecology has only recently begun to be studied in South Africa and much more work remains to be done to elucidate the ecology of urban invaders (see Potgieter et al. 2020, Chap. 11).
2 Parasitism
2.1 Plants
As mentioned above, phylogenetic similarity of recipient communities to alien species may impact on the rate and nature of interaction reassembly. Under ecological fitting , recipient communities harbouring phylogenetically closely related taxa, or alien species with high eco-evolutionary experience, could possibly facilitate spillovers of enemies from the recipient community onto the alien species, except if these lack high eco-evolutionary experience to the invader. Evidence for such spillover in South Africa comes from Crous et al. (2017). These authors found that, irrespective of residence time , pathogen accumulation of alien pines (genus Pinus), Australian wattles (genus Acacia), and eucalypts (genus Eucalyptus) was highest in taxa most closely related to the South African flora. That is, pines, with no confamilial relatives in South Africa, have acquired only one highly polyphagous pathogen despite the long residence time of the genus in the country (>300 years). On the other hand, wattles and eucalypts, both with confamilial relatives in South Africa have accumulated many pathogens since their introduction (Crous et al. 2017). For example, the fungus Chrysoporthe austroafricana, a pathogen of the South African Water Berry Tree, Syzygium cordatum (Heath et al. 2006), has caused serious stem canker disease on introduced eucalypts (Wingfield et al. 1989). In contrast, patterns of accumulation of insect pests in these three plant genera do not seem to be associated with the phylogenetic relatedness of these genera to South African plants. In line with the New Associations Hypothesis, these associations appear random and exclusively involve generalist (highly polyphagous) insect pests (Crous et al. 2017). Despite this, high abundances of South African herbivores have been found in association with alien trees (Procheş et al. 2008). For example, the native Keurboom (Virgilia divaricata) shares up to 30% of its associated arthropod community with the confamilial invasive Black Wattle, Acacia mearnsii (van der Colff et al. 2015), while the native polyphagous moth, Imbrasia cytherea (Pine Tree Emperor Moth), is a common pest on introduced pines (Roux et al. 2012). Similarly, the native seed-feeding alydid bug, Zulubius acaciaphagus, is commonly found feeding on the invasive Rooikrans, Acacia cyclops (Holmes and Rebelo 1988).
In some instances, co-xenic associations may exacerbate invader ecosystem impacts or even facilitate secondary invasions by other species, i.e. Invasional Meltdown (Simberloff and Von Holle 1999). For example, Puccinia psidii, a myrtle rust fungus native to South and Central America (Coutinho et al. 1998) is now commonly associated with alien Myrtaceae taxa, including eucalypts (Glen et al. 2007). This pathogen has now spilled over onto native forest Myrtaceae species in South Africa (Roux et al. 2015).
2.2 Marine Ecosystems
In South Africa, alien marine molluscs are often parasitised by endolithic bacteria resulting in bioerosion and causing severe shell damage (Prenter et al. 2004), often leading to lethal and sub-lethal impacts (Kaehler and McQuaid 1999). Along the South African coastline, high rates of endolithic parasitism have been reported in the widespread invasive Mediterranean Mussel, Mytilus galloprovincialis (Fig. 14.3d; Zardi et al. 2009; Marquet et al. 2013). Studies comparing the effects of endoliths between the native South African Green Mussel, Perna perna, and M. galloprovincialis found infected individuals of the latter to be more negatively impacted (Zardi et al. 2009), with both higher endolith incidence and greater reductions in shell thickness, shell strength, and overall condition (Zardi et al. 2009). Infected mussels also have lower attachment strength, probably because more energy is being directed toward shell repair and away from the production of byssus threads that are responsible for securing them to substrates (Kaehler and McQuaid 1999). This, and other mechanisms such as wave action and emersion stress (see Rius and McQuaid 2006, 2009), are thought to mediate competition , promoting co-existence between P. perna and M. galloprovincialis on South African rocky shores . In particular, wave action favours the abundance of P. perna on the low shore, while P. perna facilitates the establishment of M. galloprovincialis in the mid shore (resulting in mixed mussel beds). On the high shore, P. perna is excluded due to emersion stress, leaving M. galloprovincialis to dominate (Rius and McQuaid 2006, 2009).

Examples of biotic interactions during biological invasion in South Africa. (a) Hovering native Malachite Sunbird (Nectarinia famosa) pollinating invasive tree tobacco (Nicotiana glauca). (b) Root nodules formed by co-introduced nitrogen-fixing Bradyrhizobium strains on invasive Golden Wattle (Acacia pycnantha). (c) The native Citrus Swallowtail, Papilio demodocus, pollinating invasive Devil’s Beard (Centranthus ruber). (d) Invasive Mediterranean Mussels (Mytilus galloprovincialis) showing extensive shell damage and bioerosion resulting from parasitism by possibly native endolithic bacteria. (e) A native Grey-Headed Albatross (Thalassarche chrysostoma) attacked by invasive House Mice on Marion Island . (f) The cosmopolitan endoparasitoid, Dinocampus coccinellae, targeting the invasive Harlequin Ladybird, Harmonia axyridis. Photographs courtesy of (a, c) Sjirk Geerts; (b) Jan-Hendrik Keet; (d) Lisa Skein; (e) Andrea Angel; (f) Ingrid Minnaar
Although four out of the seven endolithic species parasitising M. galloprovincialis in South Africa are also found in the species’ native range (Marquet et al. 2013), it is unlikely that they were co-introduced with their host during the 1970s. This mussel likely invaded the South African coastline after being released as larvae from ballast water (Grant and Cherry 1985), which cannot vector endolithic bacteria. These parasites are therefore thought to be cosmopolitan in their distribution and native to South Africa (Marquet et al. 2013). Interestingly, endolithic parasitism appears to have a greater impact on South African M. galloprovincialis populations than on native populations in Portugal (Marquet et al. 2013), possibly due to the low genetic variability of the mussel in South Africa (Zardi et al. 2009; Marquet et al. 2013). However, despite the negative effects of shell parasites on M. galloprovincialis, this mussel persists as the most successful marine invasive species along the South African coastline (Robinson et al. 2005, 2020 , Chap. 9). Characteristics such as high fecundity and recruitment rates (van Erkom Schurink and Griffiths 1991; Harris et al. 1998), fast growth (Griffiths et al. 1992), and high desiccation tolerance (Hockey and van Erkom Schurink 1992), enable it to overcome the negative impacts imposed by parasites like endolithic bacteria. Mytilus galloprovincialis invasions also had some positive impacts on native species in South Africa. This invasive mussel now makes up a large part of the diet of the endemic African black oystercatcher (Haematopus moquini), southern Africa’s second-rarest coastal bird (Coleman and Hockey 2008).
In contrast to the post-introduction accumulation of parasites in M. galloprovincialis, the intentional introduction of molluscs for aquaculture often leads to the co-introduction of their parasites (Naylor et al. 2001). For example, ten shell-boring polychaete worm species are known to infect shells of cultured molluscs, mainly oysters and abalone, along the South African coastline (Simon and Sato-Okoshi 2015). Two of these parasites are invasive in South Africa, namely Polydora hoplura and Boccardia proboscidia (Simon et al. 2006, 2009; David and Simon 2014). The former was detected in the 1950s, while B. proboscidia was first recorded in 2004 (Simon et al. 2006, 2009). The ability of females of these two polychaetes to produce multiple larval types (poecilogonous), and to survive and reproduce across a wide range of temperatures and substrates, all contribute to their invasion success in South Africa (David and Simon 2014). The excavation of burrows on shell surfaces of molluscs by polydorid annelids such as P. hoplura and B. proboscidia, not only leads to shell damage, but also causes reduced growth and condition, and ultimately increased mortality rates (Simon et al. 2006). While such parasitism is initially limited to cultured molluscs, these organisms can escape from aquaculture facilities and infect wild molluscs. The parasites thus represent economic and ecological threats. Both species are now found along most of the South African coastline (David and Simon 2014), and although transport of cultured animals among aquaculture facilities is being more strictly regulated, the threats posed to both farmed and wild molluscs remain.
2.3 Freshwater Fish
Because of their importance in aquaculture, fisheries and the global pet trade, freshwater fishes are frequently introduced outside their native ranges as live adults or young. Therefore, at least historically, co-introductions of novel parasites and diseases into environments where they have not previously occurred were common. Some of the parasites are so specialised that they are unable to infect native fishes and their presence in the recipient environment is dependent on the presence of their co-introduced host. Those that are able to infect native hosts can have severe consequences, as native fish, lacking evolutionary history with alien parasites, do not possess immune responses to infection (Taraschewski 2006).
The introduction of 27 alien fishes to South Africa has provided opportunities for at least 23 parasitic co-introductions of ten monogeneans, eight ciliates, two cestodes, a copepod, a flagellate, and a branchiuran (Smit et al. 2017). Most (16) of these parasites are not known to have infected native fishes (Smit et al. 2017). For example, five ancyrocephalid monogeneans are found only on the alien Largemouth Bass, Micropterus salmoides. Despite the almost ubiquitous presence of bass in South African rivers (Ellender et al. 2014), these parasites have not been observed to infect native fishes to date (Truter et al. 2017 ). The other seven co-introduced parasites, however, have formed new associations with native hosts, probably because of broader levels of generalism in their symbiotic requirements (Smit et al. 2017). While significant impacts on the health status of novel hosts have been documented (see Weyl et al. 2020, Sect. 6.2, on freshwater biota and impacts), the influence of parasites on the invasion process has not been investigated in any detail in South Africa.
Some alien fishes in South Africa have considerably lower parasite loads than in their native ranges. In an assessment of the parasitism of largemouth bass for example, Truter et al. (2017) documented lower parasite abundance and richness in South Africa in comparison with native range populations. This may explain why this species managed to invade a wide range of habitats throughout southern Africa despite extremely low genetic diversity , resulting from a very limited number of propagules introduced into South Africa in the late 1920s (Hargrove et al. 2017). Similar mechanisms might be responsible for the success of rainbow trout, Onchorhynchus mykiss, and brown trout, Salmo trutta, for which there are no records of co-introduced parasites (see Weyl et al. 2020, Sect. 6.2).
2.4 Insects
South Africa’s alien entomofauna has assembled as a result of complex introduction pathways (Giliomee 2010; Garnas et al. 2016; Janion-Scheepers and Griffiths 2020, Chap. 7; Faulkner et al. 2020, Chap. 12). Most species were accidentally introduced, but a small proportion were intentionally introduced, mainly as biological control agents (Hill et al. 2020, Chap. 19). Introduction pathways have important implications for biotic interactions in the receiving environment, for example host-parasite relationships. Many other factors also play a role, including residence time of different partners in host-parasite relationships, whether the parasite has had any co-evolutionary history with the host, competition among hosts or parasites, complexity in food webs such as cascading effects in multi-trophic systems, and hosts shifts. While some alien insect hosts are parasitised by alien or native parasites, the opposite is also true, and there are examples of biological control agents infiltrating native communities. Thus, the relationships between parasite and host for insect invaders are complex, and include direct and indirect effects that shape the dynamics of whole communities. Here we focus on parasitoids, since the information on pathogens , fungi and other parasites such as nematodes in South Africa has either been reviewed elsewhere (Wingfield et al. 2001), is very scarce (Haelewaters et al. 2016), or has focused on the selection of biological control agents under controlled experimental conditions (e.g., nematodes, Malan and Moore 2016). We provide some examples of biotic interactions involving insect hosts and parasitoids for biological invasions in South Africa.
Alien insects can be parasitised by native or alien parasitoids. In South Africa, the invasive Harlequin Ladybeetle, Harmonia axyridis, a notorious predator of aphids and other coccinellid species and native to Asia (see Janion-Scheepers and Griffiths 2020, Box 7.4, Chap. 7), was first detected in the Western Cape Province in the early 2000s and then spread rapidly across the country (Stals and Prinsloo 2007; Roy et al. 2016). Beetles sourced in the USA, but originating from Japan, were intentionally introduced to South Africa to control an aphid pest in 1980, but failed to establish. Invasive populations of H. axyridis in the country are thought to have originated from a separate subsequent and accidental introduction (Roy et al. 2016). Population genetic analyses revealed that Western Cape populations originated from an invasive population in eastern North America, described as a bridgehead for the worldwide invasion of this species (Lombaert et al. 2010). The wasp Dinocampus coccinellae (Hymenoptera: Braconidae), a koinobiont endoparasitoid of coccinellid species, with a widespread global distribution, was later reported to utilise H. axyridis as a host (Fig. 14.3f), in addition to three other native and one alien host (Minnaar et al. 2014). The wasp was initially collected from native hosts in South Africa in the late 1940s and 1960s, suggesting that its occurrence preceded the introduction of H. axyridis. Interestingly, the level of parasitism by this parasitoid on H. axyridis was much lower than rates found on native hosts (Minnaar et al. 2014), and is consistent with findings from other global regions in the species’ invasive range (Comont et al. 2014; Ceryngier et al. 2018). Despite this, further work is needed to identify the mechanisms underlying the release from this native enemy in South Africa. A possible explanation is that the invasive ecotype of H. axyridis garners higher immunity or resistance to parasitoids than native species, as several studies highlight the diversity of chemical defences (harmonine and antimicrobial peptides; Röhrich et al. 2012; Vilcinskas et al. 2013a) and prevalence of obligate parasitic microsporidia in this species (Vilcinskas et al. 2013b). Nonetheless, the parasitoid may adapt further via changes in host location mechanisms or parasite developmental growth strategies (Firlej et al. 2007), increasing host suitability in the future. The fact that D. coccinellae has been consistently detected on this invasive species suggests that it may benefit from a marginal host, when for example, native species fluctuate in numbers.
Invasive insect species are often pests of agricultural plants and plantation trees, and in South Africa, alien biological control agents (including parasitic wasps) have been introduced to regulate them or have been accidentally co-introduced (e.g. in Eucalyptus plantations: Wingfield et al. 2008; Garnas et al. 2012; Bush et al. 2016). These species are also often reported to harbour a high diversity of natural enemies. For example, 22 species of parasitoids and hyperparasitoids emerged from Diamondback Moth (Plutella xylostella, a notorious pest of cultivated and native brassicas in the region) larvae and pupae sampled in South Africa (Kfir 1998). A few of the parasitoids had restricted distributions in South Africa and, together with their degree of host-specificity, suggests that host and parasite had time to co-evolve (Kfir 1998). Similarly, P. xylostella monitored on canola revealed novel associations with a large diversity of native larval and pupal parasitoids, infecting the host distinctively in terms of extent and timing of parasitism (Mosiane et al. 2003). Hyperparasitoids were also found to feed on the cocoons of primary parasitoid larvae and were influenced by the abundance and timing of the latter (Mosiane et al. 2003; Nofemela and Kfir 2005). Therefore, the complexity of these tri-trophic relationships, including potential density-dependent and cascading top-down effects, are likely to modulate the dynamics of these pests and invasive populations (Nofemela 2013).
Besides direct effects across trophic levels as discussed above, there are also examples of indirect effects in biotic interactions that affect the invasive host or, alternatively, an invasive species can also be a key player in the regulation of host-parasite interactions. For example, inter-specific competition between native and invasive ant species, including the Argentine Ant (Linepithema humile), can disrupt associations between parasitoids and hemipteran pests that produce honeydew sought by the ants (Mgocheki and Addison 2010). Argentine Ant invasions may also disrupt native plant-ant interactions, such as myrmecochorous seed dispersal , as has been found in many parts of the world where this invasive ant is present (reviewed in Traveset and Richardson 2014; see also Janion-Scheepers and Griffiths 2020, Chap. 7, Box 7.6).
Lastly, insect biotic interactions include non-target associations of insect herbivores introduced as biological control agents of invasive plants. In this case, the alien herbivore is attacked by native parasitoids, potentially reducing the level of biological control achieved on the target alien plant. For example, the bud-galling wasp, Trichilogaster acaciaelongifoliae, introduced in 1982 to South Africa from Australia to control Acacia longifolia quickly acquired novel communities of natural parasitoids (Manongi and Hoffmann 1995; McGeoch and Wossler 2000; Veldtman et al. 2011). Veldtman et al. (2011) showed that 33% of novel natural enemies found in the introduced range belong to the same families as its native enemies in Australia, supporting parallels in food web dynamics between the two regions. Similarly, several native parasitoids have been found to parasitise the larvae and pupae of the Bruchid Beetle, Acanthoscelides macrophthalmus, a biological control agent introduced to South Africa in 1999 against the River Tamarind shrub, Leucaena leucocephala (Fabaceae) (Olckers 2011; Sharrat and Olckers 2012; Ramanand and Olckers 2013). Native bruchinid beetles that target native mimosoid Fabaceae in South Africa are also known to host native parasitoid communities (Impson et al. 1999), thus parasitism of the introduced host by native parasitoids may reflect parasitoid eco-evolutionary experience to mimosoid-associated beetles. However, it should be cautioned that, in general, there is insufficient knowledge and quantification of the extent of shared parasitoids between alien and native hosts and of food-web interactions in these systems to confidently invoke eco-evolutionary processes. It is clear, however, that biotic resistance has played a major role in limiting the ability of some candidate biological control agents from establishing populations in South Africa (see Hill et al. 2020, Chap. 19).
3 Mutualism
3.1 Plants and Soil Bacteria
Mutualisms can play key roles in mediating not only the establishment success of alien species (Richardson et al. 2000a), but also their ecological impacts once they become invasive (Traveset and Richardson 2006, 2011). Legumes (family Fabaceae) are over-represented in regional invasive floras in many parts of the world (Pyšek et al. 2017), and 73 legume species are naturalised in South Africa (Richardson et al. 2020, Chap. 3, Sect. 3.3). The widespread success of legumes as invasive species has been attributed partly to their ability to form symbioses with soil bacteria known as rhizobia (Parker 2001). Rhizobia are bacteria capable of forming specialised structures called root nodules on the roots of most legumes. Rhizobia fix atmospheric nitrogen into ammonium that legumes can utilise. In return, legumes provide rhizobia with various sources of carbon through photosynthate. This symbiosis allows legumes to colonise nutrient-poor environments and often impacts these environments through nitrogen enrichment of soils (Parker 2001; Yelenik et al. 2004).
Invasive Australian wattles in South Africa’s Cape Floristic Region (CFR) , a global biodiversity hotspot, are a good study system for exploring how interaction reassembly pathway (i.e. co-introduction vs. ecological fitting vs. co-xenic) and interaction specialisation affect the way in which invaders infiltrate ecological networks (Fig. 14.2) and their subsequent impacts on native species. Molecular evidence has revealed that invasive wattles in the CFR have often been co-introduced with their rhizobia, primarily from the genus Bradyrhizobium (Fig. 14.3b; Ndlovu et al. 2013; Le Roux et al. 2016; Warrington et al. 2019). However, this is not the case for all alien wattles in South Africa. A recent survey of rhizobial communities associated with 19 invasive Acacia species in South Africa showed that wattles often share highly abundant Bradyrhizobium strains across wide geographic regions (Keet et al. 2017). These observations indicate that host-switching between co-introduced rhizobia and wattles may allow those wattles not co-introduced with their Australian bradyrhizobia to overcome the potential negative effects associated with ecological fitting , potentially resulting in a form of invasion meltdown (Le Roux et al. 2017; Warrington et al. 2019).
3.2 Pollination
Almost 90% of all flowering plant species rely to some extent on pollinators for seed set (Ollerton et al. 2011). Pollination is, therefore, a potentially important barrier to establishment and subsequent invasion for alien plants (Blackburn et al. 2011). Intuitively, autonomous self-fertilisation should be less likely to limit invasiveness as it allows plants to escape the negative consequences of small population sizes, mate availability, and Allee effects (Baker 1955; Stebbins 1957). Nonetheless, many invasive plant species are pollinator-dependent (e.g. van Kleunen and Johnson 2005).
It has been argued that native pollinator systems in South Africa are more specialised than the global average (Johnson and Steiner 2003). This suggests that introduced plants requiring specialist pollinators are less likely to receive pollinator services, in line with the Missed Mutualisms Hypothesis (Catford et al. 2009). In contrast, alien plants that attract a wide range of pollinators, i.e. generalists, are expected to easily form novel interactions with pollinators in the introduced range (the New Associations Hypothesis) , which will enhance establishment and invasion success (Baker and Stebbins 1965 ; Baker 1974). However, these expectations do not always hold up. For example, Tree Tobacco, Nicotiana glauca, is pollinated by hovering hummingbirds in its native range in the Americas (Nattero and Cocucci 2007; Ollerton et al. 2012) and its tubular flowers exclude insects and other potential pollinators. In South Africa, N. glauca is pollinated by hovering sunbirds, which is surprising (Fig. 14.3a). Sunbirds have a perching lifestyle and native plants provide them with perches (Anderson et al. 2005), suggesting that a switch to a hovering lifestyle in response to novel resources (N. glauca nectar) might be adaptive (Geerts and Pauw 2009). It is likely that the outcome of N. glauca introductions may have been dramatically different if native pollinators did not adopt it as a resource. For example, in countries like Greece where bird pollinators are absent, N. glauca has adapted increased selfing ability (shorter stigma-to-anther distances) compared to plants in the native range (Ollerton et al. 2012).
Another alien plant in South Africa that requires specialist pollinators is the Formosa Lily, Lilium formosanum. In its native range in Taiwan, the lily is pollinated by the Long-tongued Convolvulus Hawkmoth, Agrius convolvuli. In South Africa, the species experiences reduced pollination in small populations, but self-fertilization sufficiently compensates for this, alleviating any potential Allee effect (Rodger et al. 2013). In denser populations in South Africa, L. formosanum is readily pollinated by A. convolvuli, since this hawkmoth is native to much of the Old World, including South Africa (Rodger et al. 2010). This example illustrates how the wide native range distributions of pollinators may facilitate reproductive success of an alien species. Similarly, invasive Peanut-butter Cassia, Senna didymobotrya in South Africa, a shrub from tropical Africa that relies on buzz pollinators (where pollinators must buzz at a specific frequency for pollen release and cross-pollination, Dulberger 1981; van Kleunen and Johnson 2005), is pollinated in South Africa by the carpenter bee (Xylocopa flavorufa). As the bee’s native range includes both South Africa (where the plant is alien) and tropical Africa (where the plant is native), this is neither a novel association nor a co-introduction. In contrast, other specialised species such as the Moth Catcher (Araujia sericifera), which, as its common name suggests, is moth pollinated, is largely visited by native honeybees, Apis mellifera, in South Africa. South African honeybees have learnt to access the nectar of the large moth catcher flowers (Coombs and Peter 2010). Despite the expectation that highly specialised mutualistic interactions may hamper establishment success (i.e. Missed Mutualisms Hypothesis , Catford et al. 2009), this example supports the emerging view that specialised pollination requirements are not necessarily a barrier to plant invasiveness (Richardson et al. 2000a). The examples discussed above show that reproductive barriers can be overcome when the same pollinators, or functionally similar pollinators are present, or if local pollinators can adapt to new resources provided by invasive populations. On the other hand, alien plants with generalist pollination requirements are expected to find pollinators more easily than their specialist counterparts, whether in urban (Geerts et al. 2017) or natural environments (Gibson et al. 2011). Generalist alien plants are assured of pollination when native generalist pollinators are abundant. Honeybees in South Africa are important pollinators for many alien plants. Examples of invasive alien plant genera with generalist flowers that are pollinated by honeybees in South Africa include Acacia, Banksia, Hakea and Pueraria (Gibson et al. 2011, 2013; Moodley et al. 2016; Geerts et al. 2016).
Although generalist pollination systems promote invasiveness , Baker (1955) postulated that selfing enhances the chances of establishment success of introduced species as it assures reproduction when mates and/or pollinators are limited. Globally, it appears that selfing rates are higher in invasive plants than for native plants (Richardson et al. 2000a; Burns et al. 2011). Support for this pattern in South African comes from a study of 17 invasive woody species which showed that all were either self-compatible or apomictic (reproducing asexually, without fertilisation) (Rambuda and Johnson 2004). Similarly, Moodley et al. (2016) found that, although pollinators increased seed set in four out of the five invasive Australian Banksia species they studied, all species were capable of autonomous selfing. Interestingly, in the Willow-leafed Hakea (Hakea salicifolia) naturalised populations received almost four times the number of pollinator visits compared to populations that had not naturalised (Moodley et al. 2016). This should not prevent invasion, since H. salicifolia produces fruits via selfing in the absence of pollinators, but such spatial variation in reproduction may explain some of the variation in the extent and rate of naturalisation (Moodley et al. 2016). Geerts et al. (2016) found that invasive Kudzu Vine, Pueraria montana (native to Asia), produces seed autonomously in South Africa. This is not the case in the USA where the species is also highly invasive. Kudzu Vine flowers are frequently visited by pollinators in both the USA and South Africa. However, in the USA only 3.3% of pollinated flowers produce pods, whereas 72% of pollinated flowers do in South Africa (Geerts et al. 2016). Despite the evident role of selfing in alien plant establishment and invasiveness , it may come at a cost. Less reliance on pollinators due to high selfing can impede invasion through higher inbreeding depression. For example, Rodger and Johnson (2013) found that for the highly invasive Silver Wattle, Acacia dealbata, selfed seedlings experienced significantly higher inbreeding depression than naturally cross-pollinated treatments.
Even if an invasive plant species has a negative effect on a specific native plant or pollinator, the effect on the community may be neutral or positive. This context dependency is due to factors such as community species richness, and the abundance of pollinators and flowers (Traveset and Richardson 2014). Further work, using pollination network analyses, is needed to advance our understanding of the resilience of South African pollinator communities to infiltration by invasive species. We know of only one non-South African study that has addressed this topic. This study found that specialist flower-visiting species are lost from pollinator webs in areas impacted by invasive brambles (Hansen et al. 2018).
Although natural ecosystems in South Africa have a few well-known invasive insect species (e.g. the Argentine Ant (Linepithema humile) and the European Wasp (Vespula germanica)), very little is known about invasive invertebrates compared to other taxonomic groups (McGeoch et al. 2011; Janion-Scheepers and Griffiths 2020, Chap. 7). Although insects with negative impacts on agricultural production are generally well-studied, very little is known about alien pollinators. However, there are some examples of alien pollinators such as the Large Cabbage White Butterflies (Pieris brassicae) and its association with Devil’s Beard (Centranthus ruber; Geerts et al. 2017) and Purple Loosestrife (Lythrum salicaria; S. Geerts unpublished data).
3.3 Seed Dispersal
As with pollination , alien plants benefit from associations with native seed dispersers, and their successful spread during invasion is often enhanced by these mutualisms (Richardson et al. 2000a; Traveset and Richardson 2006, 2014). Alien plants have become thoroughly integrated in seed dispersal networks involving native birds (Middlemiss 1963; Glyphis et al. 1981; Knight 1986, 1988; Knight and Macdonald 1991; Dean and Milton 2000; Milton et al. 2007; Underhill and Hofmeyr 2007; Mokotjomela et al. 2013a, b, 2015; Dlamini et al. 2018) and mammals (Middlemiss 1963; Kerley et al. 1996; Hill 1999; Lotter et al. 1999; Richardson et al. 2000b; Foxcroft and Rejmánek 2007; Mokotjomela and Hoffmann 2013; Tew et al. 2018) in South Africa. South Africa has a rich flora of plants adapted for seed dispersal by animals (e.g. Knight and Siegfried 1983; Knight 1988) and a rich vertebrate fauna to provide generalist seed-dispersal services.
Native South African ants also play an important role in the invasion of alien plants adapted for myrmecochory. For example, they are responsible for short-distance dispersal and seed burial of the Port Jackson Willow, Acacia saligna (Holmes 1990). While other agents are more important for long-distance dispersal in this species, burial protects seeds from predation and fire (Richardson et al. 2000a). Introduced livestock are key agents for the dispersal of many widespread invasive plant species, especially in rangelands, notably species of the genus Prosopis in South Africa (Richardson et al. 2000a). Dispersal mutualisms recorded in South Africa include several novel interactions involving native bird functional groups not recorded to disperse the plant species elsewhere, e.g. Barn Swallows (Hirundo rustica; Underhill and Hofmeyr 2007) and Black Korhaans (Eupodotis afra; Knight and Macdonald 1991) dispersing Acacia cyclops seeds, and Pied Crows (Corvus albus) dispersing Opuntia seeds (Dean and Milton 2000). The presence of wide-ranging native mammals such as African Elephants (Loxodonta africana) in some of South Africa’s protected areas has resulted in unique patterns of invasion. For example, long-distance dispersed seeds of Prickly Pear, Opuntia stricta, by elephants and Chacma Baboons (Papio ursinus) from a few initial foci in the Kruger National Park , facilitated the rapid spread of the species; a very different invasion scenario compared to that in other parts of the invasive range of this cactus (Foxcroft et al. 2004; Foxcroft and Rejmánek 2007) .
Several factors that influence competition for dispersal agents have been identified in South Africa. For example, Knight (1986) reported that bird-dispersed alien fleshy-fruited plants in the CFR have fruit displays that are more conspicuous and more attractive to native birds than those of co-occurring native plants. Another factor promoting the preference of fruits of invasive species over those produced by native species by birds is that some invasive species offer higher nutritional rewards (e.g. Cinnamomum camphora, Lantana camara, Morus alba, Psidium guajava, Solanum mauritianum; Jordaan et al. 2011; Mokotjomela et al. 2013a; Thabethe et al. 2015). The reproductive phenologies of some invasive plant species also ensure that their fruits or seeds are available for longer periods compared to many native species (Knight 1988; Mokotjomela 2012). For example, the invasion of Sand Blackberry (Rubus cuneifolius) in South Africa depends on dispersal by frugivorous birds and mammals (Denny and Goodall 1991), which exploit its prolific fruit crop throughout the year (van Kleunen and Johnson 2007). Similarly, invasion of Bugweed (S. mauritianum) in South Africa is driven by the abundance of fruit, small seediness, and high sugar content of its berries, making the species’ fruit a more attractive resource than that provided by co-occurring natives (Mokotjomela et al. 2013a).
The importance of habitat quality in the assembly of mutualisms has been well documented (Muller-Landau and Hardesty 2005). In South Africa, Schor et al. (2015) found that preferential foraging on berries of the invasive S. mauritianum by native birds declined with increasing presence of native resources (fruits) in farmlands in KwaZulu-Natal . Rejmánek (1996) argued that such context-dependency of novel resource utilisation may explain why tropical forest habitats suffer less from plant invasions than other vegetation types.
Long-distance dispersal (LDD) or stratified dispersal (a combination of long- and short-distance dispersal) is essential for species to cross environmental barriers to new recruitment sites, and therefore for subsequent naturalisation and the development of independent outlying foci that generates invasive spread (Trakhtenbrot et al. 2005). LDD facilitates establishment far from parent plants where competition , predation and/or fungal attack might be lower (Chimera and Drake 2010; Jordaan et al. 2011). Birds are important vectors for LDD of plants, as they spread ingested seeds between roosting and foraging sites (Mokotjomela et al. 2013c, 2016). Behavioural patterns, such as local and regional migrations, may also influence the extent of LDD (Mokotjomela et al. 2013c). In the CFR , Red-winged Starlings (Onychognathus morio) populations consist of resident pairs and nomadic flocks; and flocks’ movements are determined by changes in local food resources (Rowan 1955; Hockey et al. 2005). Indeed, large flocks of wintering Red-winged Starlings shuttle between home gardens and montane environments searching for fruits, which results in ingested seeds being dispersed over considerable distances (Mokotjomela 2012).
Generally, frugivorous birds and mammals determine the effectiveness of dispersing the seeds of alien plants, i.e. successful dispersal and germination (Mokotjomela et al. 2016). Indeed, native frugivorous species are often responsible for the increased invasiveness of many alien plants in South Africa (Jordaan et al. 2011; Wilson and Downs 2012; Thabethe et al. 2015; Mokotjomela et al. 2016). For example, for the highly invasive Acacia cyclops, germination is greatly enhanced following ingestion of its seed by two native frugivorous birds, the Knysna Turaco, Tauraco corythaix, and the Red-winged Starling (Mokotjomela et al. 2015, 2016). Similarly, Thabethe et al. (2015) reported enhanced seed germination for S. mauritianum, C. camphora, P. guajava, and M. alba as a result of ingestion by two native Tauraco species. On the other hand, highly invasive species like the Peruvian Pepper Tree (Schinus molle) and Syringa (Melia azedarach), even though dispersed by native frugivores in South Africa, show no germination enhancement following passage of seeds through the gut of their novel vectors (Iponga et al. 2009; Voigt et al. 2011). Wahlberg’s Epauletted Fruit Bats, Epomophorus wahlbergi, consume large numbers of fruits of four invasive plant species in South Africa (Eriobotrya japonica, M. azedarach, M. alba, and P. guajava), and with the exception of M. azedarach, this increases seed germination rates of ingested seeds (Jordaan et al. 2012). Two alien bird species, the Common Starling (Sturnus vulgaris) and House Sparrow (Passer domesticus), have been recorded feeding on the fruits of less widespread invasive species such as Pittosporum undulatum and Myoporum tenuifolium (Mokotjomela et al. 2013b). Although the impacts of these co-xenic associations on seed germination remain unknown, they suggest these plant species may become widespread invaders in the future, aided by these bird dispersers. Limited evidence suggests that co-xenic associations hamper invasiveness . We know of one example from South Africa, where invasive Rose-ringed Parakeets, Psittacula krameri, may impede establishment of alien plants due to reduced germination of ingested seeds (Thabethe et al. 2015).
For successful establishment and invasion, alien species must compete with native species for available resources. Following LDD , germination and establishment of alien seeds depends on an array of factors, ranging from availability of suitable conditions (such as those created by biophysical disturbance , e.g. anthropic habitats), to inter-specific competition . Few studies have reported on the complete seed dispersal cycle of both native and alien plant species in the same environment (Wang and Smith 2002). Nonetheless, the increasing number of invasive fleshy-fruited plants in South Africa indicates that their seeds are effectively dispersed, and establishment success is high (Mokotjomela et al. 2015). Because of the commonly smaller seed size of invasive alien fruits (Gosper and Vivian-Smith 2010; Mokotjomela et al. 2013a), more seeds can be dispersed by vertebrates than those of native species, implying that each dispersal event will likely carry more seeds of alien than native species. High propagule pressure has been reported to drive rates of recruitment of many bird-dispersed invasive species such as Schinus molle (Iponga et al. 2009). Similarly, the rapid spread of invasive Prickly Pear in South Africa’s Kruger National Park was mainly driven through seed dispersal by elephants and baboons (Lotter et al. 1999; Foxcroft and Rejmánek 2007) . Sixty percent of Prickly Pear seeds sampled from baboon faeces led to successful seed germination and seedling establishment (Lotter et al. 1999).
Patterns of seed dispersal of alien plants also influence the impacts that the alien species may have in invaded ecosystems. For example, dispersal of alien Schinus molle seeds by native birds in semi-arid savannas in South Africa has resulted in recruitment of this species mainly beneath native acacias (Vachellia tortilis), the dominant tree in this vegetation type. Initially V. tortilis may act as nurse plants for S. molle seedlings (Iponga et al. 2008). Subsequently, growth of S. molle and its superiority in competition for light over V. tortilis trees results in the gradual replacement of V. tortilis by S. molle, leading to a change in woodland structure and altered ecosystem processes (Iponga et al. 2008). Seed dispersal dynamics are altered when alien plants replace native plants in South African ecosystems. An example of this is where invasive alien Prosopis trees replace native Vachellia species in arid savanna. Differences in branch height and angle between Prosopis and Vachellia alter the habitat for birds, resulting in the loss of suitable perch sites for key frugivorous birds (Dean et al. 2002).
4 Selected Examples of Impacts on Native Species Biotic Interactions and Ecological Networks
Irrespective of the pathways and dynamics underlying interaction reassembly of alien species, it will certainly have consequences for native taxa. That is, native species may experience altered biotic interactions as invaders increase in abundance and range, which may include losses and gains of old and new associations, respectively. Establishing interactions is one thing, but their effectiveness is equally important. For example, invasive legumes may not cause legume-rhizobium associations of native host plants to collapse but may impact on the identity and effectiveness of the rhizobia they associate with. In Portugal for example, the performance of Acacia longifolia which was co-introduced with its bradyrhizobia (Rodríguez-Echeverría 2010) was much higher than it would have been had it relied on Portuguese bradyrhizobia (Rodríguez-Echeverría et al. 2012). These invasive rhizobial strains may outcompete native strains for associations with native legumes (e.g. Rodríguez-Echeverría et al. 2012), and may result in a reduction in the performance of these legumes (Rodríguez-Echeverría et al. 2012). Similar data are scarce for South Africa. Recently, Warrington et al. (2019) confirmed that invasive acacias in South Africa associate with a bradyrhizobial strains that were co-introduced from Australia. Le Roux et al. (2018) also found that acacia invasions affect both the diversity and structure of whole soil rhizobial communities in CFR soils by lowering diversity and homogenising community structure in invaded compared to uninvaded soils. They also found that overall acacia-induced changes to soil abiotic conditions further benefit their invasive performance. These changes may impact co-occurring native species in a similar way to what has been previously documented in Portugal. Such impacts may explain Le Roux et al.’s (2016) observations that native CFR legumes and invasive wattles interact with distinct rhizobial assemblages, most likely due to the phylogenetic distance between these host plant groups and the co-introduction of acacias and their symbionts (Warrington et al. 2019). Moreover, rhizobia associated with native CFR legumes sampled from wattle-invaded and uninvaded sites showed strong compositional turnover. Specialised natives appear unable to persist in wattle-invaded areas, while generalist natives could persist, but only in association with compositionally different rhizobia. This South African example illustrates that specialist native legumes may be more severely impacted by invasive acacias than generalist native congeners. Whether these perceived impacts by acacias translate into lowered symbiotic effectiveness (i.e. nitrogen fixation) of native legumes remain unknown.
The legume-rhizobium example above illustrates how invasive species can interrupt mutualistic interactions of native species. A more dramatic example comes from the disruption of ant mutualist interactions with native myrmecochorous CFR species. Bond and Slingsby (1984) found that when native ants are outcompeted and displaced by invasive Argentine Ants, the overall recruitment of myrmecochorous native plants (Mimetes cucullatus, M. pauciflorus and Leucospermum glabrum) were severely impacted. Unlike native ants, Argentine Ants are slow to discover the seeds of these plants, move them over shorter distances, and do not store them in below-ground nests. This leads to the majority of seeds being consumed by rodents. This, in turn, translated to a 50-fold reduction in the seedling emergence of Mimetes cucullatus (Red-crested Pagoda) compared to areas where no Argentine Ants were present (Bond and Slingsby 1984). It has subsequently been shown that these impacts can cause shifts in CFR plant community composition, owing to a disproportionate reduction in the densities of large-seeded species that are not being dispersed by Argentine Ants (Christian 2001). Lach (2007) also found novel mutualistic associations between invasive Argentine Ants and native membracids in the CFR, which greatly increased the discovery of inflorescences of the Wagon Tree, Protea nitida, by the ants. This in turn, led to decreased visitation rates of P. nitida flowers by several native arthropods and potential pollinators (Lach 2007).
It is well known that invasive plants can disrupt native plant-pollinator interactions (Traveset and Richardson 2014). We know that native plant-pollinator networks are highly specialised in South Africa (Pauw and Stanway 2015). Invasive plant species can influence these networks indirectly, for example by competing with native plants for pollinators and acting as ‘magnet species’, attracting pollinators away from native species (Biotic Indirect Effects Hypothesis; Catford et al. 2009) . Alternatively, invasive plants may increase the “overall attraction” and increase pollinator visitation to only certain native species. Gibson et al. (2013) asked whether the prolific flowering of invasive Acacia saligna acts as a magnet for pollinators in South Africa. They found a large overlap in floral visitors between one native species, Roepera fulva, and A. saligna. Moreover, visitation rates to R. fulva were significantly lower in invaded than in uninvaded sites. This observation was mainly due to visits of native honeybees. Whether lower visitation rates resulted in lower fitness of R. fulva (e.g. reduced seed set) was not tested. In contrast, no effects on the efficiency of bird pollination of native species was caused by the presence of the invasive Showy Banksia, Banksia speciosa (Erckie 2017). Banksia species are known to add significant amounts of nectar to the landscape during the peak flowering time of native CFR Proteaceae (Geerts et al. 2013). Erckie (2017) compared visitation rates by nectar feeding-birds and subsequent seed set, between Sugarbush, Protea compacta, populations adjacent to, and far away from, invasive B. speciosa plants. Although B. speciosa attracted significantly more sugarbirds and significantly fewer sunbirds than P. compacta, it did not reduce sugarbird numbers or visitation rates in P. compacta populations, and therefore had no impact on seed set for this species.
The Banksia speciosa example illustrates that pollination impacts on native plants may be neutral, but that native pollinators may well benefit. Evidence for such impacts comes from invasive eucalypts (genus Eucalyptus) and kangaroo paws (genus Anigozanthos) in South Africa. In its Australian range, the Evergreen Kangaroo Paw (A. flavidus) is mainly pollinated by perching Western Spinebills (Acanthorhynchus supercilliosus) and New Holland Honeyeaters (Phylidonyris novaehollandiae) (Phillips et al. 2014). Given this eco-evolutionary experience , and as expected, this species is pollinated by perching sunbirds and sugarbirds in South Africa’s CFR (Le Roux et al. 2010). Anigozanthos flavidus produces rich nectar for these birds during late summer, when nectar is generally scarce in the CFR . This was evident when, following the mechanical removal of invasive A. flavidus populations, sugarbird visitation dropped from 425 visits per hour (with sometimes more than ten birds observed at any given time) to only three sugarbirds per hour. With no overlap between the flowering times of native species and A. flavidus, it is unlikely that this negatively effects the pollination services of co-occurring native plants. Similar to kangaroo paws, Australian eucalypts are in important nectar source for honeybees during summer months in the CFR. This benefits mainly managed honeybees, which are used for crop pollination during spring (de Lange et al. 2013). The impacts of alien pollinators on plant communities in South Africa remains largely unknown. However, impacts are conceivable. For example, social bumblebees from the genus Bombus are often used for agricultural pollination services. These bees are similar to native honeybees in that they are super-generalists and will be effective pollinators of many plants in South Africa, including invasive species like Paterson’s Curse (Echium plantagineum). Furthermore, if they were to escape into natural environments, bumblebees are likely to compete with functionally similar native carpenter bees (Xylocopa spp.), which could disrupt native plant communities through inter-specific competition (Pauw 2013).
It is now also becoming evident that familiar associations under novel environmental conditions may lead to altered native species interactions. Veldtman et al. (2011) found that two gall-forming biological control agents released in South Africa against Acacia longifolia and A. saligna can affect native species interactions. Galls formed on these two invaders accumulated multi-trophic food chain links (with South African inquilines, parasitoids, and hyperparasitoids) similar to those observed in their native Australian range. Theoretically, these novel interactions can lead to apparent competition and losses of native biodiversity if the biological control agent shares these natural enemies (predators and parasitoids) with herbivores of native plants, as has been found elsewhere (Carvalheiro et al. 2008). However, it might be more appropriate to classify these interactions as commensal, whereby the introduction of the biological control agents has created a resource allowing for greater population sizes than would otherwise be maintained. Lastly, in some instances biological control might also facilitate novel co-xenic associations , such as the important agricultural pest False Codling Moth (Thaumatotibia leucotreta) utilising the galls formed on A. saligna as a larval food resource in agricultural ecosystems in South Africa (Seymour and Veldtman 2010). Notably, if biological control were to provide complete control, such cases are expected to decline over time.
Oceanic islands often suffer more severe ecological impacts from invasive species than mainland areas (see Greve et al. 2020, Chap. 8). This can, in part, be explained by the isolation and evolutionary naivety of island biotas to novel biotic interactions, e.g. extensive grazing by large herbivores or predation by mesopredators. South Africa’s sub-Antarctic Marion Island not only provides an example of this, but also illustrates the complexity and unforeseen outcomes of these novel interactions. House Mice (Mus musculus) reached Marion Island some time before 1818 (Watkins and Cooper 1986), and Domestic Cats (Felis catus) were intentionally introduced in 1951 to control them (Anderson and Condy 1974). The cat population grew rapidly, with an estimated population size of 2100 by 1970 (van Aarde 1979). Cats found burrowing seabirds to be easier prey than mice, and in the mid-1970s they were killing an estimated minimum of 635,000 petrels and prions each year (van Aarde 1980). This led to decreased breeding success of these birds and caused the local extinction of one species (Berruti et al. 1981). These impacts on birds may have also led to changes in soil nutrient fluxes (through bird manuring), in turn leading to multi-trophic cascades through their knock-on effects. Indeed, since the early 1970s, nutrient-loving tussock grasslands of Cook’s Tussock-grass, Poa cookii, also showed a rapid decline, and habitats where tussock grassland previously occurred could no longer support many animal colonies (Smith 1976; Smith and Mucina 2006). The influence of domestic cats on seabirds, and thus soil nutrient inputs, was likely the reason for the shrinkage of tussock grasslands during this period. A successful cat eradication program launched on Marion Island in 1974 led to the complete eradication of cats by 1991 (Bester et al. 2000; Greve et al. 2020, Chap. 8). Since then, seabird populations have recovered, and tussock grasslands seem to follow suite (Cooper et al. 1995). On the other hand, mice have not yet been eradicated on Marion Island (see Greve et al. 2020, Chap. 8). Mice harvest up to 100% of the seed crop produced by some plant species (Chown and Smith 1993) and can cause severe structural damage to keystone species such as the cushion plant, Azorella selago (Phiri et al. 2009). They have also begun to prey on seabird chicks (Fig. 14.3e; Jones and Ryan 2010) and consume large numbers of native insects, including important keystone species like the flightless moth, Pringleophaga marioni (Chown and Smith 1993). The knock-on effects of these disruptions to multi-tropic interactions remain unknown.
5 Synthesis and Food for Thought
The South African situation provides unique circumstances to understand the role of various ecological and evolutionary hypotheses related to biotic interactions in facilitating or impeding the spread of non-native species. The country’s exceptional biodiversity and environmental heterogeneity is reflected by an equally diverse and impressive array of invasive organisms from all over the planet. This provides unique opportunities to understand how different interaction types (e.g. familiar vs. novel associations) and their evolutionary context (i.e. eco-evolutionary experience) shape the outcomes of biological invasions. We found ample examples of studies that addressed specific ecological interactions of invasive species in South Africa. Despite very few of these explicitly testing any of the main hypotheses in invasion ecology that invoke biotic interactions, we found indirect evidence supporting some of these hypotheses.
Disruption of key mutualistic requirements, such as pollination and mycorrhization , are expected to impede invasion success. Surprisingly, in South Africa, there is no indication that any plant invasion ever failed due to a lack of pollinators, and at best, we speculate that plant invasions might only be slowed down due to pollination limitation. This observation might reflect a research bias towards species that have already become widespread, implicating that barriers to reproduction have already been crossed. Future research should therefore focus on comparative analyses of the pollination requirements and limitations of non-invasive and invasive congeners that share similar introduction histories in South Africa. Available data for insect invasions in South Africa not only support the notion that interaction reassembly pathways (novel associations, co-introduction , and co-xenic) may differently impact invasion outcomes, but also caution that these effects may be specific to the system studied. Nevertheless, the diversity of multi-trophic level networks that incorporate insects in South Africa makes them great model systems to test invasion hypotheses invoking biotic interactions.
Understanding the role of biotic interactions in mediating invasions is complicated and not a trivial task. The expectation that individual biotic interactions, or even interaction guilds (e.g. dispersal, pollination , parasitism ), can mediate the outcomes of invasions may be unrealistic under many circumstances. Mollusc invasions in South Africa exemplify this. Despite the obvious and severe negative impacts imposed by shell parasites on some of these species, their unabated spread in South Africa probably reflects the fact that other biotic and abiotic interactions, in combination with unique life-history traits, aid their invasiveness . On the other hand, one or two interaction types can have massive, and often unforeseen, consequences for invasions and their ecological impacts. Predation on breeding seabird colonies by invasive cats and mice on South Africa’s Marion Island is an example of such unforeseen impacts on multi-trophic interactions, where a decline in birds led to altered soil nutrient cycling, in turn, reducing plant cover. Reduced plant cover led to reduced habitat of other marine animals. This serves as a powerful example of how a keystone species can be indirectly impacted by the presence of one or two invasive species, with multiple knock-on effects on native species interactions.
The biodiversity consequences of co-introduction , ecological fitting , and co-xenic associations , as different pathways for interaction reassembly needs urgent attention, not only in South Africa, but globally. Using legume-rhizobium associations, Le Roux et al. (2017) recently hypothesised that the severity and rate of accrual of impacts will be higher on native plants when invasive plants are co-introduced with their co-evolved mutualists. Testing these theoretical expectations across various plant-mutualism types provide exciting future research opportunities. On the other hand, the prevalence of ecological fitting for some biotic interactions suggest that certain life-history traits predispose invasive species to infiltrate native ecological interaction networks . For example, most alien plants are readily integrated into plant–pollinator and plant-seed disperser networks as generalists. Moreover, high levels of selfing might explain why non-native plants rarely experience pollen limitation (van Kleunen et al. 2018). These general trends appear to hold for South Africa, but there are many opportunities to compare aspects of the reproductive biology of invasive species in their native and introduced ranges. A number of well-studied plant genera that are invasive in South Africa lend themselves to such studies, including Acacia, Banksia, Eucalyptus, Hakea, and Melaleuca.
While much research has been focused on biotic interactions during invasion in South Africa, there are still major gaps in our understanding. In particular, little is known of the contribution of soil organisms and microbes to South African invasions, but the experiences from elsewhere, e.g. from invasive earthworms altering forest dynamics (Bohlen et al. 2004) to the widespread loss of biodiversity due to fungal infections (e.g. Kilpatrick et al. 2010), suggest that we have but scratched the surface of this fascinating topic (see Janion-Scheepers and Griffiths 2020, Chap. 7). Also, the environmental dependency of biotic interactions during invasions remains understudied, not only in South Africa, but globally. For example, agricultural pesticides and climate change (Schweiger et al. 2010) are likely to change key biotic interactions, which could cause major shifts in the trajectories of some invasions. Similarly, the biotic-dependency of interaction assembly is often neglected. We need a better understanding of how phylogenetic relatedness between invaders and native communities is linked with interaction reassembly and what the consequences are of novel interactions for both invasive and native species. Globally, there are still a number of knowledge gaps regarding invader-resident species (e.g. symbiont) interactions and their roles in facilitating establishment and invasion (Richardson et al. 2000a). South Africa provides an ideal natural laboratory to fill some of these gaps (van Wilgen et al. 2020). A large number of good ‘model systems’ have been identified in South Africa, and in many instances their invader-mutualist/parasite interactions have been well-studied, providing ideal situations to address some of the issues outlined above.
References
Aizen MA, Sabatino M, Tylianakis JM (2012) Specialization and rarity predict non-random loss of interactions from mutualistic networks. Science 335:1486–1489. https://doi.org/10.1126/science.1215320
Alston KP, Richardson DM (2006) The roles of habitat features, disturbance, and distance from putative source populations in structuring alien plant invasions at the urban/wildland interface on the Cape Peninsula, South Africa. Biol Conserv 132:183–198. https://doi.org/10.1016/j.biocon.2006.03.023
Anderson GD, Condy PR (1974) A note on the feral house cat and mouse on Marion Island. S Afr J Antarct Res 4:58–61
Anderson BW, Spencer WW, Barrett CH (2005) Specialized bird perch aids cross-pollination: a plant scores by providing an access point for visiting sunbirds to feed on its nectar. Nature 435:41. https://doi.org/10.1038/435041a
Baker H (1955) Self-compatibility and establishment after “long-distance” dispersal. Evolution 9:347–349. https://doi.org/10.1111/j.1558-5646.1955.tb01544.x
Baker HG (1974) The evolution of weeds. Annu Rev Ecol Evol Syst 5:1–25. https://doi.org/10.1146/annurev.es.05.110174.000245
Baker HG, Stebbins GL (1965) The genetics of colonizing species. Academic, London
Bascompte J (2009) Mutualistic networks. Front Ecol Environ 7:429–436. https://doi.org/10.1890/080026
Berruti A, Griffiths AM, Imber MJ et al (1981) Status of seabirds at Prince Edward Island. S Afr J Antarct Res 10:32–33
Bester MN, Bloomer JP, Bartlett PA et al (2000) Final eradication of feral cats from sub-Antarctic Marion Island, southern Indian Ocean. S Afr J Wildl Res 30:53–57
Bezeng SB, Davies JT, Yessoufou K et al (2015) Revisiting Darwin’s naturalization conundrum: explaining invasion success of non-native trees and shrubs in southern Africa. J Ecol 103:871–879. https://doi.org/10.1111/1365-2745.12410
Blackburn TM, Pyšek P, Bacher S et al (2011) A proposed unified framework for biological invasions. Trends Ecol Evol 26:333–339. https://doi.org/10.1016/j.tree.2011.03.023
Bohlen PJ, Scheu S, Hale CM et al (2004) Non-native invasive earthworms as agents of change in northern temperate forests. Front Ecol Environ 2:427–435. https://doi.org/10.1890/1540-9295(2004)002[0427:NIEAAO]2.0.CO;2
Bond W, Slingsby P (1984) Collapse of an ant-plant mutualism: the Argentine ant (Iridomyrmex humilis) and myrmecochorous Proteaceae. Ecology 65:1031–1037. https://doi.org/10.2307/1938311
Burns JH, Ashman TL, Steets JA et al (2011) A phylogenetically controlled analysis of the roles of reproductive traits in plant invasions. Oecologia 166:1009–1017. https://doi.org/10.1007/s00442-011-1929-9
Bush SJ, Slippers B, Neser S et al (2016) Six recently recorded Australian insects associated with Eucalyptus in South Africa. Afr Entomol 24:539–544. https://doi.org/10.4001/003.024.0539
Carroll SP, Loye JE, Dingle H et al (2005) And the beak shall inherit–evolution in response to invasion. Ecol Lett 8:944–951. https://doi.org/10.1111/j.1461-0248.2005.00800.x
Carvalheiro LG, Buckley YM, Ventim R et al (2008) Apparent competition can compromise the safety of highly specific biocontrol agents. Ecol Lett 11:690–700. https://doi.org/10.1111/j.1461-0248.2008.01184.x
Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 15:22–40. https://doi.org/10.1111/j.1472-4642.2008.00521.x
Ceryngier P, Nedved O, Grez AA et al (2018) Predators and parasitoids of the harlequin ladybird, Harmonia axyridis, in its native range and invaded areas. Biol Invasions 20:1009–1031. https://doi.org/10.1007/s10530-017-1608-9
Chimera CG, Drake DR (2010) Effects of pulp removal on seed germination of five invasive plants in Hawaii. Plant Prot Q 25:137–140
Chown SL, Smith VR (1993) Climate change and the short-term impact of feral house mice at the Sub-Antarctic Prince Edward Islands. Oecologia 96:508–516. https://doi.org/10.1007/BF00320508
Christian CE (2001) Consequences of a biological invasion reveal the importance of mutualism for plant communities. Nature 413:635–639. https://doi.org/10.1038/35098093
Colautti RI, Ricciardi A, Grigorovich IA et al (2004) Is invasion success explained by the enemy release hypothesis? Ecol Lett 7:721–733. https://doi.org/10.1111/j.1461-0248.2004.00616.x
Coleman RA, Hockey PAR (2008) Effects of an alien invertebrate species and wave action on prey selection by African black oystercatchers (Haematopus moquini). Austral Ecol 33:232–240. https://doi.org/10.1111/j.1442-9993.2008.01864.x
Comont RF, Purse BV, Phillips W et al (2014) Escape from parasitism by the invasive alien ladybird, Harmonia axyridis. Insect Conserv Divers 7:334–342. https://doi.org/10.1111/icad.12060
Coombs G, Peter CI (2010) The invasive “mothcatcher” (Araujia sericifera; Asclepiadoideae) co-opts native honeybees as its primary pollinator in South Africa. AoB Plants plq021:1–14. https://doi.org/10.1093/aobpla/plq021
Cooper J, Marais AVN, Bloomer JP et al (1995) A success story: breeding of burrowing petrels (Procellariidae) before and after the extinction of feral cats Felis catus at sub-Antarctic Marion Island. Mar Ornithol 23:33–37
Coutinho TA, Wingfield MJ, Alfenas AC et al (1998) Eucalyptus rust: a disease with the potential for serious international implications. Plant Dis 82:819–825. https://doi.org/10.1094/PDIS.1998.82.7.819
Crous CJ, Burgess TI, Le Roux JJ et al (2017) Pest and pathogen accumulation in non-native trees: a case of eco-evolutionary driven non-equilibria? AoB Plants 9:plw081. https://doi.org/10.1093/aobpla/plw081
Daehler CC (2001) Darwin’s naturalization hypothesis revisited. Am Nat 158:324–330
Darwin C (1859) On the origin of species by means of natural selection. John Murray, London. https://doi.org/10.1086/321316
David AA, Simon CA (2014) The effect of temperature on larval development of two non-indigenous poecilogonous polychaetes (Annalida: Spionidae) with implications for life history theory, establishment and range expansion. J Exp Mar Bio Ecol 461:20–30. https://doi.org/10.1016/j.jembe.2014.07.012
de Lange WJ, Veldtman R, Allsopp MH (2013) Valuation of pollinator forage services provided by Eucalyptus cladocalyx. J Environ Manag 125:12–18. https://doi.org/10.1016/j.jenvman.2013.03.027
Dean WRJ, Milton SJ (2000) Directed dispersal of Opuntia species in the Karoo, South Africa: are crows the responsible agents? J Arid Environ 45:305–314. https://doi.org/10.1006/jare.2000.0652
Dean WRJ, Anderson MD, Milton SJ et al (2002) Avian assemblages in native Acacia and alien Prosopis drainage line woodland in the Kalahari, South Africa. J Arid Environ 51:1–19
Denny R, Goodall J (1991) Variable effects of glyphosate and triclopyr used for the control of American bramble, Rubus cuneifolius Agg, in pine plantations. South Afr For J 159:11–15. https://doi.org/10.1080/00382167.1991.9630389
Dlamini P, Zacharides C, Downs CT (2018) The effect of frugivorous birds on seed dispersal and germination of the invasive Brazilian pepper tree (Schinus terebinthifolius) and Indian laurel (Litsea glutinosa). S Afr J Bot 114:61–68. https://doi.org/10.1016/j.sajb.2017.10.009
Donaldson JE, Hui C, Richardson DM et al (2014) Invasion trajectory of alien trees: the role of introduction pathway and planting history. Global Chang Biol 20:1527–1537. https://doi.org/10.1111/gcb.12486
Dulberger R (1981) The floral biology of Cassia didymobotrya and C. auriculata (Caesalpiniaceae). Am J Bot 68:1350–1360. https://doi.org/10.1002/j.1537-2197.1981.tb07846.x
Ellender BR, Woodford DJ, Weyl OLF et al (2014) Managing conflicts arising from fisheries enhancements based on non-native fishes in southern Africa. J Fish Biol 85:1890–1906. https://doi.org/10.1111/jfb.12512
Enders M, Hütt MT, Jeschke JM (2018) Drawing a map of invasion biology based on a network of hypotheses. Ecosphere 9:e02146. https://doi.org/10.1002/ecs2.2146
Erckie LNK (2017) Impacts and control of alien Proteaceae invasion in the Western Cape Province, South Africa. MSc thesis, University of the Western Cape, Cape Town, South Africa
Faulkner KT, Burness A, Byrne MJ et al (2020) South Africa’s pathways of introduction and dispersal and how they have changed over time. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 311–352. https://doi.org/10.1007/978-3-030-32394-3_12
Firlej A, Lucas E, Coderre D et al (2007) Teratocytes growth pattern reflects host suitability in a host-parasitoid assemblage. Physiol Entomol 32:181–187. https://doi.org/10.1111/j.1365-3032.2006.00548.x
Foxcroft LC, Rejmánek M (2007) What helps Opuntia stricta invade Kruger National Park, South Africa: Baboons or elephants? Appl Veg Sci 10:265–270. https://doi.org/10.1111/j.1654-109X.2007.tb00525.x
Foxcroft LC, Rouget M, Richardson DM et al (2004) Reconstructing fifty years of Opuntia stricta invasion in the Kruger National Park: environmental determinants and propagule pressure. Divers Distrib 10:427–437. https://doi.org/10.1111/j.1366-9516.2004.00117.x
Foxcroft LC, Richardson DM, Wilson JRU (2008) Ornamental plants as invasive aliens: problems and solutions in the Kruger National Park, South Africa. Environ Manag 41:32–51. https://doi.org/10.1007/s00267-007-9027-9
Garnas JR, Hurley BP, Slippers B et al (2012) Biological control of forest plantation pests in an interconnected world requires greater international focus. Int J Pest Manag 58:211–223. https://doi.org/10.1080/09670874.2012.698764
Garnas JR, Aúgr-Rozenberg MA, Roques A et al (2016) Complex patterns of global spread in invasive insects: eco-evolutionary and management consequences. Biol Invasions 18:935–952. https://doi.org/10.1007/s10530-016-1082-9
Geerts S, Pauw A (2009) African sunbirds hover to pollinate an invasive hummingbird-pollinated plant. Oikos 118:573–579. https://doi.org/10.1111/j.1600-0706.2008.17167.x
Geerts S, Botha PW, Visser V et al (2013) Montpellier broom (Genista monspessulana) and Spanish broom (Spartium junceum) in South Africa: an assessment of invasiveness and options for management. S Afr J Bot 87:134–145. https://doi.org/10.1016/j.sajb.2013.03.019
Geerts S, Mashele BV, Visser V et al (2016) Lack of human-assisted dispersal means Pueraria montana var. lobata (kudzu vine) could still be eradicated from South Africa. Biol Invasions 18:3119–3126. https://doi.org/10.1007/s10530-016-1226-y
Geerts S, Rossenrode T, Irlich UM et al (2017) Emerging ornamental plant invaders in urban areas—Centranthus ruber in Cape Town, South Africa as a case study. Invasive Plant Sci Manag 10:322–331. https://doi.org/10.1017/inp.2017.35
Gibson MR, Richardson DM, Marchante E et al (2011) Reproductive biology of Australian acacias: important mediator of invasiveness? Divers Distrib 17:911–933. https://doi.org/10.1111/j.1472-4642.2011.00808.x
Gibson MR, Pauw A, Richardson DM (2013) Decreased insect visitation to a native species caused by an invasive tree in the Cape Floristic Region. Biol Conserv 157:196–203. https://doi.org/10.1016/j.biocon.2012.07.011
Giliomee JH (2010) Recent establishment of many alien insects in South Africa – a cause for concern. Afr Entomol 19:151–155. https://doi.org/10.4001/003.019.0105
Glen M, Alfenas AC, Zauza EAV et al (2007) Puccinia psidii: a threat to the Australian environment and economy – a review. Australas Plant Pathol 36(1):16. https://doi.org/10.1071/AP06088
Glyphis PJ, Milton JS, Siegfried RW (1981) Dispersal of Acacia cyclops by birds. Oecologia 48:138–141. https://doi.org/10.1007/BF00347002
Gosper C, Vivian-Smith G (2010) Fruit traits of vertebrate-dispersed alien plants: smaller seeds and more pulp sugar than indigenous species. Biol Invasions 12:2153–2163. https://doi.org/10.1007/s10530-009-9617-y
Grant WS, Cherry MI (1985) Mytilus galloprovincialis Lmk. in southern Africa. J Exp Mar Bio Ecol 90:179–191. https://doi.org/10.1016/0022-0981(85)90119-4
Greve M, Eric C, von der Meden O et al (2020) Biological invasions in South Africa’s offshore sub-Antarctic territories. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 205–226. https://doi.org/10.1007/978-3-030-32394-3_8
Griffiths CL, Hockey PAR, van Erkom Schurink C et al (1992) Marine invasive aliens on South African shores: implications for community structure and tropillc functioning. Afr J Mar Sci 12:713–722. https://doi.org/10.2989/02577619209504736
Haelewaters D, Minnaar I, Clusella-Trullas S (2016) First finding of the parasitic fungus Hesperomyces virescens (Laboulbeniales) on native and invasive ladybirds (Coleoptera, Coccinellidae) in South Africa. Parasite 23:5. https://doi.org/10.1051/parasite/2016005
Hansen S, Roets F, Seymour CL et al (2018) Alien plants have greater impact than habitat fragmentation on native insect flower visitation networks. Divers Distrib 24:58–68. https://doi.org/10.1111/ddi.12656
Hargrove J, Weyl OLF, Austin J (2017) Reconstructing the introduction history of an invasive fish predator in South Africa. Biol Invasions 19:2261–2276. https://doi.org/10.1007/s10530-017-1437-x
Harris JM, Branch GM, Elliot BL et al (1998) Spatial and temporal variability in recruitment of intertidal mussels around the coast of southern Africa. Afr Zool 33:1–11. https://doi.org/10.1080/02541858.1998.11448447
Heath RN, Gryzenhout M, Roux J et al (2006) Discovery of the canker pathogen Chrysoporthe austroafricana on native Syzygium spp. in South Africa. Plant Dis 90:433–438. https://doi.org/10.1094/PD-90-0433
Heleno RH, Olesen JM, Nogales M et al (2012) Seed dispersal networks in the Galápagos and the consequences of alien plant invasions. Proc R Soc Lond B Biol 280:2012–2112. https://doi.org/10.1098/rspb.2012.2112
Hill RA (1999) Ecological and demographic determinants of time budgets in baboons: implications for cross-populational models of baboon socioecology. PhD thesis, University of Liverpool
Hill MP, Moran VC, Hoffmann JH et al (2020) More than a century of biological control against invasive alien plants in South Africa: a synoptic view of what has been accomplished. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 549–568. https://doi.org/10.1007/978-3-030-32394-3_19
Hockey PAR, van Erkom Schurink C (1992) The invasive biology of the mussel Mytilus galloprovincialis on the southern African coast. Trans R Soc S Afr 48:123–139. https://doi.org/10.1080/00359199209520258
Hockey PAR, Dean WRJ, Ryan PG (2005) Roberts’ birds of Southern Africa. John Voelcker Bird Book Fund, Cape Town
Holmes PM (1990) Dispersal and predation of alien Acacia seeds: effects of season and invading stand density. S Afr J Bot 56:428–434. https://doi.org/10.1016/S0254-6299(16)31037-7
Holmes PM, Rebelo AG (1988) The occurrence of seed-feeding Zulubius acaciaphagus (Hemiptera, Alydidae) and its effects on Acacia cyclops seed germination and seed banks in South Africa. S Afr J Bot 54:319–324. https://doi.org/10.1016/S0254-6299(16)31298-4
Holmes PM, Rebelo AG, Irlich UM (2018) Invasive potential and management of naturalised ornamentals across an urban environmental gradient with a focus on Centranthus ruber. Bothalia-Afr Biodivers Conserv 48:a2345. https://doi.org/10.4102/abc.v48i1.2345
Impson FAC, Moran VC, Hoffmann JH (1999) A review of the effectiveness of seed feeding bruchid beetles in the biological control of mesquite, Prosopis species (Fabaceae), in South Africa. Afr Entomol Mem 1:81–88
Iponga DM, Milton SJ, Richardson DM (2008) Superiority in competition for light: a crucial attribute defining the impact of the invasive alien tree Schinus molle (Anacardiaceae) in South African savanna. J Arid Environ 72:612–623. https://doi.org/10.1016/j.jaridenv.2007.10.001
Iponga DM, Milton SJ, Richardson DM (2009) Reproductive potential and seedling establishment of the invasive alien tree Schinus molle (Anacardiaceae) in South Africa. Austral Ecol 34:678–687. https://doi.org/10.1111/j.1442-9993.2009.01975.x
Janion-Scheepers C, Griffiths CL (2020) Alien terrestrial invertebrates in South Africa. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 183–204. https://doi.org/10.1007/978-3-030-32394-3_7
Johnson SD, Steiner KE (2003) Specialized pollination systems in southern Africa. S Afr J Sci 99:345–348
Jones MGW, Ryan PG (2010) Evidence of mouse attacks on albatross chicks on sub-Antarctic Marion Island. Antarct Sci 22:39–42. https://doi.org/10.1017/S0954102009990459
Jordaan LA, Johnson SD, Downs CT (2011) Digestion of fruit of invasive alien plants by three southern African avian frugivores. Ibis 153:863–867. https://doi.org/10.1111/j.1474-919X.2011.01157.x
Jordaan LA, Johnson SD, Downs CT (2012) Wahlberg’s epauletted fruit bat (Epomophorus wahlbergi) as a potential dispersal agent for fleshy-fruited invasive alien plants: effects of handling behaviour on seed germination. Biol Invasions 14:959–968. https://doi.org/10.1007/s10530-011-0131-7
Kaehler S, McQuaid CD (1999) Lethal and sub-lethal effects of phototrophic endoliths attacking the shell of the intertidal mussel Perna perna. Mar Biol 135:497–503. https://doi.org/10.1007/s002270050650
Keet JH, Ellis AG, Hui C et al (2017) Legume–rhizobium symbiotic promiscuity and effectiveness do not affect plant invasiveness. Ann Bot 119:1319–1331. https://doi.org/10.1093/aob/mcx028
Kerley GIH, McLachlan A, Castley JG (1996) Diversity and dynamics of bush-pockets in the Alexandria Coastal Dunefield, South Africa. Landscape Urban Plan 34:255–266. https://doi.org/10.1016/0169-2046(95)00224-3
Kfir R (1998) Origin of the diamondback moth (Lepidoptera: Plutellidae). Ann Entom Soc Am 91:164–167. https://doi.org/10.1093/aesa/91.2.164
Kilpatrick AM, Briggs CJ, Daszak P (2010) The ecology and impact of chytridiomycosis: an emerging disease of amphibians. Trends Ecol Evol 25:109–118. https://doi.org/10.1016/j.tree.2009.07.011
Knevel IC, Lans T, Menting FBJ et al (2004) Release from native root herbivores and biotic resistance by soil pathogens in a new habitat both affect the alien Ammophila arenaria in South Africa. Oecologia 141:502–510. https://doi.org/10.1007/s00442-004-1662-8
Knight RS (1986) A comparative analysis of fleshy fruit displays in alien and indigenous plants. In: Macdonald IAW, Kruger FJ, Ferrar AA (eds) The ecology and management of biological invasions in Southern Africa. Oxford University Press, Cape Town, pp 171–178
Knight RS (1988) Aspects of plant dispersal in the south-western Cape with particular reference to the roles of birds as dispersal agents. Ph.D. Thesis, University of Cape Town, South Africa
Knight RS, Macdonald IAW (1991) Acacias and korhaans: an artificially assembled seed dispersal system. S Afr J Bot 57:220–225. https://doi.org/10.1016/S0254-6299(16)30942-5
Knight RS, Siegfried WR (1983) Inter-relationships between type, size and colour of fruits and dispersal in southern African trees. Oecologia 56:405–412. https://doi.org/10.1007/BF00379720
Lach L (2007) A mutualism with native membracid facilitates pollinator displacement by Argentine ants. Ecology 88:1994–2004. https://doi.org/10.1890/06-1767.1
Le Roux JJ, Geerts S, Ivey P et al (2010) Molecular systematics and ecology of invasive Kangaroo Paws in South Africa: management implications for a horticulturally important genus. Biol Invasions 12:3989–4002. https://doi.org/10.1007/s10530-010-9818-4
Le Roux JJ, Mavengere N, Ellis AG (2016) The structure of legume-rhizobium interaction networks and their response to tree invasions. AoB Plants 8:plw038. https://doi.org/10.1093/aobpla/plw038
Le Roux JJ, Hui C, Keet JH et al (2017) Co-introduction vs ecological fitting as pathways to the establishment of effective mutualisms during biological invasions. New Phytol 215:1354–1360. https://doi.org/10.1111/nph.14593
Le Roux JJ, Ellis AG, van Zyl LM et al (2018) Importance of soil legacy effects and successful mutualistic interactions during Australian acacia invasions in nutrient poor environments. J Ecol 106:2071–2081. https://doi.org/10.1111/1365-2745.12965
Lombaert E, Guillemaud T, Cornuet JM et al (2010) Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird. PLoS One 5:e9743. https://doi.org/10.1371/journal.pone.0009743
Lotter WD, Thatcher L, Rossouw L et al (1999) The influence of baboon predation and time in water on germination and early establishment of Opuntia stricta (Australian pest pear) in Kruger National Park. Koedoe 42:43–50. https://doi.org/10.4102/koedoe.v42i1.220
Malan AP, Moore SD (2016) Evaluation of local entomopathogenic nematodes for the control of false codling moth, Thaumatotibia leucotreta (Meyrick, 1913), in a citrus orchard in South Africa. Afr Entomol 24:489–501. https://doi.org/10.4001/003.024.0489
Manongi FS, Hoffmann JH (1995) The incidence of parasitism in Trichilogaster acaciaelongifoliae (Hymenoptera: Pteromalidae), a gall-forming biological control agent of Acacia longifolia (Fabaceae) in South Africa. Afr Entomol 3:147–151
Marquet N, Nicastro KR, Gektidis M et al (2013) Comparison of phototrophic shell-degrading endoliths in invasive and native populations of the intertidal mussel Mytilus galloprovincialis. Biol Invasions 15:1253–1272. https://doi.org/10.1007/s10530-012-0363-1
McGeoch MA, Wossler TC (2000) Range expansion and success of the weed biocontrol agent Trichilogaster acaciaelongifoliae (Froggatt) (Hymenoptera: Pteromalidae) in South Africa. Afr Entomol 8:273–280
McGeoch MA, Sithole H, Samways MJ et al (2011) Conservation and monitoring of invertebrates in terrestrial protected areas. Koedoe 53:1–13. https://doi.org/10.4102/koedoe.v53i2.1000
Measey J, Hui C, Somers M (2020) Terrestrial vertebrate invasions in South Africa. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 113–150. https://doi.org/10.1007/978-3-030-32394-3_5
Mgocheki N, Addison P (2010) Spatial distribution of ants (Hymenoptera: Formicidae), vine mealybugs and mealybug parasitoids in vineyards. J Appl Entomol 134:285–295. https://doi.org/10.1111/j.1439-0418.2009.01494.x
Middlemiss E (1963) The distribution of Acacia cyclops in the Cape Peninsula area by birds and other animals. S Afr J Sci 59:419–420
Milton SJ, Wilson JRU, Richardson DM et al (2007) Invasive alien plants infiltrate bird-mediated shrub nucleation processes in arid savanna. J Ecol 95:648–661. https://doi.org/10.1111/j.1365-2745.2007.01247.x
Minnaar IA, Shinner R, van Noort S et al (2014) New records of the parasitic wasp Dinocampus coccinellae (Hymeoptera: Braconidae) and its hosts in South Africa. Afr Entomol 22:226–229
Mokotjomela TM (2012) A comparison of bird foraging preferences for fruits of indigenous and alien shrubs and seed dispersal potentials in the Cape Floristic Region. PhD Thesis, Stellenbosch University. https://doi.org/10.4001/003.022.0124
Mokotjomela TM, Hoffmann JH (2013) Removal of post-dispersed seeds in Acacia cyclops thickets under biological control in South Africa. S Afr J Bot 88:260–264. https://doi.org/10.1016/j.sajb.2013.08.004
Mokotjomela TM, Musil CF, Esler KJ (2013a) Do frugivorous birds concentrate their foraging activities on those alien plants with the most abundant and nutritious fruits in the South African Mediterranean-climate region? Plant Ecol 214:49–59. https://doi.org/10.1007/s11258-012-0145-y
Mokotjomela TM, Musil CF, Esler KJ (2013b) Frugivorous birds visit fruits of emerging alien shrub species more frequently than those of native shrub species in the South African Mediterranean climate region. S Afr J Bot 86:73–78. https://doi.org/10.1016/j.sajb.2013.02.004
Mokotjomela TM, Musil CF, Esler KJ (2013c) Potential seed dispersal distances of native and non-native fleshy fruiting shrubs in the South African Mediterranean climate region. Plant Ecol 214:1127–1137. https://doi.org/10.1007/s11258-013-0237-3
Mokotjomela TM, Hoffmann JH, Downs CT (2015) The potential for birds to disperse the seeds of Acacia cyclops, an invasive alien plant in South Africa. Ibis 157:449–458. https://doi.org/10.1111/ibi.12260
Mokotjomela TM, Downs CT, Esler KS et al (2016) Seed dispersal effectiveness: a comparison of four bird species feeding on seeds of invasive Acacia cyclops in South Africa. S Afr J Bot 105:259–263. https://doi.org/10.1016/j.sajb.2016.04.005
Moodley D, Geerts S, Richardson DM et al (2016) The importance of pollinators and autonomous self-fertilisation in the early stages of plant invasions: Banksia and Hakea (Proteaceae) as case studies. Plant Biol 18:124–131. https://doi.org/10.1111/plb.12334
Mosiane SM, Kfir R, Villet MH (2003) Seasonal phenology of the diamondback moth, Plutella xylostella (L.) (Lepidoptera: Plutellidae), and its parasitoids on canola, Brassica napus (L.), in Gauteng province, South Africa. Afr Entomol 11:277–285
Muller-Landau HC, Hardesty BD (2005) Seed dispersal of woody plants in tropical forests: concepts, examples, and future directions. In: Burslem D, Pinard M, Hartley S (eds) Biotic interactions in the tropics: their role in the maintenance of species diversity. Cambridge University Press, Cambridge, pp 267–309. https://doi.org/10.1017/CBO9780511541971.012
Nattero J, Cocucci AA (2007) Geographical variation in floral traits of the tree tobacco in relation to its hummingbird pollinator fauna. Biol J Linn Soc 90:657–667. https://doi.org/10.1111/j.1095-8312.2007.00756.x
Naylor RL, Williams SL, Strong DR (2001) Aquaculture – a gateway for exotic species. Science 294:1655–1656. https://doi.org/10.1126/science.1064875
Ndlovu J, Richardson DM, Wilson JRU et al (2013) Co-invasion of South African ecosystems by an Australian legume and its rhizobial symbionts. J Biogeogr 40:1240–1251. https://doi.org/10.1111/jbi.12091
Noble IR (1989) Attributes of invaders and the invading process: terrestrial and vascular plants. In: Drake JA, Mooney HA, di Castri F, Groves RH, Kruger FJ, Rejmanek M, Williamson M (eds) Biological invasions. Wiley, Chichester, pp 301–313
Nofemela RS (2013) The effect of obligate hyperparasitoids on biological control: differential vulnerability of primary parasitoids to hyperparasitism can mitigate trophic cascades. Biol Control 65:218–224. https://doi.org/10.1016/j.biocontrol.2013.02.003
Nofemela RS, Kfir R (2005) The role of parasitoids in suppressing diamondback moth, Plutella xylostella (L) (Lepidoptera: Plutellidae), populations on unsprayed cabbage in the North West Province of South Africa. Afr Entomol 13:71–83
Nuñez MA, Dickie IA (2014) Invasive belowground mutualists of woody plants. Biol Invasions 16:645–661. https://doi.org/10.1007/s10530-013-0612-y
Olckers T (2011) Biological control of Leucaena leucocephala (Fabaceae) in South Africa: a tale of opportunism, seed feeders and unanswered questions. Afr Entomol 19:356–365. https://doi.org/10.4001/003.019.0219
Ollerton J, Winfree R, Tarrant S (2011) How many flowering plants are pollinated by animals? Oikos 120:321–326. https://doi.org/10.1111/j.1600-0706.2010.18644.x
Ollerton J, Watts S, Connerty S et al (2012) Pollination ecology of the invasive tree tobacco Nicotiana glauca: comparisons across native and non-native ranges. J Pollinat Ecol 9:85–95. https://doi.org/10.26786/1920-7603(2012)12
Parker MA (2001) Mutualism as a constraint on invasion success for legumes and rhizobia. Divers Distrib 7:125–136. https://doi.org/10.1046/j.1472-4642.2001.00103.x
Pauw A (2013) Can pollination niches facilitate plant coexistence? Trends Ecol Evol 28:30–37. https://doi.org/10.1016/j.tree.2012.07.019
Pauw A, Stanway R (2015) Unrivalled specialization in a pollination network from South Africa reveals that specialization increases with latitude only in the Southern Hemisphere. J Biogeogr 42:652–661. https://doi.org/10.1111/jbi.12453
Phillips RD, Steinmeyer F, Menz MHM et al (2014) Changes in the composition and behaviour of a pollinator guild with plant population size and the consequences for plant fecundity. Funct Ecol 28:846–856. https://doi.org/10.1111/1365-2435.12237
Phiri EE, McGeoch MA, Chown SL (2009) Spatial variation in structural damage to a keystone plant species in the sub-Antarctic: interactions between Azorella selago and invasive house mice. Antarct Sci 21:189–196. https://doi.org/10.1017/S0954102008001569
Post DM, Palkovacs EP (2009) Eco-evolutionary feedbacks in community and ecosystem ecology: interactions between the ecological theater and the evolutionary play. Philos Trans R Soc B 364:1629–1640. https://doi.org/10.1098/rstb.2009.0012
Potgieter LJ, Douwes E, Gaertner M et al (2020) Biological invasions in South Africa’s urban ecosystems: patterns, processes, impacts and management. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 273–310. https://doi.org/10.1007/978-3-030-32394-3_11
Prenter J, MacNeil C, Dick JTA et al (2004) Roles of parasites in invasions. Trends Ecol Evol 19:385–390. https://doi.org/10.1016/j.tree.2004.05.002
Procheş S, Wilson JRU, Richardson DM et al (2008) Herbivores, but not other insects, are scarce on alien plants. Austral Ecol 33:691–700. https://doi.org/10.1111/j.1442-9993.2008.01836.x
Pyšek P, Pergl J, Essl F et al (2017) Naturalized alien flora of the world: species diversity, taxonomic and phylogenetic patterns, geographic distribution and global hotspots of plant invasion. Preslia 89:203–274. https://doi.org/10.23855/preslia.2017.203
Ramanand H, Olckers T (2013) Does height of exposure in the canopy influence egg mortality in Acanthoscelides macrophthalmus (Coleoptera: Chrysomelidae), a biological control agent of Leucaena leucocephala in South Africa? Biocontrol Sci Technol 23:545–554. https://doi.org/10.1080/09583157.2013.790344
Rambuda TD, Johnson SD (2004) Breeding systems of invasive alien plants in South Africa: does Baker’s rule apply? Divers Distrib 10:409–416. https://doi.org/10.1111/j.1366-9516.2004.00100.x
Rejmánek M (1996) A theory of seed plant invasiveness: the first sketch. Biol Conserv 78:171–181
Richardson DM, Pyšek P (2012) Naturalization of introduced plants: ecological drivers of biogeographic patterns. New Phytol 196:383–396. https://doi.org/10.1111/j.1469-8137.2012.04292.x
Richardson DM, van Wilgen BW (2004) Invasive alien plants in South Africa: how well do we understand the ecological impacts? Working for water. S Afr J Sci 100:45–52
Richardson DM, Williams PA, Hobbs RJ (1994) Pine invasions in the Southern Hemisphere: determinants of spread and invasibility. J Biogeogr 21:511–527. https://doi.org/10.2307/2845655
Richardson DM, Allsopp N, D’Antonio CM et al (2000a) Plant invasions—the role of mutualisms. Biol Rev 75:65–93. https://doi.org/10.1017/S0006323199005435
Richardson DM, Bond WJ, Dean WRJ et al (2000b) Invasive alien organisms and global change: a South African perspective. In: Mooney HA, Hobbs RJ (eds) Invasive species in a changing world. Island Press, Washington, DC, pp 303–349
Richardson DM, Pyšek P, Rejmánek M et al (2000c) Naturalization and invasion of alien plants – concepts and definitions. Divers Distrib 6:93–107. https://doi.org/10.1046/j.1472-4642.2000.00083.x
Richardson DM, Rouget M, Ralston SJ et al (2005) Species richness of alien plants in South Africa: environmental correlates and the relationship with indigenous plant species richness. EcoScience 12:391–402. https://doi.org/10.2980/i1195-6860-12-3-391.1
Richardson DM, Wilson JRU, Weyl OLF et al (2011) South Africa: invasions. In: Simberloff D, Rejmánek M (eds) Encyclopedia of biological invasions. University of California Press, Berkeley, pp 643–651
Richardson DM, Foxcroft LC, Latombe G et al (2020) The biogeography of South African terrestrial plant invasions. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 65–94. https://doi.org/10.1007/978-3-030-32394-3_3
Rius M, McQuaid CD (2006) Wave action and competitive interaction between the invasive mussel Mytilus galloprovincialis and the indigenous Perna perna in South Africa. Mar Biol 150:69–78. https://doi.org/10.1007/s00227-006-0322-4
Rius M, McQuaid CD (2009) Facilitation and competition between invasive and indigenous mussels over a gradient of physical stress. Basic Appl Ecol 10:607–613. https://doi.org/10.1016/j.baae.2009.03.008
Robinson TB, Griffiths CL, McQuaid CD et al (2005) Marine alien species of South Africa – status and impacts. Afr J Mar Sci 27:297–306. https://doi.org/10.2989/18142320509504088
Robinson TB, Peters K, Brooker B (2020) Coastal invasions: the South African context. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 227–246. https://doi.org/10.1007/978-3-030-32394-3_9
Rodger JG, Johnson SD (2013) Self-pollination and inbreeding depression in Acacia dealbata: can selfing promote invasion in trees? S Afr J Bot 88:252–259. https://doi.org/10.1016/j.sajb.2013.08.002
Rodger JG, van Kleunen M, Johnson SD (2010) Does specialized pollination impede plant invasions? Int J Plant Sci 171:382–391. https://doi.org/10.1086/651226
Rodger JG, van Kleunen M, Johnson SD (2013) Pollinators, mates and Allee effects: the importance of self-pollination for fecundity in an invasive lily. Funct Ecol 27:1023–1033. https://doi.org/10.1111/1365-2435.12093
Rodríguez-Echeverría S (2010) Rhizobial hitchhikers from down under: invasional meltdown in a plant–bacteria mutualism? J Biogeogr 37:1611–1622. https://doi.org/10.1111/j.1365-2699.2010.02284.x
Rodríguez-Echeverría S, Fajardo S, Ruiz-Díez B, Fernández-Pascual M (2012) Differential effectiveness of novel and old legume-rhizobia mutualisms: implications for invasion by exotic legumes. Oecologia 170:253–261. https://doi.org/10.1007/s00442-012-2299-7
Röhrich CR, Ngwa CJ, Wiesner J et al (2012) Harmonine, a defence compound from the harlequin ladybird, inhibits mycobacterial growth and demonstrates multi-stage antimalarial activity. Biol Lett 8:308–311. https://doi.org/10.1098/rsbl.2011.0760
Roux J, Hurley BP, Wingfield MJ (2012) Disease and pests of eucalypts, pine and wattle. In: South African forestry handbook. Pretoria, The South African Institute of Forestry, pp 303–335
Roux J, Germishuizen I, Nadel R et al (2015) Risk assessment for Puccinia psidii becoming established in South Africa. Plant Pathol 64:1326–1335. https://doi.org/10.1111/ppa.12380
Rowan MK (1955) The breeding biology and behaviour of the Redwinged Starling, Onychognathus morio. Ibis 97:663–705. https://doi.org/10.1111/j.1474-919X.1955.tb01926.x
Roy HE, Brown PJ, Adriaens T et al (2016) The harlequin ladybird, Harmonia axyridis: global perspectives on invasion history and ecology. Biol Invasions 18:997–1044. https://doi.org/10.1007/s10530-016-1077-6
Saul WC, Jeschke JM, Heger T (2013) The role of eco-evolutionary experience in invasion success. NeoBiota 17:57–74. https://doi.org/10.3897/neobiota.17.5208
Schor J, Farwig N, Berens DG (2015) Intensive land-use and high native fruit availability reduce fruit removal of the invasive Solanum mauritianum in South Africa. S Afr J Bot 96:6–12. https://doi.org/10.1016/j.sajb.2014.11.004
Schweiger O, Biesmeijer JC, Bommarco R et al (2010) Multiple stressors on biotic interactions: how climate change and alien species interact to affect pollination. Biol Rev 85:777–795. https://doi.org/10.1111/j.1469-185X.2010.00125.x
Seymour CL, Veldtman R (2010) Ecological role of control agent, and not just host-specificity, determine risks of biological control. Austral Ecol 35:704–711. https://doi.org/10.1111/j.1442-9993.2009.02077.x
Sharrat MEJ, Olckers T (2012) The biological control agent Acanthoscelides macrophthalmus (Chrysomelidae: Bruchinae) inflicts moderate levels of seed damage on its target, the invasive tree Leucaena leucocephala (Fabaceae) in the KwaZulu-Natal coastal region of South Africa. Afr Entomol 20:44–51. https://doi.org/10.4001/003.020.0106
Simberloff D, Von Holle B (1999) Positive interactions of non-indigenous species: invasional meltdown? Biol Invasions 1:21–32. https://doi.org/10.1023/A:1010086329619
Simon CA, Sato-Okoshi W (2015) Polydorid polychaetes on farmed molluscs: distribution, spread and factors contributing to their success. Aquac Environ Interact 7:147–166. https://doi.org/10.3354/aei00138
Simon CA, Ludford A, Wynne S (2006) Spionid polychaetes infesting cultured abalone Haliotis midae in South Africa. Afr J Mar Sci 28:167–171. https://doi.org/10.2989/18142320609504141
Simon CA, Thornhill DJ, Oyarzun F et al (2009) Genetic similarity between Boccardia proboscidea from western North America and cultured abalone, Haliotis midae, in South Africa. Aquaculture 294:18–24. https://doi.org/10.1016/j.aquaculture.2009.05.022
Smit NJ, Malherbe W, Hadfield KA (2017) Alien freshwater fish parasites from South Africa: diversity, distribution, status and the way forward. Int J Parasitol Parasites Wildl 6:386–401. https://doi.org/10.1016/j.ijppaw.2017.06.001
Smith VR (1976) The effect of burrowing species of Procellariidae on the nutrient status of inland tussock grasslands on Marion Island. S Afr J Bot 42:265–272
Smith VR, Mucina L (2006) Vegetation of sub-Antarctic Marion and Prince Edward Islands. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19:698–723
Stals R, Prinsloo G (2007) Discovery of an alien invasive, predatory insect in South Africa: the multi-coloured Asian ladybird beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). S Afr J Sci 103:123–126
Stebbins GL (1957) Self fertilization and population variability in the higher plants. Am Nat 91:337–354. https://doi.org/10.1086/281999
Strauss SY, Lau JA, Carroll SP (2006) Evolutionary responses of natives to introduced species: what do introductions tell us about natural communities? Ecol Lett 9:357–374. https://doi.org/10.1111/j.1461-0248.2005.00874.x
Stricker KB, Harmon PF, Goss EM et al (2016) Emergence and accumulation of novel pathogens suppress an invasive species. Ecol Lett 19:469–477. https://doi.org/10.1111/ele.12583
Taraschewski H (2006) Hosts and parasites as aliens. J Helminthol 80:99–128. https://doi.org/10.1079/JOH2006364
Tew E, Landman M, Kerley GIH (2018) The contribution of the chacma baboon to seed dispersal in the eastern Karoo, South Africa. Afr J Wildl Res 48(2):1–8. https://doi.org/10.3957/056.048.023002
Thabethe V, Wilson AL, Hart L et al (2015) Ingestion by an invasive parakeet species reduces germination success of invasive alien plants relative to ingestion by indigenous turaco species in South Africa. Biol Invasions 17:1–11. https://doi.org/10.1007/s10530-015-0932-1
Trakhtenbrot A, Nathan R, Perry G et al (2005) The importance of long distance dispersal in biodiversity conservation. Divers Distrib 11:173–181. https://doi.org/10.1111/j.1366-9516.2005.00156.x
Traveset A, Richardson DM (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol Evol 21:208–216. https://doi.org/10.1016/j.tree.2006.01.006
Traveset A, Richardson DM (2011) Mutualisms – key drivers of invasions… key casualties of invasions. In: Richardson DM (ed) Fifty years of invasion ecology. The legacy of Charles Elton. Wiley-Blackwell, Oxford, pp 143–160. https://doi.org/10.1002/9781444329988.ch12
Traveset A, Richardson DM (2014) Mutualistic interactions and biological invasions. Annu Rev Ecol Evol Syst 45:89–113. https://doi.org/10.1146/annurev-ecolsys-120213-091857
Truter M, Přikrylová E, Weyl OLF et al (2017) Co-introduction of ancyrocephalid monogeneans on their invasive host, the largemouth bass, Micropterus salmoides (Lacepéde, 1802) in South Africa. Int J Parasitol Parasites Wildl 6:420–429. https://doi.org/10.1016/j.ijppaw.2017.06.002
Underhill GL, Hofmeyr HJ (2007) Barn swallows Hirundo rustica disperse seeds of rooikrans Acacia cyclops, an invasive alien plant in the fynbos biome. Ibis 149:468–471. https://doi.org/10.1111/j.1474-919X.2007.00598.x
van Aarde RJ (1979) Distribution and density of the feral house cat on Marion Island. S Afr J Antarc Res 9:14–19
van Aarde RJ (1980) The diet and feeding behaviour of feral cats at Marion Island. Afr J Wildl Res 101:123–128
van der Colff D, Dreyer LL, Valentine A et al (2015) Invasive plant species may serve as a biological corridor for the invertebrate fauna of naturally isolated hosts. J Insect Conserv 19:863–875. https://doi.org/10.1007/s10841-015-9804-3
van Erkom Schurink C, Griffiths CL (1991) A comparison of reproductive cycles and reproductive output in four southern African mussel species. Mar Ecol Prog Ser 76:123–134. https://doi.org/10.3354/meps076123
van Kleunen M, Johnson SD (2005) Testing for ecological and genetic Allee effects in the invasive shrub Senna didymobotrya (Fabaceae). Am J Bot 92:1124–1130. https://doi.org/10.3732/ajb.92.7.1124
van Kleunen M, Johnson SD (2007) South African Iridaceae with rapid and profuse seedling emergence are more likely to become naturalized in other regions. J Ecol 95:674–681. https://doi.org/10.1111/j.1365-2745.2007.01250.x
van Kleunen M, Bossdorf O, Dawson W (2018) The ecology and evolution of alien plants. Annu Rev Ecol Evol Syst 49:25–47. https://doi.org/10.1146/annurev-ecolsys-110617-062654
van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya T (2020) Biological invasions in South Africa: an overview. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 3–30. https://doi.org/10.1007/978-3-030-32394-3_1
Veldtman R, Lado TF, Botes A et al (2011) Creating novel food webs on introduced Australian acacias: indirect effects of galling biological control agents. Divers Distrib 17:958–967. https://doi.org/10.1111/j.1472-4642.2011.00781.x
Vilcinskas A, Mukherjee K, Vogel H (2013a) Expansion of the antimicrobial peptide repertoire in the invasive ladybird Harmonia axyridis. Proc R Soc Lond B Biol Sci 280:20122113. https://doi.org/10.1098/rspb.2012.2113
Vilcinskas A, Stoecker K, Schmidtberg H et al (2013b) Invasive harlequin ladybird carries biological weapons against native competitors. Science 340:862–863. https://doi.org/10.1126/science.1234032
Voigt FA, Farwig N, Johnson SD (2011) Interactions between the invasive tree Melia azedarach (Meliaceae) and native frugivores in South Africa. J Trop Ecol 27:355–363. https://doi.org/10.1017/S0266467410000702
Walker G, Robertson MP, Gaertner M, Gallien L et al (2017) Assessing the potential range of Ailanthus altissima (Simaroubaceae; tree of heaven) in South Africa: the role of climate and disturbance. Biol Invasions 19:3675–3690. https://doi.org/10.1007/s10530-017-1597-8
Wang BC, Smith TB (2002) Closing the seed dispersal loop. Trends Ecol Evol 17:379–386. https://doi.org/10.1016/S0169-5347(02)02541-7
Warrington S, Ellis A, Novoa A, Wandrag EM, Hulme PE, Duncan RP, Valentine A, Le Roux JJ (2019) Cointroductions of Australian acacias and their rhizobial mutualists in the Southern Hemisphere. J Biogeogr 46:1519-1531. https://doi.org/10.1111/jbi.13602
Watkins BP, Cooper J (1986) Introduction, present status and control of alien species at the Prince Edward islands, Sub-Antarctic. S Afr J Antarct Res 16:86–94
Weyl OLF, Ellender B, Wassermann RJ et al (2020) Alien freshwater fauna in South Africa. In: van Wilgen BW, Measey J, Richardson DM, Wilson JR, Zengeya TA (eds) Biological invasions in South Africa. Springer, Berlin, pp 151–182. https://doi.org/10.1007/978-3-030-32394-3_6
Wilson AL, Downs CT (2012) Knysna Turacos (Tauraco corythaix) do not improve seed germination of ingested fruit of some indigenous South African tree species. S Afr J Bot 78:55–62. https://doi.org/10.1016/j.sajb.2011.05.006
Wingfield MJ, Swart WJ, Abear BJ (1989) First record of Cryphonectria canker of Eucalyptus in South Africa. Phytophylactica 21:311–313
Wingfield MJ, Slippers B, Roux J et al (2001) Worldwide movement of exotic forest fungi, especially in the tropics and the southern hemisphere. Bioscience 51:134–140. https://doi.org/10.1641/0006-3568(2001)051[0134:WMOEFF]2.0.CO;2
Wingfield MJ, Slippers B, Hurley BP et al (2008) Eucalypt pests and diseases: growing threats to plantation productivity. Sth For 70:139–144
Yelenik SG, Stock WD, Richardson DM (2004) Ecosystem-level impacts of invasive alien nitrogen-fixing plants. Ecosystem and community-level impacts of invasive alien Acacia saligna in the fynbos vegetation of South Africa. Restor Ecol 12:44–51. https://doi.org/10.1111/j.1061-2971.2004.00289.x
Zachariades C, Paterson ID, Strathie LW et al (2017) Assessing the status of biological control as a management tool for suppression of invasive alien plants in South Africa. Bothalia-Afr Biodivers Conserv 47:1–9. https://doi.org/10.4102/abc.v47i2.2142
Zardi GI, Nicastro KR, McQuaid CD et al (2009) Effects of endolithic parasitism on invasive and indigenous mussels in a variable physical environment. PLoS One 4:e6560. https://doi.org/10.1371/journal.pone.0006560
Acknowledgements
We acknowledge support from the DSI-NRF Centre of Excellence for Invasion Biology. JLR acknowledges startup funds received from Macquarie University’s Faculty of Science and Engineering and Department of Biological Sciences as well as South Africa’s National Research Foundation (NRF, grant no. 112097). Additional funding from the NRF is acknowledged (grant no. 85417 to DMR, grant no. 115093 to SG, grant no. 110507 to OLFW). The South African Department of Environment, Forestry, and Fisheries (DEFF) are thanked for funding the South African National Biodiversity Institute noting that this publication does not necessarily represent the views or opinions of DEFF or its employees.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2020 The Author(s)
About this chapter

Cite this chapter
Le Roux, J.J. et al. (2020). Biotic Interactions as Mediators of Biological Invasions: Insights from South Africa. In: van Wilgen, B., Measey, J., Richardson, D., Wilson, J., Zengeya, T. (eds) Biological Invasions in South Africa. Invading Nature - Springer Series in Invasion Ecology, vol 14. Springer, Cham. https://doi.org/10.1007/978-3-030-32394-3_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-32394-3_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-32393-6
Online ISBN: 978-3-030-32394-3
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)